Olfactory Bulb D2/D3 Receptor Availability after Intrastriatal Botulinum Neurotoxin-A Injection in a Unilateral 6-OHDA Rat Model of Parkinson’s Disease

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
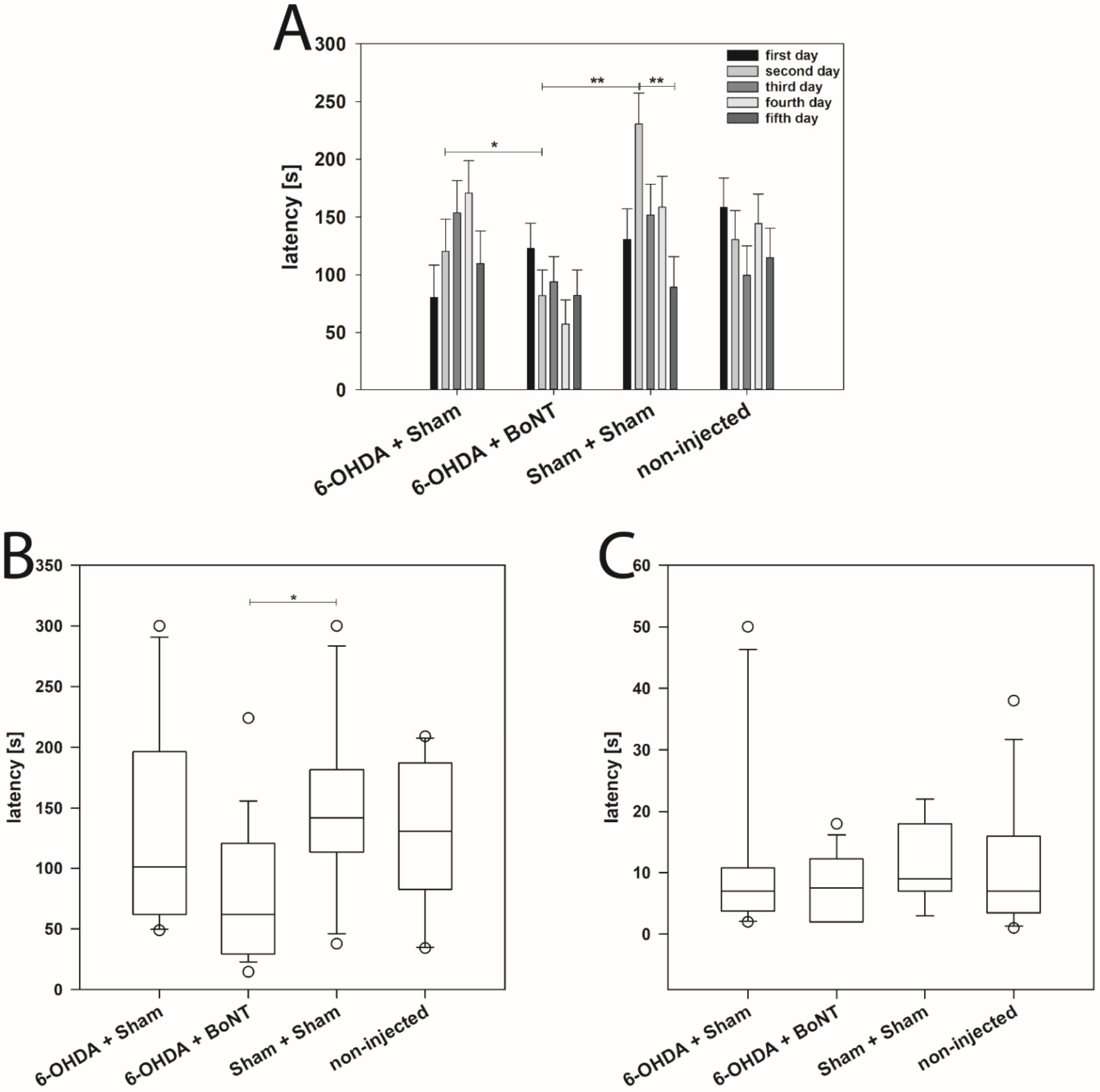
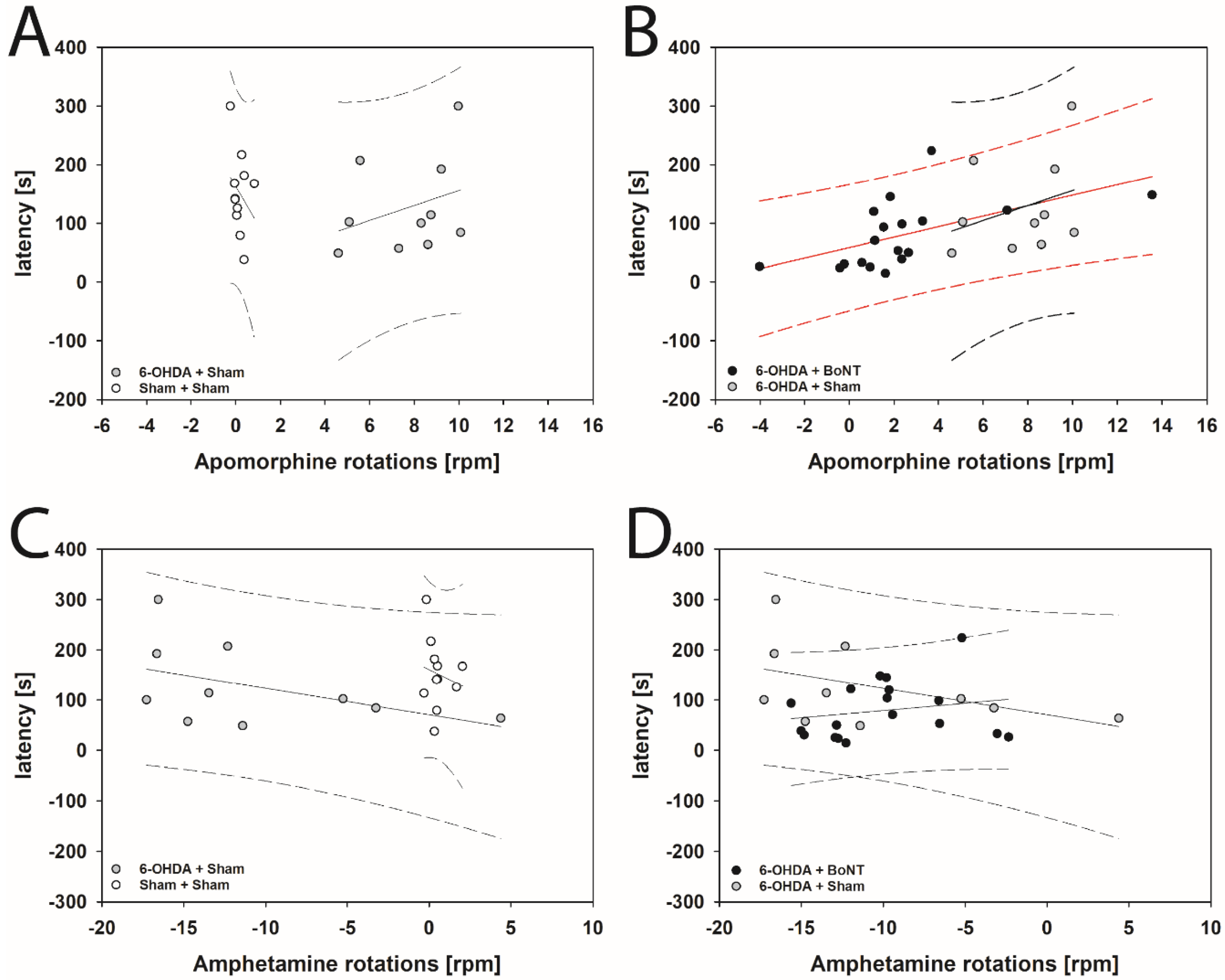
2.1. Drug-Induced Behavior Tests
2.1.1. Apomorphine-Induced Rotation Test
2.1.2. Amphetamine-Induced Rotation Test
2.2. Hemi-PD Rats Do Not Show Olfactory Deficits, but BoNT-A-Injected Hemi-PD Rats Improve Performance in the Buried Pellet Test
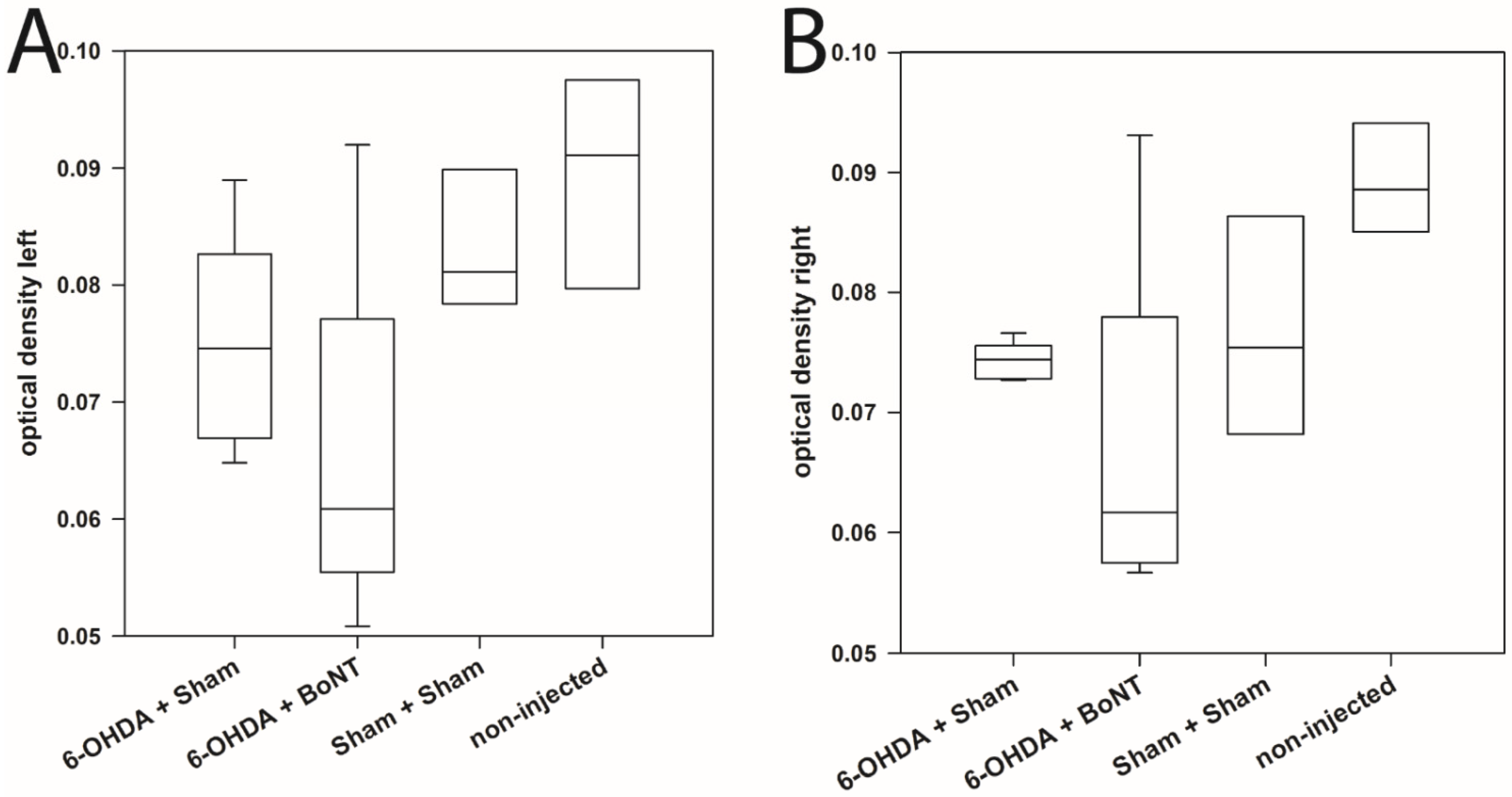
2.3. There Is No Densitometric Interhemispheric Difference of TH-Immunoreactivity in the OB
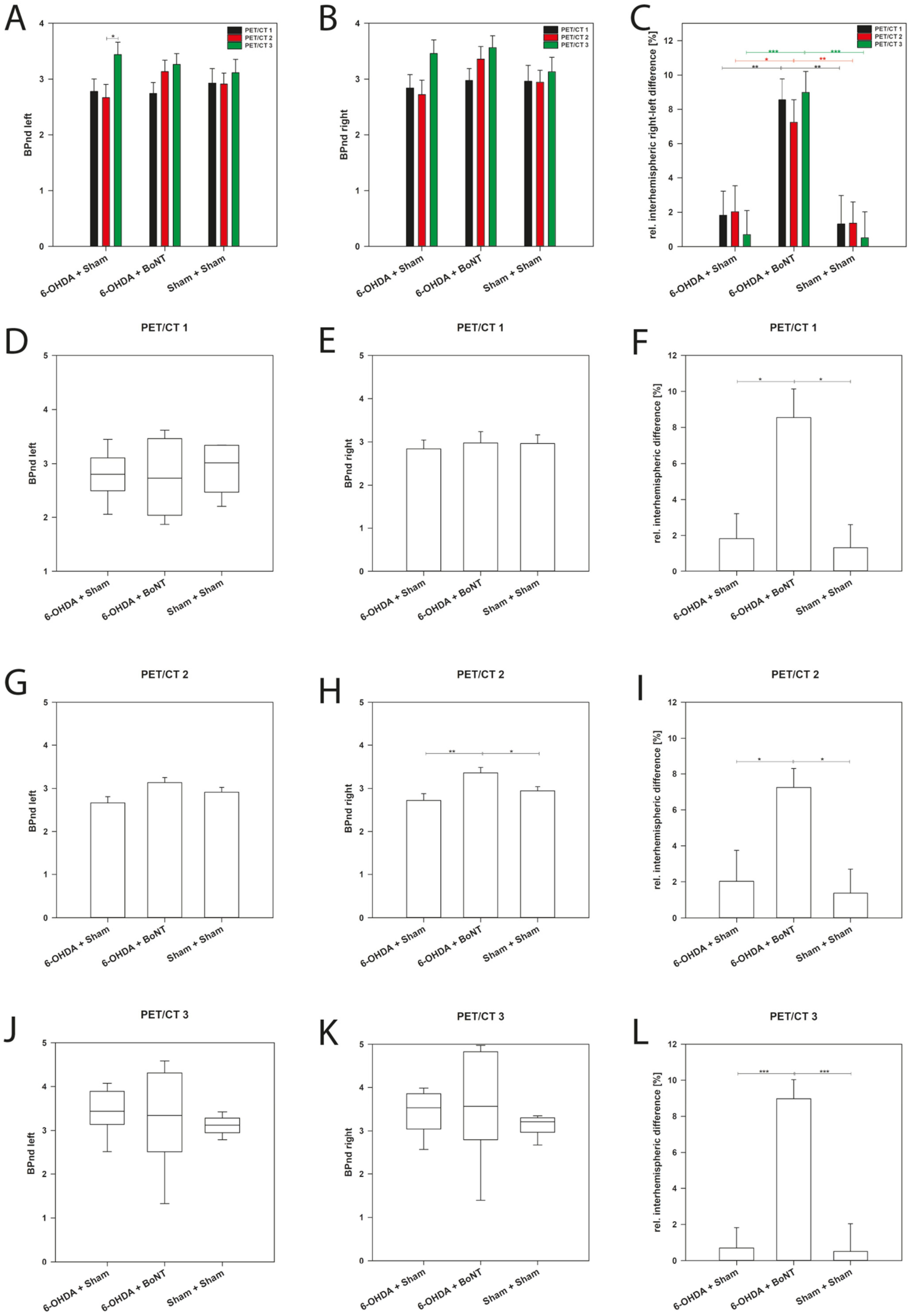
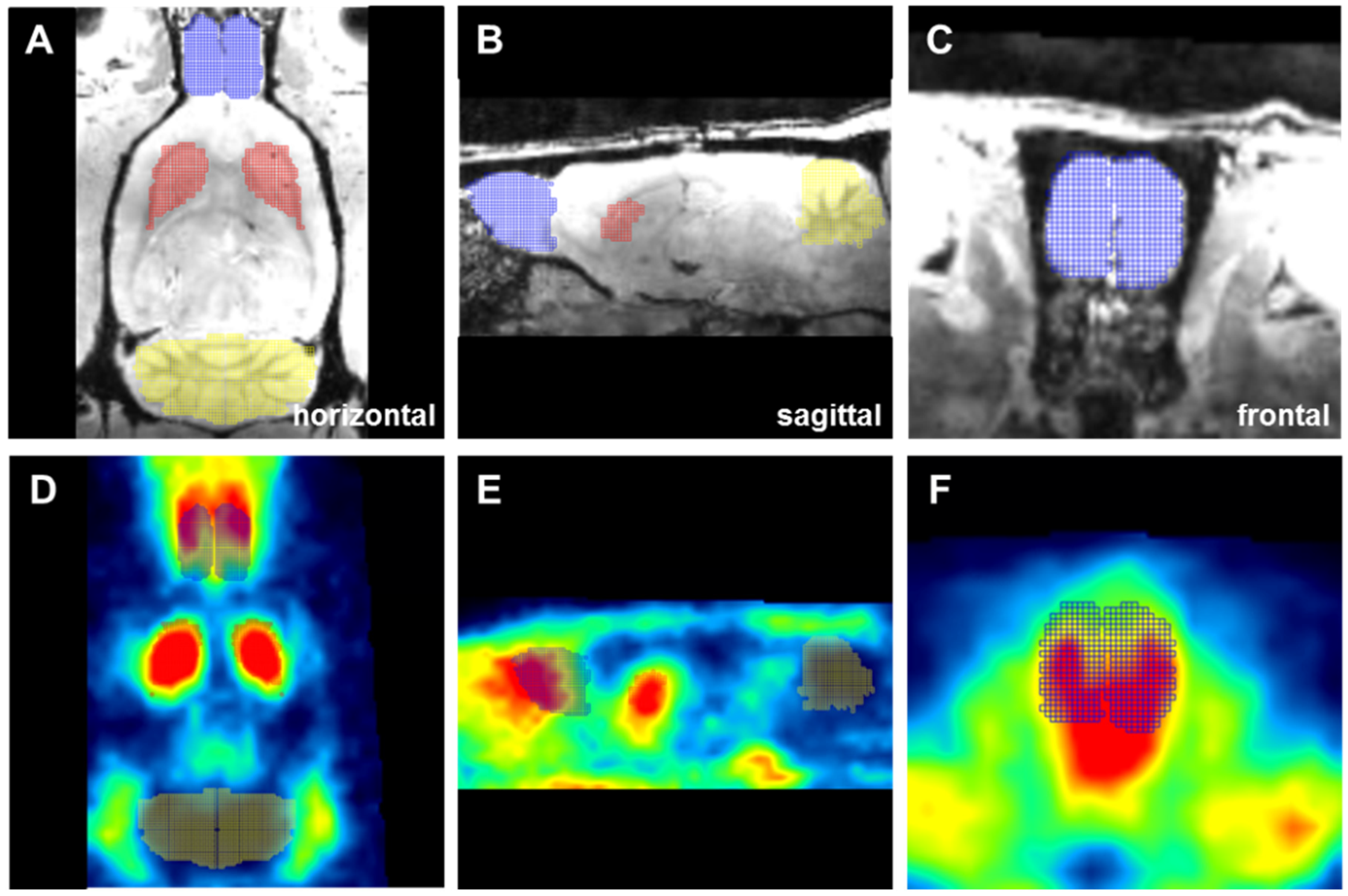
2.4. Olfactory Bulb D2/D3R Availability—Receptor Upregulation after Ipsilateral Intrastriatal BoNT-A Injection
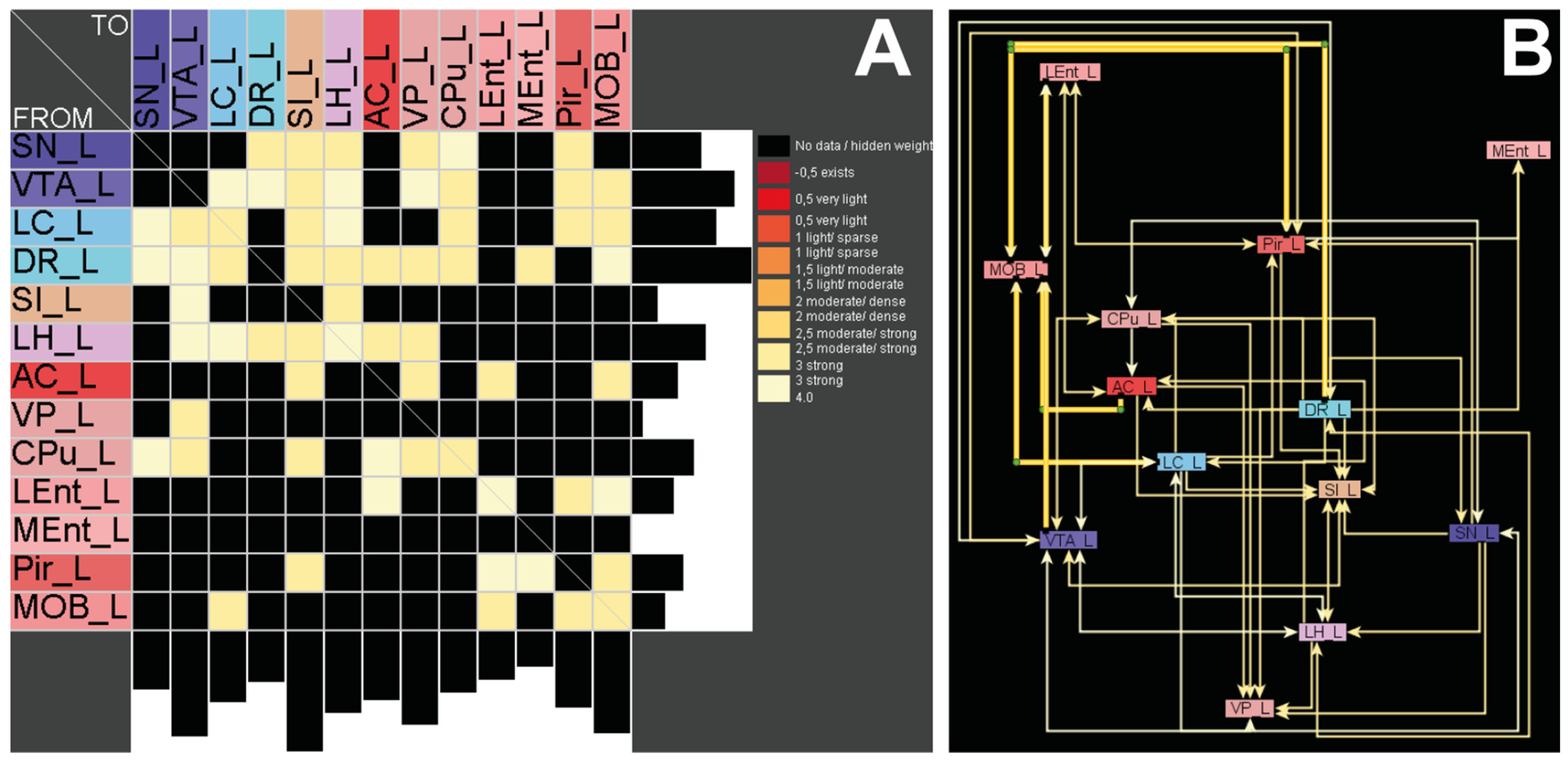
2.5. Connectomics—There Are No Direct Pathways between the Main Olfactory Bulb and Basal Ganglia
2.6. Correlating Buried Pellet Test, Apomorphine- and Amphetamine-Induced Rotations, and Optical Densites of Glomerular Layers of the Left and Right OB
3. Discussion
3.1. Olfactory Deficits in PD
3.2. Hemi-PD Rats Show No Olfactory Deficits in the Buried Pellet Test
3.3. Dopamine as a Key Transmitter in Processing of Olfactory Information
3.3.1. Hemi-PD Rats Do Not Show Altered TH-Immunoreactivity in the Glomerular Layer of the OB
3.3.2. Intrastriatal BoNT-A Injection Led to an Upregulation of D2/D3 Receptor Availability in the Ipsilateral OB of Hemi-PD Rats
3.4. Connectomics Reveal Indirect Projections between the OB and the CPu
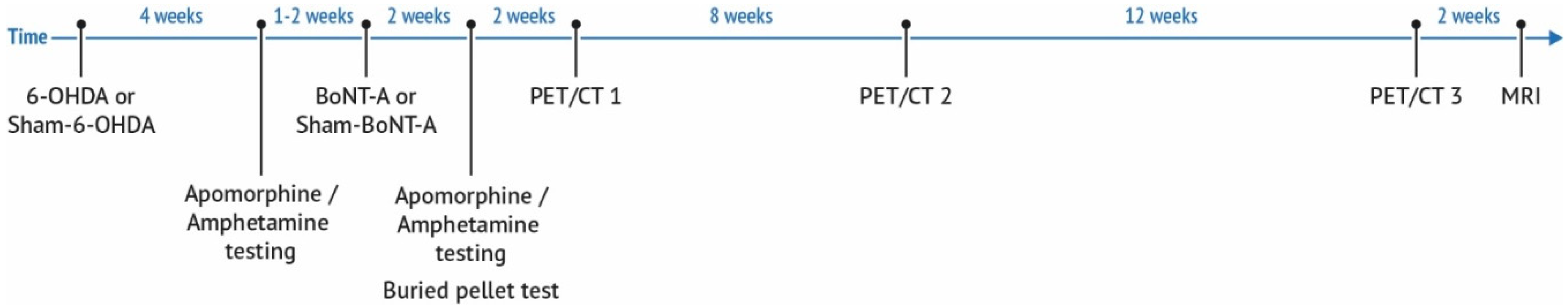
4. Materials and Methods
4.1. Animals
4.2. Induction of Hemiparkinsonism
4.3. Injection of BoNT-A into the Striatum
4.4. Behavioral Testing
4.4.1. Drug-Induced Rotation Tests (Apomorphine, Amphetamine)
Apomorphine-Induced Rotation Test
Amphetamine-Induced Rotational Test
4.4.2. Buried Pellet Test
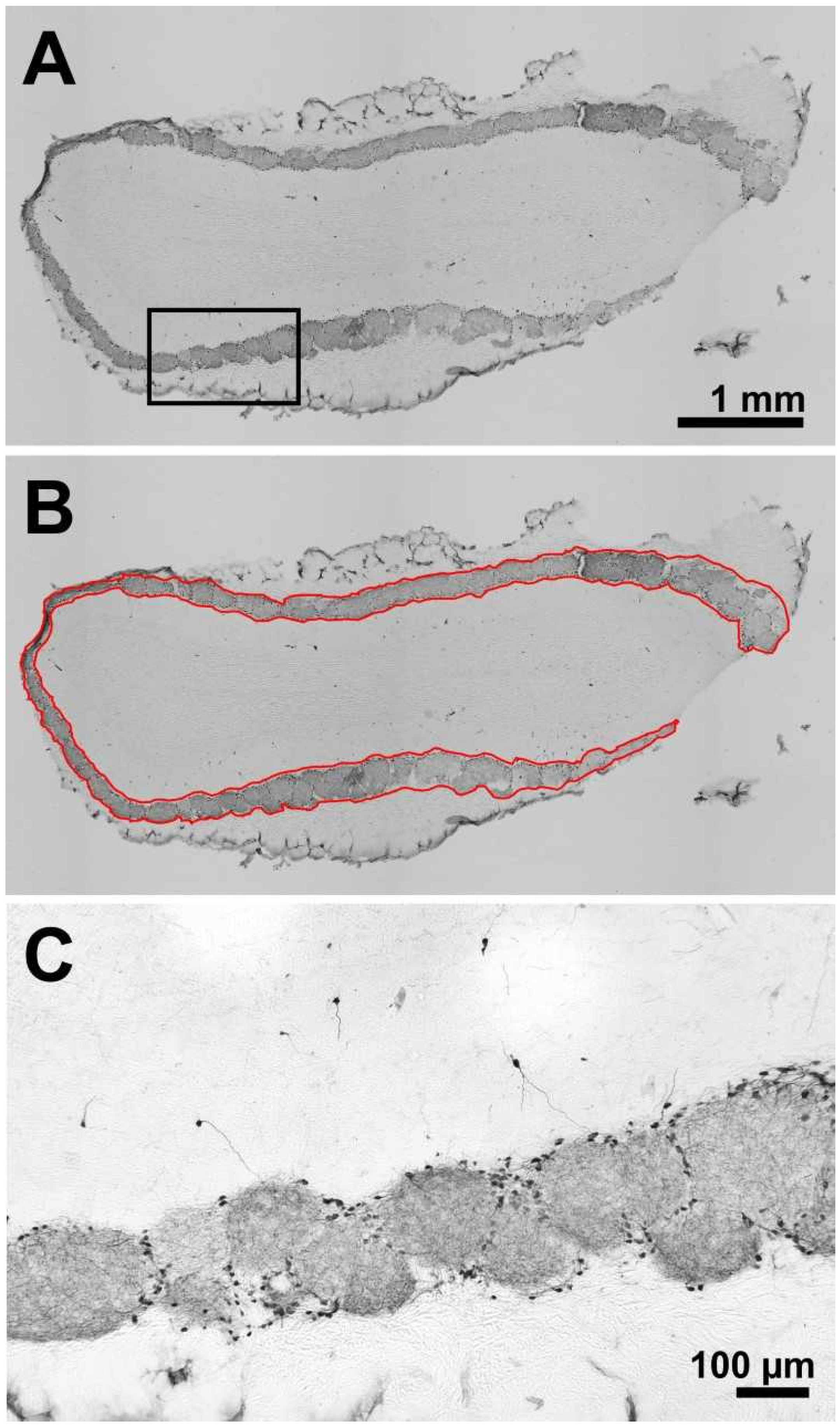
4.5. TH-Immunohistochemistry of the Olfactory Bulb
4.6. Densitometric Measurement
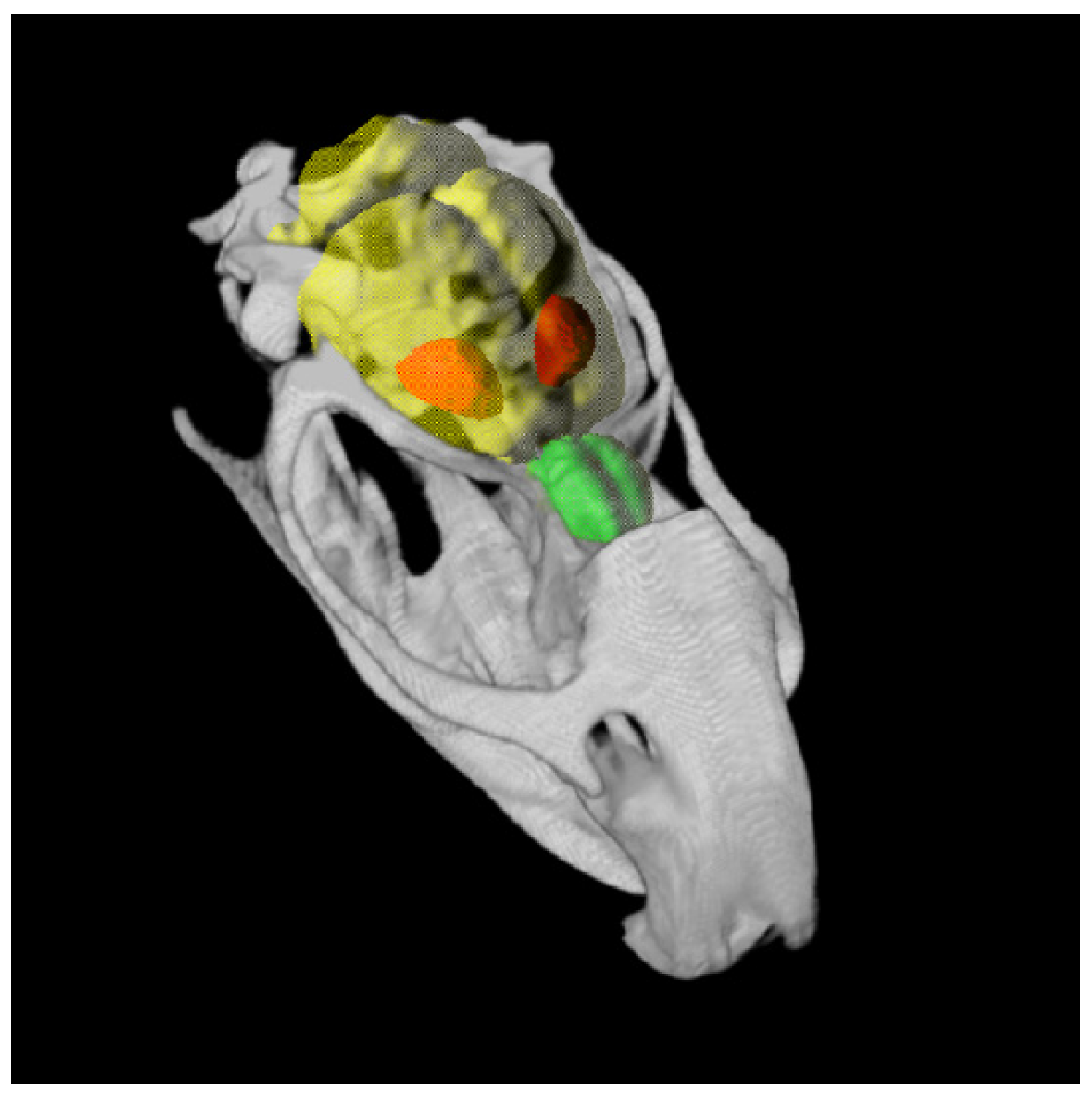
4.7. MRI
4.8. Atlas Generation and PET/CT Data Analysis
4.9. Tract-Tracing-Based Generation of Connectome Data
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BoNT-A | Botulinum neurotoxin-A |
| BPnd | non-displaceable Binding Potential |
| BSA | bovine serum albumin |
| CPu | striatum (caudate-putamen) |
| CT | computed tomography |
| GABA | gamma-aminobutyric acid |
| hemi-PD | hemiparkinsonian |
| D2/D3R | D2/D3 receptor |
| MFB | medial forebrain bundle |
| MOB | main olfactory bulb |
| MRI | magnetic resonance imaging |
| OB | olfactory bulb |
| OBG | olfactory-basal ganglia-connectivity |
| 6-OHDA | 6-hydroxydopamine |
| PBS | phosphate-buffered saline |
| PD | Parkinson’s disease |
| PET | Positron emission tomography |
| PVE | partial volume effect |
| SNpc | substantia nigra pars compacta |
| SRTM2 | Simplified Reference Tissue Model 2 |
| TAC | time-active hydroxylase |
| TH | tyrosine hydroxylase |
| VOI | voxels of interest |
| VTA | ventral tegmental area |
References
- Eskow Jaunarajs, K.L.; Dupre, K.B.; Ostock, C.Y.; Button, T.; Deak, T.; Bishop, C. Behavioral and neurochemical effects of chronic L-DOPA treatment on nonmotor sequelae in the hemiparkinsonian rat. Behav. Pharmacol. 2010, 21, 627–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eskow Jaunarajs, K.L.; Angoa-Perez, M.; Kuhn, D.M.; Bishop, C. Potential mechanisms underlying anxiety and depression in Parkinson’s disease: Consequences of l-DOPA treatment. Neurosci. Biobehav. Rev. 2011, 35, 556–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jungnickel, J.; Kalve, I.; Reimers, L.; Nobre, A.; Wesemann, M.; Ratzka, A.; Halfer, N.; Lindemann, C.; Schwabe, K.; Töllner, K.; et al. Topology of intrastriatal dopaminergic grafts determines functional and emotional outcome in neurotoxin-lesioned rats. Behav. Brain Res. 2011, 216, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Tibar, H.; El Bayad, K.; Bouhouche, A.; Ben Haddou, E.H.A.; Benomar, A.; Yahyaoui, M.; Benazzouz, A.; Regragui, W. Non-motor symptoms of Parkinson’s Disease and their impact on quality of life in a cohort of Moroccan patients. Front. Neurol. 2018, 9, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Politis, M.; Wu, K.; Molloy, S.; Bain, P.G.; Chaudhuri, K.R.; Piccini, P. Parkinson’s disease symptoms: The patient’s perspective. Mov. Disord. 2010, 25, 1646–1651. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, C.H.; Shephard, B.C. Olfactory evoked responses and identification tests in neurological disease. Ann. N. Y. Acad. Sci. 1998, 855, 608–615. [Google Scholar] [CrossRef]
