A Novel Microbial Zearalenone Transformation through Phosphorylation
by
 ,
,
Yan Zhu
1, 
Pascal Drouin
2,
Dion Lepp
1,
Xiu-Zhen Li
1,
Honghui Zhu
1,
Mathieu Castex
2 and
Ting Zhou
1,* 1
Guelph Research and Development Centre, Agriculture and Agri-Food Canada, Guelph, ON N1G 5C9, Canada
2
Lallemand Inc., Montréal, QC H1W 2N8, Canada
*
Author to whom correspondence should be addressed.
Toxins 2021, 13(5), 294; https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13050294
Submission received: 1 April 2021
/
Revised: 16 April 2021
/
Accepted: 19 April 2021
/
Published: 21 April 2021
(This article belongs to the Special Issue Innovative Mycotoxin Detoxification: Discoveries, Mechanisms and Applications)
Abstract
:Zearalenone (ZEA) is a mycotoxin widely occurring in many agricultural commodities. In this study, a purified bacterial isolate, Bacillus sp. S62-W, obtained from one of 104 corn silage samples from various silos located in the United States, exhibited activity to transform the mycotoxin ZEA. A novel microbial transformation product, ZEA-14-phosphate, was detected, purified, and identified by HPLC, LC-MS, and NMR analyses. The isolate has been identified as belonging to the genus Bacillus according to phylogenetic analysis of the 16S rRNA gene and whole genome alignments. The isolate showed high efficacy in transforming ZEA to ZEA-14-phosphate (100% transformation within 24 h) and possessed advantages of acid tolerance (work at pH = 4.0), working under a broad range of temperatures (22–42 °C), and a capability of transforming ZEA at high concentrations (up to 200 µg/mL). In addition, 23 Bacillus strains of various species were tested for their ZEA phosphorylation activity. Thirteen of the Bacillus strains showed phosphorylation functionality at an efficacy of between 20.3% and 99.4% after 24 h incubation, suggesting the metabolism pathway is widely conserved in Bacillus spp. This study established a new transformation system for potential application of controlling ZEA although the metabolism and toxicity of ZEA-14-phosphate requires further investigation.
Key Contribution: The discovery of the new ZEA transformation system, including the novel transformation pathway and the bacterial isolate (Bacillus sp. S62-W), provides a potential for a new ZEA detoxification strategy.
1. Introduction
Zearalenone (ZEA), chemically characterized as a macrocyclic β-resorcyclic acid lactone, is an estrogenic mycotoxin produced by Fusarium spp. [1]. ZEA contamination occurs in maize and other crops such as barley, oat, wheat, sorghum, millet and rice [2]. Silage, moldy hay and grain used as concentrates in animal feeds may also contain the toxin carried over from contaminated crops [3,4]. According to the European Food Safety Authority (EFSA) [5], ZEA was detected in 15% of 9877 unprocessed grains and 13,075 of food samples collected in 19 European countries between 2005 and 2010. In that survey the average ZEA level detected in unprocessed corn was 87 µg/kg with the maximal concentration as high as 2700 µg/kg. Ensiling is a common agricultural practice for the preservation of green fodders intend for livestock feed by fermentation. Although ZEA-producing fungi are inhibited by low pH and anaerobic conditions, contamination by ZEA is not mitigated during the process [6]. Moreover, these conditions (i.e., acidic and anaerobic environment) bring challenges for ZEA detoxification by microorganisms.
Strategies for ZEA decontamination have been investigated for several decades and can be categorized into chemical, physical, and biological approaches. Among them, efficient, specific, and environmentally friendly biological detoxification methods, which transform ZEA into less toxic metabolites, are being actively investigated [7,8]. Biological transformation of ZEA was reported in a variety of microorganisms such as bacteria, yeasts, or filamentous fungi (molds). These organisms included strains of Bacillus [9,10,11,12,13], Pseudomonas [14,15], Acinetobacter [16], Rhodococcus [17], Saccharomyces [18], Aspergillus [19,20,21], Rhizopus [19,22], Trichosporon [23], Clonostachys [24], and Thamnidium [25]. Among the identified metabolites, α-zearalenone (α-ZOL), β-zearalenone (β- ZOL), α-zearalanone (α-ZAL), and β-zearalanone (β- ZAL) have been shown to possess similar or higher estrogenic activity than ZEA. Only two conjugate metabolites (zearalenone-14-O-β-glucoside, and zearalenone-sulfate) [20,25] and two macrocyclic ring cleavage products [23,24] were demonstrated to have lower or no estrogenic effects using in vitro approaches.
The present work reported the discovery of a novel ZEA transformation pathway, ZEA phosphorylation, and the isolation, identification and characterization of a new strain of Bacillus sp. with ZEA phosphorylation functionality. In addition, the research demonstrated that multiple Bacillus species/strains also possess such functionality, opening the door for future applications of this newly discovered ZEA transformation mechanism.
2. Results
2.1. Isolation of ZEA Transforming Bacteria
ZEA reduction (>50%) was detected in seven of the 104 corn silage samples tested. One sample, labelled S62, showed 100% ZEA reduction. When tested in nutrient broth with mineral salts (NBM) at 28 °C for 5 days, the microbial consortium prepared from S62 transformed ZEA completely to a product identified below, even after being diluted 104-fold. The active microbial consortium was then used for isolating single colonies by dilution plating on NBM plates. One of the selected single colonies, named S62-W, showed 100% ZEA transformation activity after 5 d incubation at 28 °C. When tested under the same temperature conditions with 120 RPM shaking, S62-W completely transformed ZEA within only 24 h.
2.2. Taxonomic Identity of the Active Isolate S62-W
Phylogenetic analysis based on the 16S rRNA gene sequence putatively identified the active isolate S62-W to the genus Bacillus, with 99% identity to sequences from the Bacillus pumilis group [26], including B. aerius, B. stratosphericus, B. altitudinis, B. pumilus, and B. safensis (Table 1). Further identification of S62-W using whole genome sequencing was initially performed with the JSpeciesWS Tetra correlation search (TCS) tool, which compares a query genome against a database of reference genomes based on tetranucleotide frequencies. This method identified 19 reference genomes with a Z-score higher than 0.999, which were all identified as Bacillus spp., including B. stratosphericus, B. pumilus, B. altitudinis and B. cellulasensis. The average nucleotide identities between S62-W and the top fourteen TCS hits, based on BLAST alignments are given in Table 2. Phylogenetic analysis based on core genome sequence alignments of S62-W and 39 Bacillus reference genomes placed S62-W in a separate cluster from B. pumilus and B. safensis, in contrast to the 16S rRNA gene-based results, and within a cluster consisting mainly of B. stratosphericus and B. altitudinis strains, indicating that it is more closely related to these two species (Figure 1).
2.3. Identification of ZEA Transformation Product
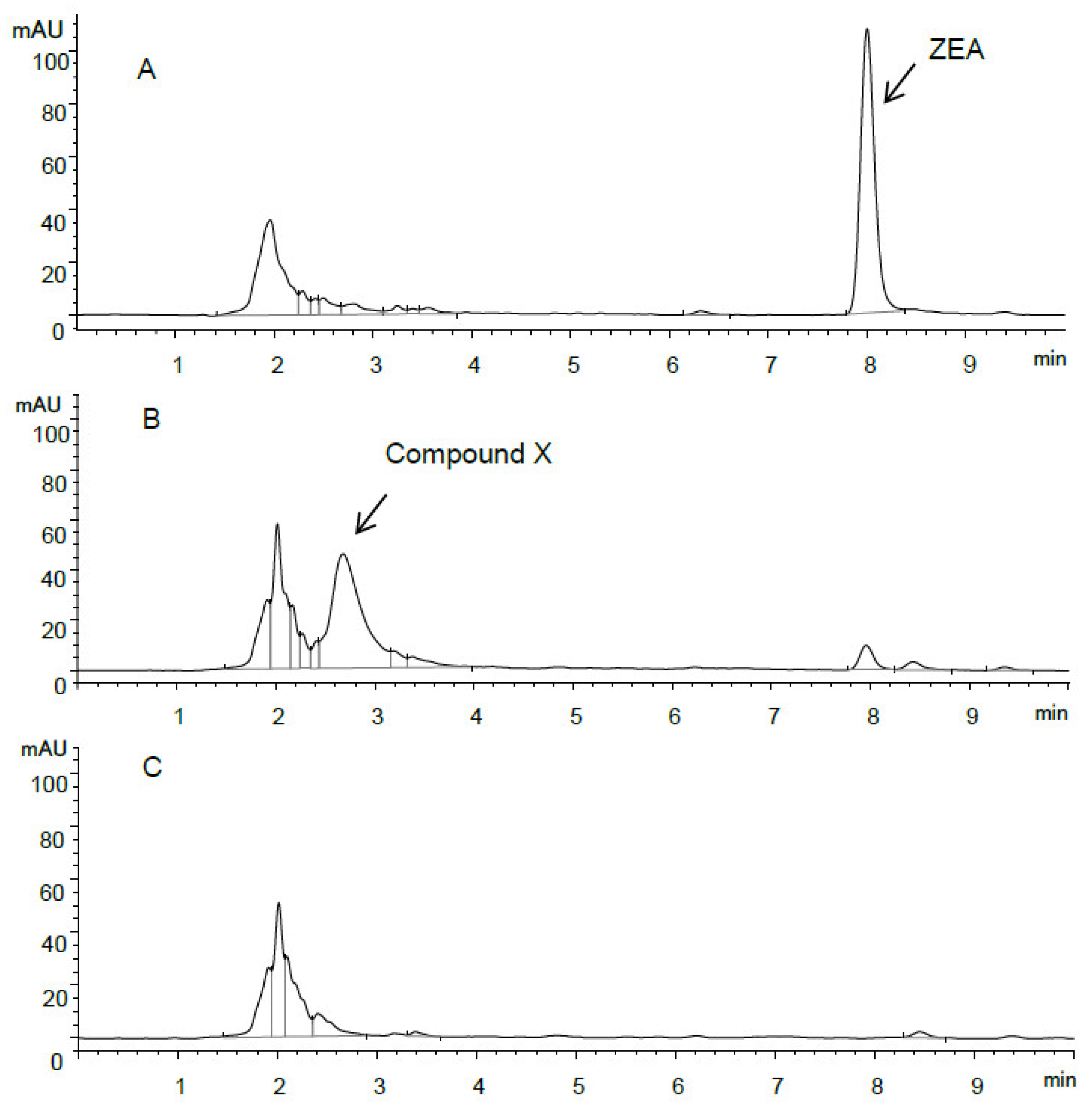
HPLC chromatograms of ZEA and ZEA transformation products are shown in Figure 2. Compared to the initial culture spiked with 50 µg/mL of ZEA (Figure 2A), the presence of an unknown peak was reported together with the decrease of the peak corresponding to ZEA after 24 h incubation (Figure 2B). Given that the peak was not observed in the control sample without ZEA (Figure 2C), this indicated that the corresponding ZEA transformation product, compound X, was related to the metabolism of ZEA, resulting from ZEA transformation by the isolate S62-W. The earlier retention time suggested that compound X was characterized as having a higher polarity than ZEA.
