In-Host Emergence of Linezolid Resistance in a Complex Pattern of Toxic Shock Syndrome Toxin-1-Positive Methicillin-Resistant Staphylococcus aureus Colonization in Siblings with Cystic Fibrosis

 , and
, and
Abstract
:1. Introduction
2. Results
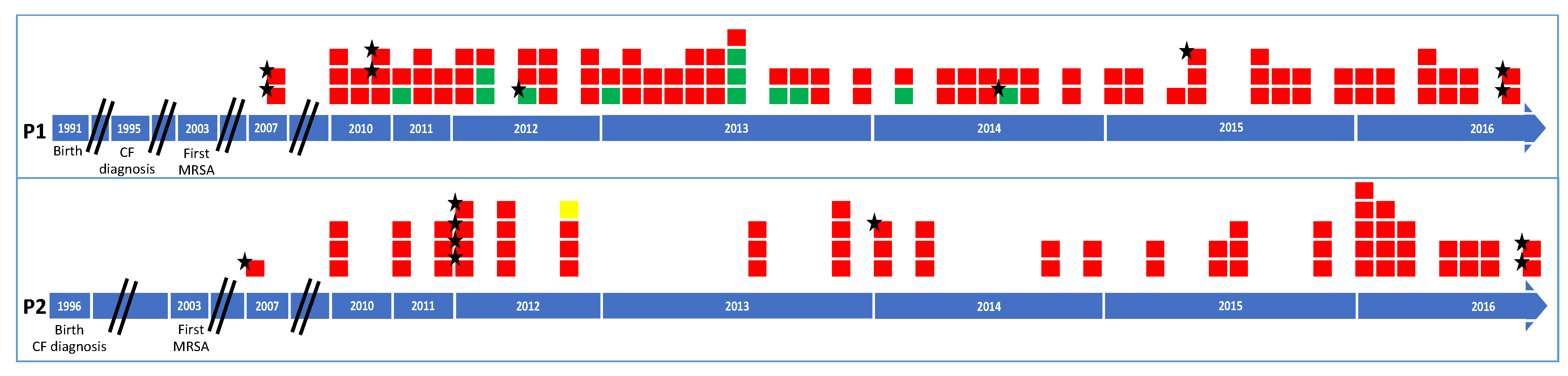
2.1. Patient Follow-Up and Microbiological Data
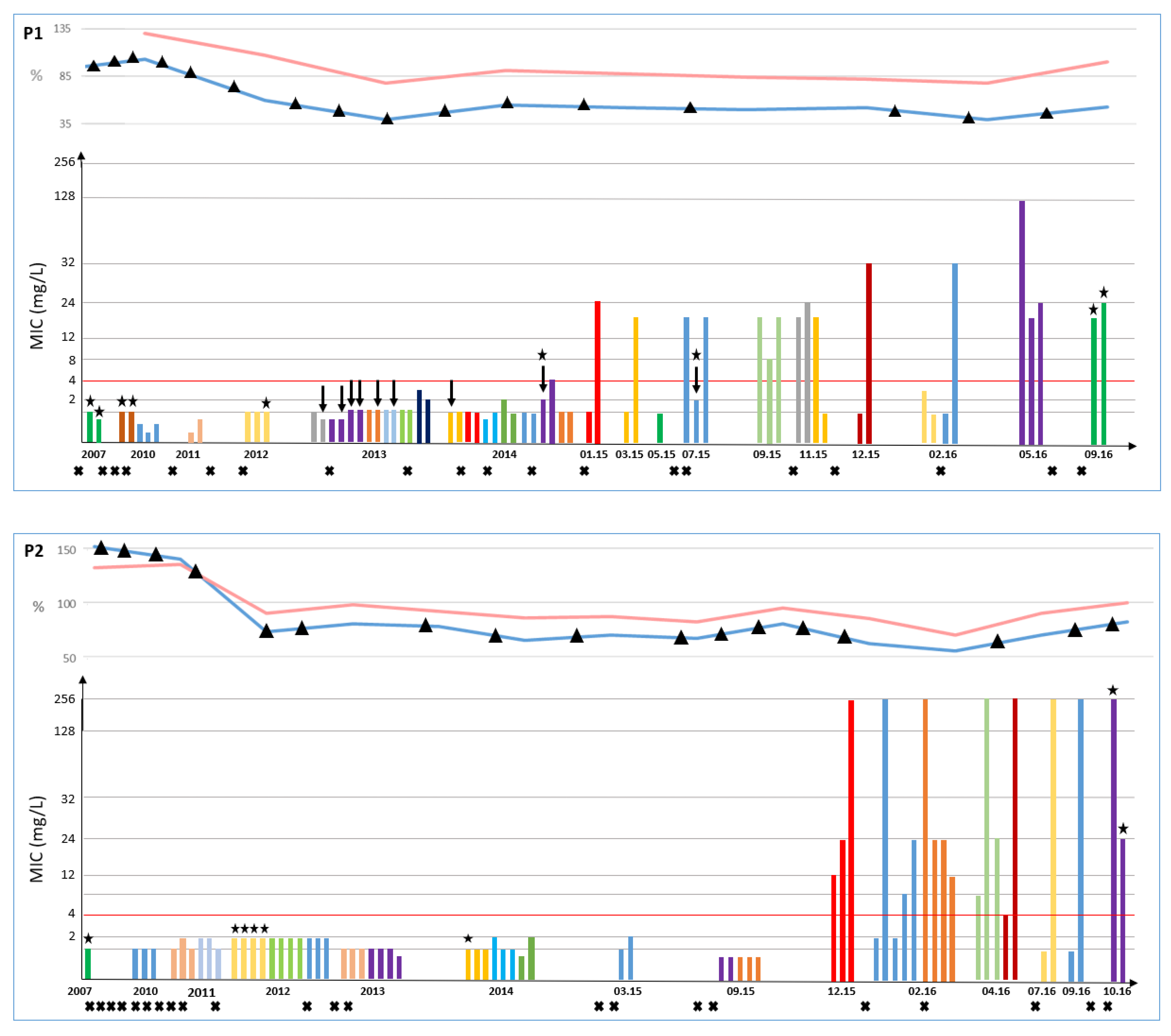
2.2. Antimicrobial Susceptibility Profile and Linezolid Resistance
2.3. Strain Genotyping
2.4. Identifying of the Genetic Determinant of Linezolid Resistance
2.5. Relationship between Isolates
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Patients, Bacterial Strains and Data Collection
5.2. Culture Conditions and Antimicrobial Susceptibility Testing
5.3. DNA Extraction, Analysis of the Staphylococcal Protein A (spa) Locus and Multi Locus Sequence Typing
5.4. Determining the Rrn Copy Number
5.5. Whole Genome Sequencing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Registre français de la mucoviscidose –Bilan des données 2018, Vaincre la Mucoviscidose Paris, Décembre 2020. Available online: https://www.vaincrelamuco.org/sites/default/files/registre_francais_de_la_mucoviscidose_bilan_donnees_2018.pdf (accessed on 20 February 2021).
- Cystic fibrosis foundation patient registry 2019 annual data report. Available online: https://www.cff.org/Research/Researcher-Resources/Patient-Registry/2019-Patient-Registry-Annual-Data-Report.Pdf (accessed on 20 February 2021).
