Ciguatera in the Indian Ocean with Special Insights on the Arabian Sea and Adjacent Gulf and Seas: A Review




Abstract
:1. Introduction
2. Results
2.1. Environmental and Global Pressures in the Northern Part of the Indian Ocean
2.2. Ciguatera Causative Organisms Occurrence
3. Discussion
3.1. Toxin Production by Ciguatera in Indian Ocean Region and Adjoining Marginal Seas
3.2. Ciguatera Causative Organism’s Abundance
3.3. Ciguatera Causative Organisms’ Substrates and Co-Occurring Species
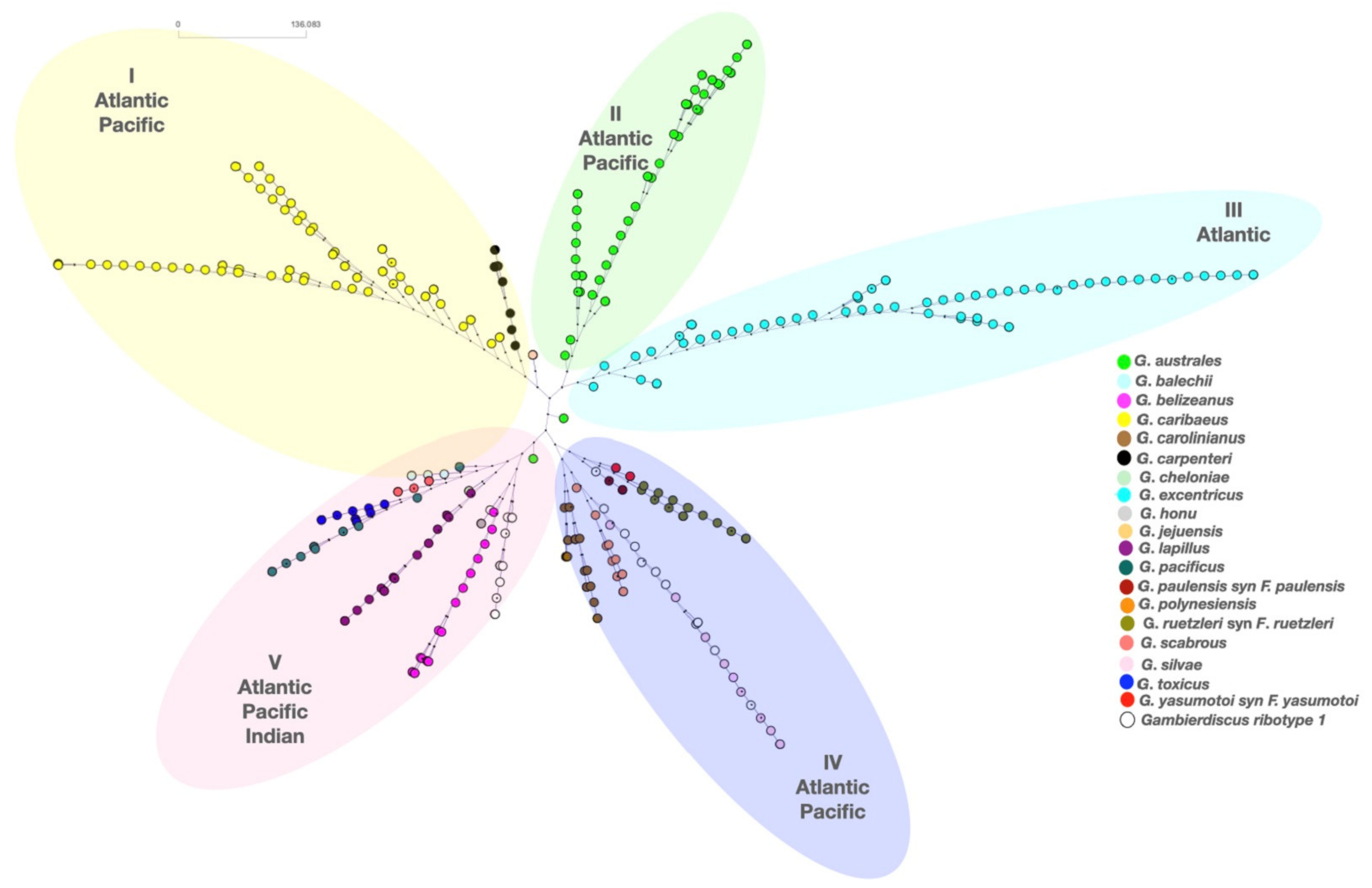
3.4. Ciguatera Causative Organism’s Morphology and Phylogeny
3.5. Vectors of Ciguatera in Indian Ocean Region
3.6. Ciguatera Monitoring in Indian Ocean Region
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Senthilkumaran, S.; Meenakshisundaram, R.; Michaels, A.; Suresh, P.; Thirumalaikolundusubramanian, P. Cardiovascular complications in ciguatera fish poisoning: A wake-up call. Heart Views Off. J. Gulf Heart Assoc. 2011, 12, 166. [Google Scholar] [CrossRef] [PubMed]
- Albinali, H. Ciguatera fish poisoning. Heart Views 2011, 12, 165. [Google Scholar] [CrossRef] [PubMed]
- Accoroni, S.; Romagnoli, T.; Colombo, F.; Pennesi, C.; Di Camillo, C.G.; Marini, M.; Battocchi, C.; Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; et al. Ostreopsis cf. ovata bloom in the northern Adriatic Sea during summer 2009: Ecology, molecular characterization and toxin profile. Mar. Pollut. Bull. 2011, 62, 2512–2519. [Google Scholar] [CrossRef] [PubMed]
- Chinain, M.; Darius, H.; Ung, A.; Cruchet, P.; Wang, Z.; Ponton, D.; Laurent, D.; Pauillac, S. Growth and toxin production in the ciguatera-causing dinoflagellate Gambierdiscus polynesiensis (Dinophyceae) in culture. Toxicon 2010, 56, 739–750. [Google Scholar] [CrossRef]
- Chinain, M.; Gatti, C.M.i.; Darius, H.T.; Quod, J.P.; Tester, P.A. Ciguatera poisonings: A global review of occurrences and trends. Harmful Algae 2021, 102, 101873. [Google Scholar] [CrossRef]
- Friedman, M.; Fleming, L.; Fernandez, M.; Bienfang, P.; Schrank, K.; Dickey, R.; Bottein, M.; Backer, L.; Ayyar, R.; Weisman, R. Ciguatera fish poisoning: Treatment, prevention and management. Mar. Drugs 2008, 6, 456–479. [Google Scholar] [CrossRef]
- Fleming, L.; Baden, D.; Bean, J.; Weisman, R.; Blythe, D. Marine Seafood Toxin Diseases: Issues in Epidemiology & Community Outreach; National Institute of Environmental Health Sciences: Durham, NC, USA, 1998. [Google Scholar]
- WHO, F.A. Report of the Expert Meeting on Ciguatera poisoning: Rome, 19–23 November 2018; Food Safety and Quality Series; World Health Organization: Rome, Italy, 2020. [Google Scholar]
- Parsons, M.; Richlen, M. An overview of ciguatera fish poisoning in the Bahamas. In The 15th Symposium on the Natural History of the Bahamas; Gerace Research Centre San Salvador Bahamas; A & A Printing Inc.: Tampa, Florida, USA, 2016. [Google Scholar]
- Chinain, M.; Gatti, C.; Roué, M.; Darius, H. Ciguatera-causing dinoflagellates in the genera gambierdiscus and fukuyoa: Distribution, ecophysiology and toxicology. In Dinoflagellates: Morphology, Life History and Ecological Significance; Subba Rao, D.V., Ed.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2020; pp. 405–457. [Google Scholar]
- Litaker, R.W.; Vandersea, M.W.; Faust, M.A.; Kibler, S.R.; Nau, A.W.; Holland, W.C.; Chinain, M.; Holmes, M.J.; Tester, P.A. Global distribution of ciguatera causing dinoflagellates in the genus Gambierdiscus. Toxicon 2010, 56, 711–730. [Google Scholar] [CrossRef]
- Holmes, M. Gambierdiscus yasumotoi sp. nov.(Dinophyceae), a toxic benthic dinoflagellate from southeastern Asia. J. Phycol. 1998, 34, 661–668. [Google Scholar] [CrossRef]
- Kashiwada, S.; Ishikawa, H.; Miyamoto, N.; Ohnishi, Y.; Magara, Y. Fish test for endocrine-disruption and estimation of water quality of Japanese rivers. Water Res. 2002, 36, 2161–2166. [Google Scholar] [CrossRef]
- Nishimura, T.; Sato, S.; Tawong, W.; Sakanari, H.; Uehara, K.; Shah, M.; Suda, S.; Yasumoto, T.; Taira, Y.; Yamaguchi, H. Genetic diversity and distribution of the ciguatera-causing dinoflagellate Gambierdiscus spp.(Dinophyceae) in coastal areas of Japan. PLoS ONE 2013, 8, e60882. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, L.; Gimenez, P.; Smith, K.; Harwood, T. Gambierdiscus cf. yasumotoi (Dinophyceae) isolated from New Zealand’s sub-tropical northern coastal waters. N. Z. J. Mar. Freshw. Res. 2014, 48, 303–310. [Google Scholar] [CrossRef] [Green Version]
- Casabianca, S.; Perini, F.; Casabianca, A.; Battocchi, C.; Giussani, V.; Chiantore, M.; Penna, A. Monitoring toxic Ostreopsis cf. ovata in recreational waters using a qPCR based assay. Mar. Pollut. Bull. 2014, 88, 102–109. [Google Scholar] [CrossRef]
- Cohu, S.; Thibaut, T.; Mangialajo, L.; Labat, J.P.; Passafiume, O.; Blanfune, A.; Simon, N.; Cottalorda, J.M.; Lemee, R. Occurrence of the toxic dinoflagellate Ostreopsis cf. ovata in relation with environmental factors in Monaco (NW Mediterranean). Mar. Pollut. Bull. 2011, 62, 2681–2691. [Google Scholar] [CrossRef]
- Parsons, M.L.; Brandt, A.L.; Ellsworth, A.; Leynse, A.K.; Rains, L.K.; Anderson, D.M. Assessing the use of artificial substrates to monitor Gambierdiscus populations in the Florida Keys. Harmful Algae 2017, 68, 52–66. [Google Scholar] [CrossRef]
- Yasumoto, T.; Nakajima, I.; Bagnis, R.; Adachi, R. Finding of a dinoflagellate as a likely culprit of ciguatera. Bull. Jpn. Soc. Sci. Fish. 1977, 43, 1021–1026. [Google Scholar] [CrossRef] [Green Version]
- Yasumoto, T.; Inoue, A.; Bagnis, R. Ecological survey of a toxic dinoflagellate associated with ciguatera. In Toxic Dinoflagellate Blooms; Taylor, D.L., Seliger, H.H., Eds.; Elsevier North Holland, Inc.: New York, NY, USA, 1979; pp. 221–224. [Google Scholar]
- Yasumoto, T.; Inoue, A.; Ochi, T.; Fujimoto, K.; Oshima, Y.; Fukuyo, Y.; Adachi, R.; Bagnis, R. Environmental studies on a toxic dinoflagellate responsible for ciguatera. Bull. Jpn. Soc. Sci. Fish. 1980, 46, 1397–1404. [Google Scholar] [CrossRef] [Green Version]
- Ballantine, D.L.; Tosteson, T.R.; Bardales, A.T. Population dynamics and toxicity of natural populations of benthic dinoflagellates in southwestern Puerto Rico. J. Exp. Mar. Biol. Ecol. 1988, 119, 201–212. [Google Scholar] [CrossRef]