- Haehner, A.; Boesveldt, S.; Berendse, H.W.; Mackay-Sim, A.; Fleischmann, J.; Silburn, P.A.; Johnston, A.N.; Mellick, G.D.; Herting, B.; Reichmann, H.; et al. Prevalence of smell loss in Parkinson’s disease—A multicenter study. Park. Relat. Disord. 2009, 15, 490–494. [Google Scholar] [CrossRef]
- Braak, H.; Ghebremedhin, E.; Rüb, U.; Bratzke, H.; Del Tredici, K. Stages in the development of Parkinson’s disease-related pathology. Cell Tissue Res. 2004, 318, 121–134. [Google Scholar] [CrossRef]
- Haehner, A.; Masala, C.; Walter, S.; Reichmann, H.; Hummel, T. Incidence of Parkinson’s disease in a large patient cohort with idiopathic smell and taste loss. J. Neurol. 2019, 266, 339–345. [Google Scholar] [CrossRef]
- Reichmann, H. Clinical criteria for the diagnosis of Parkinson’s disease. Neurodegener. Dis. 2010, 7, 284–290. [Google Scholar] [CrossRef] [Green Version]
- Haehner, A.; Hummel, T.; Reichmann, H. Olfactory loss in parkinson’s disease. Parkinsons. Dis. 2011, 2011, 450939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hüttenbrink, K.B.; Hummel, T.; Berg, D.; Gasser, T.; Hähner, A. Riechstörungen: Häufig im alter und wichtiges frühsymptom neurodegenerativer erkrankungen. Dtsch. Arztebl. Int. 2013, 110, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berendse, H.W.; Ponsen, M.M. Detection of preclinical Parkinson’s disease along the olfactory trac(t). J. Neural Transm. Suppl. 2006, 2006, 321–325. [Google Scholar] [CrossRef]
- Haehner, A.; Tosch, C.; Wolz, M.; Klingelhoefer, L.; Fauser, M.; Storch, A.; Reichmann, H.; Hummel, T. Olfactory Training in Patients with Parkinson’s Disease. PLoS ONE 2013, 8, e61680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haehner, A.; Hummel, T.; Reichmann, H. A clinical approach towards smell loss in Parkinson’s disease. J. Parkinsons. Dis. 2014, 4, 189–195. [Google Scholar] [CrossRef]
- Doty, R.L.; Deems, D.A.; Stellar, S. Olfactory dysfunction in parkinsonism: A general deficit unrelated to neurologic signs, disease stage, or disease duration. Neurology 1988, 38, 1237–1244. [Google Scholar] [CrossRef] [Green Version]
- Alexander, G.E. Biology of Parkinson’s disease: Pathogenesis and pathophysiology of a multisystem neurodegenerative disorder. Dialogues Clin. Neurosci. 2004, 6, 259–280. [Google Scholar] [CrossRef]
- Emre, M. What causes mental dysfunction in Parkinson’s disease? Mov. Disord. 2003, 18, 63–71. [Google Scholar] [CrossRef]
- Melis, M.; Haehner, A.; Mastinu, M.; Hummel, T.; Barbarossa, I.T. Molecular and genetic factors involved in olfactory and gustatory deficits and associations with microbiota in parkinson’s disease. Int. J. Mol. Sci. 2021, 22, 4286. [Google Scholar] [CrossRef]
- Jankovic, J.; Tan, E.K. Parkinson’s disease: Etiopathogenesis and treatment. J. Neurol. Neurosurg. Psychiatry 2020, 91, 795–808. [Google Scholar] [CrossRef]
- Doty, R.L. Olfaction in Parkinson’s disease and related disorders. Neurobiol. Dis. 2012, 46, 527–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rietdijk, C.D.; Perez-Pardo, P.; Garssen, J.; van Wezel, R.J.A.; Kraneveld, A.D. Exploring Braak’s hypothesis of parkinson’s disease. Front. Neurol. 2017, 8, 37. [Google Scholar] [CrossRef] [PubMed]
- Lerner, A.; Bagic, A. Olfactory pathogenesis of idiopathic parkinson disease revisited. Mov. Disord. 2008, 23, 1076–1084. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Del Tredici, K.; Rüb, U.; De Vos, R.A.I.; Jansen Steur, E.N.H.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Braak, H.; Rüb, U.; Gai, W.P.; Del Tredici, K. Idiopathic Parkinson’s disease: Possible routes by which vulnerable neuronal types may be subject to neuroinvasion by an unknown pathogen. J. Neural Transm. 2003, 110, 517–536. [Google Scholar] [CrossRef]