- Ren, C.L.; Morgan, W.J.; Konstan, M.W.; Schechter, M.S.; Wagener, J.S.; Fisher, K.A.; Regelmann, W.E. Presence of methicillin resistant Staphylococcus aureus in respiratory cultures from cystic fibrosis patients is associated with lower lung function. Pediatr. Pulmonol. 2007, 42, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Dasenbrook, E.C.; Checkley, W.; Merlo, C.A.; Konstan, M.W.; Lechtzin, N.; Boyle, M.P. Association Between Respiratory Tract Methicillin-Resistant Staphylococcus aureus and Survival in Cystic Fibrosis. JAMA 2010, 303, 2386–2392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasenbrook, E.C.; Merlo, C.A.; Diener-West, M.; Lechtzin, N.; Boyle, M.P. Persistent Methicillin-resistant Staphylococcus aureus and Rate of FEV1 Decline in Cystic Fibrosis. Am. J. Respir. Crit. Care Med. 2008, 178, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Akil, N.; Muhlebach, M.S. Biology and management of methicillin resistant Staphylococcus aureus in cystic fibrosis. Pediatr. Pulmonol. 2018, 53, S64–S74. [Google Scholar] [CrossRef] [Green Version]
- Ferrin, M.; Zuckerman, J.B.; Meagher, A.; Blumberg, E.A. Successful treatment of methicillin-resistant Staphylococcus aureus pulmonary infection with linezolid in a patient with cystic fibrosis. Pediatr. Pulmonol. 2002, 33, 221–223. [Google Scholar] [CrossRef]
- Serisier, D.J.; Jones, G.; Carroll, M. Eradication of pulmonary methicillin-resistant Staphylococcus aureus (MRSA) in cystic fibrosis with linezolid. J. Cyst. Fibros. 2004, 3, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gales, A.C.; Sader, H.S.; Andrade, S.S.; Lutz, L.; Machado, A.; Barth, A.L. Emergence of linezolid-resistant Staphylococcus aureus during treatment of pulmonary infection in a patient with cystic fibrosis. Int. J. Antimicrob. Agents 2006, 27, 300–302. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Mendes, R.E.; Streit, J.M.; Hogan, P.A.; Flamm, R.K. Five-Year Summary of In Vitro Activity and Resistance Mechanisms of Linezolid against Clinically Important Gram-Positive Cocci in the United States from the LEADER Surveillance Program (2011 to 2015). Antimicrob. Agents Chemother. 2017, 61, e00609-17. [Google Scholar] [CrossRef] [Green Version]
- Endimiani, A.; Blackford, M.; Dasenbrook, E.C.; Reed, M.D.; Bajaksouszian, S.; Hujer, A.M.; Rudin, S.D.; Hujer, K.M.; Perreten, V.; Rice, L.B.; et al. Emergence of Linezolid-Resistant Staphylococcus aureus after Prolonged Treatment of Cystic Fibrosis Patients in Cleveland, Ohio. Antimicrob. Agents Chemother. 2011, 55, 1684–1692. [Google Scholar] [CrossRef] [Green Version]
- Champion, E.; Miller, M.; Popowitch, E.; Hobbs, M.; Saiman, L.; Muhlebach, M.; for the STAR-CF Study Team. Antimicrobial susceptibility and molecular typing of MRSA in cystic fibrosis. Pediatr. Pulmonol. 2013, 49, 230–237. [Google Scholar] [CrossRef]
- Société Française de Microbiologie, Staphylococcus. CASFM/EUCAST. Société Française de Microbiologie. 2020, pp. 59–67. Available online: https://www.sfm-microbiologie.org/2020/10/02/casfm-eucast-v1-2-octobre-2020/ (accessed on 20 February 2021).