- Lobel, P.S.; Anderson, D.M.; Durand-Clement, M. Assessment of Ciguatera Dinoflagellate Populations: Sample Variability and Algal Substrate Selection. Biol. Bull. 1988, 175, 94–101. [Google Scholar] [CrossRef]
- Bomber, J.W.; Rubio, M.G.; Norris, D.R. Epiphytism of dinoflagellates associated with the disease ciguatera: Substrate specificity and nutrition. Phycologia 1989, 28, 360–368. [Google Scholar] [CrossRef]
- Litaker, R.; Vandersea, M.; Faust, M.; Kibler, S.; Chinain, M.; Holmes, M.; Holland, W.; Tester, P. Taxonomy of Gambierdiscus including four new species, Gambierdiscus caribaeus, Gambierdiscus carolinianus, Gambierdiscus carpenteri and Gambierdiscus ruetzleri (Gonyaulacales, Dinophyceae). Phycologia 2009, 48, 344–390. [Google Scholar] [CrossRef]
- Rhodes, L.; Smith, K.; Murray, J.; Nishimura, T.; Finch, S. Ciguatera fish poisoning: The risk from an Aotearoa/New Zealand perspective. Toxins 2020, 12, 50. [Google Scholar] [CrossRef] [Green Version]
- Biotoxins, F.M. FAO Food and Nutrition Paper 80; Food and Agriculture Organization of the United Nations: Rome, Italy, 2004. [Google Scholar]
- De Rota, F. URDANETA¿ CIENTÍFICO? Institute de Historia and Cultura Naval: Madrid, Spain, 2009; 38p. [Google Scholar]
- Pearn, J. Around the rim: The role of surgeons in discovery and research in the Pacific rim. Part I. Surgeons in the Pacific: Expeditioners and expedition leaders. Aust. New Zealand J. Surg. 1994, 64, 38–44. [Google Scholar] [CrossRef]
- Lee, C. Fish poisoning with particular reference to ciguatera. J. Trop. Med. Hyg. 1980, 83, 93–97. [Google Scholar] [PubMed]
- Silva, S. Contribution al’etude du microplankton de dakar et des regions maritimes voisines. Bull. Inst. Fr. Afr. Noire. Ser. A. Sci. Nat. 1956, 18, 335–371. [Google Scholar]
- Guzman-Perez, S.; Park, D. Ciguatera toxins: Chemistry and detection. Food Sci. Technol. N.Y. Marcel Dekker 2000, 401–418. [Google Scholar]
- Tester, P.A.; Litaker, R.W.; Berdalet, E. Climate change and harmful benthic microalgae. Harmful Algae 2020, 91, 101655. [Google Scholar] [CrossRef]
- Landrigan, P.J.; Stegeman, J.J.; Fleming, L.E.; Allemand, D.; Anderson, D.M.; Backer, L.C.; Brucker-Davis, F.; Chevalier, N.; Corra, L.; Czerucka, D.; et al. Human Health and Ocean Pollution. Ann. Glob. Health 2020, 86, 1–64. [Google Scholar] [CrossRef]
- Kibler, S.; Litaker, R.; Holland, W.; Vandersea, M.; Tester, P. Growth of eight Gambierdiscus (Dinophyceae) species: Effects of temperature, salinity and irradiance. Harmful Algae 2012, 19, 1–14. [Google Scholar] [CrossRef]
- Gingold, D.B.; Strickland, M.J.; Hess, J.J. Ciguatera fish poisoning and climate change: Analysis of National Poison Center Data in the United States, 2001–2011. Environ. Health Perspect. 2014, 122, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Kibler, S.R.; Tester, P.A.; Kunkel, K.E.; Moore, S.K.; Litaker, R.W. Effects of ocean warming on growth and distribution of dinoflagellates associated with ciguatera fish poisoning in the Caribbean. Ecol. Model. 2015, 316, 194–210. [Google Scholar] [CrossRef] [Green Version]
- Yong, H.L.; Mustapa, N.I.; Lee, L.K.; Lim, Z.F.; Tan, T.H.; Usup, G.; Gu, H.; Litaker, R.W.; Tester, P.A.; Lim, P.T. Habitat complexity affects benthic harmful dinoflagellate assemblages in the fringing reef of Rawa Island, Malaysia. Harmful Algae 2018, 78, 56–68. [Google Scholar] [CrossRef]
- Hallegraeff, G.M.; Anderson, D.M.; Belin, C.; Bottein, M.-Y.D.; Bresnan, E.; Chinain, M.; Enevoldsen, H.; Iwataki, M.; Karlson, B.; McKenzie, C.H.; et al. Perceived global increase in algal blooms is attributable to intensified monitoring and emerging bloom impacts. Commun. Earth Environ. 2021, 2, 117. [Google Scholar] [CrossRef]
- Rhodes, L.; Smith, K.; Munday, R.; Selwood, A.; McNabb, P.; Holland, P.; Bottein, M. Toxic dinoflagellates (Dinophyceae) from Rarotonga, Cook Islands. Toxicon 2010, 56, 751–758. [Google Scholar] [CrossRef]
- Uddin, S.; Sultan, M.; Behbehani, M.; Al-Rashed, W.; Al-Shamroukh, D.; Al-Khabbaz, A.; Al-Yaegoub, A.; Al-Bahloul, M. A Remote Sensing-Based Early Warning System for Algal Blooms in Kuwait Bay and Coastal Waters; Project EM033C, Final Report, KISR12320; Kuwait Institute for Scientific Research, 2014; 145p. [Google Scholar]
- Saburova, M.; Polikarpov, I.; Al-Yamani, F. New records of the genus Gambierdiscus in marginal seas of the Indian Ocean. Mar. Biodivers. Rec. 2013, 6, e91. [Google Scholar] [CrossRef]
- Catania, D. The Prevalence of Benthic Dinoflagellates Associated with Ciguatera in the Central Red Sea. M.Sc. Thesis, King Abdullah University of Science and Technology, Thuwal, Saudi Arabia, 2012; 40p. [Google Scholar]
- Catania, D.; Richlen, M.; Mak, Y.; Morton, S.; Laban, E.; Xu, Y.; Anderson, D.; Chan, L.; Berumen, M. The prevalence of benthic dinoflagellates associated with ciguatera fish poisoning in the central Red Sea. Harmful Algae 2017, 68, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Manche, C. A Remote Sensing Based Early Warning System for Algal Blooms in Kuwait Bay and Coastal Waters. Master’s Thesis, Western Michigan University, Kalamazoo, MI, USA, 2014. Available online: https://scholarworks.wmich.edu/masters_theses/546 (accessed on 1 June 2021).
- Quod, J.-P.; Turquet, J.; Conejero, S.; Ralijaona, C. Ciguatera risk assessment in the Indian Ocean following the 1998 coral bleaching event. Coral Reef Degrad. Indian Ocean. Status Rep. 2000, 166–168. [Google Scholar]
- Bagnis, R. L’ichtyosarcotoxisme de type ciguatera: Phénomène complexe de biologie marine et humaine. Oceanol. Acta 1981, 4, 375–387. [Google Scholar]
- Behbehani, M.; Uddin, S.; Dupont, S.; Sajid, S.; Al-Musalam, L.; Al-Ghadban, A. Response of corals Acropora pharaonis and Porites lutea to changes in pH and temperature in the Gulf. Sustainability 2019, 11, 3156. [Google Scholar] [CrossRef] [Green Version]
- Reguera, B.; Alonso, R.; Moreira, A.; Méndez, S.; Dechraoui-Bottein, M.-Y. Guide for Designing and Implementing a Plan to Monitor Toxin-Producing Microalgae. In Intergovermental Oceanographic Commission Manuals and Guides, 2nd ed.; UNESCO: Paris, France; IAEA: Vienna, Austria, 2016; 66p, Available online: http://hdl.handle.net/11329/304 (accessed on 1 June 2021).
