1. Introduction
Fusarium head blight (FHB) is a global disease affecting small grain cereals including wheat, barley and oat [
1,
2,
3,
4] The disease is caused by several
Fusarium species of which
Fusarium graminearum Schwabe is the primary etiological agent in wheat crops in Canada based on the frequency of isolation and impact on grain yield and quality [
5,
6,
7,
8]. In addition to a reduction in yield,
F. graminearum produces the type B trichothecene mycotoxins, deoxynivalenol (DON) and its acetylated derivatives (15-acetyl-DON (15ADON) and 3-acetyl-DON (3ADON)) in Canada [
7,
9]. The contamination of wheat by mycotoxins is a serious health concern for both humans and animals. This has led to strict DON tolerance limits in Canada that are established at 2 mg kg
−1 body weight for unprocessed grain and 0.2 mg kg
−1 for baby food [
10].
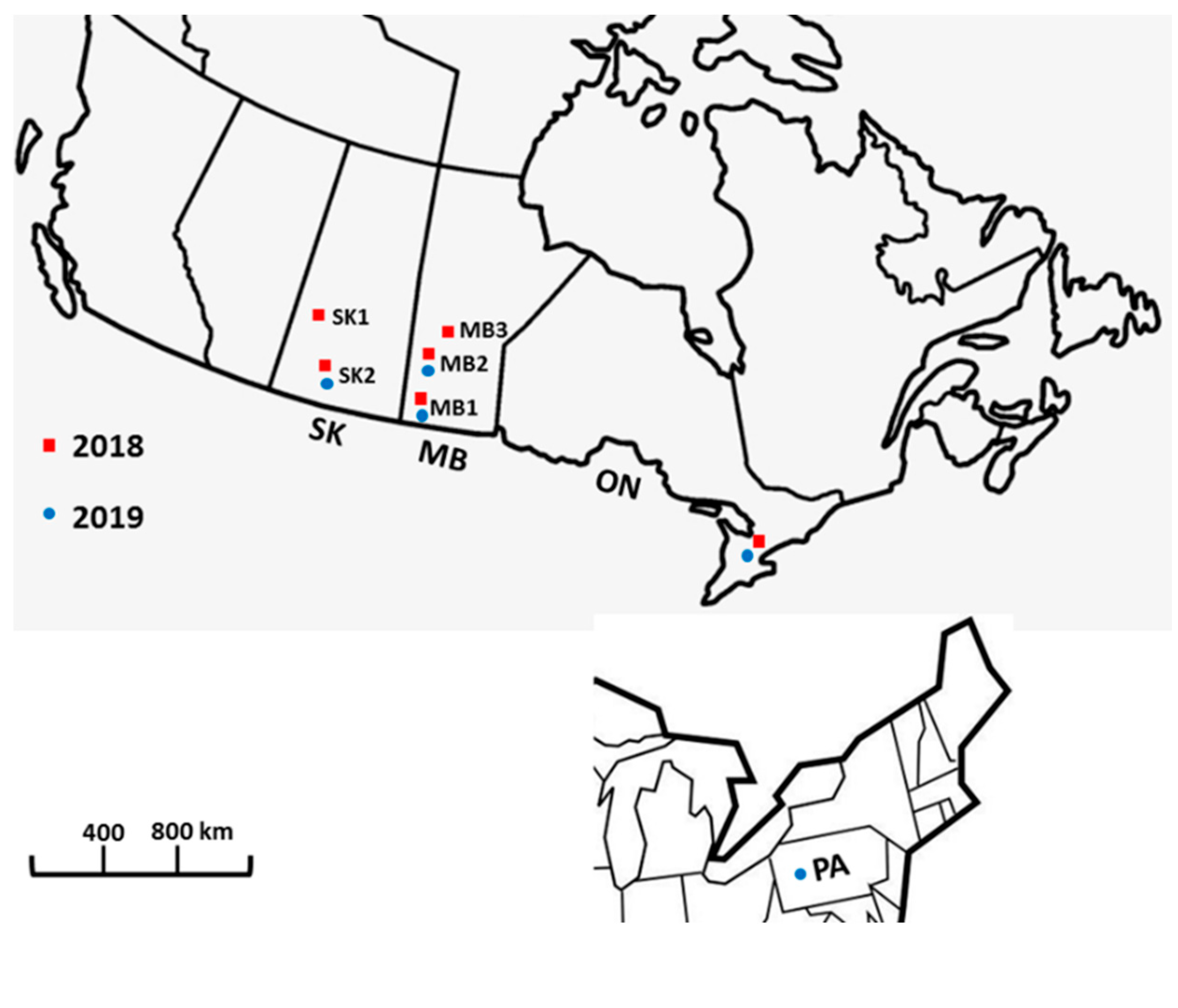
Epidemics of FHB in western Canada started in the early 1980s when grain samples of wheat from Manitoba showed symptoms of bleached kernels and detectable levels of DON mycotoxin [
11,
12]. Ever since, increasingly frequent FHB epidemic years occurred in Canadian prairies with the worst outbreak in 2016 in Saskatchewan (SK) and Manitoba (MB) [
13]. More recently, outbreaks have extended to the Province of Alberta. Comprehensive surveys of MB and SK wheat fields in the last three years have shown an FHB prevalence (proportion of fields affected) ranging from mild to very serious. One of the most recent epidemics occurred in 2016 with 87% FHB prevalence among wheat fields in Manitoba [
8,
14,
15]. High precipitation, relative humidity and warm temperatures during and after anthesis favors FHB development in wheat [
16]. Epidemics of FHB is also enhanced by rapid evolution of
Fusarium pathogens and the ability of the disease to spread through airborne ascospores over large areas [