- Ankrum, A.; Hall, B.G. Population Dynamics of Staphylococcus aureus in Cystic Fibrosis Patients to Determine Transmission Events by Use of Whole-Genome Sequencing. J. Clin. Microbiol. 2017, 55, 2143–2152. [Google Scholar] [CrossRef] [Green Version]
- Hill, R.L.R.; Kearns, A.M.; Nash, J.; North, S.E.; Pike, R.; Newson, T.; Woodford, N.; Calver, R.; Livermore, D.M. Linezolid-resistant ST36 methicillin-resistant Staphylococcus aureus associated with prolonged linezolid treatment in two paediatric cystic fibrosis patients. J. Antimicrob. Chemother. 2010, 65, 442–445. [Google Scholar] [CrossRef]
- Román, F.; Roldán, C.; Trincado, P.; Ballesteros, C.; Carazo, C.; Vindel, A. Detection of Linezolid-Resistant Staphylococcus aureus with 23S rRNA and Novel L4 Riboprotein Mutations in a Cystic Fibrosis Patient in Spain. Antimicrob. Agents Chemother. 2013, 57, 2428–2429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tazi, A.; Chapron, J.; Touak, G.; Longo, M.; Hubert, D.; Collobert, G.; Dusser, D.; Poyart, C.; Morand, P.C. Rapid Emergence of Resistance to Linezolid and Mutator Phenotypes in Staphylococcus aureus Isolates from an Adult Cystic Fibrosis Patient. Antimicrob. Agents Chemother. 2013, 57, 5186–5188. [Google Scholar] [CrossRef] [Green Version]
- Locke, J.B.; Zuill, D.E.; Scharn, C.R.; Deane, J.; Sahm, D.F.; Denys, G.A.; Goering, R.V.; Shaw, K.J. Linezolid-Resistant Staphylococcus aureus Strain 1128105, the First Known Clinical Isolate Possessing the cfr Multidrug Resistance Gene. Antimicrob. Agents Chemother. 2014, 58, 6592–6598. [Google Scholar] [CrossRef] [Green Version]
- Caballero, J.D.D.; Pastor, M.D.; Vindel, A.; Máiz, L.; Yagüe, G.; Salvador, C.; Cobo, M.; Morosini, M.-I.; Del Campo, R.; Cantón, R.; et al. Emergence of cfr-Mediated Linezolid Resistance in a Methicillin-Resistant Staphylococcus aureus Epidemic Clone Isolated from Patients with Cystic Fibrosis. Antimicrob. Agents Chemother. 2015, 60, 1878–1882. [Google Scholar] [CrossRef] [Green Version]
- López-Hernández, I.; Valverde, M.D.; Díaz, N.B.; Pascual, A. First report of linezolid dependence in methicillin-resistant Staphylococcus aureus. Clin. Microbiol. Infect. 2015, 21, 650.e1–650.e4. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Stach, L.M.; Newland, J.G. Linezolid-Resistant Staphylococcus aureus in Children with Cystic Fibrosis: Table 1. J. Pediatr. Infect. Dis. Soc. 2015, 4, e163–e165. [Google Scholar] [CrossRef] [Green Version]
- Antonelli, A.; D’Andrea, M.M.; Borchi, B.; Vaggelli, G.; Cavallo, A.; Giovanetti, E.; Galano, A.; Brenciani, A.; Bartoloni, A.; Rossolini, G.M. Linezolid-resistant cfr-positive MRSA, Italy. J. Antimicrob. Chemother. 2016, 71, 2349–2351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouard, C.; Garnier, F.; Leraut, J.; Lepainteur, M.; Rahajamananav, L.; Languepin, J.; Ploy, M.-C.; Bourgeois-Nicolaos, N.; Doucet-Populaire, F. Emergence and Within-Host Genetic Evolution of Methicillin-Resistant Staphylococcus aureus Resistant to Linezolid in a Cystic Fibrosis Patient. Antimicrob. Agents Chemother. 2018, 62, e00720-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Angulo, V.A.; Herve, B.; Melo, J.; Sanhueza, C.; De La Fuente, S.; Aguirre, L.L.; Baysdorfer, C.; Ulloa, M.T. Isolation and first draft genome sequence of a linezolid-dependent Staphylococcus aureus clinical strain. Future Microbiol. 2020, 15, 1123–1129. [Google Scholar] [CrossRef]