In the UV spectra of ZEA and compound X (Figure 3), both compounds showed similar triple absorption peaks in the UV wavelength range, although the hypsochromic shift of compound X was observed. This observation suggested that compound X shares a similar structure to ZEA.
The LC-MS/MS analyses of the standard ZEA and purified compound X are presented in Figure 4. Compared to the major m/z signal of ZEA (317.14), a higher m/z signal of 397.10 was observed in the MS spectrum of compound X, which indicated that it is probably a ZEA conjugate. The major m/z signal of 78.96 in the MS/MS spectrum of fragment ions originally from the precursor ion of ([C18H23O8P]-H)− revealed a substitution of a phosphate group of ZEA.
The purified compound X was analyzed by nuclear magnetic resonance (NMR). The 1D1H and 1D13C NMR spectra, along with correlations in the associated 2D NMR spectra, were used to assign resonances to all non-exchangeable 1H and 13C nuclei. The 2D correlations in the associated COSY, HSQC, HMBC and ROESY spectra were used to determine the structure of compound X. The results determined that the main structure of compound X (except for the phosphate group) is identical to that of ZEA (Table 3). A detail of particular importance was a ROESY correlation between H11 and H13 and an HMBC correlation between H12 and C13. These were used to distinguish the olefin substitution site at the phenyl ring of compound X and to confirm the assignment of the H13 and H15 phenyl ring protons at 6.75 ppm and 6.65 ppm, respectively. HMBC correlations between both H13 and H15 to the 13C resonance at 157.33 ppm were used to positively identify this resonance as arising from the C14, the oxygen-substituted phenyl ring carbon that is para to the ester substitution site on the phenyl ring of compound X. The 3JHH COSY correlations, along with HMBC correlations between H6 to C8 and C7, H8 to C7, and H19 to C4, helped to differentiate the methylene 1H and 13C resonances in the aliphatic portions of compound X and to distinguish the C7 ketone 13C resonance (216.18 ppm). Further HMBC correlations between H13 and a 13C resonance at 131.54 ppm helped to establish this 13C resonance as belonging to the olefinic C12 carbon directly bound to the phenyl ring of compound X. The ester carbonyl resonance at 171.27 ppm was identified via HMBC correlations to H3 (5.01 ppm), H15 (6.65 ppm), and H13 (6.75 ppm). In general, HSQC 1JCH crosspeaks were used to assign the remaining 13C resonances. The 1D31P NMR spectrum revealed a resonance of phosphate at 31P nucleus although the phosphate substitution site on the phenyl ring of compound X was not identified. The phosphate substitution site could be at either C14 or C16.
The retention time of compound X, synthesized ZEA-14-phosphate, and ZEA-16-phosphate in HPLC chromatograms were 17.8, 17.8, and 10.1 min, respectively, which indicated that the structure of compound X and ZEA-14-phosphate is probably identical. With the combination of HPLC, LC-MS/MS and NMR analyses, the compound X is confidently identified as the phosphate conjugate at C14 position of ZEA (Figure 5), which is named ZEA-14-phosphate (ZEA-14-P).
2.4. ZEA Transformation by Bacillus sp. S62-W in Media
When tested with the initial concentration of ZEA and bacterial cells at 25 µg/mL (78.5 µmol/L) and 2.5 × 107 cfu/mL, respectively, the decrease of ZEA was found to be positively correlated with the optical density value of Bacillus sp. S62-W cells in the culture in both static and shaken incubations (Figure 6A). This observation indicated that the transformation was mainly related to the primary metabolism of the bacterial isolate, indicating that the cell growth of Bacillus sp. S62-W could facilitate ZEA transformation.
The transformation product, ZEA-14-phosphate (i.e., compound X), was detected and quantified by HPLC. A negative correlation between the concentrations of ZEA and ZEA-14-phosphate was observed (Figure 6B). Since no intermediate compounds were detected, this suggests that ZEA was fully transformed into ZEA-14-phosphate by Bacillus sp. S62-W. The efficiency of this transformation was high since 100% of ZEA was transformed within 24 h.
2.5. Effects of Environmental Factors on ZEA Transformation
Environmental factors, including culture media, pH, temperature, oxygen requirement and initial mycotoxin concentration, had different effects on the ZEA transformation by Bacillus sp. S62-W. Figure 7A shows the level of ZEA transformation in different media. Full transformation activities were observed in corn meal broth (CMB) and alfalfa silage extract (ASE). Corn silage extract (CSE) supported a lower transformation rate (80%), likely due to its higher concentration of organic acids and lower level of nutrients.
The pH values affected the ZEA transformation capacity (Figure 7B). Complete transformation was achieved within 24 h at 28 °C under pH values ranging between 6.0 and 10.0. At a lower pH of 4.0, however, noteworthy transformation appeared only after 120 h of incubation. The results revealed that the acid tolerance of Bacillus sp. S62-W was greatly reduced at pH below 5.0 and the ZEA transformation activity by the bacterial isolate was low in acid medium.
Since temperature in silage may reach as high as 40 °C upon oxygen ingress, the capability of Bacillus sp. S62-W to transform ZEA was evaluated at a broad range of temperatures between 22 °C and 42 °C (Figure 7C). Under all temperatures tested, more that 95% of ZEA was transformed to ZEA-14-phosphate after 24 h static incubation, indicating that the bacterial isolate can still be active under relatively high temperatures.
The oxygen requirement for ZEA transforming activity was evaluated (Figure 7D) as the ensiling process, comprising both aerobic (aerobic phase and feed-out phase) and anaerobic stages. The full transformation activity was reached under aerobic incubation. On the contrary, less than 10% ZEA was transformed under anaerobic conditions. However, transformation activity can be restored by aerobic incubation. Complete transformation was achieved after incubating the bacterial strain under anaerobic condition for 6 d following by a 2 d aerobic incubation. Although the isolate may not function during the absolute anaerobic stage, it may potentially exhibit transformation activity in the early stages of ensiling immediately after inoculation of the harvested forage, as well as during short exposure to air once the silage is open at feed-out.
Although the ZEA contamination levels in crops rarely exceed 5 mg/kg, the contaminated areas invaded by mycotoxin-producing fungi may accumulate higher concentrations of ZEA. Thus, the maximum capacity of ZEA transformation of S62-W was investigated. After 96 h aerobic incubation in NBM, more than 80% of ZEA transformation was observed in the samples spiked with up to 100 µg/mL of ZEA. In the sample with 200 µg/mL of ZEA, the transformation ratio was still greater than 50% (Figure 7E).
2.6. Determination of ZEA Phosphorylation in Various Bacillus Strains
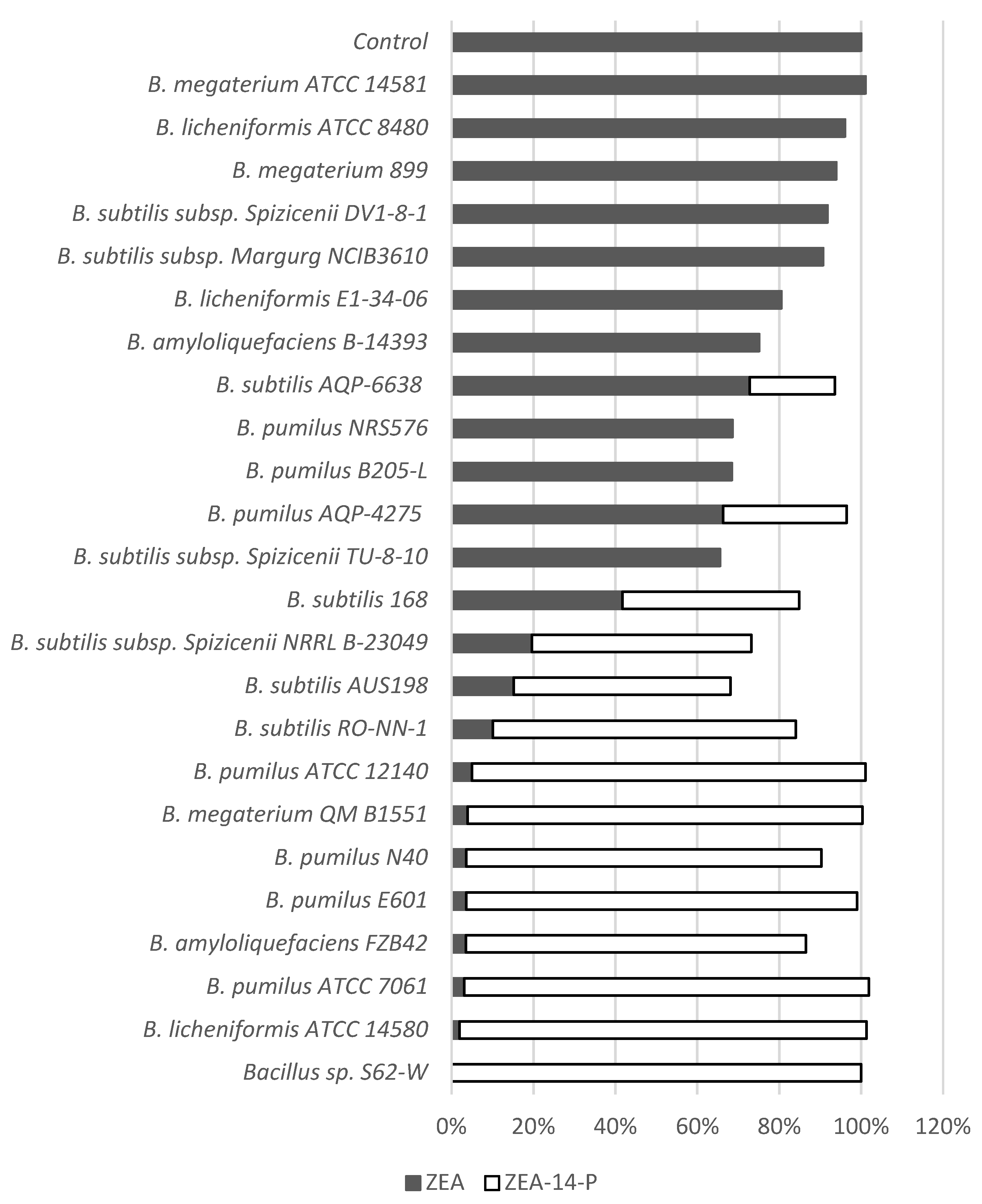
Among the 23 strains of Bacillus tested, 13 exhibited phosphorylation activity at an efficacy between 20.3% and 99.4% after 24 h incubation at 28 °C, suggesting that the metabolism pathway is widely conserved among Bacillus spp. Five Bacillus spp. including B. licheniformis ATCC 14580, B. pumilus ATCC 7061, B. megaterium QM B1551, B. pumilus ATCC 12140, and B. pumilus E601 possess high phenotypic capability (>95%) to transform ZEA to ZEA-14-phosphate (Figure 8). Two commercial strains, B. pumilus AQP-4275 and B. subtilis AQP-6638 displayed partial transformation of 30.2% and 20.8%, respectively. B. subtilis subsp. Spizicenii NRRL B-23049 and B. subtilis AUS198, reduced 80.4% and 84.8% of spiked ZEA, but only 53.6% and 52.9% of ZEA-14-phospate were detected after incubation. The notable difference (more than 20%) between the decrease of ZEA and production of ZEA-14-phosphate indicated unidentified intermediate or final products in the culture.