17]. These important factors increase the risk of the development of FHB epidemics in the future.
In Canada, FHB outbreaks lead to serious losses in terms of yield coupled with grain contaminated by mycotoxins. Moderately resistant FHB cultivars have been developed through breeding efforts and are used extensively in western Canada; these include AAC Brandon, Cardale and Carberry [
18] with AAC Brandon being the predominant cultivar with 66.2% of the acreage in Manitoba in 2019 [
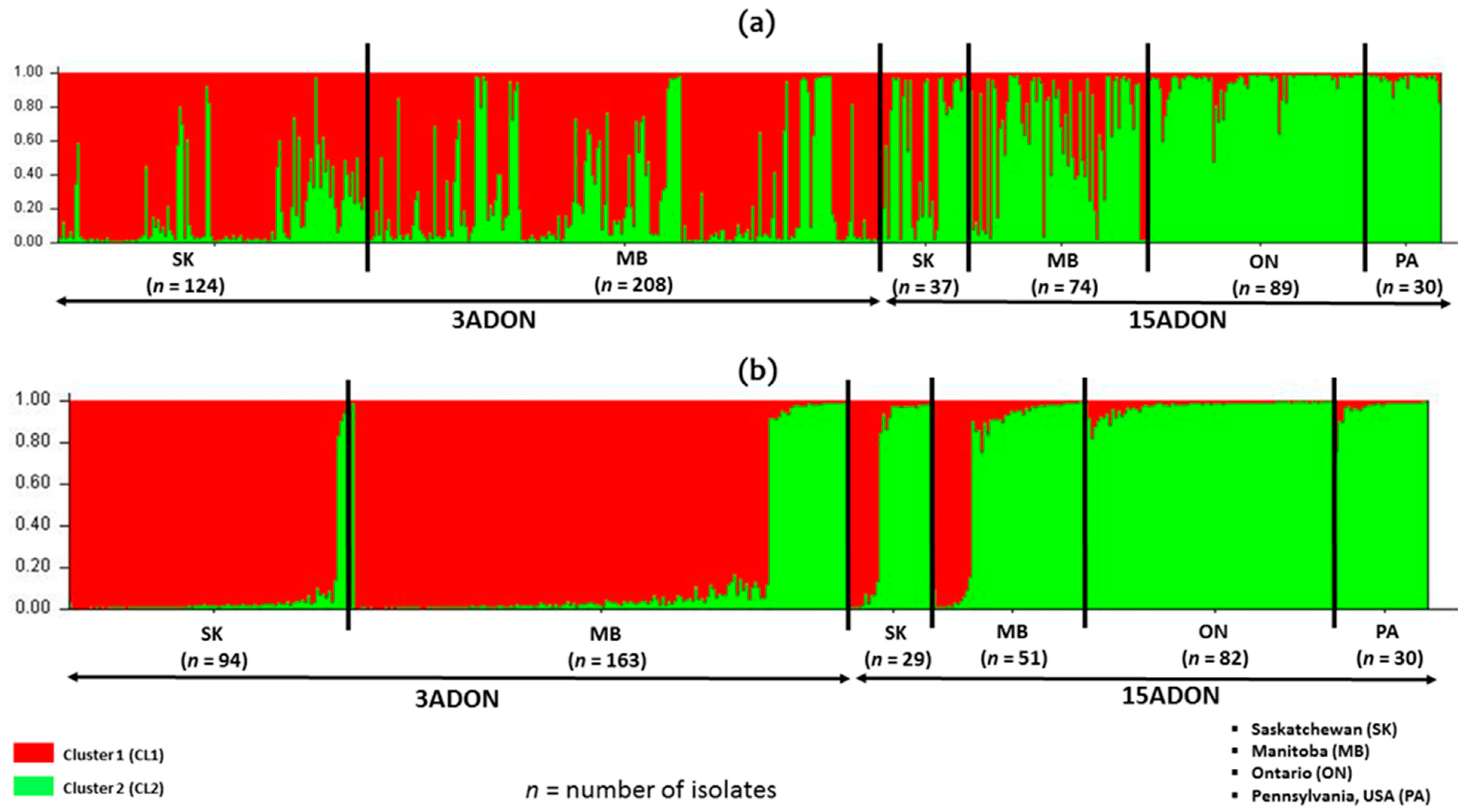
19]. Despite the moderate resistance achieved in many cultivars, the percentage of diseased spikes can increase dramatically in severe epidemics. Several surveys and molecular mycotoxin profiling over the last 20 years have shown a temporal and spatial increase in the 3ADON isolates compared to the 15ADON isolates, with a 3ADON vs 15ADON frequency of 10.7% and 31% 3ADON in Saskatchewan and Manitoba, respectively, between 1984 and 2004 [
7,
9,
20]. However, these studies did not consider the mycotoxin sub-structuring profile within the geographic locations in the provinces in Canada.
Population biology studies are critical in providing information that can be utilized to predict disease epidemics and also assist in setting up trials in developing FHB resistant cultivars. Genetic variation in a population is a strong indicator of frequent sexual reproduction and the rapid evolution of a pathogen. Such populations with high genetic variation can quickly adapt to resistant cultivars and fungicide modes of action [
21]. The ability of genes or genotypes to migrate to new geographic areas can be predicted by the amount of gene flow between populations [
22]. Surveys of
Fusarium species associated with FHB in wheat across Canada have shown