- Quiles-Melero, I.; García-Perea, A.; De Pablos, M.; Gómez-Gil, R.; Mingorance, J. Resistance to linezolid in a methicillin-susceptible Staphylococcus aureus clinical isolate without previous exposure to oxazolidinones. Int. J. Med. Microbiol. 2012, 302, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Bosso, J.A.; Flume, P.A.; Gray, S.L. Linezolid Pharmacokinetics in Adult Patients with Cystic Fibrosis. Antimicrob. Agents Chemother. 2004, 48, 281–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durand, G.; Bes, M.; Meugnier, H.; Enright, M.C.; Forey, F.; Liassine, N.; Wenger, A.; Kikuchi, K.; Lina, G.; Vandenesch, F.; et al. Detection of New Methicillin-Resistant Staphylococcus aureus Clones Containing the Toxic Shock Syndrome Toxin 1 Gene Responsible for Hospital- and Community-Acquired Infections in France. J. Clin. Microbiol. 2006, 44, 847–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dauwalder, O.; Lina, G.; Durand, G.; Bes, M.; Meugnier, H.; Jarlier, V.; Coignard, B.; Vandenesch, F.; Etienne, J.; Laurent, F. Epidemiology of Invasive Methicillin-Resistant Staphylococcus aureus Clones Collected in France in 2006 and 2007. J. Clin. Microbiol. 2008, 46, 3454–3458. [Google Scholar] [CrossRef] [Green Version]
- Dodémont, M.; Argudín, M.; Willekens, J.; Vanderhelst, E.; Pierard, D.; Deyi, V.M.; Hanssens, L.; Franckx, H.; Schelstraete, P.; Leroux-Roels, I.; et al. Emergence of livestock-associated MRSA isolated from cystic fibrosis patients: Result of a Belgian national survey. J. Cyst. Fibros. 2019, 18, 86–93. [Google Scholar] [CrossRef]
- Winstanley, C.; O’Brien, S.; Brockhurst, M.A. Pseudomonas aeruginosa Evolutionary Adaptation and Diversification in Cystic Fibrosis Chronic Lung Infections. Trends Microbiol. 2016, 24, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Dupont, C.; Michon, A.-L.; Jumas-Bilak, E.; Nørskov-Lauritsen, N.; Chiron, R.; Marchandin, H. Intrapatient diversity of Achromobacter spp. involved in chronic colonization of Cystic Fibrosis airways. Infect. Genet. Evol. 2015, 32, 214–223. [Google Scholar] [CrossRef]
- Esposito, A.; Pompilio, A.; Bettua, C.; Crocetta, V.; Giacobazzi, E.; Fiscarelli, E.; Jousson, O.; Di Bonaventura, G. Evolution of Stenotrophomonas maltophilia in cystic fibrosis lung over chronic infection: A genomic and phenotypic population study. Front. Microbiol. 2017, 8, 1590. [Google Scholar] [CrossRef]
- Clark, S.T.; Diaz Caballero, J.; Cheang, M.; Coburn, B.; Wang, P.W.; Donaldson, S.L.; Zhang, Y.; Liu, M.; Keshavjee, S.; Yau, Y.C.W.; et al. Phenotypic diversity within a Pseudomonas aeruginosa population infecting an adult with cystic fibrosis. Sci. Rep. 2015, 5, 10932. [Google Scholar] [CrossRef]
- Dupont, C.; Jumas-Bilak, E.; Michon, A.-L.; Chiron, R.; Marchandin, H. Impact of High Diversity of Achromobacter Populations within Cystic Fibrosis Sputum Samples on Antimicrobial Susceptibility Testing. J. Clin. Microbiol. 2016, 55, 206–215. [Google Scholar] [CrossRef] [Green Version]
- Hogardt, M.; Heesemann, J. Microevolution of Pseudomonas aeruginosa to a Chronic Pathogen of the Cystic Fibrosis Lung. Curr. Top. Microbiol. Immunol. 2011, 358, 91–118. [Google Scholar] [CrossRef]
- Goerke, C.; Gressinger, M.; Endler, K.; Breitkopf, C.; Wardecki, K.; Stern, M.; Wolz, C.; Kahl, B.C. High phenotypic diversity in infecting but not in colonizing Staphylococcus aureus populations. Environ. Microbiol. 2007, 9, 3134–3142. [Google Scholar] [CrossRef] [PubMed]