- Pearce, R.K.B.; Hawkes, C.H.; Daniel, S.E. The anterior olfactory nucleus in Parkinson’s disease. Mov. Disord. 1995, 10, 283–287. [Google Scholar] [CrossRef]
- Giguère, N.; Nanni, S.B.; Trudeau, L.-E. On Cell Loss and Selective Vulnerability of Neuronal Populations in Parkinson’s Disease. Front. Neurol. 2018, 9, 455. [Google Scholar] [CrossRef]
- Atasoy, H.T.; Nuyan, O.; Tunc, T.; Yorubulut, M.; Unal, A.E.; Inan, L.E. T2-weighted MRI in Parkinson’s disease; substantia nigra pars compacta hypointensity correlates with the clinical scores. Neurol. India 2004, 52, 332–337. [Google Scholar]
- Drui, G.; Carnicella, S.; Carcenac, C.; Favier, M.; Bertrand, A.; Boulet, S.; Savasta, M. Loss of dopaminergic nigrostriatal neurons accounts for the motivational and affective deficits in Parkinson’s disease. Mol. Psychiatry 2014, 19, 358–367. [Google Scholar] [CrossRef] [Green Version]
- Sonne, J.; Beato, M.R. Neuroanatomy, Substantia Nigra; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Duty, S.; Jenner, P. Animal models of Parkinson’s disease: A source of novel treatments and clues to the cause of the disease. Br. J. Pharmacol. 2011, 164, 1357–1391. [Google Scholar] [CrossRef] [Green Version]
- Tieu, K. A guide to neurotoxic animal models of Parkinson’s disease. Cold Spring Harb. Perspect. Med. 2011, 1, a009316. [Google Scholar] [CrossRef] [PubMed]
- Stayte, S.; Vissel, B. Advances in non-dopaminergic treatments for Parkinson’s disease. Front. Neurosci. 2014, 8, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stott, S.R.W.; Barker, R.A. Time course of dopamine neuron loss and glial response in the 6-OHDA striatal mouse model of Parkinson’s disease. Eur. J. Neurosci. 2014, 39, 1042–1056. [Google Scholar] [CrossRef] [PubMed]
- Lledo, P.M.; Merkle, F.T.; Alvarez-Buylla, A. Origin and function of olfactory bulb interneuron diversity. Trends Neurosci. 2008, 31, 392–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsia, A.Y.; Vincent, J.D.; Lledo, P.M. Dopamine depresses synaptic inputs into the olfactory bulb. J. Neurophysiol. 1999, 82, 1082–1085. [Google Scholar] [CrossRef] [PubMed]
- Maher, B.J.; Westbrook, G.L. Co-transmission of dopamine and GABA in periglomerular cells. J. Neurophysiol. 2008, 99, 1559–1564. [Google Scholar] [CrossRef]
- Pignatelli, A.; Belluzzi, O. Dopaminergic neurones in the main olfactory bulb: An overview from an electrophysiological perspective. Front. Neuroanat. 2017, 11, 7. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Guzman, J.N.; Tkatch, T.; Chen, S.; Goldberg, J.A.; Ebert, P.J.; Levitt, P.; Wilson, C.J.; Hamm, H.E.; Surmeier, D.J. RGS4-dependent attenuation of M4 autoreceptor function in striatal cholinergic interneurons following dopamine depletion. Nat. Neurosci. 2006, 9, 832–842. [Google Scholar] [CrossRef]
- Oldenburg, I.A.; Ding, J.B. Cholinergic modulation of synaptic integration and dendritic excitability in the striatum. Curr. Opin. Neurobiol. 2011, 21, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Pisani, A.; Bernardi, G.; Ding, J.; Surmeier, D.J. Re-emergence of striatal cholinergic interneurons in movement disorders. Trends Neurosci. 2007, 30, 545–553. [Google Scholar] [CrossRef]
- Ztaou, S.; Maurice, N.; Camon, J.; Guiraudie-Capraz, G.; Kerkerian-Le Goff, L.; Beurrier, C.; Liberge, M.; Amalric, M. Involvement of Striatal Cholinergic Interneurons and M1 and M4 Muscarinic Receptors in Motor Symptoms of Parkinson’s Disease. J. Neurosci. 2016, 36, 9161–9172. [Google Scholar] [CrossRef] [PubMed]
- Antipova, V.; Hawlitschka, A.; Mix, E.; Schmitt, O.; Dräger, D.; Benecke, R.; Wree, A. Behavioral and structural effects of unilateral intrastriatal injections of botulinum neurotoxin a in the rat model of Parkinson’s disease. J. Neurosci. Res. 2013, 91, 838–847. [Google Scholar] [CrossRef] [PubMed]
- Antipova, V.; Holzmann, C.; Schmitt, O.; Wree, A.; Hawlitschka, A. Botulinum Neurotoxin A Injected Ipsilaterally or Contralaterally into the Striatum in the Rat 6-OHDA Model of Unilateral Parkinson’s Disease Differently Affects Behavior. Front. Behav. Neurosci. 2017, 11, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antipova, V.; Wree, A.; Holzmann, C.; Mann, T.; Palomero-Gallagher, N.; Zilles, K.; Schmitt, O.; Hawlitschka, A. Unilateral Botulinum Neurotoxin-A Injection into the Striatum of C57BL/6 Mice Leads to a Different Motor Behavior Compared with Rats. Toxins 2018, 10, 295. [Google Scholar] [CrossRef] [Green Version]