F. graminearum to be predominant at up to 90% of the
Fusarium spp. [
7,
23,
24]. Population genetic studies of
F. graminearum collected from wheat in Canada are few and either limited to a single province or performed with isolates collected about 15 years ago. A large-scale population structure study of Canadian isolates collected in 2005–2007 was investigated by [
7] revealing a one-fold increase in the 3ADON populations in 2005–2007. Population genetic studies in Manitoba was carried out on isolates collected in 2004–2005 [
23,
25]. A more recent study in the Province of Ontario utilized isolates collected in 2010–2012 [
24]. Pathogen populations can be dynamic in terms of evolutionary rates over space and time, and coupled with changing agronomic practices, can have a huge influence on pathogen population dynamics in a particular geographic location. It is therefore necessary to monitor
F. graminearum populations in wheat to detect changes in population structure and understand how these changes can influence the proliferation and development of FHB.
Molecular markers that are unambiguous and polymorphic are important tools in the study of the population biology of plant pathogens [
21]. Several genetic markers have been utilized to study genetic diversity in
F. graminearum populations. These include random amplified polymorphic DNA, restriction fragment length polymorphism, amplified fragment length polymorphism, microsatellite or simple-sequence repeats and sequence-related amplified polymorphisms [
23,
26,
27,
28,
29]. For this study, we used the variable number of tandem repeats (VNTR) developed by [
30] from the genome of
F. graminearum PH−1 (NRRL31084). These markers are more suited to genetic diversity studies than other markers because of the simplicity of polymorphic data collection [
30].