3. Discussion
The disappearance of ZEA through the metabolism of Bacillus spp. has been reported in many studies [9,10,11,12,13]. However, the lack of a degradation mechanism limited further investigations and application since the identification and safety assessments of the transformation products are essential to develop an effective biological detoxification system [27]. In our study, the phosphorylation of ZEA revealed a novel transformation mechanism through Bacillus spp. Phosphorylation is critical for many cellular processes through the addition of a phosphoryl group to proteins, sugars, lipids, and other molecules, during many metabolic and cell signaling pathways [28,29]. However, the metabolism of xenobiotic compounds through phosphorylation has rarely been reported compared to other conjugations [28]. Two isoflavones, daidzein and genistein, were found to be transformed to daidzein-7-O-phosphate and genistein-7-O-phosphate through attachment of a phosphate moiety to the phenolic hydroxyl group by B. subtilis [30], which represents some of the limited examples of phosphate conjugates produced by bacteria. The well-known phase II reactions of xenobiotic metabolism in mammals include glucuronidation, glycosidation, sulfation, methylation, acetylation, amino acid conjugation, glutathione conjugation, and fatty acid conjugation [31]. Phosphorylation is not typically included in these reactions, although rare cases (e.g., phosphorylation of chloramphenicol and 2-acetamidofluorene) have been reported [28]. Conjugated products of ZEA through microbial metabolic processes are limited to ZEA-14-O-β-glucoside and ZEA-sulfate, which were transformed by Rhizopus sp. and Aspergillus niger, respectively [20,25]. In this study, for the first time, the phosphorylated product of ZEA resulting from bacterial metabolism has been identified and purified. This distinctive and abnormal xenobiotic transformation enriches our knowledge of microbial conjugation reactions.
One issue that hinders the successful application of mycotoxin detoxification using a biological strategy is the assessment of functional microbes for safety. Bacilli and most lactic acid bacteria are good candidates for food-related applications, since several species are included in the EFSA Qualified Presumption of Safety list, which can be used as a source of feed additives [32]. In the current study, the bacterial strain S62-W belongs to the genus Bacillus and exhibits efficient ZEA transformation activity. More significantly, the main transformation product has been clearly characterized. Combined with its advantages of acid tolerance (down to a pH of 4.0), working at a range of temperatures (22-42 °C), and the capability of transforming high levels of ZEA (up to 200 µg/mL), Bacillus sp. S62-W may be applied to various feedstuffs including silage in the postharvest phase and during feed-out. In order to investigate the specificity of this phosphorylation mechanism in Bacillus, 23 Bacillus strains representing various species were selected and their transformation capacities tested in vitro. The widely positive results among various strains suggest that ZEA phosphorylation activity is common to phylogenetically related strains in the same sub-groups of Bacillus, based on 16S rDNA analysis [33]. However, several strains of the same species differed in their transformation capacity, exhibiting either low or high activity, indicating that the function is also strain specific.
Bacillus spp. are ubiquitous in soil and are frequently present in silage [34]. The spore-forming capacity of Bacillus contributes to its survival under ensiling conditions, including a prolonged period in a low pH environment as well as anaerobic conditions. The endospores germinate and the cells grow in the silage when the pH and oxygen concentration are appropriate [35,36]. Queiroz et al. [37] proposed that exposure to air results in a greater amount of Bacillus endospores in the outer layers of silage bales than the central and tightly-packed layers. The aerobic deterioration caused by the oxidation of lactic acid by yeasts also supports the growth of Bacillus spp. [37]. Although the role of Bacillus spp. in silage has been generally considered to be negative, they may improve the aerobic stability of silage and produce antimicrobials to inhibit fungal growth [35]. Improvements in aerobic stability were observed after inoculation of B. subtilis along with Lactobacillus plantarum [38]. In addition to the adaptation in the ensiling procedure, the Bacillus spp. also provided supplementary benefits as feed additives. Some Bacillus spp. with ZEA mitigation activities have been shown to possess probiotic characteristics, including tolerance to the gut environment and anti-pathogenic capabilities [11,39]. As an affordable source of enzymes, Bacillus spp. have been widely applied in enzyme production [40]. B. licheniformis CK1, another reported ZEA degrading microorganism, displayed high levels of xylanase, CMCase and protease activities, which improved the digestibility of nutrients in feed [9]. From a safety perspective, Bacillus spp. are considered ideal mycotoxin mitigation agents due to their perceived probiotic properties, although some species (e.g., B. anthracis and B. cereus) are widely known as pathogens because of the production of enterotoxins [27,41]. Since Bacillus sp. S62-W was originally isolated from silage, it may be adapted to silage conditions and contribute to the ensiling process and quality. Meanwhile, the variously functional Bacillus spp. described in the present study, particularly the two commercial Bacillus strains, provide multiple options for potential industrial application based on their safety profile and additional benefits.
The toxicities of conjugated ZEA transformation products have been reported in both in vitro and in vivo studies. El-Shakawy and Abul-Hajj [42] carried out an estrogen receptor affinity assay using rat uterine cells to evaluate the estrogenic activities of various ZEA derivatives. The metabolites with a blocked 14-phenolic group (14,16-dimethoxy-ZEA and ZEA-14-O-β-glucoside) were found to be biologically inactive. Similar observations were reported for less estrogenic activities of ZEA-sulfate through MCF-7 cell proliferation assays [20]. Conjugated mycotoxins are often considered “masked mycotoxins” (i.e., biological modification of mycotoxins which may release parent mycotoxins through mammalian metabolism) although the term was recently defined as biologically modified mycotoxins that were conjugated by plants [43]. If conjugation results in decreased toxicity, however, it should be designated as detoxification rather than masking [44]. For example, the mammalian phase II reactions use a strategy to detoxify xenobiotic compounds by conjugating endogenous groups, which creates more acidic and polar compounds that have lower lipid-solubility, preventing them from diffusing into cell membranes and facilitating their elimination from the body [31]. An in vitro investigation of the stability of ZEA-14-O-β-glucoside in the human gut was performed [45]. Neither hydrolysis in the digestion juice of the upper gut, nor gut epithelial absorption in Caco-2/TC7 cells, were observed. Nevertheless, the conjugated ZEA was fully hydrolyzed to ZEA by the human gut microbiota and 40%–70% of the transformed ZEA was further metabolized to unknown compounds, which revealed the potential risks of conjugated mycotoxins [45]. As a novel conjugated mycotoxin, a comprehensive toxicity assessment of ZEA-14-phosphate will need to be performed not only to understand the in vitro estrogenic activity, but also its fate following ingestion by animals.
4. Conclusions
The discovery of a new ZEA microbial transformation system, including the novel transformation reaction (i.e., phosphorylation) and bacterial isolate (i.e., Bacillus sp. S62-W) provides an alternative strategy for ZEA detoxification. The current data demonstrates that there is great potential to use such system for commercial application to mitigate the ZEA contamination problem in silage and other agricultural commodities, although further research is necessary. The toxicity reduction of ZEA-14-phosphate needs to be demonstrated using cell culture assays and animal trials. Strategies to apply the transformation system under commercial conditions need to be developed in future studies.
5. Materials and Methods
5.1. Chemicals and Culture Media
ZEA was obtained from Sigma-Aldrich (St. Louis, MO, USA). ZEA-14-phosphate and ZEA-16-phosphate were chemically synthesized by TripleBond (Guelph, ON, Canada) according to the proposed ZEA transformation products in this study. NBM contained 8 g nutrient broth (BD, Mississauga, Canada), 2.5 g K2HPO4, 2.5 g KH2PO4, 1.0 g (NH4)2HPO4, 0.2 g MgSO4·7H2O, 0.01 g FeSO4, and 0.007 g MnSO4 per liter. Nutrient agar (NA) plates were prepared using 8 g of nutrient broth and 15 g agar per liter. CSE or ASE was prepared by blending 50 g corn or alfalfa silage with 200 mL of deionized water for two minutes and was filtered through a Whatman No. 1 filter paper (Whatman, Maidstone, Kent, UK). CMB was prepared by soaking 40 g of corn meal in 1 L of deionized water at 58 °C for 4 h. After standing for 2 h, the broth was filtered through a Whatman No. 1 filter paper and 3 g (NH4)2SO4, 1 g K2HPO4, 0.5 g MgSO4·7H2O, 0.5 g K2SO4, 0.01 g FeSO4, and 0.007 g MnSO4 were added. All the media were autoclaved at 121 °C for 15 min before using.
5.2. Screening of Corn Silage Samples
A total of 104 corn silage samples were collected from silos of various farms located in the states of Wisconsin, New York, Vermont, Michigan and Maine. Silage samples were screened for the ZEA transformation activity by spiking ZEA in the silage extracts. The silage extracts were prepared by weighing 50 g of each silage sample and mixing with 200 mL of sterile deionized water in a blender. The mixture was blended for 2 min and filtrated through a coarse filter paper. One hundred µL of sample was transferred into 900 µL of NBM or CSE containing 25 µg/mL of ZEA. Two incubation conditions (aerobic at 28 °C and 5% CO2 at 30 °C) have been applied for the screening. ZEA concentration and accumulation of ZEA transformation products were analyzed by HPLC and LC-MS methods after the incubation up to 30 days.