- Jean Turquet, J.-P.Q.; Ten-Hage, L.; Dahalani, Y.; Wendling, B. Example of a Gambierdiscus toxicus flare-up following the 1998 coral bleaching event in Mayotte Island (Comoros, south-west Indian Ocean). In Proceedings of the Harmful Algal Blooms 2000: Proceedings of the Ninth International Conference on Harmful Algal Blooms, Hobart, Australia, 7–11 February 2000. [Google Scholar]
- Uddin, S.; Gevao, B.; Al-Ghadban, A.; Nithyanandan, M.; Al-Shamroukh, D. Acidification in Arabian Gulf–Insights from pH and temperature measurements. J. Environ. Monit. 2012, 14, 1479–1482. [Google Scholar] [CrossRef]
- Al-Musalam, L.; Uddin, S. Effect of Ocean Acidification on Growth and Abundance of Penaeus Semisulcatus in the Northern Arabian Gulf; Annual Progress Report No. 1(EM078C); Kuwait Insitute for Scientific Research: Kuwait City, Kuwait, 2018; 27p. [Google Scholar]
- Al-Musallam, L.; Uddin, S.; Al-Dakkor, S.; Kumar, V. Effect of ocean acidification and ocean warming on the growth and survival of Penaeus semisulcatus Post-Larvae. J Earth Env. Sci 2019, 3. [Google Scholar] [CrossRef]
- Elkadiri, R.; Manche, C.; Sultan, M.; Al-Dousari, A.; Uddin, S.; Chouinard, K.; Abotalib, A.Z. Development of a Coupled Spatiotemporal Algal Bloom Model for Coastal Areas: A Remote Sensing and Data Mining-Based Approach. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2016, 9, 5159–5171. [Google Scholar] [CrossRef]
- Heil, C.A.; Glibert, P.M.; Al-Sarawi, M.A.; Faraj, M.; Behbehani, M.; Husain, M. First record of a fish-killing Gymnodinium sp. bloom in Kuwait Bay, Arabian Sea: Chronology and potential causes. Mar. Ecol. Prog. Ser. 2001, 214, 15–23. [Google Scholar] [CrossRef]
- Glibert, P.; Landsberg, J.; Evans, J.; Al-Sarawi, M.; Faraj, M.; Al-Jarallah, M.; Haywood, A.; Ibrahem, S.; Klesius, P.; Powell, C. A fish kill of massive proportion in Kuwait Bay, Arabian Gulf, 2001: The roles of bacterial disease, harmful algae, and eutrophication. Harmful Algae 2002, 1, 215–231. [Google Scholar] [CrossRef]
- Walsh, J.; Steidinger, K. Saharan dust and Florida red tides: The cyanophyte connection. J. Geophys. Res. Ocean. 2001, 106, 11597–11612. [Google Scholar] [CrossRef] [Green Version]
- Richlen, M.L.; Morton, S.L.; Jamali, E.A.; Rajan, A.; Anderson, D.M. The catastrophic 2008–2009 red tide in the Arabian gulf region, with observations on the identification and phylogeny of the fish-killing dinoflagellate Cochlodinium polykrikoides. Harmful Algae 2010, 9, 163–172. [Google Scholar] [CrossRef]
- Berland, B.; Grzebyk, D.; Thomassin, B.-A. Benthic dinoflagellates from the coral reef lagoon of Mayotte Island (SW Indian Ocean); identification, toxicity and preliminary ecophysiological study. Bull.-Soc. Pathol. Exot. 1992, 85, 453. [Google Scholar] [PubMed]
- Grzebyk, D.; Berland, B.; Thomassin, B.A.; Bosi, C.; Arnoux, A. Ecology of ciguateric dinoflagellates in the coral reef complex of Mayotte Island (SW Indian Ocean). J. Exp. Mar. Biol. Ecol. 1994, 178, 51–66. [Google Scholar] [CrossRef]
- Thomassin, B.; ME, A.H.; Quod, J.; Maggiorani, J.; Berland, B.; Grzebyk, D.; Coqueugniot, J. Evolution of Gambierdiscus toxicus populations in the coral reef complex of Mayotte Island (SW Indian Ocean) during the 1985–1991 period. Bull. Soc. Pathol. Exot. 1992, 85, 449–452. [Google Scholar]
- Quod, J.; Prunaux, O.; Guignard, A. Les empoisonnements par poissons tropicaux à La Réunion: Synthèse et perspectives. Rev. Méd. Vét. 1990, 141, 1005–1009. [Google Scholar]
- Quod, J.; Bourdeau, P.; Turquet, J.; Guignard, A. La ciguatera dans les DOM-TOM: Aspects épidémiologiques et physiopathologiques. Rev. Méd. Vét. 1994, 170, 141–146. [Google Scholar]
- Quod, J.; Turquet, J. Ciguatera in Reunion Island (SW Indian Ocean): Epidemiology and clinical patterns. Toxicon 1996, 34, 779–785. [Google Scholar] [CrossRef]
- Ten-Hage, L.; Turquet, J.; Quod, J.-P.; Couté, A. An overview of the biodiversity of benthic diniflagellates from La Reunion Island (France, South-west Indian Ocean). In 9th Int Conference on Harmful Algal Blooms (HAB 2000); Hobart, Tasmania, 2000. [Google Scholar]
- Lugomela, C. Autecology of the Toxic Dinoflagellate Gambierdiscus toxicus Adachi et Fukyo (Dinophyceae) in Central Coastal Areas of Tanzania. West. Indian Ocean J. Mar. Sci. 2006, 5, 213–221. [Google Scholar] [CrossRef]
- Gómez, F.; Qiu, D.; Lopes, R.M.; Lin, S. Fukuyoa paulensis gen. et sp. nov., a New Genus for the Globular Species of the Dinoflagellate Gambierdiscus (Dinophyceae). PLoS ONE 2015, 10, e0119676. [Google Scholar] [CrossRef]
- Saburova, M.; Igor, P.; Al-Yamani, F. Gambierdiscus in Kuwait. Harmful Algae News 2013, 47, 22–23. [Google Scholar]
- Munir, S.; Siddiqui, P.; Morton, S.L. The occurrence of the ciguatera fish poisoning producing dinoflagellate genus Gambierdiscus in Pakistan waters. Algae 2011, 26, 317–325. [Google Scholar] [CrossRef]
- Polikarpov, I.; Saburova, M.; Al-Yamani, F. Diversity and distribution of winter phytoplankton in the Arabian Gulf and the Sea of Oman. Cont. Shelf Res. 2016, 119, 85–99. [Google Scholar] [CrossRef]
- Naik, R.K.; Hegde, S.; Anil, A.C. Dinoflagellate community structure from the stratified environment of the Bay of Bengal, with special emphasis on harmful algal bloom species. Environ. Monit. Assess. 2011, 182, 15–30. [Google Scholar] [CrossRef]
- Rajeish, M.; Shekar, M.; Madhushree, H.; Venugopal, M. Presumptive case of ciguatera fish poisoning in Mangalore, India. Curr. Sci. 2016, 111, 1543–1547. [Google Scholar] [CrossRef]
- Rajisha, R.; Kishore, P.; Panda, S.K.; Ravishankar, C.N.; Kumar, K.A. Confirmation of ciguatoxin fish poisoning in red snapper, Lutjanus bohar (Forsskål, 1775) by mouse bioassay. Fish. Technol. 2017, 54, 287–290. [Google Scholar]
- Karunasagar, I.; Turner, A.D.; Maskrey, B.; Robertson, A.; Shivanagouda Hosagoudar, S.; Rai, P.; Adappa, S.; Hiremath, S.; Kogaluru Shivakumaraswamy, S.; Dechraoui Bottein, M.Y.; et al. Report of a major outbreak of ciguatera fish poisoning in Mangalore, India. In Proceedings of the 18th International Conference on Harmful Algae; Rome, Italy, 2018; p. 482. [Google Scholar]
- Rajisha, R.; Kishore, P.; Panda, S.K.; Harikrisnan, G.; Ajitha, K.C.; Suresh, M.K.; Chowdhury, L.M.; Ravishankar, C.N.; Kumar, K.A. Incidence of ciguatoxin fish poisoning in Trivandrum, India. Indian J. Fish. 2017, 64, 129–133. [Google Scholar] [CrossRef]
- Hamilton, B.; Hurbungs, M.; Jones, A.; Lewis, R.J. Multiple ciguatoxins present in Indian Ocean reef fish. Toxicon 2002, 40, 1347–1353. [Google Scholar] [CrossRef]
- Hamilton, B.; Hurbungs, M.; Vernoux, J.-P.; Jones, A.; Lewis, R.J. Isolation and characterisation of Indian Ocean ciguatoxin. Toxicon 2002, 40, 685–693. [Google Scholar] [CrossRef]
- Diogène, J.; Reverté, L.; Rambla-Alegre, M.; Del Río, V.; De La Iglesia, P.; Campàs, M.; Palacios, O.; Flores, C.; Caixach, J.; Ralijaona, C. Identification of ciguatoxins in a shark involved in a fatal food poisoning in the Indian Ocean. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef]
- Roeder, K.; Erler, K.; Kibler, S.; Tester, P.; Van The, H.; Nguyen-Ngoc, L.; Gerdts, G.; Luckas, B. Characteristic profiles of Ciguatera toxins in different strains of Gambierdiscus spp. Toxicon 2010, 56, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Pisapia, F.; Holland, W.C.; Hardison, D.R.; Litaker, R.W.; Fraga, S.; Nishimura, T.; Adachi, M.; Nguyen-Ngoc, L.; Séchet, V.; Amzil, Z.; et al. Toxicity screening of 13 Gambierdiscus strains using neuro-2a and erythrocyte lysis bioassays. Harmful Algae 2017, 63, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Friedemann, M. Ciguatera fish poisoning outbreaks from 2012 to 2017 in Germany caused by snappers from India, Indonesia, and Vietnam. J. Consum. Prot. Food Saf. 2019, 14, 71–80. [Google Scholar] [CrossRef]
- Yogi, K.; Oshiro, N.; Inafuku, Y.; Hirama, M.; Yasumoto, T. Detailed LC-MS/MS Analysis of Ciguatoxins Revealing Distinct Regional and Species Characteristics in Fish and Causative Alga from the Pacific. Anal. Chem. 2011, 83, 8886–8891. [Google Scholar] [CrossRef] [PubMed]
- Clausing, R.; Chinain, M.; Bottein, M. Practical Sampling Guidance for Determination of Ciguatoxin in Fish. In Guide for Designing and Implementing a Plant to Monitor Toxin-Producing Microalgae, 2nd ed.; IOC Manuals and Guides, no. 59; Intergovernmental Oceanographic Commission (IOC) of UNESCO: Paris, France; International Atomic Energy Agency (IAEA): Vienna, Austria, 2016; pp. 51–63. [Google Scholar]
- Hurbungs, M.D.; Jayabalanm, N.; Chineah, V. Seasonal distribution of potentially toxic benthic dinoflagellates in the lagoon of Trou aux Biches, Mauritius. In Proceedings of the 5th Annual Meeting of Agricultural Scientists, Reduit, Mauritius, 3–4 May 2001; Lalouette, J.A., Bachraz, D.Y., Eds.; The Food and Agricultural Research Council: Reduit, Mauritius, 2002; pp. 211–217. [Google Scholar]
- Quod, J.; Turquet, J.; Diogene, G.; Fessard, V. Screening of extracts of dinoflagellates from coral reefs (Reunion Island, SW Indian Ocean), and their biological activities. Harmful Mar. Algal Bloom. 1995, 815–820. [Google Scholar]