We hypothesize that the mycotoxin chemotype pattern of F. graminearum in wheat is reflected in the population structure within and between provinces in western Canada. The specific objectives were to: (i) investigate the genetic variability and population structure of F. graminearum, and the patterns of relatedness of the populations in Manitoba and Saskatchewan, (ii) investigate the level of gene flow and to determine the correlation of genetic distance with genetic identity, and (iii) determine the DON chemotype distribution within and between the provinces and their relationship with population genetic structure.
3. Discussion
Fusarium graminearum is an economically important plant pathogenic fungus because it is the primary causal agent of FHB in small grain cereals. In Canada, it causes substantial damage to wheat. The optimum conditions for the proliferation of FHB are warm temperatures and high (>90%) relative humidity during flowering [
34]. This is a strong indication that the pathogen will continue to be a subject of interest in the future due to the impact of climate change and global warming [
35]. In the present study, we used VNTR markers to analyze and characterize the population genetic diversity and structure of
F. graminearum isolates collected in three provinces in Canada and in the northeastern USA.
The
F. graminearum VNTR markers have been widely used in
F. graminearum population genetic analysis [
7,
9,
24,
36,
37,
38,
39,
40,
41,
42,
43]. In the current study, a high level of polymorphism was detected as indicated by the relatively high Shannon Index values across the geographic regions. This was also identified in the ten VNTR markers used in this study, which proved that it is an appropriate type of marker system for population genetic studies of
F. graminearum. The high polymorphism and unique haplotypes (98.6%) identified showed the high genetic diversity and very limited clonal populations of
F. graminearum in wheat in Canada.
The present work is the most recent wide-scale study of
F. graminearum population structure in Canada. Previous population genetic analysis of
F. graminearum in Canada has been done on isolates collected in 2001–2008 [
7,
9,