5.3. Single Colony Isolation and Purification
Samples showing decreased ZEA concentration were selected for single colony isolation. A series of 10-fold dilution was applied using saline solution (0.85% NaCl) and 100 µL of diluted culture was spread on NA plates. Plates were incubated aerobically at 28 °C for 2 days. A total of 100 colonies were randomly selected from the 10−5 dilution plates and transferred into NBM medium containing 25 µg/mL of ZEA. Samples were incubated aerobically at 28 °C for 5 days. The decrease of ZEA after incubation was detected by HPLC.
5.4. Taxonomic Identification of the Active Isolate
Genomic DNA was extracted from a single colony using InstaGene™ matrix (Bio-Rad, Mississauga, ON, Canada). The PCR primers used for 16S rRNA gene amplification were universal primers of 16S8F primer 5′ AGA GTT TGA TCC TGG CTC AG 3′ and 16S1391R primer 5′ GAC GGG CGG TGW GTR CA 3′. PCR was performed in a total volume of 25 µL, which contained 0.6 µmol/L of each primer, 1× of HotStarTaq® Master Mix (QIAGEN, Toronto, ON, Canada) and 20–40 ng of DNA. Amplification was carried out in a GeneAmp® PCR System 9700 (Applied Biosystems, Waltham, MA, USA) under the following conditions: 95 °C (15 min); 35 cycles of 94 °C (20 s), 52 °C (15 s), 72 °C (1 min 30 s); and a 72 °C (7 min) extension step.
Amplified PCR products with a single fragment were confirmed on a 2.0% agarose gel and purified using NucleoFast 96 PCR clean up plate (Macherey-Nagel, Düren, Germany). Purified PCR fragments were sequenced using BigDye® Terminator v3.1 Cycle Sequencing method (Applied Biosystems). DNA sequencing was performed with an automated ABI 3730 DNA analyzer (Applied Biosystems) following the manufacturer’s instruction (ABI Prism® DNA Sequencing Analysis Software Version 3.7, Applied Biosystems). Forward and reverse sequences were edited and aligned. Aligned sequences were compared to the NCBI database using the BLAST tool.
Genomic DNA was extracted from S62-W broth and plate cultures for high-throughput sequencing using QIAGEN® Gentra Puregene Yeast/Bact. Kit. Sequencing libraries were prepared from 1 ng of genomic DNA with the Nextera XT kit (Illumina) according to the manufacturer’s instructions, and subsequently sequenced on a MiSeq instrument (Illumina, San Diego, CA, USA) using a 600-cycle v3 kit (Illumina). The resulting 300 bp paired-end reads were filtered to remove phiX, adapter and human sequence contamination using BBTools (https://jgi.doe.gov/data-and-tools/bbtools/ (accessed on 20 April 2021)) and assembled with SPAdes v. 3.11.1 [46].
Whole-genome based identification of S62-W was initially performed with the JSpeciesWS online service [47]. Further phylogenetic analysis based on core genome alignments of the S62-W sequenced genome and 39 Bacillus spp. reference genomes was performed with RealPhy v. 1.12 [48]. Bacillus spp. were selected for phylogenetic comparison based on putative taxonomic assignments from JSpeciesWS, as well as the KmerFinder tool (https://cge.cbs.dtu.dk/services/KmerFinder/) (accessed on 20 April 2021). Five representative genomes were chosen for each species, wherever available, in the event that the species in some cases had been assigned incorrectly. A maximum likelihood tree was built with RAxML v 8.1.17 [49] using rapid Bootstrap analysis based on 400 replicates, and visualized with the Interactive Tree of Life (iTOL) web service [50].
5.5. ZEA Transformation Activities of the Active Isolate
ZEA transformation activities of the isolate were evaluated under various media (CMB, ASE, and CSE), temperature (22, 28, 37, and 42 °C), pH (3.0, 4.0, 5.0, 6.0, 7.0, 8.0, 9.0, and 10.0), oxygen requirement (aerobic and anaerobic), and initial ZEA concentration (25, 50, 100, and 200 µg/mL). The pure culture stored in −80 °C freezer was pre-incubated in the NBM media at 28 °C overnight before carrying each transformation evaluation. The initial cell concentration in all experiments mentioned above was adjusted to 2.5 × 107 cfu/mL. The percentage of ZEA transformation was calculated with the Equation (1).
5.6. Analysis of ZEA by HPLC
Samples and standards were analyzed using an HPLC system (Agilent Technology 1200 Series, Palo Alto, CA, USA) equipped with a quaternary pump, an inline degasseµr, and a diode array detector set at 235 nm. A Phenomenex® 4 µ Jupiter Proteo 90A (250 × 4.6 mm) with a C18 guard column (Torrance, CA, USA) was used for the separation. The compound of interest, ZEA in this case, was eluted using binary mobile phase set at a flow rate of 1.0 mL/min. The mobile phase was acetonitrile:water (60:40 by volume) and the injection volume was 10 μL. The retention time of ZEA was around 8.2 min.
5.7. Extraction and Purification of ZEA Transformation Product
The isolate with ZEA transformation activity stored in −80 °C was centrifuged at 3500× g for 10 min. The pellet was collected and dissolved in 1 mL of CMB. A 100 µL volume of culture was transferred into a 10 mL of CMB and pre-incubated at 28 °C with shaking at 120 RPM for 24 h. Then pre-incubated culture (10 mL) was transferred in 400 mL of CMB media spiked with 10 mg of ZEA to reach a final concentration of 25 µg/mL. The culture was aerobically incubated at 28 °C with shaking at 120 RPM for 48 h. The decrease of ZEA and increase of ZEA transformation product (i.e., compound X) were detected by HPLC.
After incubation, the culture was centrifuged at 3500× g for 10 min to remove the cells and particles. The supernatant was freeze-dried, and the compound X was extracted by 10 mL of 50% v/v acetonitrile. The acetonitrile extraction solution was centrifuged at 3500× g for 10 min to remove the precipitation. The supernatant was passed through 0.45 µm syringe filter before HPLC collection.
The filtered extraction solution was fractionally collected using an HPLC system (Agilent Technology 1200 Series, Palo Alto, CA, USA) equipped with a quaternary pump, an inline degasser, and a diode array detector set at 270 nm. A Luna 5 µ C18 (2) 100A (250 × 10.0 mm) semi preparative column with a C18 guard column (Phenomenex, Torrance, CA, USA) was used for the separation. The compound of interest was eluted using binary mobile phase at a flow rate of 1.0 mL/min with run time of 23 min. The mobile phase was acetonitrile:water (32:68 by volume) with 0.5% formic acid. A volume of 100 µL were injected. The retention time of the compound X ranged between 17 and 21 min. This elution fraction was collected using an Agilent 1200 series fraction collector. The collected fraction was concentrated by a rotary evaporator (Heidolph, Elk Grove Village, IL, USA) to remove solvent. Then it was freeze-died (Labconco, Kansas City, MO, USA) and the pure compound X was obtained.
5.8. Identification of ZEA Transformation Product by LC-MS/MS and NMR
LC/MS analysis of ZEA and compound X were carried out by using a Thermo Scientific Q-Exactive Benchtop Orbitrap Mass Spectrometer connected with a Vanquish™ Flex Binary UPLC System (Thermo Scientific, Waltham, MA, USA). An Agilent ZORBAX SB-C18 column (2.1 × 100 mm, 3.5 µm) was used for separation. Two mobile phases, solvent A (99.9% H2O + 0.1% formic acid) and solvent B (99.9% acetonitrile + 0.1% formic acid) were used. The chromatographic elution condition was as following: 0–10 min, isocratic 40% B. The column compartment was controlled at 25 °C, the flow rate was set at 0.4 mL/min, the injection amount was 10 µL and the UV absorption wavelength is 270 nm. The negative heated-electrospray ionization (HESI) mode was used for the data collection. The optimized HESI conditions were as follows: sheath gas, 58 arbitrary units; auxiliary gas, 16 arbitrary units; sweep gas, 3 arbitrary units; spray voltage, 3.5 kV; S-lens RF level, 50%; capillary temperature, 281 °C and auxiliary heater temperature, 463 °C. The normalized collision energy (NCE) used for parallel reaction monitoring (PRM) mode was set at 20%. The automatic gain control target and maximum injection time were 1e6 and 50 ms.
1D 1H and 2D spectra, such as Correlated Spectroscopy (COSY), Heteronuclear Single-Quantum Correlation (HSQC), Heteronuclear Multiple Bond Correlation (HMBC), and Rotating-frame Overhauser and Exchange Spectroscopy (ROESY), were acquired at 25 °C on an Agilent DD2 spectrometer (υ(1H) = 699.803 MHz, υ(13C) = 175.982; Agilent, Walnut Creek, CA, USA) equipped with a 5 mm HFCN cold probe. The 1D 13C spectrum was acquired at 25 °C on an Agilent DD2 spectrometer (υ (1H) = 499.662 MHz, υ (13C) = 125.653; Agilent, Walnut Creek, CA, USA) equipped with a 5 mm XSens cold probe. The 1D 31P and 2D Heteronuclear NOESY (HP-HOESY) spectra were acquired at 25 °C on an Agilent DD2 spectrometer (υ (1H) = 599.823 MHz, υ (31P) = 242.812 MHz; Agilent, Walnut Creek, CA, USA) equipped with a 5 mm HFX probe. Purified compound X (1.5 mg) was dissolved in a 1:1 mixture of acetonitrile-d3: D2O (550 µL total volume). The mixture was vortexed for 1 min and then transferred to a 5 mm tube for analysis by NMR. NMR spectra were processed using MestReNova (version 10.0.2-15465, Mestrelab Research S.L., Santiago de Compostela, Spain). 1H and 13C chemical shifts were referenced relative to acetonitrile- d3 (δ(1H) = 1.94 ppm, δ(13C) = 1.320 ppm).
5.9. Identification of ZEA Transformation Product by Comparing Standard ZEA-14-Phosphate and ZEA-16-Phosphate
Compound X was purified from the metabolites of Bacillus sp. S62-W using the above-mentioned method. Since neither compound of ZEA phosphate conjugation was reported, nor were such products available, ZEA-14-phosphate and ZEA-16-phosphate were artificially synthesized based on our hypothesized ZEA transformation products. In order to identify the position of phosphate conjugation, 50 µg/mL for each compound were analyzed using an HPLC system described above. The compound of interest was eluted using binary mobile phase at a flow rate of 1.0 mL/min. The mobile phase was acetonitrile:water with 0.5% formic acid (60:40 by volume) and the injection volume was 10 μL.