- Antipova, V.; Holzmann, C.; Hawlitschka, A.; Wree, A. Botulinum neurotoxin-a injected intrastriatally into hemiparkinsonian rats improves the initiation time for left and right forelimbs in both forehand and backhand directions. Int. J. Mol. Sci. 2019, 20, 992. [Google Scholar] [CrossRef] [Green Version]
- Antipova, V.; Holzmann, C.; Hawlitschka, A.; Witt, M.; Wree, A. Antidepressant-like properties of intrastriatal botulinum neurotoxin-a injection in a unilateral 6-ohda rat model of parkinson’s disease. Toxins 2021, 13, 505. [Google Scholar] [CrossRef]
- Hawlitschka, A.; Antipova, V.; Schmitt, O.; Witt, M.; Benecke, R.; Mix, E.; Wree, A. Intracerebrally applied botulinum neurotoxin in experimental neuroscience. Curr. Pharm. Biotechnol. 2013, 14, 124–130. [Google Scholar] [CrossRef]
- Hawlitschka, A.; Holzmann, C.; Wree, A.; Antipova, V. Repeated Intrastriatal Botulinum Neurotoxin-A Injection in Hemiparkinsonian Rats Increased the Beneficial Effect on Rotational Behavior. Toxins 2018, 10, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawlitschka, A.; Berg, C.; Schmitt, O.; Holzmann, C.; Wree, A.; Antipova, V. Repeated intrastriatal application of botulinum neurotoxin-A did not influence choline acetyltransferase-immunoreactive interneurons in hemiparkinsonian rat brain—A histological, stereological and correlational analysis. Brain Res. 2020, 1742, 146877. [Google Scholar] [CrossRef]
- Holzmann, C.; Dräger, D.; Mix, E.; Hawlitschka, A.; Antipova, V.; Benecke, R.; Wree, A. Effects of intrastriatal botulinum neurotoxin A on the behavior of Wistar rats. Behav. Brain Res. 2012, 234, 107–116. [Google Scholar] [CrossRef]
- Wree, A.; Mix, E.; Hawlitschka, A.; Antipova, V.; Witt, M.; Schmitt, O.; Benecke, R. Intrastriatal botulinum toxin abolishes pathologic rotational behaviour and induces axonal varicosities in the 6-OHDA rat model of Parkinson’s disease. Neurobiol. Dis. 2011, 41, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Wree, A.; Hawlitschka, A.; Holzmann, C.; Witt, M.; Kurth, J.; Lindner, T.; Mann, T.; Antipova, V. Interlinking potential therapy with botulinum neurotoxin A and Parkinson’s disease. In The Neuroscience of Parkinson’s Disease; Colin, R., Martin Preedy, V., Eds.; Elsevier Academic Press: London, UK, 2020; pp. 665–681. ISBN 9780128159583. [Google Scholar]
- Mehlan, J.; Brosig, H.; Schmitt, O.; Mix, E.; Wree, A.; Hawlitschka, A. Intrastriatal injection of botulinum neurotoxin-A is not cytotoxic in rat brain—A histological and stereological analysis. Brain Res. 2016, 1630, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Mann, T.; Zilles, K.; Dikow, H.; Hellfritsch, A.; Cremer, M.; Piel, M.; Rösch, F.; Hawlitschka, A.; Schmitt, O.; Wree, A. Dopamine, Noradrenaline and Serotonin Receptor Densities in the Striatum of Hemiparkinsonian Rats following Botulinum Neurotoxin-A Injection. Neuroscience 2018, 374, 187–204. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, J.; Yang, Z.Y.; Brown, T.; Lew, R.; Wernick, M.; Ouyang, X.; Yasillo, N.; Chen, C.T.; Mintzer, R.; Cooper, M. Preliminary assessment of extrastriatal dopamine D-2 receptor binding in the rodent and nonhuman primate brains using the high affinity radioligand, 18F-fallypride. Nucl. Med. Biol. 1999, 26, 519–527. [Google Scholar] [CrossRef]
- Rominger, A.; Cumming, P.; Xiong, G.; Koller, G.; Böning, G.; Wulff, M.; Zwergal, A.; Förster, S.; Reilhac, A.; Munk, O.; et al. Fallypride PET measurement of striatal and extrastriatal dopamine D 2/3 receptor availability in recently abstinent alcoholics. Addict. Biol. 2012, 17, 490–503. [Google Scholar] [CrossRef] [PubMed]