- Turquet, J. Assemblage of benthic dinofagellates and monitoring of harmful species in Reunion Island, SW Indian Ocean, 1993–1996. Harmful Algae 1998, 44–47. [Google Scholar]
- Roué, M.; Smith, K.; Sibat, M.; Viallon, J.; Henry, K.; Ung, A.; Biessy, L.; Hess, P.; Darius, H.; Chinain, M. Assessment of Ciguatera and Other Phycotoxin-Related Risks in Anaho Bay (Nuku Hiva Island, French Polynesia): Molecular, Toxicological, and Chemical Analyses of Passive Samplers. Toxins 2020, 12, 321. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.; Kohli, G.; Murray, S.; Rhodes, L. Assessment of the metabarcoding approach for community analysis of benthic-epiphytic dinoflagellates using mock communities. N. Z. J. Mar. Freshw. Res. 2017, 51, 555–576. [Google Scholar] [CrossRef]
- Rhodes, L.; Smith, K.; Verma, A.; Murray, S.; Harwood, D.; Trnski, T. The dinoflagellate genera Gambierdiscus and Ostreopsis from subtropical Raoul Island and North Meyer Island, Kermadec Islands. N. Z. J. Mar. Freshw. Res. 2017, 51, 490–504. [Google Scholar] [CrossRef]
- Hoppenrath, M.; Kretzschmar, A.; Kaufmann, M.; Murray, S. Morphological and molecular phylogenetic identification and record verification of Gambierdiscus excentricus (Dinophyceae) from Madeira Island (NE Atlantic Ocean). Mar. Biodivers. Rec. 2019, 12, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, L.; Smith, K. A checklist of the benthic and epiphytic marine dinoflagellates of New Zealand, including Rangitāhua/Kermadec Islands. N. Z. J. Mar. Freshw. Res. 2019, 53, 258–277. [Google Scholar] [CrossRef]
- Faust, M. Observation of sand-dwelling toxic dinoflagellates (Dinophyceae) from widely differing sites, including two new species. J. Phycol. 1995, 31, 996–1003. [Google Scholar] [CrossRef]
- Adachi, R.; Fukuyo, Y. The thecal structure of a marine toxic dinoflagellate Gambierdiscus toxicus gen. et sp. nov. collected in a ciguatera endemic area. Bull. Jpn. Soc. Sci. Fish. 1979, 45, 67–71. [Google Scholar] [CrossRef] [Green Version]
- Chinain, M.; Faust, M.A.; Pauillac, S. Morphology and molecular analyses of three toxic species of Gambierdiscus (Dinophyceae): G. pacificus sp. nov., G. australes, sp. nov. and G. polynesiensis, sp. nov. J. Phycol. 1999, 35, 1282–1296. [Google Scholar] [CrossRef]
- Xu, Y.; Richlen, M.L.; Morton, S.L.; Mak, Y.L.; Chan, L.L.; Tekiau, A.; Anderson, D.M. Distribution, abundance and diversity of Gambierdiscus spp. from a ciguatera-endemic area in Marakei, Republic of Kiribati. Harmful Algae 2014, 34, 56–68. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Mak, Y.; Lu, C.; Mei, H.; Wu, J.; Lee, W.; Chan, L.; Lim, P.; Mustapa, N.; Lim, H. Taxonomic assignment of the benthic toxigenic dinoflagellate Gambierdiscus sp. type 6 as Gambierdiscus balechii (Dinophyceae), including its distribution and ciguatoxicity. Harmful Algae 2017, 67, 107–118. [Google Scholar] [CrossRef]
- Richlen, M.; Morton, S.; Barber, P.; Lobel, P. Phylogeography, morphological variation and taxonomy of the toxic dinoflagellate Gambierdiscus toxicus (Dinophyceae). Harmful Algae 2008, 7, 614–629. [Google Scholar] [CrossRef]
- Fraga, S.; Rodríguez, F.; Caillaud, A.; Diogène, J.; Raho, N.; Zapata, M. Gambierdiscus excentricus sp. nov.(Dinophyceae), a benthic toxic dinoflagellate from the Canary Islands (NE Atlantic Ocean). Harmful Algae 2011, 11, 10–22. [Google Scholar] [CrossRef] [Green Version]
- Fraga, S.; Rodríguez, F. Genus Gambierdiscus in the Canary Islands (NE Atlantic Ocean) with description of Gambierdiscus silvae sp. nov., a new potentially toxic epiphytic benthic dinoflagellate. Protist 2014, 165, 839–853. [Google Scholar] [CrossRef] [PubMed]
- Fraga, S.; Rodríguez, F.; Riobó, P.; Bravo, I. Gambierdiscus balechii sp. nov (Dinophyceae), a new benthic toxic dinoflagellate from the Celebes Sea (SW Pacific Ocean). Harmful Algae 2016, 58, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, S.; Melo, G.; Salgueiro, F.; Diniz, B.; Fraga, S. Morphology of Gambierdiscus excentricus (Dinophyceae) with emphasis on sulcal plates. Phycologia 2015, 54, 628–639. [Google Scholar] [CrossRef]
- Rodríguez, F.; Fraga, S.; Ramilo, I.; Rial, P.; Figueroa, R.; Riobó, P.; Bravo, I. Canary Islands (NE Atlantic) as a biodiversity ‘hotspot’of Gambierdiscus: Implications for future trends of ciguatera in the area. Harmful Algae 2017, 67, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Leung, P.; Yan, M.; Lam, V.; Yiu, S.; Chen, C.; Murray, J.; Harwood, D.; Rhodes, L.; Lam, P.; Wai, T. Phylogeny, morphology and toxicity of benthic dinoflagellates of the genus Fukuyoa (Goniodomataceae, Dinophyceae) from a subtropical reef ecosystem in the South China Sea. Harmful Algae 2018, 74, 78–97. [Google Scholar] [CrossRef]
- Rhodes, L.; Smith, K.; Verma, A.; Curley, B.; Harwood, D.; Murray, S.; Kohli, G.; Solomona, D.; Rongo, T.; Munday, R. A new species of Gambierdiscus (Dinophyceae) from the south-west Pacific: Gambierdiscus honu sp. nov. Harmful Algae 2017, 65, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.; Biessy, L.; Argyle, P.; Trnski, T.; Halafihi, T.; Rhodes, L. Molecular identification of Gambierdiscus and Fukuyoa (Dinophyceae) from environmental samples. Mar. Drugs 2017, 15, 243. [Google Scholar] [CrossRef] [Green Version]
- Tudó, À.; Toldrà, A.; Rey, M.; Todolí, I.; Andree, K.; Fernández-Tejedor, M.; Campàs, M.; Sureda, F.; Diogène, J. Gambierdiscus and Fukuyoa as potential indicators of ciguatera risk in the Balearic Islands. Harmful Algae 2020, 99, 101913. [Google Scholar] [CrossRef]
- Bravo, I.; Rodríguez, F.; Ramilo, I.; Afonso-Carrillo, J. Epibenthic Harmful Marine Dinoflagellates from Fuerteventura (Canary Islands), with Special Reference to the Ciguatoxin-Producing Gambierdiscus. J. Mar. Sci. Eng. 2020, 8, 909. [Google Scholar] [CrossRef]
- Jang, S.H.; Jeong, H.J.; Yoo, Y.D. Gambierdiscus jejuensis sp. nov., an epiphytic dinoflagellate from the waters of Jeju Island, Korea, effect of temperature on the growth, and its global distribution. Harmful Algae 2018, 80, 149–157. [Google Scholar] [CrossRef]
- Nishimura, T.; Sato, S.; Tawong, W.; Sakanari, H.; Yamaguchi, H.; Adachi, M. Morphology of G ambierdiscus scabrosus sp. nov.(G onyaulacales): A new epiphytic toxic dinoflagellate from coastal areas of Japan. J. Phycol. 2014, 50, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Tawong, W.; Nishimura, T.; Sakanari, H.; Sato, S.; Yamaguchi, H.; Adachi, M. Characterization of G ambierdiscus and C oolia (D inophyceae) isolates from T hailand based on morphology and phylogeny. Phycol. Res. 2015, 63, 125–133. [Google Scholar] [CrossRef]
- Larsson, M.; Harwood, T.; Lewis, R.; SWA, H.; Doblin, M. Toxicological characterization of Fukuyoa paulensis (Dinophyceae) from temperate Australia. Phycol. Res. 2019, 67, 65–71. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, Z.; Cen, J.; Li, Y.; Wang, H.; Lu, S. First report of three benthic dinoflagellates, Gambierdiscus pacificus, G. australes and G. caribaeus (Dinophyceae), from Hainan Island, South China Sea. Phycol. Res. 2016, 64, 259–273. [Google Scholar] [CrossRef]
- Kretzschmar, A.; Verma, A.; Harwood, T.; Hoppenrath, M.; Murray, S. Characterization of Gambierdiscus lapillus sp. nov.(Gonyaulacales, Dinophyceae): A new toxic dinoflagellate from the Great Barrier Reef (Australia). J. Phycol. 2017, 53, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.; Momigliano, P.; Heimann, K.; Blair, D. Molecular phylogenetics and morphology of Gambierdiscus yasumotoi from tropical eastern Australia. Harmful Algae 2014, 39, 242–252. [Google Scholar] [CrossRef]
- Dickey, R.; Plakas, S. Ciguatera: A public health perspective. Toxicon 2010, 56, 123–136. [Google Scholar] [CrossRef]
- Wasay, M.; Sarangzai, A.; Siddiqi, A.; Nizami, Q. Ciguatera fish poisoning with elevated muscle enzymes and abnormal spinal MRI. Southeast Asian J. Trop. Med. Public Health 2008, 39, 307. [Google Scholar]
- Rajisha, R.; Kishore, P.; Panda, S.; Kumar, K.A. Ciguatoxin-An Emerging Biological Hazard among Reef Fishes of India. Fish. Tech. 2018, 55, 153–167. [Google Scholar]
- Saraya, A.; Sintunawa, C.; Wacharapluesadee, S.; Swangpun, K.; Dumrongchua, S.; Wilde, H.; Hemachudha, T. Marine Fish Toxins in Thailand: Report of 6 Suspected Ciguatera Cases. Case Rep. Clin. Med. 2014, 3, 7. [Google Scholar] [CrossRef] [Green Version]
- Sozzi, G.; Marotta, P.; Aldeghi, D.; Tredici, G.; Calvi, L. Polyneuropathy Secondary to Ciguatoxin Poisoning. Ital. J. Neurol. Sci. 1988, 9, 491–495. [Google Scholar] [CrossRef] [PubMed]
- De Haro, L.; Pommier, P.; Valli, M. Emergence of Imported Ciguatera in Europe: Report of 18 Cases at the Poison Control Centre of Marseille. J. Toxicol. Clin. Toxicol. 2003, 41, 927–930. [Google Scholar] [CrossRef]
- Habermehl, G.G.; Krebs, H.C.; Rasoanaivo, P.; Ramialiharisoa, A. Severe ciguatera poisoning in Madagascar: A case report. Toxicon 1994, 32, 1539–1542. [Google Scholar] [CrossRef]
- Anonymous. Mangaluru: Khader Warns against Consumption of Red Snapper, Puffer Fish Heads, in Daijiworld Published on 6 October 2016: Mangaluru, India. Available online: https://www.daijiworld.com/news/newsDisplay?newsID=417564 (accessed on 1 June 2021).