5.10. Determination of ZEA Phosphorylation in Various Bacillus Strains
Multiple Bacillus spp. were selected to test ZEA phosphorylation activities. Twenty-one strains (B. amyloliquefaciens, B. licheniformis, B. megaterium, B. pumilus, and B. subtilis; see Figure 8 for complete list of the strains) were obtained from the laboratory culture collection of the Guelph Research and Development Centre, Agriculture and Agri-Food Canada. Two strains, B. pumilus AQP-4275 and B. subtilis AQP-6638, were isolated by Lallemand Inc. (Montréal, QC, Canada), which are exempted of Nagoya protocol. The ZEA detoxification activity was identified using the techniques described above. Briefly, bacteria from −80 °C freezer were initially enriched in NA plates and were incubated for 3 days at 28 °C. The colonies were transferred into 1 mL of CMB and were incubated overnight at 28 °C with shaking at 120 RPM. In order to ensure the similar initial concentration of inoculum, the enriched cultures were normalized to the OD600 = 0.1. Then 50 µL of normalized culture was inoculated to 945 µL of fresh CMB and 5 µL of ZEA (5000 µg/mL). The final concentration of ZEA was 25 µg/mL. The control sample was prepared in the same method without inoculation of bacteria. Samples and control were incubated at 28 °C with shaking at 120 RPM. One hundred microliter of each sample was taken out after 24 h. The transformation was terminated by adding 600 µL of acetonitrile and 300 µL of water. All samples were filtered by 0.45 µm syringe filter before HPLC analysis.
5.11. Statistical Analysis
ZEA transformation activities of Bacillus sp. S62-W were performed in triplicate with completely independent and randomized design. The statistical analyses of experimental data were carried out by SPSS version 20 (IBM, Armonk, NY, USA). Statistical significance of ZEA concentration between control and treatment were evaluated through paired T-test. The percentages of ZEA transformation were evaluated through one-way ANOVA with post-hoc Tukey’s HSD test.
Author Contributions
Conceptualization, Y.Z., T.Z. and M.C.; methodology, Y.Z., P.D., D.L., X.-Z.L. and H.Z.; software, D.L.; formal analysis, Y.Z., D.L. and H.Z.; resources, T.Z.; writing, Y.Z., P.D., D.L., X.-Z.L., H.Z., T.Z.; supervision, T.Z.; project administration, T.Z.; funding acquisition, T.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by collaborative framework projects, jointly supported by Agriculture and Agri-Food Canada and Lallemand Inc. (Project J-000653).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zinedine, A.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef]
- Tanaka, T.; Hasegawa, A.; Yamamoto, S.; Lee, U.S.; Sugiura, Y.; Ueno, Y. Worldwide contamination of cereals by the fusarium mycotoxins nivalenol, deoxynivalenol, and zearalenone. 1. Survey of 19 countries. J. Agric. Food Chem. 1988, 36, 979–983. [Google Scholar] [CrossRef]
- Tangni, E.K.; Pussemier, L.; Van Hove, F. Mycotoxin contaminating maize and grass silages for dairy cattle feeding: Current state and challenges. J. Anim. Sci. Adv. 2013, 3, 492–511. [Google Scholar]
- Ogunade, I.M.; Martinez-Tuppia, C.; Queiroz, O.C.M.; Jiang, Y.; Drouin, P.; Wu, F.; Vyas, D.; Adesogan, A.T. Silage review: Mycotoxins in silage: Occurrence, effects, prevention, and mitigation. J. Dairy Sci. 2018, 101, 4034–4059. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Scientific opinion on the risks for public health related to the presence of zearalenone in food. EFSA J. 2011, 9, 2197. [Google Scholar] [CrossRef]
- Pereyra, M.L.G.; Sulyok, M.; Baralla1, V.; Dalcero1, A.M.; Krska, R.; Chulze, S.; Cavaglieri, L.R. Evaluation of zearalenone, α-zearalenol, β-zearalenol, zearalenone 4-sulfate and β-zearalenol 4-glucoside levels during the ensiling process. World Mycotoxin J. 2014, 7, 291–295. [Google Scholar] [CrossRef]
- He, J.; Zhou, T. Patented techniques for detoxification of mycotoxins in feeds and food matrices. Recent Pat. Food Nutr. Agric. 2010, 2, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Hassan, Y.I.; Watts, C.; Zhou, T. Innovative technologies for the mitigation of mycotoxins in animal feed and ingredients—A review of recent patents. Anim. Feed Sci. Technol. 2016, 216, 19–29. [Google Scholar] [CrossRef]
- Yi, P.; Pai, C.; Liu, J. Isolation and characterization of a bacillus licheniformis strain capable of degrading zearalenone. World J. Microbiol. Biotechnol. 2011, 27, 1035–1043. [Google Scholar] [CrossRef]
- Cho, K.J.; Kang, J.S.; Cho, W.T.; Lee, C.H.; Ha, J.K.; Song, K.B. In vitro degradation of zearalenone by bacillus subtilis. Biotechnol. Lett. 2010, 32, 1921–1924. [Google Scholar] [CrossRef]
- Lei, Y.; Zhao, L.; Ma, Q.; Zhang, J.; Zhou, T.; Gao, C.; Ji, C. Degradation of zearalenone in swine feed and feed ingredients by bacillus subtilis ansb01g. World Mycotoxin J. 2014, 7, 143–151. [Google Scholar] [CrossRef]
- Tinyiro, S.E.; Yao, W.; Sun, X.; Wokadala, C.; Wang, S. Scavenging of zearalenone by bacillus strains-in vitro. Res. J. Microbiol. 2011, 6, 304–309. [Google Scholar] [CrossRef] [Green Version]
- Tinyiro, S.E.; Wokadala, C.; Xu, D.; Yao, W. Adsorption and degradation of zearalenone by bacillus strains. Folia Microbiol. 2011, 56, 321–327. [Google Scholar] [CrossRef]
- Tan, H.; Hu, Y.; He, J.; Wu, L.; Liao, F.; Luo, B.; He, Y.; Zuo, Z.; Ren, Z.; Zhong, Z.; et al. Zearalenone degradation by two pseudomonas strains from soil. Mycotoxin Res. 2014, 30, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Zhang, Z.; Hu, Y.; Wu, L.; Liao, F.; He, J.; Luo, B.; He, Y.; Zuo, Z.; Ren, Z.; et al. Isolation and characterization of pseudomonas otitidis th-n1 capable of degrading zearalenone. Food Control 2015, 47, 285–290. [Google Scholar] [CrossRef]
- Yu, Y.; Qiu, L.; Wu, H.; Tang, Y.; Yu, Y.; Li, X.; Liu, D. Degradation of zearalenone by the extracellular extracts of acinetobacter sp. Sm04 liquid cultures. Biodegradation 2011, 22, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Kriszt, R.; Krifaton, C.; Szoboszlay, S.; Cserháti, M.; Kriszt, B.; Kukolya, J.; Czéh, A.; Fehér-Tóth, S.; Török, L.; Szőke, Z.; et al. A new zearalenone biodegradation strategy using non-pathogenic rhodococcus pyridinivorans k408 strain. PLoS ONE 2012, 7, e43608. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Dong, M.; Yang, Q.; Apaliya, M.T.; Li, J.; Zhang, X. Biodegradation of zearalenone by saccharomyces cerevisiae: Possible involvement of zen responsive proteins of the yeast. J. Proteom. 2016, 143, 416–423. [Google Scholar] [CrossRef]
- Brodehl, A.; Möller, A.; Kunte, H.J.; Koch, M.; Maul, R. Biotransformation of the mycotoxin zearalenone by fungi of the genera rhizopus and aspergillus. FEMS Microbiol. Lett. 2014, 359, 124–130. [Google Scholar] [CrossRef] [Green Version]
- Jard, G.; Liboz, T.; Mathieu, F.; Guyonvarc’h, A.; André, F.; Delaforge, M.; Lebrihi, A. Transformation of zearalenone to zearalenone-sulfate by aspergillus spp. World Mycotoxin J. 2010, 3, 183–191. [Google Scholar] [CrossRef]
- Sun, X.; He, X.; Xue, K.; Li, Y.; Xu, D.; Qian, H. Biological detoxification of zearalenone by aspergillus niger strain fs10. Food Chem. Toxicol. 2014, 72, 76–82. [Google Scholar] [CrossRef]