- Mann, T.; Kurth, J.; Hawlitschka, A.; Stenzel, J.; Lindner, T.; Polei, S.; Hohn, A.; Krause, B.J.; Wree, A. [18F]fallypride-PET/CT analysis of the dopamine D2/D3receptor in the hemiparkinsonian rat brain following intrastriatal botulinum neurotoxin a injection. Molecules 2018, 23, 587. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.Y.; Kim, C.H.; Jeon, T.J.; Cho, W.G.; Lee, J.D.J.S.; Lee, S.J.; Choi, T.H.; Kim, B.S.; Yi, C.H.; Seo, Y.; et al. Evaluation of dopamine transporters and D2 receptors in hemiparkinsonian rat brains in vivo using consecutive PET scans of [18F]FPCIT and [18F]fallypride. Appl. Radiat. Isot. 2012, 70, 2689–2694. [Google Scholar] [CrossRef]
- Vučcković, M.G.; Li, Q.; Fisher, B.; Nacca, A.; Leahy, R.M.; Walsh, J.P.; Mukherjee, J.; Williams, C.; Jakowec, M.W.; Petzinger, G.M. Exercise elevates dopamine D2 receptor in a mouse model of Parkinson’s disease: In vivo imaging with [18F]fallypride. Mov. Disord. 2010, 25, 2777–2784. [Google Scholar] [CrossRef] [Green Version]
- Björklund, A.; Dunnett, S.B. The amphetamine induced rotation test: A re-assessment of its use as a tool to monitor motor impairment and functional recovery in rodent models of Parkinson’s disease. J. Parkinsons. Dis. 2019, 9, 17–29. [Google Scholar] [CrossRef] [Green Version]
- Boix, J.; Padel, T.; Paul, G. A partial lesion model of Parkinson’s disease in mice—Characterization of a 6-OHDA-induced medial forebrain bundle lesion. Behav. Brain Res. 2015, 284, 196–206. [Google Scholar] [CrossRef]
- Grealish, S.; Mattsson, B.; Draxler, P.; Björklund, A. Characterisation of behavioural and neurodegenerative changes induced by intranigral 6-hydroxydopamine lesions in a mouse model of Parkinson’s disease. Eur. J. Neurosci. 2010, 31, 2266–2278. [Google Scholar] [CrossRef] [PubMed]
- Rosa, I.; Di Censo, D.; Ranieri, B.; Di Giovanni, G.; Scarnati, E.; Alecci, M.; Galante, A.; Florio, T.M. Comparison between tail suspension swing test and standard rotation test in revealing early motor behavioral changes and neurodegeneration in 6-OHDA hemiparkinsonian rats. Int. J. Mol. Sci. 2020, 21, 2874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delfino, M.; Kalisch, R.; Czisch, M.; Larramendy, C.; Ricatti, J.; Taravini, I.R.E.; Trenkwalder, C.; Murer, M.G.; Auer, D.P.; Gershanik, O.S. Mapping the effects of three dopamine agonists with different dyskinetogenic potential and receptor selectivity using pharmacological functional magnetic resonance imaging. Neuropsychopharmacology 2007, 32, 1911–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passamonti, L.; Salsone, M.; Toschi, N.; Cerasa, A.; Giannelli, M.; Chiriaco, C.; Cascini, G.L.; Fera, F.; Quattrone, A. Dopamine-transporter levels drive striatal responses to apomorphine in Parkinson’s disease. Brain Behav. 2013, 3, 249–262. [Google Scholar] [CrossRef]
- Paul, M.L.; Graybiel, A.M.; David, J.C.; Robertson, H.A. D1-like and D2-like dopamine receptors synergistically activate rotation and c-fos expression in the dopamine-depleted striatum in a rat model of Parkinson’s disease. J. Neurosci. 1992, 12, 3729–3742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riddle, L.R.; Kumar, R.; Griffin, S.A.; Grundt, P.; Newman, A.H.; Luedtke, R.R. Evaluation of the D3 dopamine receptor selective agonist/partial agonist PG01042 on l-dopa dependent animal involuntary movements in rats. Neuropharmacology 2011, 60, 284–294. [Google Scholar] [CrossRef] [Green Version]
- Hudson, J.L.; van Horne, C.G.; Strömberg, I.; Brock, S.; Clayton, J.; Masserano, J.; Hoffer, B.J.; Gerhardt, G.A. Correlation of apomorphine- and amphetamine-induced turning with nigrostriatal dopamine content in unilateral 6-hydroxydopamine lesioned rats. Brain Res. 1993, 626, 167–174. [Google Scholar] [CrossRef]
- Kirik, D.; Rosenblad, C.; Bjorklund, A. Preservation of a functional nigrostriatal dopamine pathway by GDNF in the intrastriatal 6-OHDA lesion model depends on the site of administration of the trophic factor. Eur. J. Neurosci. 2000, 12, 3871–3882. [Google Scholar] [CrossRef]
- Lee, C.S.; Sauer, H.; Björklund, A. Dopaminergic neuronal degeneration and motor impairments following axon terminal lesion by intrastriatal 6-hydroxydopamine in the rat. Neuroscience 1996, 72, 641–653. [Google Scholar] [CrossRef]
- Thomas, J.; Wang, J.; Takubo, H.; Sheng, J.; de Jesus, S.; Bankiewicz, K.S. A 6-hydroxydopamine-induced selective parkinsonian rat model: Further biochemical and behavioral characterization. Exp. Neurol. 1994, 126, 159–167. [Google Scholar] [CrossRef]