- Jin, D.; Kite-Powell, H.; Hoagland, P.; Solow, A. A bioeconomic analysis of traditional fisheries in the Red Sea. Mar. Resour. Econ. 2012, 27, 137–148. [Google Scholar] [CrossRef]
- Anonymous. Fishbase. Available online: https://www.fishbase.de (accessed on 1 June 2021).
- Mattei, C.; Vetter, I.; Eisenblätter, A.; Krock, B.; Ebbecke, M.; Desel, H.; Zimmermann, K. Ciguatera fish poisoning: A first epidemic in Germany highlights an increasing risk for European countries. Toxicon 2014, 91, 76–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Location | Marginal Seas | GPS Coordinates | Sampling Depth | Temperature | Associated Species | Salinities | Reference |
|---|---|---|---|---|---|---|---|---|
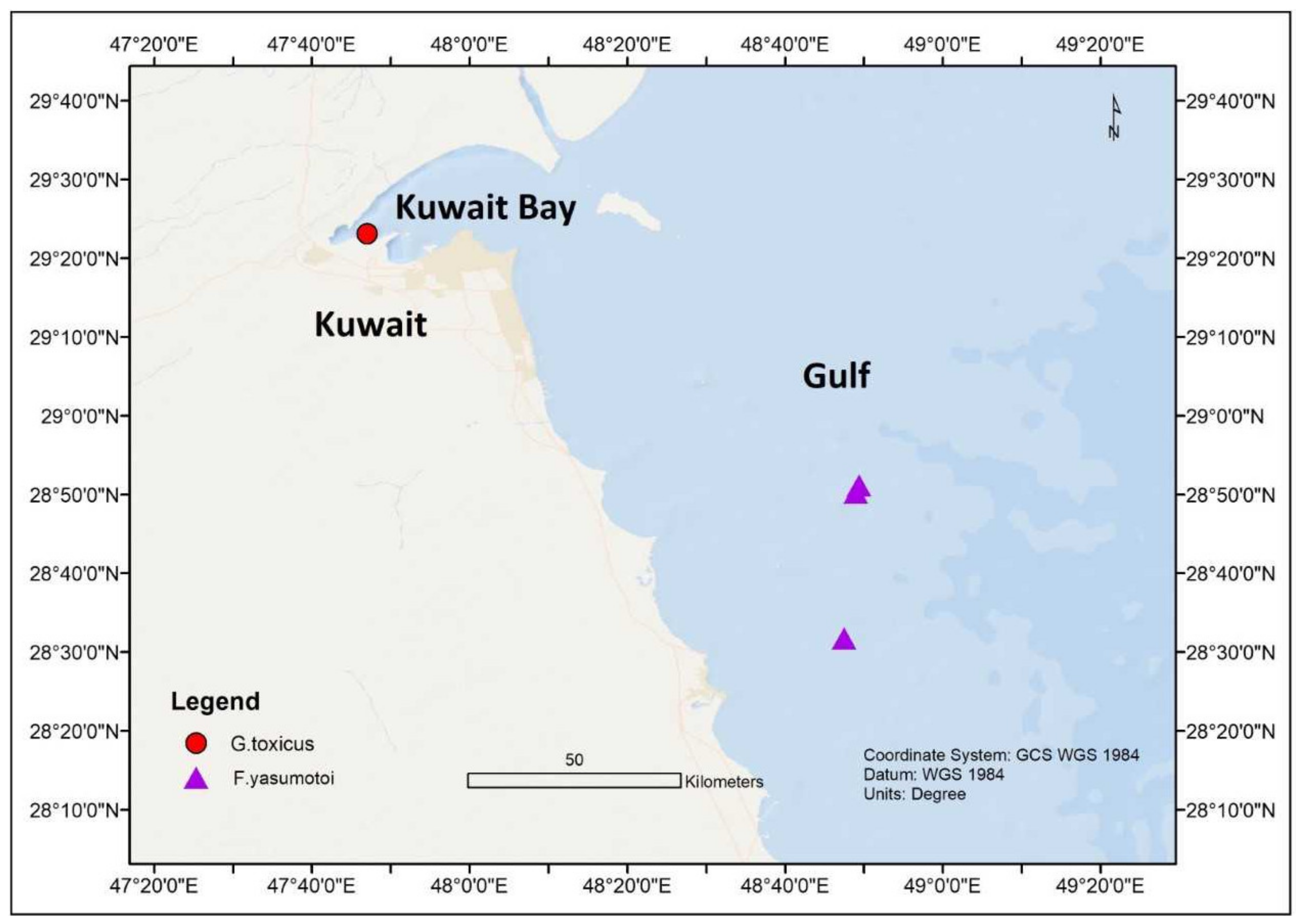
| G. toxicus | Kuwait | Persian Gulf | 29.3860° N, 47.7830° E | 17.59 °C | Karenia selliformis, Karenia brevis, Prorocentrum lima, Phalacroma rotundatum, Tripos muelleri and Mesodinium rubrum | [41,45] | ||
| F. yasumotoi | Qit’at Julai’ah, Kuwait | Persian Gulf | 28.8370° N, 48.8160° E | 1–3 m | 21–22.3 °C | Padina tetrastomatica, Sargassum oligocystum, Chaetomorpha sp., Amphidinium carterae, Coolia monotis, Ostreopsis sps., Prorocentrum formosum, P. tsawwassenense, Blixaea quinquecornis, Adenoides eludens and Cabra matta | 41.2–42.4 | [42] |
| F. yasumotoi | Semi protected shallow lagoons, Kuwait | Persian Gulf | 28.8530° N, 48.8230° E | 1–3 m | 23.5–25 °C | 41.2–42.4 | [42] | |
| F. yasumotoi | Southern coast, Kuwait | Persian Gulf | 28.8530° N, 48.8230° E | 1–3 m | 23.5–25 °C | 42.1–42.8 | [42] | |
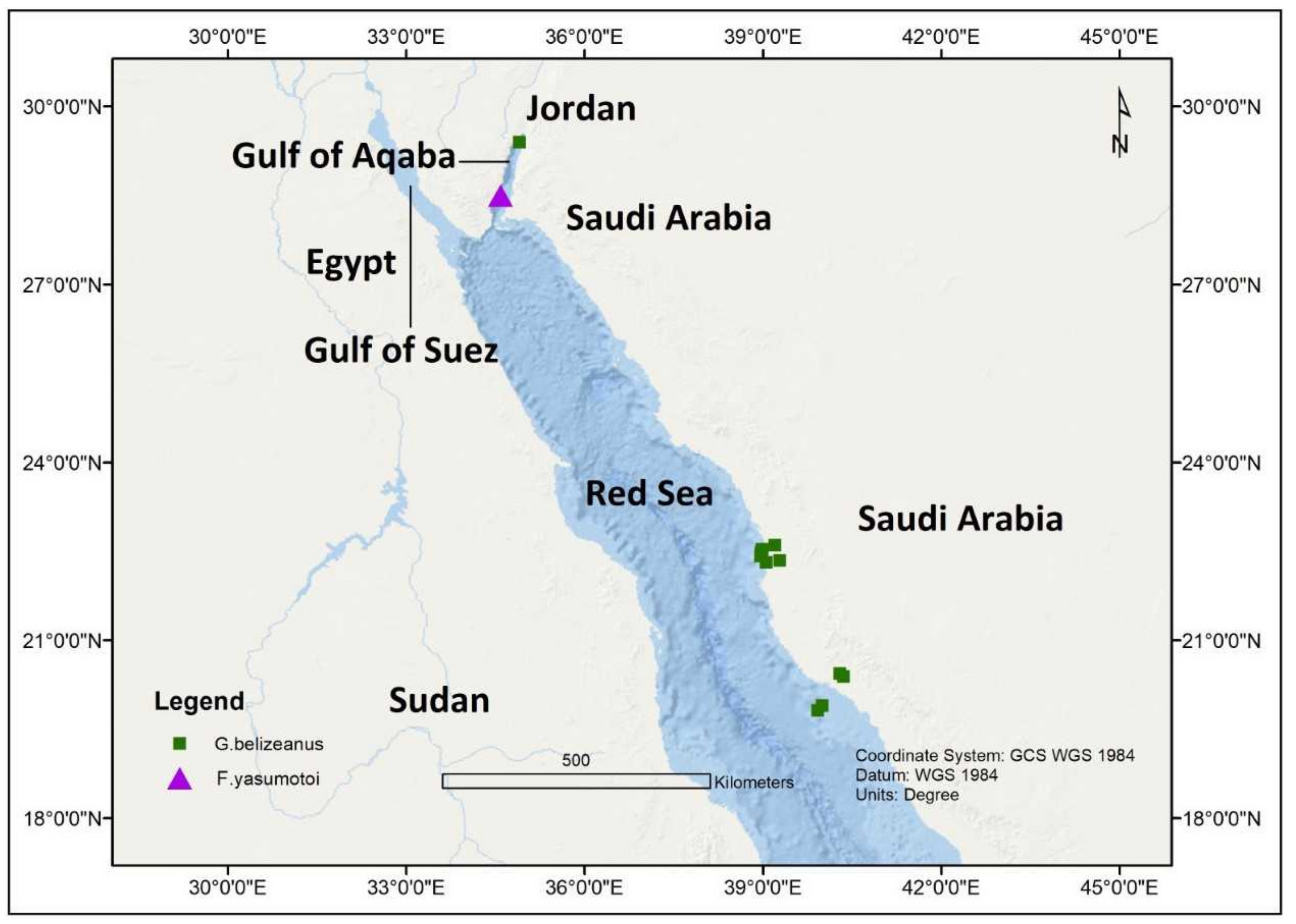
| F. yasumotoi | Jordan | Gulf of Aqaba | 29.8250° N,34.8580° E | 1.5–2 m | 24–25 °C | NA | 40.0 | [42] |
| G. belizeanus | Jordan | Gulf of Aqaba | 29.8250° N, 34.8580° E | 1.5–2 m | 24–25 °C | NA | 40.0 | [42] |
| G. belizeanus | Qita Al Kirsh, Saudi Arabia | Red Sea | 22.6058° N, 39.1980° E | 0.5–0.6 m | 29.3 °C | Turbinaria decurrens and Halimeda sp. | 38.4 | [43,44] |
| G. belizeanus | Sh’ib Nazar, Saudi Arabia | Red Sea | 22.4916° N, 38.9722° E | 0.4 m | 30.1 °C | Turbinaria decurrens | 28.9 | [43,44] |
| G. belizeanus | Malathu Reef, Saudi Arabia | Red Sea | 19.8194° N, 39.9194° E | 0.5–0.7 m | NA | Turbinaria decurrens | NA | [43,44] |
| G. belizeanus | Marmar Reef, Saudi Arabia | Red Sea | 19.9038° N, 39.9947° E | 0.8 m | NA | Turbinaria decurrens | NA | [43,44] |
| G. belizeanus | Al Fahal, Saudi Arabia | Red Sea | 22.5386° N, 38.9813° E | 0.6–0.8 m | 30.4 °C | Turbinaria decurrens and Halimeda sp. | 36.0 | [43,44] |
| G. belizeanus | Abu Shosha, Saudi Arabia | Red Sea | 22.3505° N, 39.2811° E | 0.5 m | 29.6 °C | Turbinaria decurrens and Halimeda sp. | 38.3 | [43,44] |