- El-Sharkawy, S.H.; Selim, M.I.; Afifi, M.S.; Halaweish, F.T. Microbial transformation of zearalenone to a zearalenone sulfate. Appl. Environ. Microbiol. 1991, 57, 549–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vekiru, E.; Hametner, C.; Mitterbauer, R.; Rechthaler, J.; Adam, G.; Schatzmayr, G.; Krska, R.; Schuhmacher, R. Cleavage of zearalenone by trichosporon mycotoxinivorans to a novel nonestrogenic metabolite. Appl. Environ. Microbiol. 2010, 76, 2353–2359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakeya, H.; Takahashi-Ando, N.; Kimura, M.; Onose, R.; Yamaguchi, I.; Osada, H. Biotransformation of the mycotoxin, zearalenone, to a non-estrogenic compound by a fungal strain of clonostachys sp. Biosci. Biotechnol. Biochem. 2002, 66, 2723–2726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Sharkawy, S.; Abul-Hajj, Y. Microbial transformation of zearalenone, i. Formation of zearalenone-4-o-β-glucoside. J. Nat. Prod. 1987, 50, 520–521. [Google Scholar] [CrossRef]
- Branquinho, R.; Meirinhos-Soares, L.; Carriço, J.A.; Pintado, M.; Peixe, L.V. Phylogenetic and clonality analysis of bacillus pumilus isolates uncovered a highly heterogeneous population of different closely related species and clones. FEMS Microbiol. Ecol. 2014, 90, 689–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Hassan, Y.I.; Lepp, D.; Shao, S.; Zhou, T. Strategies and methodologies for developing microbial detoxification systems to mitigate mycotoxins. Toxins 2017, 9, 130. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, S.C. Xenobiotic conjugation with phosphate—A metabolic rarity. Xenobiotica 2016, 46, 743–756. [Google Scholar] [CrossRef]
- Day, E.K.; Sosale, N.G.; Lazzara, M.J. Cell Signaling regulation by protein phosphorylation: A multivariate, heterogeneous, and context-dependent process. Curr. Opin. Biotechnol. 2016, 40, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.; Chang, C.; Wang, S.; Fang, T.; Lee, M.; Su, N. Soy isoflavone-phosphate conjugates derived by cultivating bacillus subtilis var. Natto bcrc 80517 with isoflavone. Food Res. Int. 2013, 53, 487–495. [Google Scholar] [CrossRef]
- Abou-Donia, M.B.; Elmasry, E.M.; Abu-Qare, A.W. Metabolism and toxicokinetics of xenobiotics. In Handbook of toxicology, 2nd ed.; Derelanko, M.J., Hollinger, M.A., Eds.; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- EFSA. Update of the list of qps-recommended biological agents intentionally added to food or feed as notified to efsa 4: Suitability of taxonomic units notified to efsa until March 2016. EFSA J. 2016, 14, 4522. [Google Scholar]
- Wang, W.; Sun, M. Phylogenetic relationships between Bacillus species and related genera inferred from 16s rDNA sequences. Braz. J. Microbiol. 2009, 40, 505–521. [Google Scholar] [CrossRef] [Green Version]
- Te Giffel, M.C.; Wagendorp, A.; Herrewegh, A.; Driehuis, F. Bacterial spores in silage and raw milk. Antonie Leeuwenhoek. 2002, 81, 625–630. [Google Scholar] [CrossRef]
- Ávila, C.L.S.; Carvalho, B.F. Silage fermentation—Updates focusing on the performance of micro-organisms. J. Appl. Microbiol. 2020, 128, 966–984. [Google Scholar] [CrossRef] [Green Version]
- Drouin, P.; Tremblay, J.; Renaud, J.; Apper, E. Microbiota succession during aerobic stability of maize silage inoculated with Lentilactobacillus buchneri NCIMB 40788 and Lentilactobacillus hilgardii CNCM-I-4785. MicrobiologyOpen 2021, 10, e1153. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, O.C.M.; Ogunade, I.M.; Weinberg, Z.; Adesogan, A.T. Silage review: Foodborne pathogens in silage and their mitigation by silage additives. J. Dairy Sci. 2017, 101, 4132–4142. [Google Scholar] [CrossRef] [PubMed]
- Lara Erika, C.; Basso Fernanda, C.; de Assis Flávia, B.; Souza Fernando, A.; Berchielli Telma, T.; Reis Ricardo, A. Changes in the nutritive value and aerobic stability of corn silages inoculated with Bacillus subtilis alone or combined with Lactobacillus Plantarum. Anim. Prod. Sci. 2015, 56, 1867–1874. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.; Cheng, K.C.; Liu, J.R. Isolation and characterization of a Bacillus amyloliquefaciens strain with zearalenone removal ability and its probiotic potential. PLoS ONE 2017, 12, e0182220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danilova, I.; Sharipova, M. The practical potential of Bacilli and their enzymes for industrial production. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Elshaghabee, F.M.F.; Rokana, N.; Gulhane, R.D.; Sharma, C.; Panwar, H. Bacillus as potential probiotics: Status, concerns, and future perspectives. Front. Microbiol. 2017, 8, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Sharkawy, S.H.; Abul-Hajj, Y.J. Microbial transformation of zearalenone. 2. Reduction, hydroxylation, and methylation products. J. Org. Chem. 1988, 53, 515–519. [Google Scholar] [CrossRef]
- Rychlik, M.; Humpf, H.; Marko, D.; Dänicke, S.; Mally, A.; Berthiller, F.; Klaffke, H.; Lorenz, N. Proposal of a comprehensive definition of modified and other forms of mycotoxins including “masked” mycotoxins. Mycotoxin Res. 2014, 30, 197–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthiller, F.; Crews, C.; Dall’Asta, C.; Saeger, S.D.; Haesaert, G.; Karlovsky, P.; Oswald, I.P.; Seefelder, W.; Speijers, G.; Stroka, J. Masked mycotoxins: A review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [CrossRef]
- Gratz, S.W.; Dinesh, R.; Yoshinari, T.; Holtrop, G.; Richardson, A.J.; Duncan, G.; MacDonald, S.; Lloyd, A.; Tarbin, J. Masked trichothecene and zearalenone mycotoxins withstand digestion and absorption in the upper gi tract but are efficiently hydrolyzed by human gut microbiota in vitro. Mol. Nutr. Food Res. 2016. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. Spades: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Richter, M.; Rossello-Mora, R.; Oliver Glockner, F.; Peplies, J. Jspeciesws: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef]
- Bertels, F.; Silander, O.K.; Pachkov, M.; Rainey, P.B.; van Nimwegen, E. Automated reconstruction of whole-genome phylogenies from short-sequence reads. Mol. Biol. Evol. 2014, 31, 1077–1088. [Google Scholar] [CrossRef]
- Stamatakis, A. Raxml version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive tree of life (itol) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phylogenetic analysis based on core genome sequence alignments of S62-W and 39 Bacillus reference genomes.
Figure 1.
Phylogenetic analysis based on core genome sequence alignments of S62-W and 39 Bacillus reference genomes.

Figure 2.
HPLC chromatogram of (A) S62-W culture spiked with 50 µg/mL of ZEA (0 h), (B) S62-W culture spiked with 50 µg/mL of ZEA (24 h), and (C) S62-W culture without ZEA (24 h).
Figure 2.
HPLC chromatogram of (A) S62-W culture spiked with 50 µg/mL of ZEA (0 h), (B) S62-W culture spiked with 50 µg/mL of ZEA (24 h), and (C) S62-W culture without ZEA (24 h).
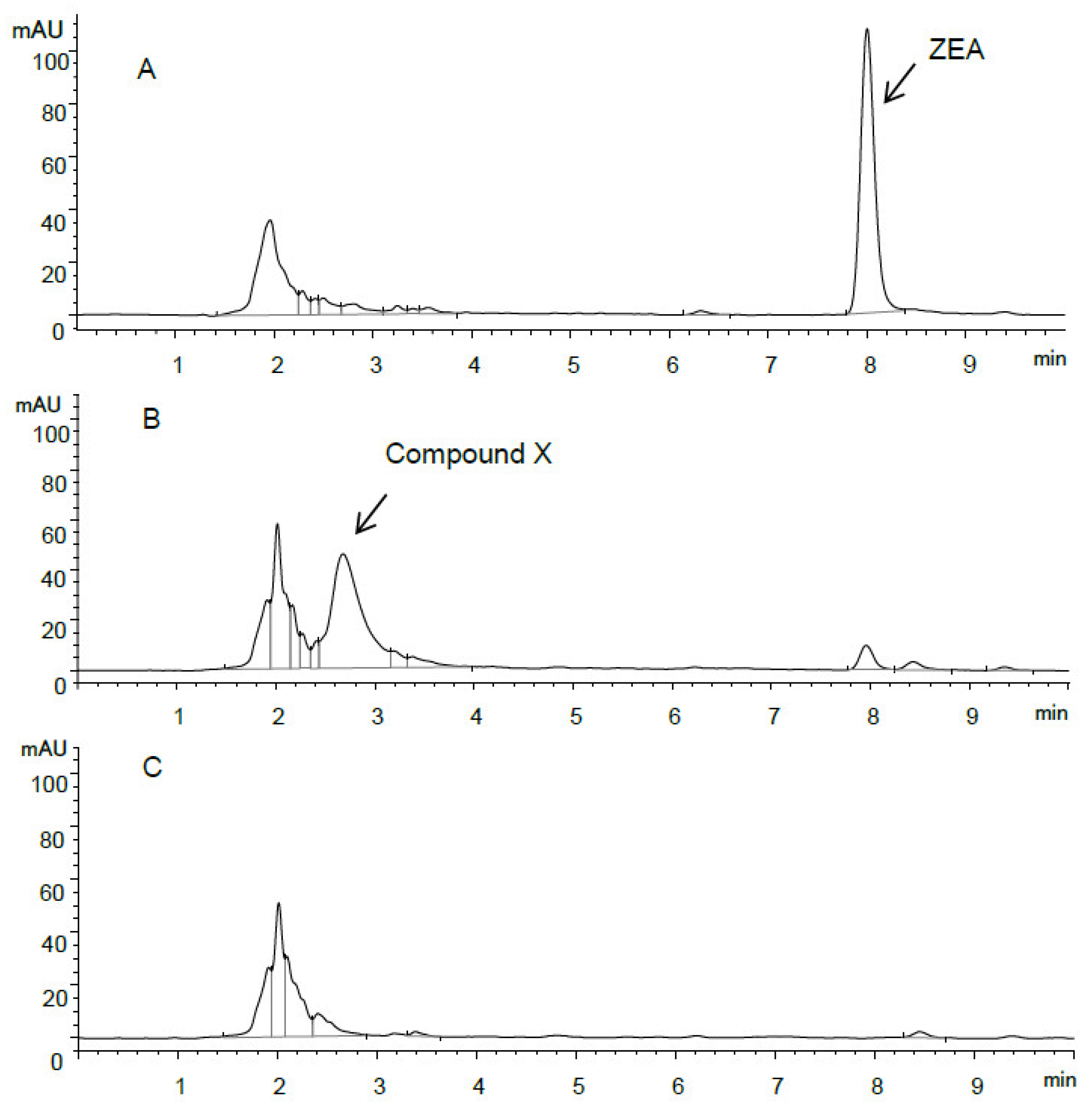
Figure 3.
HPLC UV spectra of ZEA and compound X.

Figure 4.
MS spectra of (A) standard ZEA (25 µg/mL) and (B) compound X (25 µg/mL); MS/MS spectrum of (C) compound X.
Figure 4.
MS spectra of (A) standard ZEA (25 µg/mL) and (B) compound X (25 µg/mL); MS/MS spectrum of (C) compound X.