- Ziegler, M.G.M.; Szechtman, H. Relation between motor asymmetry and direction of rotational behaviour under amphetamine and apomorphine in rats with unilateral degeneration of the nigrostriatal dopamine system. Behav. Brain Res. 1990, 39, 123–133. [Google Scholar] [CrossRef]
- Lehmkuhl, A.M.; Dirr, E.R.; Fleming, S.M. Olfactory Assays for Mouse Models of Neurodegenerative Disease. J. Vis. Exp. 2014, 90, e51804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doty, R.L. Olfactory dysfunction in Parkinson disease. Nat. Rev. Neurol. 2012, 8, 329–339. [Google Scholar] [CrossRef]
- Zhang, W.; Sun, C.; Shao, Y.; Zhou, Z.; Hou, Y.; Li, A. Partial depletion of dopaminergic neurons in the substantia nigra impairs olfaction and alters neural activity in the olfactory bulb. Sci. Rep. 2019, 9, 254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Q.; Chen, S.; Le, W. Hyposmia: A possible biomarker of Parkinson’s disease. Neurosci. Bull. 2014, 30, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Gire, D.H.; Restrepo, D.; Sejnowski, T.J.; Greer, C.; De Carlos, J.A.; Lopez-Mascaraque, L. Temporal Processing in the Olfactory System: Can We See a Smell? Neuron 2013, 78, 416–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, A.; Gire, D.H.; Bozza, T.; Restrepo, D. Precise detection of direct glomerular input duration by the olfactory bulb. J. Neurosci. 2014, 34, 16058–16064. [Google Scholar] [CrossRef]
- Wilson, C.D.; Serrano, G.O.; Koulakov, A.A.; Rinberg, D. A primacy code for odor identity. Nat. Commun. 2017, 8, 1477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinograd, A.; Livneh, Y.; Mizrahi, A. History-dependent odor processing in the mouse olfactory bulb. J. Neurosci. 2017, 37, 12018–12030. [Google Scholar] [CrossRef] [Green Version]
- Paß, T.; Aßfalg, M.; Tolve, M.; Blaess, S.; Rothermel, M.; Wiesner, R.J.; Ricke, K.M. The Impact of Mitochondrial Dysfunction on Dopaminergic Neurons in the Olfactory Bulb and Odor Detection. Mol. Neurobiol. 2020, 57, 3646–3657. [Google Scholar] [CrossRef]
- Pignatelli, A.; Kobayashi, K.; Okano, H.; Belluzzi, O. Functional properties of dopaminergic neurones in the mouse olfactory bulb. J. Physiol. 2005, 564, 501–514. [Google Scholar] [CrossRef] [PubMed]
- Cave, J.W.; Baker, H. Dopamine systems in the forebrain. Adv. Exp. Med. Biol. 2009, 651, 15–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korshunov, K.S.; Blakemore, L.J.; Bertram, R.; Trombley, P.Q. Spiking and Membrane Properties of Rat Olfactory Bulb Dopamine Neurons. Front. Cell. Neurosci. 2020, 14, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLean, J.H.; Shipley, M.T. Postmitotic, postmigrational expression of tyrosine hydroxylase in olfactory bulb dopaminergic neurons. J. Neurosci. 1988, 8, 3658–3669. [Google Scholar] [CrossRef] [Green Version]
- Panzanelli, P.; Fritschy, J.M.; Yanagawa, Y.; Obata, K.; Sassoè-Pognetto, M. GABAergic phenotype of periglomerular cells in the rodent olfactory bulb. J. Comp. Neurol. 2007, 502, 990–1002. [Google Scholar] [CrossRef]
- O’Connor, S.; Jacob, T. Neuropharmacology of the Olfactory Bulb. Curr. Mol. Pharmacol. 2010, 1, 181–190. [Google Scholar] [CrossRef]
- Liu, S.; Plachez, C.; Shao, Z.; Puche, A.; Shipley, M.T. Olfactory bulb short axon cell release of GABA and dopamine produces a temporally biphasic inhibition-excitation response in external tufted cells. J. Neurosci. 2013, 33, 2916–2926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, A.; Marbach, F.; Anselmi, F.; Koh, M.S.; Davis, M.B.; da Silva, P.G.; Delevich, K.; Oyibo, H.K.; Gupta, P.; Li, B.; et al. An Interglomerular Circuit Gates Glomerular Output and Implements Gain Control in the Mouse Olfactory Bulb. Neuron 2015, 87, 193–207. [Google Scholar] [CrossRef] [Green Version]
- Bundschuh, S.T.; Zhu, P.; Schärer, Y.P.Z.; Friedrich, R.W. Dopaminergic modulation of mitral cells and odor responses in the zebrafish olfactory bulb. J. Neurosci. 2012, 32, 6830–6840. [Google Scholar] [CrossRef] [PubMed]
- Escanilla, O.; Yuhas, C.; Marzan, D.; Linster, C. Dopaminergic Modulation of Olfactory Bulb Processing Affects Odor Discrimination Learning in Rats. Behav. Neurosci. 2009, 123, 828–833. [Google Scholar] [CrossRef] [Green Version]