| G. belizeanus | Um Al Balam, Saudi Arabia | Red Sea | 22.4277° N, 38.9652° E | 0.4–0.6 m | 29.3 °C | Turbinaria decurrens and Halimeda sp. | 36.1 | [43,44] |
| G. belizeanus | Um Al Kiethl, Saudi Arabia | Red Sea, | 22.3197° N, 39.0541° E | 0.5 m | 29.5 °C | Turbinaria decurrens and Halimeda sp. | 28.9 | [43,44] |
| G. belizeanus | Coast Guard Reef, Saudi Arabia | Red Sea | 20.3950° N, 40.3580° E | 0.5 m | NA | Turbinaria decurrens and Halimeda sp. | NA | [43,44] |
| G. belizeanus | Mangrove Reef, Saudi Arabia | Red Sea | 20.4400° N, 40.2911° E | 0.4 m | NA | Turbinaria decurrens | NA | [43,44] |
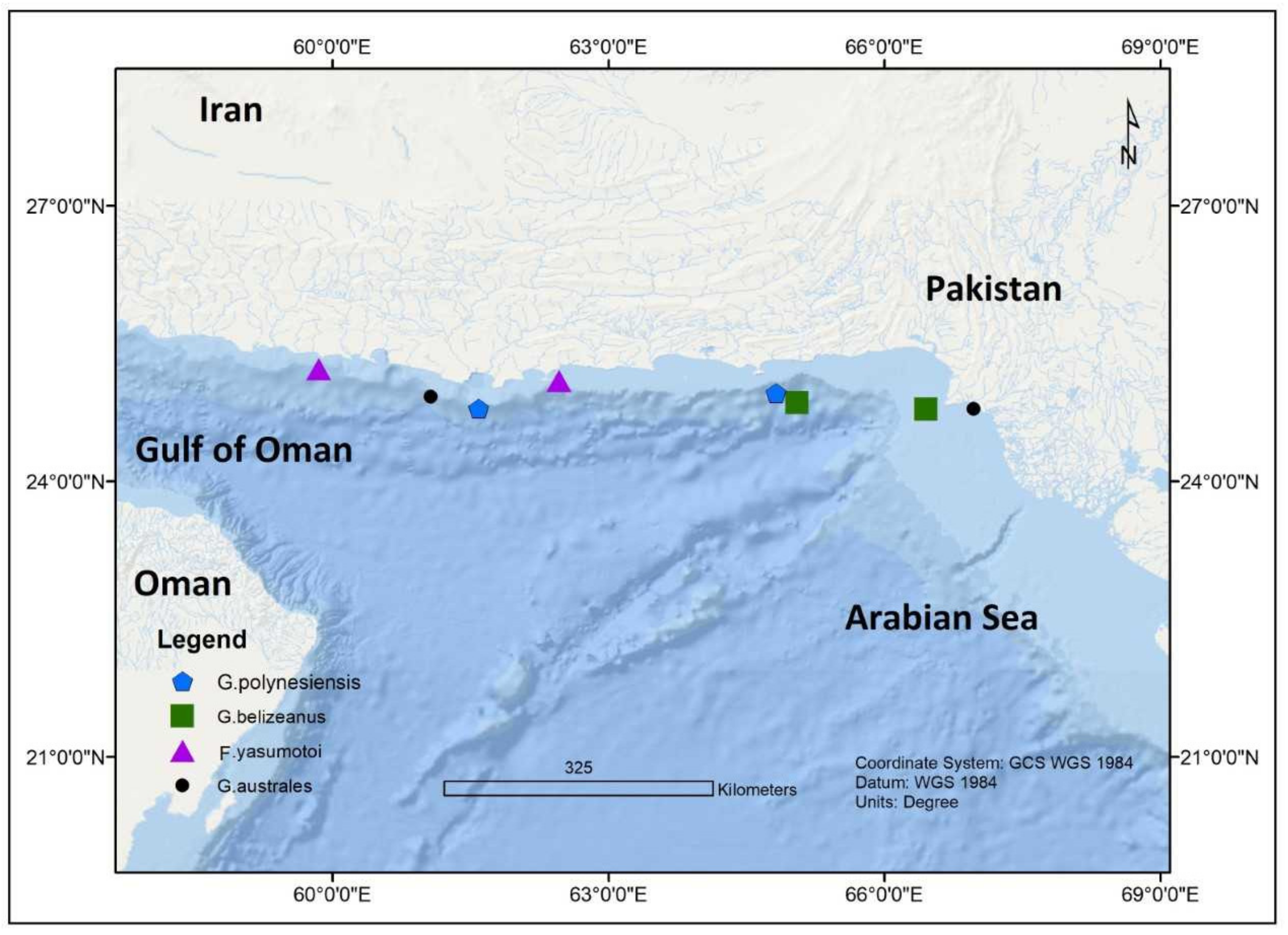
| G. toxicus | Sindh Coast, Pakistan | Manora Channel; Chuma Island | 24.8091° N, 66.9908° E | 1.0 m | 32 °C | [69] | ||
| G. toxicus | Balochistan Coast, Pakistan | Manora Channel; Chuma Island | 24.7861° N, 66.4516° E | 1.0 m | 28 °C | [69] | ||
| G. belizeanus | Sindh Coast, Pakistan | Manora Channel; Chuma Island | 24.8091° N, 66.9908° E | 1.0 m | 32 °C | [69] | ||
| G. belizeanus | Balochistan Coast | Manora Channel; Chuma Island, Pakistan | 24.7861° N, 66.4516° E | 1.0 m | 28 °C | [69] | ||
| G. australes | Sindh Coast | Manora Channel; Chuma Island, Pakistan | 24.8091° N, 66.9908° E | 1.0 m | 32 °C | [69] | ||
| G. australes | Balochistan Coast | Manora Channel; Chuma Island, Pakistan | 24.7861° N, 66.4516° E | 1.0 m | 28 °C | [69] | ||
| G. polynesiensis | Sindh Coast | Manora Channel; Chuma Island, Pakistan | 24.8091° N, 66.9908° E | 1.0 m | 32 °C | [69] | ||
| G. polynesiensis | Balochistan Coast | Manora Channel; Chuma Island, Pakistan | 24.7861° N, 66.4516° E | 1.0 m | 28 °C | [69] | ||
| F. yasumotoi | Sindh Coast | Manora Channel; Chuma Island, Pakistan | 24.8091° N, 66.9908° E | 1.0 m | 32 °C | [69] | ||
| F. yasumotoi | Balochistan Coast | Manora Channel; Chuma Island, Pakistan | 24.7861° N, 66.4516° E | 1.0 m | 28 °C | [69] | ||
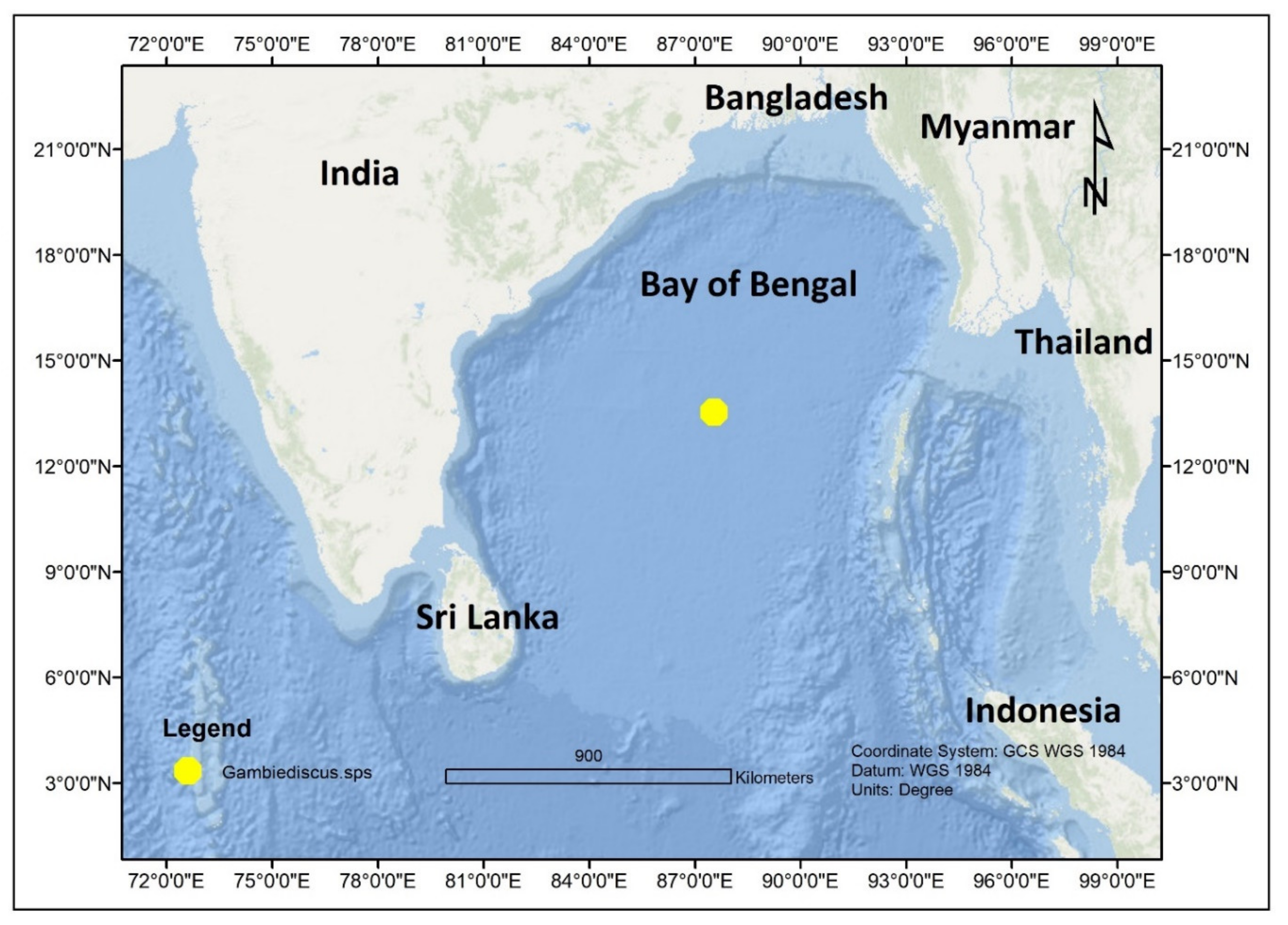
| Gambierdiscus sp. | Bay of Bengal | India | [71] | |||||
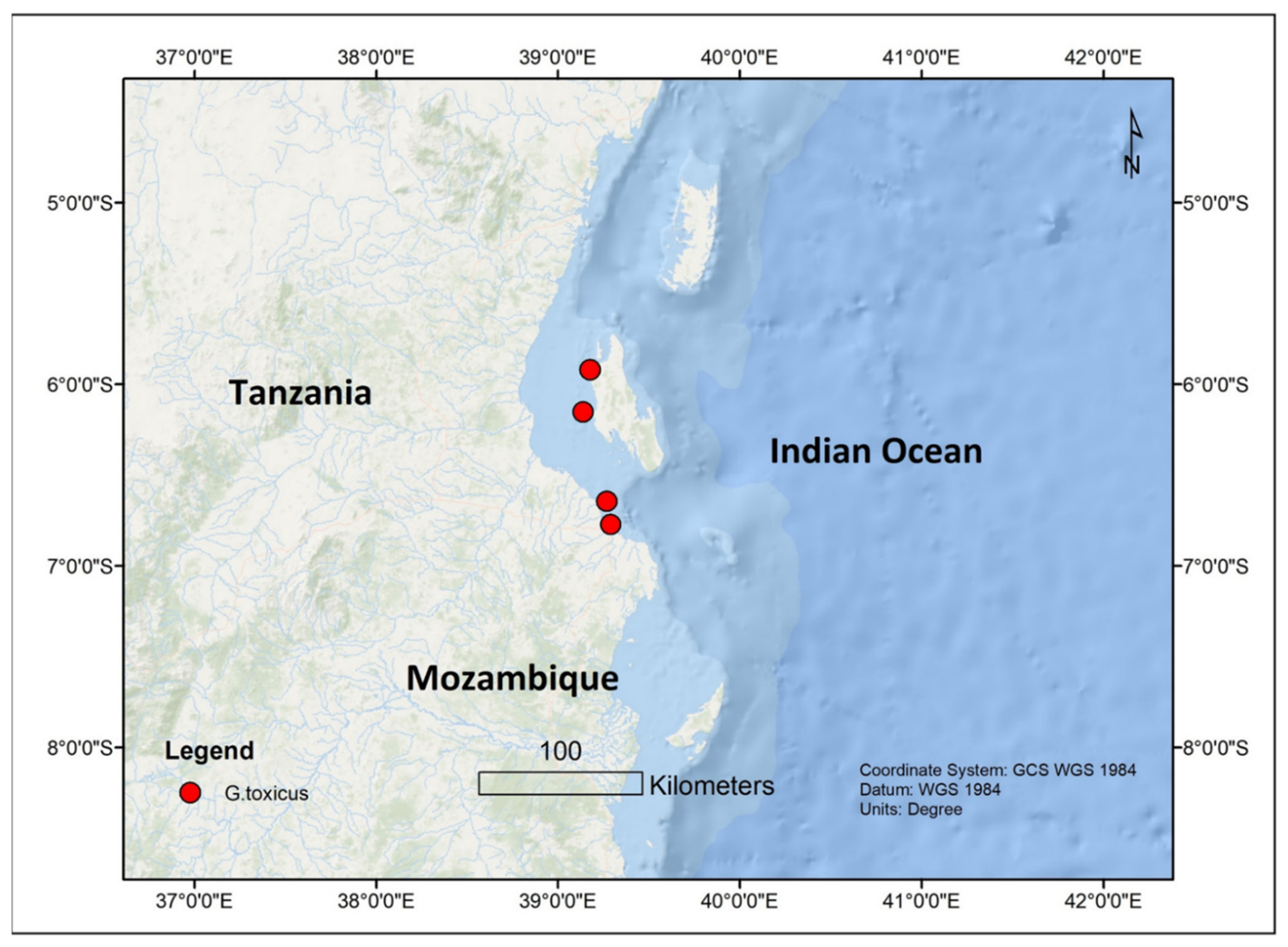
| G. toxicus | Mbudya Island, Dar es Salaam | Tanzania | 5.0 m | 25.5–31.0 °C | Seagrass—Thalassodendron ciliatum | 35–37 | [66] | |
| G. toxicus | Bawe Island, Zanzibar | Tanzania | 10.0 m | 26.0–29.0 °C | Unknown brown algae mixed with Filamentous cyanobacteria | 34–36 | [66] | |
| G. toxicus | Makoba, Zanzibar | Tanzania | 10.0 m | 26.0 °C | Gracilaria sp. | 35–43 | [66] | |
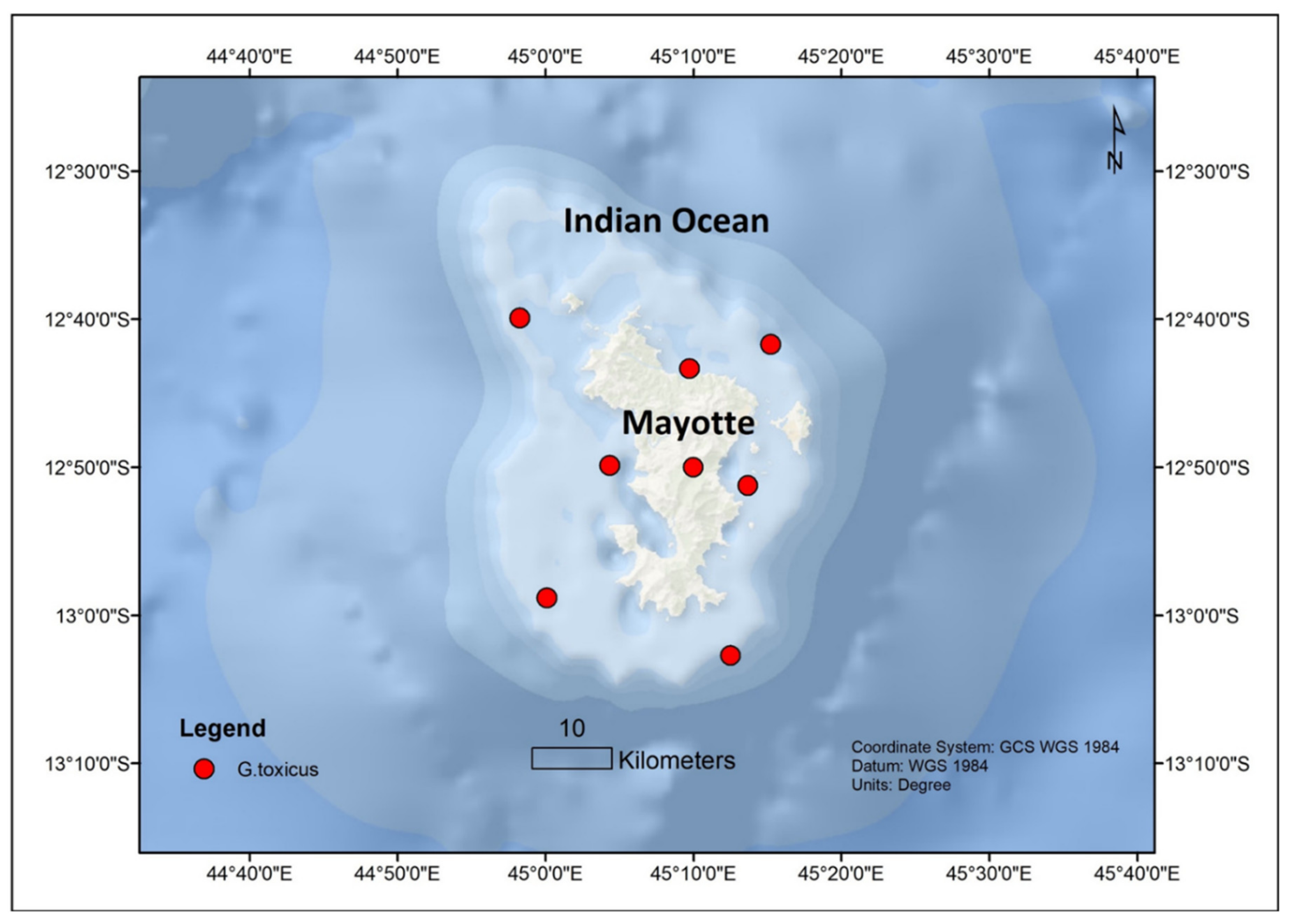
| G. toxicus | Longoni Bay (Stations 1 and 2) | Mayotte, Mozambique | 12.8333° N, 45.1666° E | Between the depth of the lower intertidal level and 30 m | 26–31 °C | Ostreopsis lenticularis; O. lenticularis; Prorocentrum lima and 4 new species of Prorocentrum | 35–37 | [60] |