23]. Our study indicates very high gene flow among isolates from different geographic regions in the two neighboring western provinces: Saskatchewan and Manitoba. The high gene flow points to subpopulations within the two provinces that are part of a larger population that allows for frequent random mating. High gene flow between the eastern Canadian province of Ontario and PA in northeast USA also indicates they probably belong to one population. However, some level of genetic structure was observed for the two trichothecene chemotypes (3ADON and 15ADON) irrespective of geographical locations. This is in agreement with previous results of the clustering of 3ADON and 15ADON chemotypes across Canada [
7,
9] and the USA [
27].
Previous reports have shown that the majority of
F. graminearum isolates in wheat in Canada exhibit two longitudinal clines explaining the distribution of the 3ADON and 15ADON chemotypes. The 15ADON chemotype is predominant in eastern Canada (Ontario and Quebec) where the ratio of 15ADON to 3ADON remained relatively constant over the last 15–20 years [
7,
24]. In contrast, the 3ADON chemotype is predominant in the western provinces of Canada (Alberta, Saskatchewan and Manitoba) and there has been a temporal and spatial increase in 3ADON isolates over the last 20 years [
7,
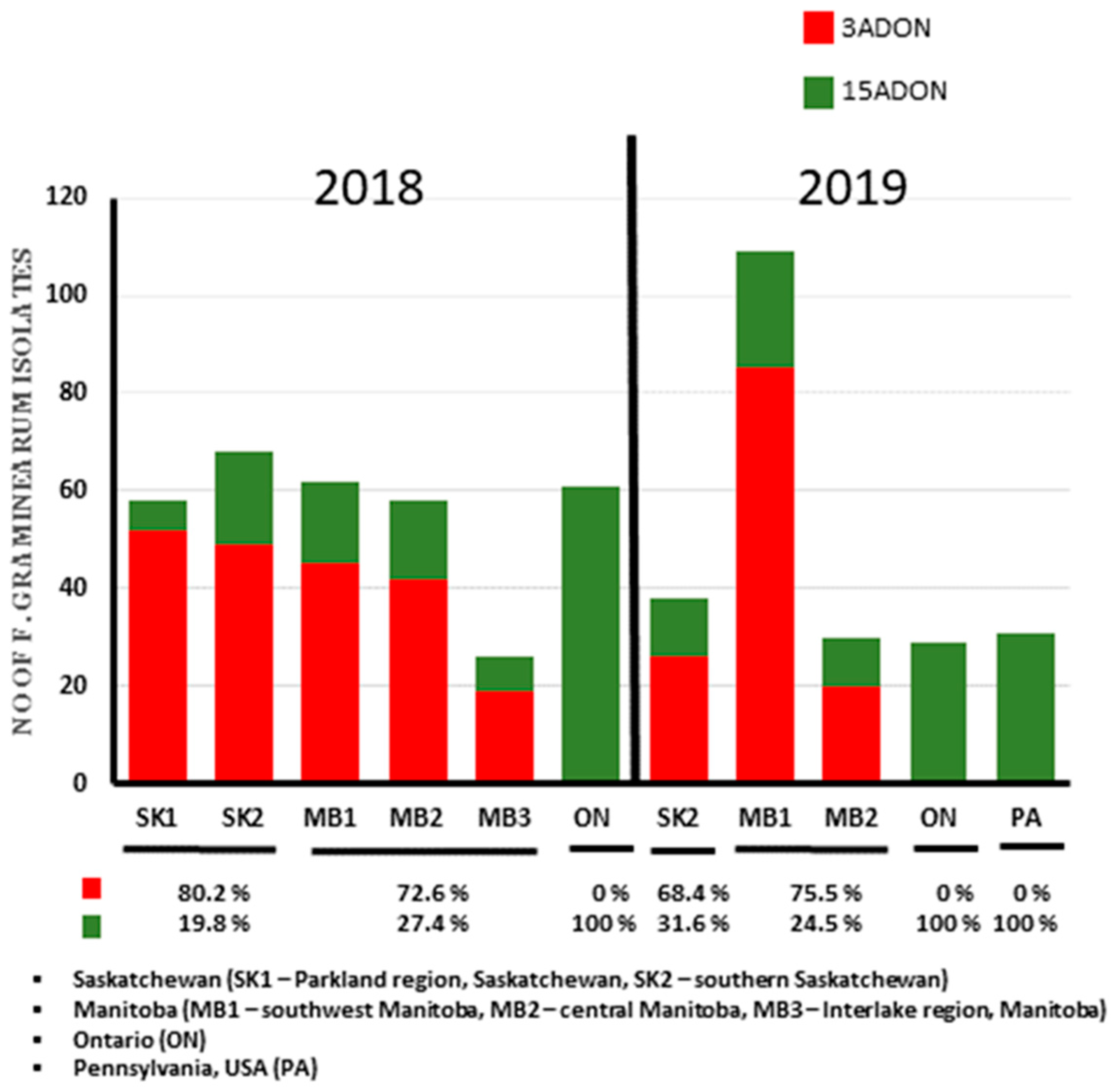
9]. Results from this study support the temporal increase in 3ADON chemotypes in western Canada as over 70% of the isolates were of the 3ADON genotypes. Ref. [
9] reported a 10.7% and 31% 3ADON chemotype percentage in 2004 for Saskatchewan and Manitoba. Breakdown of the results of this study show that the 3ADON chemotype increased in Saskatchewan more than 6-fold between 2004 (10.7%) and 2019 (68.4%) and increase in Manitoba 3ADON more than 2.5-fold between 2004 (31%) and 2019 (75.5%). Ref. [
23] reported 33.7% 3ADON isolates from 291 isolates of
F. graminearum, while the 3ADON percentage in this study of 570 isolates was above ≥70% in both 2018 and 2019. In addition, two regions sampled by [
23] from Manitoba (Towns of Killarney and Cartier), which had ≥72% 15ADON composition had a chemotype shift that is now ≥75% 3ADON as recorded in this study. The exact reason for this chemotype shift is a subject of debate but may be due to adaptation as suggested by [
9], as the 3ADON isolates have been reported to be more aggressive than their 15ADON counterparts. The higher admixture level observed in the 3ADON isolates might play a role in its higher virulence compared to the 15 ADON isolates [
39]. Fungal pathogens with high admixture can significantly influence and speed the evolution of virulence [
44].