- Tillerson, J.L.; Caudle, W.M.; Parent, J.M.; Gong, C.; Schallert, T.; Miller, G.W. Olfactory discrimination deficits in mice lacking the dopamine transporter or the D2 dopamine receptor. Behav. Brain Res. 2006, 172, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Lussier, S.; Rane, A.; Choi, S.W.; Andersen, J.K. Inducible dopaminergic glutathione depletion in an alpha-synuclein transgenic mouse model results in age-related olfactory dysfunction. Neuroscience 2011, 172, 379–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleming, S.M.; Tetreault, N.A.; Mulligan, C.K.; Hutson, C.B.; Masliah, E.; Chesselet, M.F. Olfactory deficits in mice overexpressing human wildtype α-synuclein. Eur. J. Neurosci. 2008, 28, 247–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winner, B.; Geyer, M.; Couillard-Despres, S.; Aigner, R.; Bogdahn, U.; Aigner, L.; Kuhn, G.; Winkler, J. Striatal deafferentation increases dopaminergic neurogenesis in the adult olfactory bulb. Exp. Neurol. 2006, 197, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Winner, B.; Melrose, H.L.; Zhao, C.; Hinkle, K.M.; Yue, M.; Kent, C.; Braithwaite, A.T.; Ogholikhan, S.; Aigner, R.; Winkler, J.; et al. Adult neurogenesis and neurite outgrowth are impaired in LRRK2 G2019S mice. Neurobiol. Dis. 2011, 41, 706–716. [Google Scholar] [CrossRef] [Green Version]
- Ferraris, A.; Ialongo, T.; Passali, G.C.; Pellecchia, M.T.; Brusa, L.; Laruffa, M.; Guidubaldi, A.; Paludetti, G.; Albanese, A.; Barone, P.; et al. Olfactory dysfunction in Parkinsonism caused by PINK1 mutations. Mov. Disord. 2009, 24, 2350–2357. [Google Scholar] [CrossRef]
- Von Coelln, R.; Thomas, B.; Savitt, J.M.; Lim, K.L.; Sasaki, M.; Hess, E.J.; Dawson, V.L.; Dawson, T.M. Loss of locus coeruleus neurons and reduced startle in parkin null mice. Proc. Natl. Acad. Sci. USA 2004, 101, 10744–10749. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Xiao, Q.; Le, W. Olfactory dysfunction and neurotransmitter disturbance in olfactory bulb of transgenic mice expressing human A53T mutant α-synuclein. PLoS ONE 2015, 10, e0119928. [Google Scholar] [CrossRef]
- Johnson, M.E.; Bergkvist, L.; Mercado, G.; Stetzik, L.; Meyerdirk, L.; Wolfrum, E.; Madaj, Z.; Brundin, P.; Wesson, D.W. Deficits in olfactory sensitivity in a mouse model of Parkinson’s disease revealed by plethysmography of odor-evoked sniffing. Sci. Rep. 2020, 10, 9242. [Google Scholar] [CrossRef]
- Winner, B.; Desplats, P.; Hagl, C.; Klucken, J.; Aigner, R.; Ploetz, S.; Laemke, J.; Karl, A.; Aigner, L.; Masliah, E.; et al. Dopamine receptor activation promotes adult neurogenesis in an acute Parkinson model. Exp. Neurol. 2009, 219, 543–552. [Google Scholar] [CrossRef] [Green Version]
- Ubeda-Bañon, I.; Saiz-Sanchez, D.; De La Rosa-Prieto, C.; Martinez-Marcos, A. α-Synuclein in the olfactory system of a mouse model of Parkinson’s disease: Correlation with olfactory projections. Brain Struct. Funct. 2012, 217, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Lv, D.J.; Li, L.X.; Wang, Y.L.; Qi, D.; Chen, J.; Mao, C.J.; Wang, F.; Liu, Y.; Hu, L.F.; et al. Nicotine improved the olfactory impairment in MPTP-induced mouse model of Parkinson’s disease. Neurotoxicology 2019, 73, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Vroon, A.; Drukarch, B.; Bol, J.G.J.M.; Cras, P.; Brevé, J.J.P.; Allan, S.M.; Relton, J.K.; Hoogland, P.V.J.M.; Van Dam, A.M. Neuroinflammation in Parkinson’s patients and MPTP-treated mice is not restricted to the nigrostriatal system: Microgliosis and differential expression of interleukin-1 receptors in the olfactory bulb. Exp. Gerontol. 2007, 42, 762–771. [Google Scholar] [CrossRef] [PubMed]
- Quik, M.; Di Monte, D.A. Nicotine administration reduces striatal MPP+ levels in mice. Brain Res. 2001, 917, 219–224. [Google Scholar] [CrossRef]
- Gervais, R.; Pager, J. Olfactory bulb excitability selectively modified in behaving rats after local 6-hydroxydopamine treatment. Behav. Brain Res. 1983, 9, 165–179. [Google Scholar] [CrossRef]
- Guan, X.; Blank, J.; Dluzen, D. Depletion of olfactory bulb norepinephrine by 6-OHDA disrupts chemical cue but not social recognition responses in male rats. Brain Res. 1993, 622, 51–57. [Google Scholar] [CrossRef]