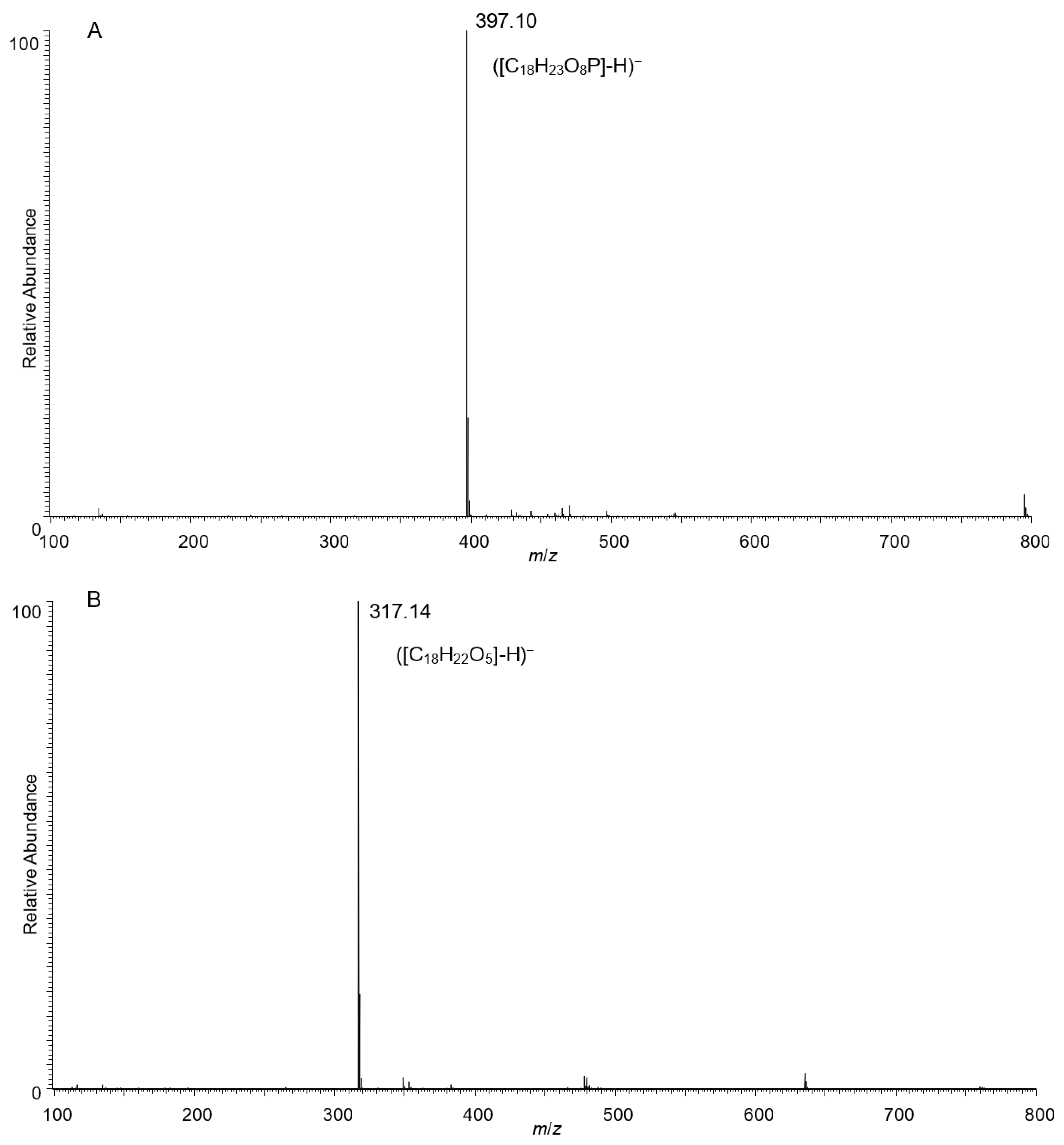

Figure 5.
Structure of ZEA-14-phosphate.

Figure 6.
Correlation (A) between ZEA decrease and growth of Bacillus sp. S62-W in CMB at 28 °C (n = 3), and transformation (B) of ZEA and production of ZEA-14-phosphate by Bacillus sp. S62-W in CMB at 28 °C (n = 3).
Figure 6.
Correlation (A) between ZEA decrease and growth of Bacillus sp. S62-W in CMB at 28 °C (n = 3), and transformation (B) of ZEA and production of ZEA-14-phosphate by Bacillus sp. S62-W in CMB at 28 °C (n = 3).

Figure 7.
(A): ZEA transformation by Bacillus sp. S62-W in various media (28 °C, 3 days, n = 3). CMB: Corn meal broth W/O yeast extract; ASE: Alfalfa silage extract; CSE: Corn silage extract. (B): ZEA transformation by Bacillus sp. S62-W in CMB with pH value between 3 and 10 (28 °C, 5 days, n = 3). (C): ZEA transformation by Bacillus sp. S62-W in CMB incubated between 22 and 42 °C (2 days, n = 3). (D): ZEA transformation by Bacillus sp. S62-W in CMB under different incubation conditions (n = 3). Aerobic: Aerobic incubation at 28 °C for 2 days; Anaerobic: Anaerobic incubation at 28 °C for 2 days; AN-A: Anaerobic incubation at 28 °C for 6 days following by aerobic incubation at 28 °C for 2 days. (E): ZEA transformation by Bacillus sp. S62-W in CMB spiked with concentration varying between 25 and 200 µg/mL of ZEA (28 °C, 24 h and 96 h, n = 3).
Figure 7.
(A): ZEA transformation by Bacillus sp. S62-W in various media (28 °C, 3 days, n = 3). CMB: Corn meal broth W/O yeast extract; ASE: Alfalfa silage extract; CSE: Corn silage extract. (B): ZEA transformation by Bacillus sp. S62-W in CMB with pH value between 3 and 10 (28 °C, 5 days, n = 3). (C): ZEA transformation by Bacillus sp. S62-W in CMB incubated between 22 and 42 °C (2 days, n = 3). (D): ZEA transformation by Bacillus sp. S62-W in CMB under different incubation conditions (n = 3). Aerobic: Aerobic incubation at 28 °C for 2 days; Anaerobic: Anaerobic incubation at 28 °C for 2 days; AN-A: Anaerobic incubation at 28 °C for 6 days following by aerobic incubation at 28 °C for 2 days. (E): ZEA transformation by Bacillus sp. S62-W in CMB spiked with concentration varying between 25 and 200 µg/mL of ZEA (28 °C, 24 h and 96 h, n = 3).
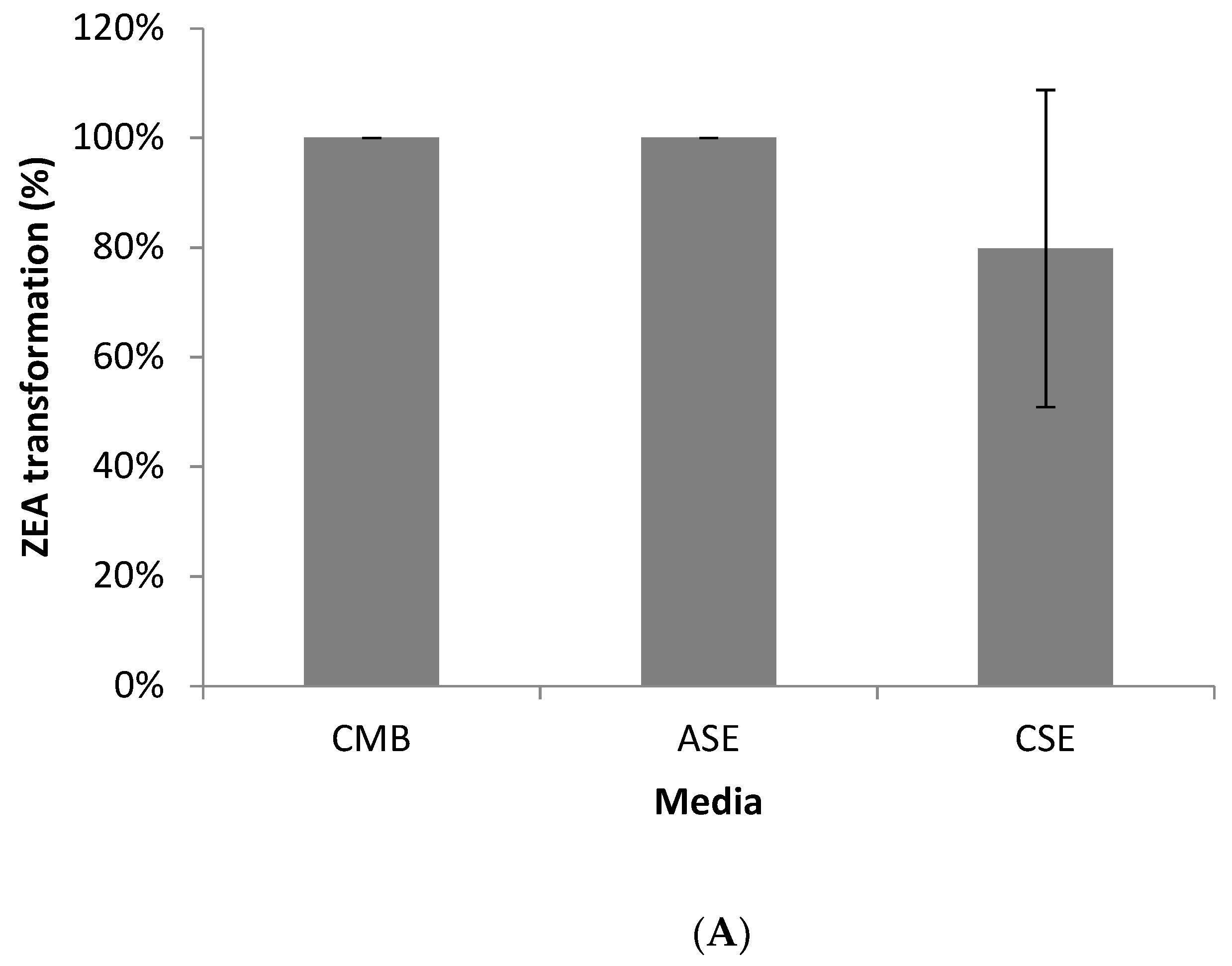
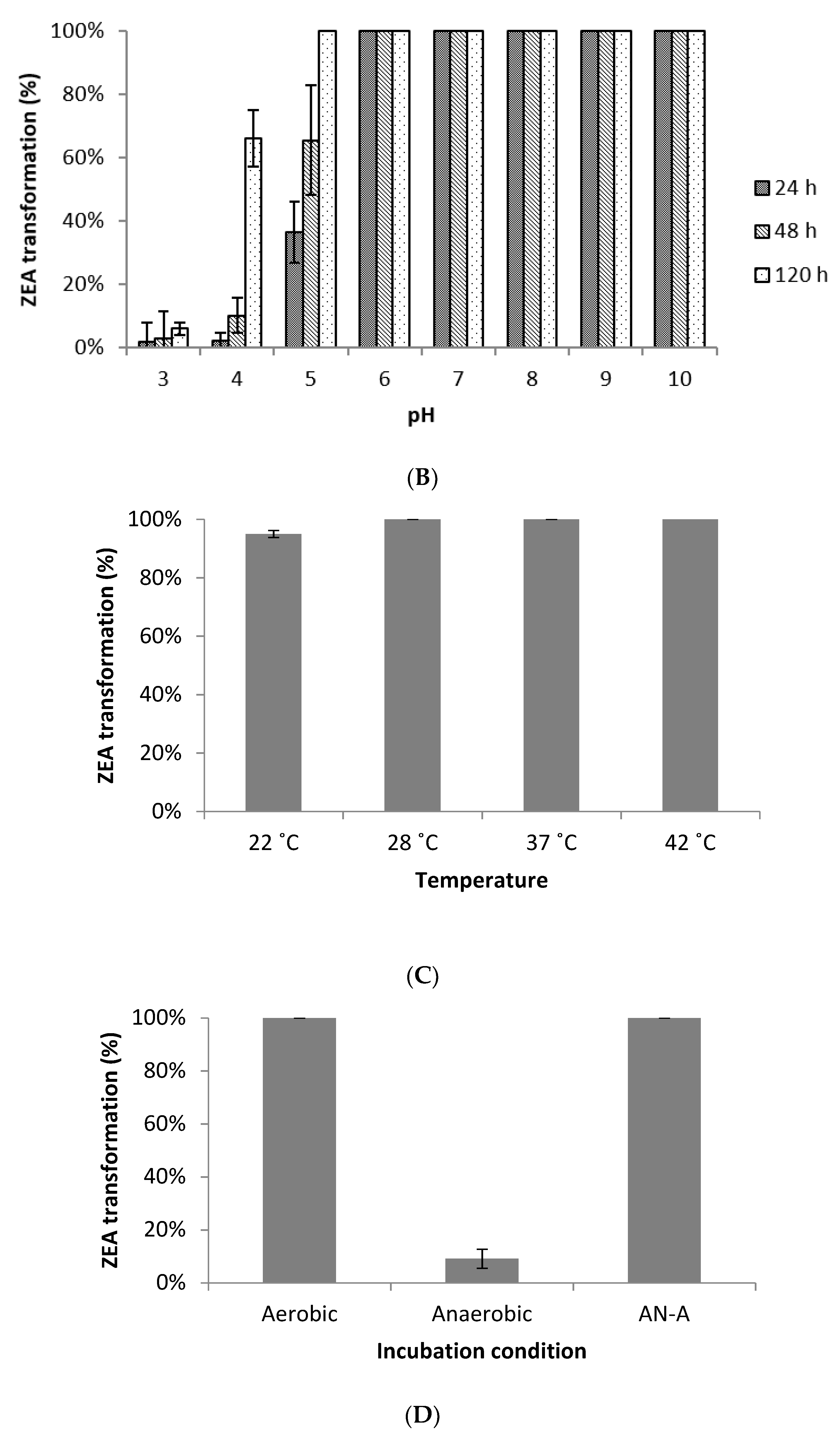

Figure 8.