The chemotype composition of
F. graminearum in western Canada was the opposite of that observed in eastern Canada. The 15ADON chemotype is predominant in eastern Canada and PA in the United States. Ref. [
24] and [
45], respectively, reported a 98% and 97% 15ADON composition of
F. graminearum isolates collected from wheat in the Province of Ontario in 2008–2013. After extensive sampling and chemotyping in the mid-west United States, researchers have reported a high 15ADON composition [
46,
47,
48]. Interestingly in this study, all isolates collected from wheat in Ontario and Pennsylvania (northeastern USA) were of the 15ADON chemotype. The high proportion of 3ADON isolates recorded in both Saskatchewan and Manitoba in this study emphasizes the need for regular temporal and spatial monitoring of trichothecene chemotypes across Canada. In addition, Bayesian model population structure analysis identified a different population structure that is associated with trichothecene chemotype differences. This study as well as previous studies on chemotype populations have demonstrated a significant temporal chemotype change in western Canada, which might be strongly linked to differences in local selective pressures among provinces in Canada. This further emphasizes the need for regional solutions to FHB management.
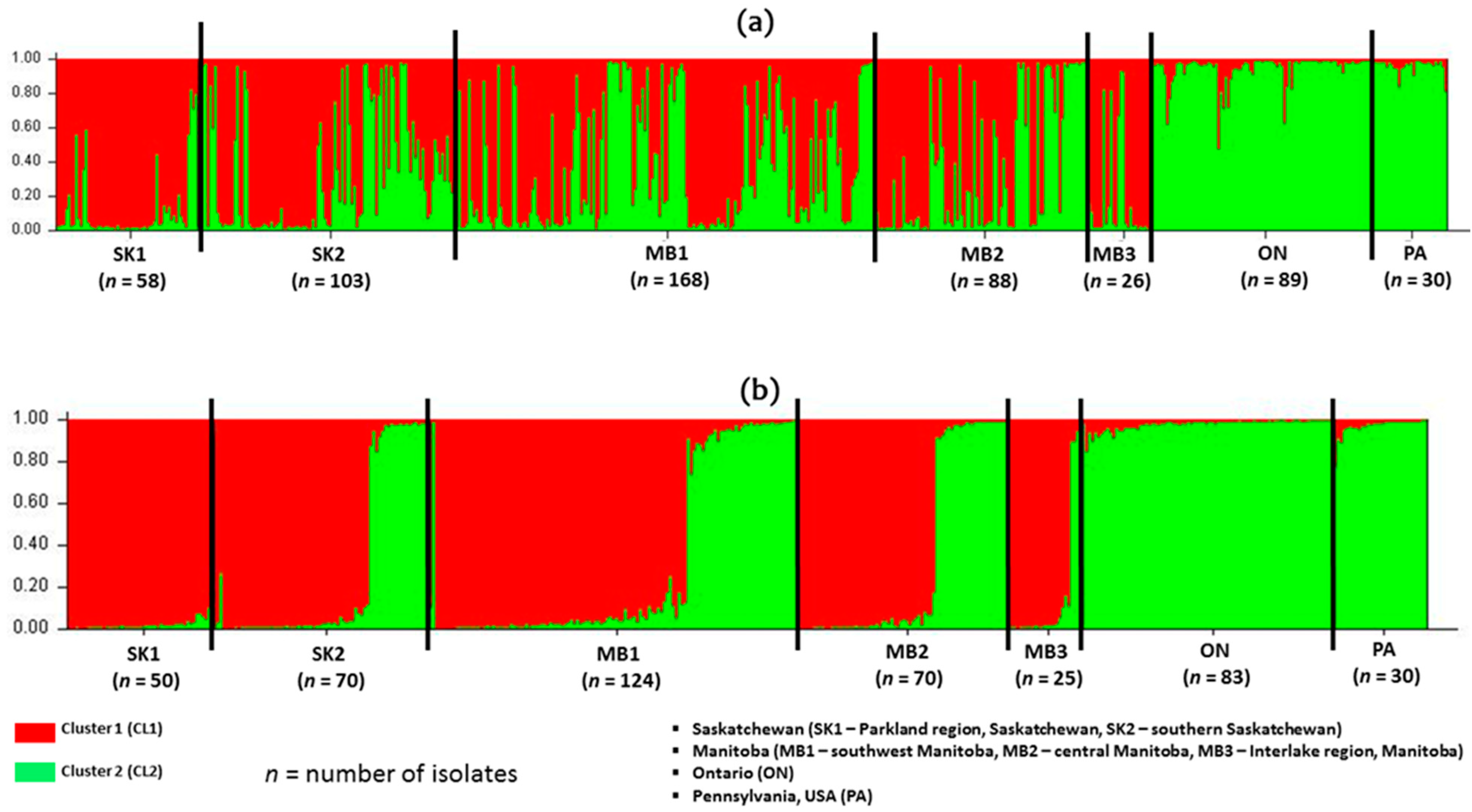
Spatial variability and relatedness of populations from the seven geographic regions were inferred from the genetic identity and genetic distance. Our population genetics analysis suggested that the five
F. graminearum populations (SK1, SK2, MB1, MB2 and MB3) from western Canada were genetically similar. This is mainly supported by the high gene flow (
Nm, 4.971–21.750) and low genetic differentiation (
FST, 0.011–0.048) among regions. Lower gene flow and relatively high genetic differentiation (
FST, 0.026–0.082) between the populations in the western provinces and that in Ontario and PA indicated a genetic structure between eastern and western regions. A similar correlation between genetic distance/identity and geographic location was reported for
F. graminearum isolates collected from Alberta, Saskatchewan and Manitoba [
49]. Ref. [
23] also found high gene flow (
Nm = 11.176) among 15 subpopulations of
F. graminearum in Manitoba. Similarly, high gene flow (
Nm = 19.483) was reported among
F. graminearum populations collected in wheat fields in Alberta, Saskatchewan and Manitoba [
49]. Similar high gene was also recorded between populations from central and southwestern Ontario [
24]. The AMOVA results buttressed the points above, since very high genetic variation (96%) was found among individuals within the subpopulations compared with a very low genetic variation (4%) among the subpopulations. This suggests a greater probability of sexual reproduction among the
F. graminearum populations in the regions studied. There was no significant difference in the population structure of the geographic regions studied, which agrees with previous studies of
F. graminearum isolates collected across Manitoba using sequence-related amplified polymorphism [