- Vu-Thien, H.; Hormigos, K.; Corbineau, G.; Fauroux, B.; Corvol, H.; Moissenet, D.; Vergnaud, G.; Pourcel, C. Longitudinal survey of Staphylococcus aureus in cystic fibrosis patients using a multiple-locus variable-number of tandem-repeats analysis method. BMC Microbiol. 2010, 10, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirschhausen, N.; Block, D.; Bianconi, I.; Bragonzi, A.; Birtel, J.; Lee, J.C.; Dübbers, A.; Küster, P.; Kahl, J.; Peters, G.; et al. Extended Staphylococcus aureus persistence in cystic fibrosis is associated with bacterial adaptation. Int. J. Med. Microbiol. 2013, 303, 685–692. [Google Scholar] [CrossRef]
- Fitzgerald, J.R. Evolution of Staphylococcus aureus during human colonization and infection. Infect. Genet. Evol. 2014, 21, 542–547. [Google Scholar] [CrossRef] [PubMed]
- Protocole National de Diagnostic et de Soins (PNDS) Mucoviscidose. 2017. Available online: https://www.has-sante.fr/upload/docs/application/pdf/2017-09/pnds_2017_vf1.pdf (accessed on 25 April 2021).
- Monecke, S.; Slickers, P.; Ehricht, R. Assignment of Staphylococcus aureus isolates to clonal complexes based on microarray analysis and pattern recognition. FEMS Immunol. Med. Microbiol. 2008, 53, 237–251. [Google Scholar] [CrossRef] [Green Version]
- Predari, S.C.; Ligozzi, M.; Fontana, R. Genotypic identification of methicillin-resistant coagulase-negative staphylococci by polymerase chain reaction. Antimicrob. Agents Chemother. 1991, 35, 2568–2573. [Google Scholar] [CrossRef] [Green Version]
- Harmsen, D.; Claus, H.; Witte, W.; Rothgänger, J.; Turnwald, D.; Vogel, U. Typing of Methicillin-Resistant Staphylococcus aureus in a University Hospital Setting by Using Novel Software for spa Repeat Determination and Database Management. J. Clin. Microbiol. 2003, 41, 5442–5448. [Google Scholar] [CrossRef] [Green Version]
- Enright, M.C.; Day, N.P.J.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus Sequence Typing for Characterization of Methicillin-Resistant and Methicillin-Susceptible Clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [CrossRef] [Green Version]
- Crisóstomo, M.I.; Westh, H.; Tomasz, A.; Chung, M.; Oliveira, D.C.; De Lencastre, H. The evolution of methicillin resistance in Staphylococcus aureus: Similarity of genetic backgrounds in historically early methicillin-susceptible and -resistant isolates and contemporary epidemic clones. Proc. Natl. Acad. Sci. USA 2001, 98, 9865–9870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchandin, H.; Teyssier, C.; De Buochberg, M.S.; Jean-Pierre, H.; Carriere, C.; Jumas-Bilak, E. Intra-chromosomal heterogeneity between the four 16S rRNA gene copies in the genus Veillonella: Implications for phylogeny and taxonomy. Microbiol. 2003, 149, 1493–1501. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 20 February 2021).
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [PubMed]
- Hasman, H.; Clausen, P.T.L.C.; Kaya, H.; Hansen, F.; Knudsen, J.D.; Wang, M.; Holzknecht, B.J.; Samulioniené, J.; Røder, B.L.; Frimodt-Møller, N.; et al. LRE-Finder, a Web tool for detection of the 23S rRNA mutations and the optrA, cfr, cfr(B) and poxtA genes encoding linezolid resistance in enterococci from whole-genome sequences. J. Antimicrob. Chemother. 2019, 74, 1473–1476. [Google Scholar] [CrossRef]
- Woodford, N.; North, S.E.; Ellington, M.J.; Sharon, M. Detecting Mutations That Confer Oxazolidinone Resistance in Gram-Positive Bacteria. In Pyrosequencing Protocols; Springer Science and Business Media LLC: Berlin, Germany, 2007; Volume 373, pp. 103–114. [Google Scholar]
- Seemann, T. Snippy: Fast bacterial variant calling from NGS reads. Available online: https://github.com/tseemann/snippy (accessed on 20 February 2021).