| G. toxicus | Surprise Reef (Station 3) | Mayotte, Mozambique | 12.8333° N, 45.1666° E | [60] | ||||
| G. toxicus | Prevoyante Reef (Station 5) | Mayotte, Mozambique | 12.8333° N, 45.1666° E | [60] | ||||
| G. toxicus | Great Barrier Reef (Station 4A–C) | Mayotte, Mozambique | 12.8333° N, 45.1666° E | [60] | ||||
| G. toxicus | Cape Andrema Station 6) | Mayotte, Mozambique | 12.8333° N, 45.1666° E | [60] | ||||
| G. toxicus | M’Songoma (Station 7) | Mayotte, Mozambique | 12.8333° N, 45.1666° E | [60] | ||||
| G. toxicus | M’Sanga Tsohole Reef (Station 8A,B) | Mayotte, Mozambique | 12.8333° N, 45.1666° E | [60] | ||||
| G. toxicus | Bambo Reef (Station 9A,B) | Mayotte, Mozambique | 12.8333° N, 45.1666° E | [60] | ||||
| G. toxicus | Mayotte | South West Indian Ocean | Prorocentrum spp., Ostreopsis sp., Amphidinium spp | [59] | ||||
| G. toxicus | Mayotte | South West Indian Ocean | [86] | |||||
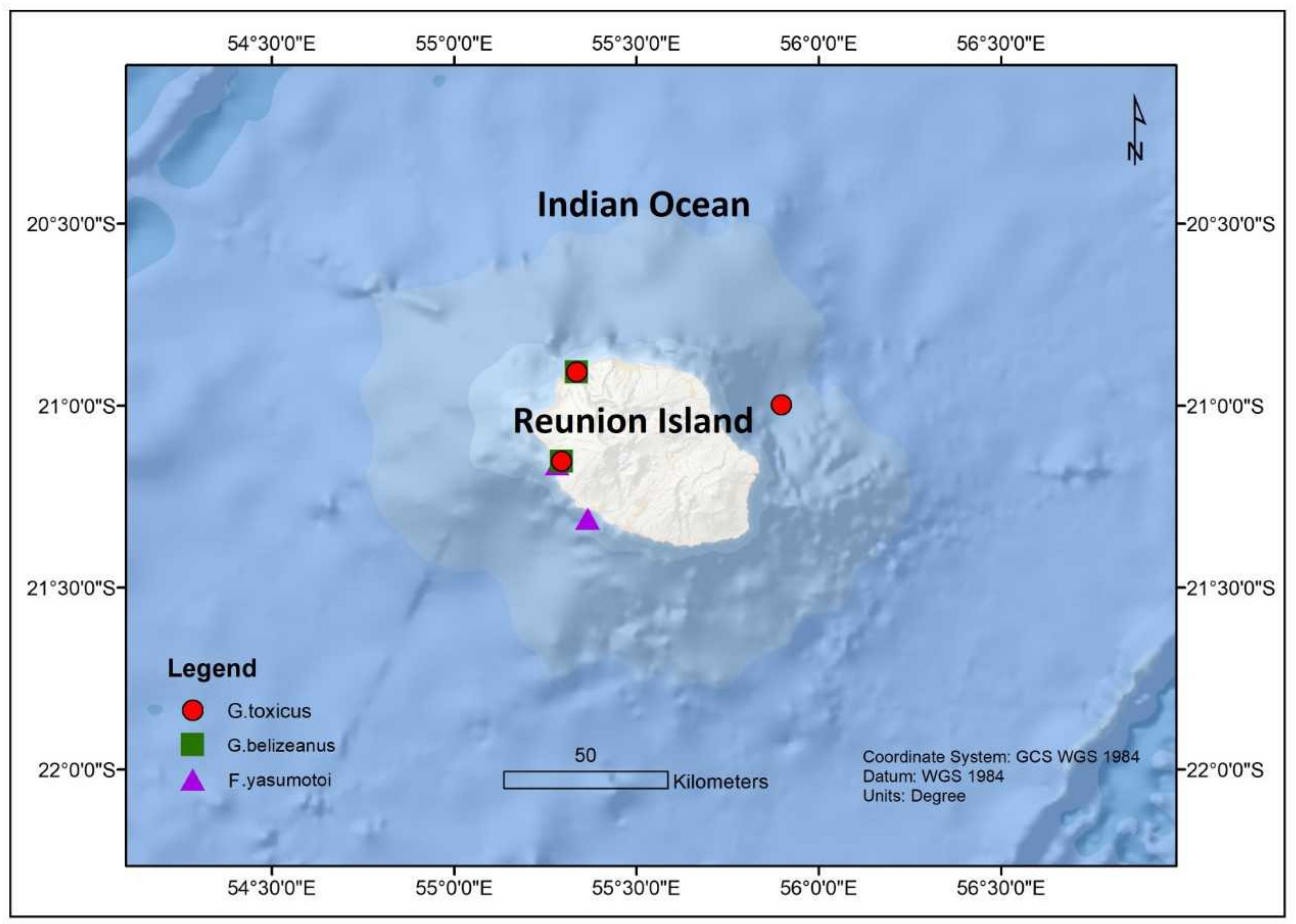
| G. belizeanus | St-Leu and Saline reef | Reunion Island | 21.1533° N, 55.2944° E | 1–5 m | P. arenarium; P. belizeanum; P. concavum; P. elegans; P. emarginatum; P. hoffmannianum; P. lima; P. mexicanum; P sculptile; P. borbonicum; O. heptagona; O. labens; O. lenticularis; O. mascarensis; O. ovata; O. siamensis; C. monotis; C. tropicalis; Sinophysis microcephalis; S. canaliculate; Amphidiniopsis sp.; Bysmatrum sps. | [65] | ||
| G. belizeanus | Possession Bay | La Reunion | 20.9347° N, 55.3491° E | 10–12 m | P. arenarium; P. emarginatum; P. concavum; P. lima; P panamensis; Coolia sps. | [65] | ||
| G. toxicus | St-Leu and Saline reef | La Reunion | 21.1533° N, 55.2944° E | 1–5 m | P. arenarium; P. belizeanum; P. concavum; P. elegans; P. emarginatum; P. hoffmannianum; P. lima; P. mexicanum; P sculptile; P. borbonicum; O. heptagona; O. labens; O. lenticularis; O. mascarensis; O. ovata; O. siamensis; C. monotis; C. tropicalis; Sinophysis microcephalis; S. canaliculate; Amphidiniopsis sp.; Bysmatrum sps. | [65] | ||
| G. toxicus | Possession Bay | La Reunion | 20.9347° N, 55.3491° E | 10–12 m | P. arenarium; P. emarginatum; P. concavum; P. lima; P panamensis; Coolia sp. | [65] | ||
| F. yasumotoi | St-Leu and Saline reef | La Reunion | 21.1533° N, 55.2944° E | 1–5 m | P. arenarium; P. belizeanum; P. concavum; P. elegans; P. emarginatum; P. hoffmannianum; P. lima; P. mexicanum; P sculptile; P. borbonicum; O. heptagona; O. labens; O. lenticularis; O. mascarensis; O. ovata; O. siamensis; C. monotis; C. tropicalis; Sinophysis microcephalis; S. canaliculate; Amphidiniopsis sp.; Bysmatrum sp. | [65] | ||