ZEA phosphorylation activity of 23 Bacillus spp. and Bacillus sp. S62-W in CMB at 28 °C for 24 h incubation.
Figure 8.
ZEA phosphorylation activity of 23 Bacillus spp. and Bacillus sp. S62-W in CMB at 28 °C for 24 h incubation.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Putative identities of S62-W based on 16S rRNA gene sequence.
| Microorganism | Accession No. | Identity (%) |
|---|---|---|
| Bacillus aerius 24K | NR118439.1 | 99 |
| Bacillus stratosphericus 41KF2a | NR042336.1 | 99 |
| Bacillus altitudinis 41KF2b | NR042337.1 | 99 |
| Bacillus pumilus NBRC 12092 | NR112637.1 | 99 |
| Bacillus pumilus SAFR-032 | NR074977.1 | 99 |
| Bacillus pumilus ATCC 7061 | NR043242.1 | 99 |
| Bacillus pumilus SBMP2 | NR118381.1 | 99 |
| Bacillus safensis NBRC 100820 | NR113945.1 | 99 |
| Bacillus safensis FO-366 | NR041794.1 | 99 |
Table 2.
Putative identities of S62-W based on whole genome analysis with JSpeciesWS Tetra correlation search (TCS) tool.
Table 2.
Putative identities of S62-W based on whole genome analysis with JSpeciesWS Tetra correlation search (TCS) tool.
| Microorganism | Z-Scores | ANIb (%) |
|---|---|---|
| Bacillus stratosphericus LK31 | 0.99986 | 98.05 |
| Bacillus stratosphericus (GCA_001043535) LK5 | 0.99984 | 98.07 |
| Bacillus stratosphericus LK23 | 0.99982 | 98.06 |
| Bacillus sp. LK10 | 0.99982 | 98.06 |
| Bacillus stratosphericus (GCA_001038775) LK33 | 0.99981 | 98.06 |
| Bacillus sp. L_1B0_12 | 0.99979 | 98.02 |
| Bacillus stratosphericus (GCA_001038845) LK18 | 0.99979 | 98.05 |
| Bacillus sp. TH007 | 0.99979 | 98.79 |
| Bacillus pumilus (GCA_000828455) B4133 | 0.99975 | 98.30 |
| Bacillus stratosphericus (GCA_001265125) M53 | 0.99975 | 98.09 |
| Bacillus cellulasensis NIO-1130 | 0.99974 | 97.91 |
| Bacillus altitudinis 41KF2b | 0.99973 | 98.01 |
| Bacillus sp. FJAT-21955 | 0.99971 | 98.51 |
| Bacillus pumilus (GCA_000972685) W3 | 0.99965 | 98.10 |
ANIb: Average Nucleotide Identities.
Table 3.
1H, 13C chemical shift assignments and 2D correlations for ZEA-phosphate.
| Atom | Chemical Shift (ppm) | COSY | HSQC | HMBC | ROESY |
|---|---|---|---|---|---|
| 1C | 171.27 | 3, 13, 15 | |||
| 3C | 74.47 | 3 | 5b, 4b, 5a, 19 | ||
| 4C | 35.22 | 4a, 4b | 19, 6a, 6b, 5b, 5a, 3 | ||
| 5C | 22.31 | 5a, 5b | 3, 19, 6a, 4b, 4a, 6b | ||
| 6C | 43.85 | 6a, 6b | 4b, 5b, 5a | ||
| 7C | 216.18 | 6a, 6b, 8b, 8a, 9b, 5b, 9a, 5a | |||
| 8C | 37.68 | 8b, 8a | 6a, 9b, 9a, 6b, 10a, 10b | ||
| 9C | 21.60 | 9b, 9a | 11, 8b, 8a, 10a, 10b, 12 | ||
| 10C | 31.65 | 10b, 10a | 11, 8a, 8b, 9b, 9a, 12 | ||
| 11C | 134.56 | 11 | 12, 9b, 9a, 10b, 10a | ||
| 12C | 131.54 | 12 | 11, 13, 10b, 10a | ||
| 13C | 112.04 | 13 | 12, 15 | ||
| 14C | 157.33 | 13, 15 | |||
| 15C | 107.35 | 15 | 13 | ||
| 16C | 161.26 | 15 | |||
| 17C | 110.12 | 15, 13, 12, 19 | |||
| 18C | 142.00 | 12, 11 | |||
| 19C | 20.41 | 19(H3) | 4b, 3 | ||
| 3H | 5.01 | 19, 4b | 3 | 5, 1, 4, 19 | 5b, 4a, 4b, 5a, 6a |
| 4Ha | 1.61 | 4b | 4 | 5 | 3, 6a |
| 4Hb | 1.51 | 3, 4a, 5a | 4 | 5, 6, 19, 3 | 12, 3, 6a, 6b |
| 5Ha | 1.62 | 5b, 4b, 6b, 6a | 5 | 7, 4, 6, 3 | 3, 8a, 6a |
| 5Hb | 1.70 | 6a, 6b, 5a | 5 | 7, 4, 6, 3 | 12, 3, 19, 8a, 8b, 6a |
| 6Ha | 2.45 | 6b, 5b, 5a | 6 | 7, 8, 4, 5 | 3, 4b, 5b, 5a, 4a |
| 6Hb | 2.17 | 11, 6a, 5b, 5a | 6 | 7, 4, 8, 5 | 8b, 4b |
| 8Ha | 2.38 | 8b, 9b, 9a | 8 | 7, 9, 10 | 11, 9b, 5b, 9a, 5a |
| 8Hb | 2.58 | 8a, 9b, 9a | 8 | 7, 9, 10 | 11, 5b, 6b, 9a |
| 9Ha | 1.63 | 9b, 8a, 8b, 10b, 10a | 9 | 7, 10, 8, 11 | 11, 8a, 8b |
| 9Hb | 1.79 | 9a, 8b, 8a, 10b | 9 | 7, 10, 8, 11 | 11, 8a, 10a |
| 10Ha | 2.11 | 11, 12, 10b, 9a | 10 | 9, 8, 12, 11 | 12, 9b |
| 10Hb | 2.15 | 11, 12, 10a, 9b, 9a | 10 | 9, 8, 12, 11 | 12 |
| 11H | 5.89 | 12, 6b, 10a, 10b | 11 | 9, 10, 18, 12 | 13, 9a, 9b, 8a, 8b |
| 12H | 6.71 | 11, 10a, 10b | 12 | 18, 11, 13, 17, 10, 9 | 4b, 10a, 10b, 5b, 19 |
| 13H | 6.75 | 15 | 13 | 12, 14, 1, 15, 17 | 11 |
| 15H | 6.65 | 13 | 15 | 17, 14, 16, 1, 13 | |
| 19H3 | 1.29 | 3 | 19 | 4, 5, 3, 17 | 12, 5b |
COSY: Correlated Spectroscopy; HSQC: Heteronuclear Single-Quantum Correlation; HMBC: Heteronuclear Multiple Bond Correlation; ROESY: Rotating-frame Overhauser and Exchange Spectroscopy.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhu, Y.; Drouin, P.; Lepp, D.; Li, X.-Z.; Zhu, H.; Castex, M.; Zhou, T. A Novel Microbial Zearalenone Transformation through Phosphorylation. Toxins 2021, 13, 294. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13050294
AMA Style
Zhu Y, Drouin P, Lepp D, Li X-Z, Zhu H, Castex M, Zhou T. A Novel Microbial Zearalenone Transformation through Phosphorylation. Toxins. 2021; 13(5):294. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13050294
Chicago/Turabian StyleZhu, Yan, Pascal Drouin, Dion Lepp, Xiu-Zhen Li, Honghui Zhu, Mathieu Castex, and Ting Zhou. 2021. "A Novel Microbial Zearalenone Transformation through Phosphorylation" Toxins 13, no. 5: 294. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13050294
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.