{kind=link}
{kind=link}
{kind=link}
| Patient 1 | Patient 2 | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Jul. 2007 | Oct. 2010 | Jul. 2012 | Oct. 2014 | Jul. 2015 | Sep. 2016 | May 2007 | Feb. 2012 | Mar. 2014 | Oct. 2016 | |||||||||
| t002 | t002 | t002 | t002 | t045 | t045 | t002 | t002 | t002 | t002 | t002 | t002 | t002 | ||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | ||
| Patient 1 | 1 | 23 | 26 | > 1.000 | 29 | 33 | 54 | 41 | 37 | 22 | 55 | 51 | 37 | 32 | 74 | 47 | 66 | |
| 2 | 43 | 45 | 49 | 69 | 57 | 54 | 18 | 42 | 38 | 54 | 49 | 62 | 67 | 80 | ||||
| 3 | 29 | 34 | 51 | 44 | 41 | 43 | 76 | 72 | 41 | 36 | 95 | 52 | 67 | |||||
| 4 | >1000 | >1000 | ||||||||||||||||
| 5 | 11 | 51 | 49 | 52 | 46 | 78 | 74 | 42 | 39 | 97 | 57 | 73 | ||||||
| 6 | 60 | 71 | 57 | 48 | 82 | 78 | 48 | 43 | 101 | 64 | 77 | |||||||
| 7 | 71 | 68 | 71 | 103 | 100 | 68 | 63 | 121 | 79 | 97 | ||||||||
| 8 | 13 | 56 | 90 | 86 | 56 | 31 | 109 | 46 | 82 | |||||||||
| 9 | 53 | 87 | 83 | 53 | 28 | 106 | 43 | 79 | ||||||||||
| Patient 2 | 10 | 53 | 47 | 54 | 49 | 72 | 64 | 82 | ||||||||||
| 11 | 70 | 86 | 81 | 90 | 100 | 115 | ||||||||||||
| 12 | 81 | 80 | 87 | 95 | 111 | |||||||||||||
| 13 | 41 | 96 | 56 | 61 | ||||||||||||||
| 14 | 101 | 27 | 76 | |||||||||||||||
| 15 | 122 | 135 | ||||||||||||||||
| 16 | 92 | |||||||||||||||||
| 17 | ||||||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boudet, A.; Jay, A.; Dunyach-Remy, C.; Chiron, R.; Lavigne, J.-P.; Marchandin, H. In-Host Emergence of Linezolid Resistance in a Complex Pattern of Toxic Shock Syndrome Toxin-1-Positive Methicillin-Resistant Staphylococcus aureus Colonization in Siblings with Cystic Fibrosis. Toxins 2021, 13, 317. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13050317
Boudet A, Jay A, Dunyach-Remy C, Chiron R, Lavigne J-P, Marchandin H. In-Host Emergence of Linezolid Resistance in a Complex Pattern of Toxic Shock Syndrome Toxin-1-Positive Methicillin-Resistant Staphylococcus aureus Colonization in Siblings with Cystic Fibrosis. Toxins. 2021; 13(5):317. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13050317
Chicago/Turabian StyleBoudet, Agathe, Alexandre Jay, Catherine Dunyach-Remy, Raphaël Chiron, Jean-Philippe Lavigne, and Hélène Marchandin. 2021. "In-Host Emergence of Linezolid Resistance in a Complex Pattern of Toxic Shock Syndrome Toxin-1-Positive Methicillin-Resistant Staphylococcus aureus Colonization in Siblings with Cystic Fibrosis" Toxins 13, no. 5: 317. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13050317