23]. Trade-in wheat seed between the two provinces is likely a major contributor to the high gene flow between isolates from the different geographic regions. Long-distance spore transfer as shown for
F. graminearum [
50] might also be a factor responsible for the high gene flow.
The 3ADON and 15ADON trichothecene chemotype populations from this study appear to be homogenous for chemotype and genetically similar with STRUCTURE Bayesian modelling analysis of cluster isolates. Homogeneity is more visible in the 15ADON chemotype isolates than the 3ADON chemotype isolates. Ref. [
51] reported a similar homogenous grouping of 15ADON isolates from Canada, but the main reason for this homogeneity is unclear. This structuring might be limited as there is continuous gene flow between the two-chemotype populations, which may explain the chemotype shifts observed over the years in western Canada. The limited genetic structuring by trichothecene chemotype suggests populations that are not specific to either province or chemotypes might not be a very good indicators of population structure at the provincial level.
This study has provided current information on the population structure and chemotype diversity of
F. graminearum isolates in wheat in three important wheat-producing provinces in Canada and on a state in the USA. The class B-Trichothecene profile of
F. graminearum isolates world-wide recently reviewed by [
52] is very variable in different regions. Management strategies should be geared towards chemotyping studies in local areas in order to correctly estimate and predicts risks in the face of climate changes and agronomic practices like crop rotation. Genetic analysis using VNTR data has revealed high levels of gene flow and genetic variation within the regions studied in the Provinces of Manitoba and Saskatchewan. The higher genetic diversity in Manitoba compared to Saskatchewan might result in more
F. graminearum populations with better adaptation to current control strategies including resistant cultivars and fungicides. This is a result of constant sexual recombination between isolates that may suggest a drive towards more pathogenic
F. graminearum isolates through mating. The
F. graminearum isolates in this study were correlated with trichothecene chemotypes (3ADON and 15ADON) according to stringent genetic structure analyses. An understanding of the correlation of chemotype between or among populations and genetic structure would require more in-depth studies of thousands of isolates collected more recently from across Canada and the United States. In addition, this should contribute to the identification of any fitness attributes that might be associated with the frequency of chemotype shifts observed in this study. Finally, this information on genetic variation among subpopulations will contribute to genetic resistance breeding in wheat, as well as help in mycotoxin risk assessment and the monitoring the of FHB infected seed across Canada.

 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}