| F. yasumotoi | Possession Bay | La Reunioin | 20.9347° N, 55.3491° E | 10–12 m | P. arenarium; P. emarginatum; P. concavum; P. lima; P panamensis; Coolia sp. | [65] | ||
| G. toxicus | Reunioin Islands | Mauritius | Prorocentrum sp.; Ostreopsis sp. | [64] | ||||
| Gambierdiscus | Mangalore | India | 12.91777° N, 74.79162° E | [72,73,74] |
| F. yasumotoi [12] | G. belizeanus [92] | G. toxicus [93] | G. polynesiensis [94] | G. australes [94] | |
|---|---|---|---|---|---|
| Cell depth | 54 to 73 µm | 59 to 73 µm | 71 to 96 µm | 65 to 87 µm | 60 to 82 µm |
| Cell width | 46 to 61 µm | 60 to 73 µm | 45 to 140 µm | 64 to 75 µm | 65 to 85 µm |
| Cell length | 49 to 70 µm | 47 to 51 µm | 50 to 150 µm | 57 to 75 µm | 76 to 93 µm |
| Cell shape | Globular | Round | Rounded/ellipsoid | Rounded/ellipsoid | Rounded/ellipsoid |
| Apical pore | Fish hook-shaped | Fish hook-shaped | Fish hook shaped | Fish hook shaped | Fish hook shaped |
| Thecal surface | Moderately aerated | Smooth | Smooth | ||
| Apical plate | elongated and teardrop-shaped | Ellipsoid | Ellipsoid | large triangular apical pore plate | broad ellipsoid apical pore plate |
| Hypotheca | Truncate-conical | Eight hypothecal plates | |||
| Epitheca | dome-shaped | Ellipsoid | Eleven plates | ||
| Sulcus | Short | ||||
| Chloroplast | elongated golden-brown chloroplasts were peripherally arranged | golden-brown drop-shaped chloroplasts radiating from the cell centre | |||
| Nucleus | large, oblong and located in the right posterior | large, oblong and crescent-shaped, and located in the right posterior cell part | Posteriorly situated nucleus | ||
| Cingulum | deeply excavated, descending, | Ascending narrow cingulum | Narrow and deep | ||
| Plate Formulae | Po, 3′, 7″, 6C, 8S, 5‴, 1p, 2⁗ | Po, 3′, 7″, 6C, 8S, 5‴, 1p et 2‴ | Po, 3′, 7″, 6C, 8S, 5‴, 1p et 2⁗ | ||
| Studied by | [42,69] | [42,43,44,69] | [66,69] | [69] | [69] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Habibi, N.; Uddin, S.; Bottein, M.-Y.D.; Faizuddin, M. Ciguatera in the Indian Ocean with Special Insights on the Arabian Sea and Adjacent Gulf and Seas: A Review. Toxins 2021, 13, 525. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13080525
Habibi N, Uddin S, Bottein M-YD, Faizuddin M. Ciguatera in the Indian Ocean with Special Insights on the Arabian Sea and Adjacent Gulf and Seas: A Review. Toxins. 2021; 13(8):525. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13080525
Chicago/Turabian StyleHabibi, Nazima, Saif Uddin, Marie-Yasmine Dechraoui Bottein, and Mohd Faizuddin. 2021. "Ciguatera in the Indian Ocean with Special Insights on the Arabian Sea and Adjacent Gulf and Seas: A Review" Toxins 13, no. 8: 525. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13080525