- Höglinger, G.U.; Alvarez-Fischer, D.; Arias-Carrión, O.; Djufri, M.; Windolph, A.; Keber, U.; Borta, A.; Ries, V.; Schwarting, R.K.W.; Scheller, D.; et al. A new dopaminergic nigro-olfactory projection. Acta Neuropathol. 2015, 130, 333–348. [Google Scholar] [CrossRef]
- Voronkov, D.N.; Khudoerkov, R.M.; Sal’nikova, O.V.; Stavrovskaya, A.V.; Ol’shanskii, A.S.; Gushchina, A.S. Immunohistochemical Assessment of the Compensatory Responses in Rat Olfactory Bulbs after 6-Hydroxydopamine-Induced Lesion of the Substantia Nigra. Bull. Exp. Biol. Med. 2019, 166, 811–815. [Google Scholar] [CrossRef]
- Femandez-Ruiz, J.; Guzman, R.; Martinez, M.D.; Miranda, M.I.; Bermudez-Rattoni, F.; Drucker-Colin, R. Adrenal medullary grafts restore olfactory deficits and catecholamine levels of 6-OHDA amygdala lesioned animals. J. Neural Transplant. Plast. 1993, 4, 289–297. [Google Scholar] [CrossRef]
- Ilkiw, J.L.; Kmita, L.C.; Targa, A.D.S.S.; Noseda, A.C.D.; Rodrigues, L.S.; Dorieux, F.W.C.C.; Fagotti, J.; dos Santos, P.; Lima, M.M.S.S. Dopaminergic Lesion in the Olfactory Bulb Restores Olfaction and Induces Depressive-Like Behaviors in a 6-OHDA Model of Parkinson’s Disease. Mol. Neurobiol. 2019, 56, 1082–1095. [Google Scholar] [CrossRef]
- Sampaio, T.B.; de Souza, B.S.; Roversi, K.; Schuh, T.; Poli, A.; Takahashi, R.N.; Prediger, R.D. Temporal development of behavioral impairments in rats following locus coeruleus lesion induced by 6-hydroxydopamine: Involvement of β 3-adrenergic receptors. Neuropharmacology 2019, 151, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.M.; Baker, L.S.; Seroogy, K.B.; Genter, M.B. Intranasal Carnosine Mitigates α-Synuclein Pathology and Motor Dysfunction in the Thy1-aSyn Mouse Model of Parkinson’s Disease. ACS Chem. Neurosci. 2021, 12, 2347–2359. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.H.; Kang, S.W.; Kim, K.; Wi, S.; Lee, J.W.; Cho, S.R. Environmental enrichment attenuates oxidative stress and alters detoxifying enzymes in an a53t α-synuclein transgenic mouse model of Parkinson’s disease. Antioxidants 2020, 9, 928. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A.; Gläser, A.; Bräuer, A.U.; Wree, A.; Strotmann, J.; Rolfs, A.; Witt, M. Olfactory performance as an indicator for protective treatment effects in an animal model of neurodegeneration. Front. Integr. Neurosci. 2018, 12, 35. [Google Scholar] [CrossRef] [PubMed]
- Gutièrrez-Mecinas, M.; Crespo, C.; Blasco-Ibáñez, J.M.; Gracia-Llanes, F.J.; Marqués-Marí, A.I.; Nácher, J.; Varea, E.; Martínez-Guijarro, F.J. Distribution of D2 dopamine receptor in the olfactory glomeruli of the rat olfactory bulb. Eur. J. Neurosci. 2005, 22, 1357–1367. [Google Scholar] [CrossRef]
- Nickell, W.T.; Norman, A.B.; Wyatt, L.M.; Shipley, M.T. Olfactory bulb DA receptors may be located on terminals of the olfactory nerve. Neuroreport 1991, 2, 9–12. [Google Scholar] [CrossRef]
- Coronas, V.; Srivastava, L.K.; Liang, J.J.; Jourdan, F.; Moyse, E. Identification and localization of dopamine receptor subtypes in rat olfactory mucosa and bulb: A combined in situ hybridization and ligand binding radioautographic approach. J. Chem. Neuroanat. 1997, 12, 243–257. [Google Scholar] [CrossRef]
- Palacios, J.M.; Niehoff, D.L.; Kuhar, M.J. [3H]Spiperone binding sites in brain: Autoradiographic localization of multiple receptors. Brain Res. 1981, 213, 277–289. [Google Scholar] [CrossRef]
- Charuchinda, C.; Supavilai, P.; Karobath, M.; Palacios, J.M. Dopamine D2 receptors in the rat brain: Autoradiographic visualization using a high-affinity selective agonist ligand. J. Neurosci. 1987, 7, 1352–1360. [Google Scholar] [CrossRef] [Green Version]
- van der Weide, J.; Camps, M.; Horn, A.S.; Palacios, J.M. Autoradiographic localization of dopamine D2 receptors in the rat brain using the new agonist [3H]N-0437. Neurosci. Lett. 1987, 83, 259–263. [Google Scholar] [CrossRef]
- Closse, A.; Camps, M.; Wanner, A.; Palacios, J.M. In vivo labeling of brain dopamine D2 receptors using the high-affinity specific D2 agonist [3H]CV 205-502. Brain Res. 1988, 440, 123–132. [Google Scholar] [CrossRef]
- Wamsley, J.K.; Gehlert, D.R.; Filloux, F.M.; Dawson, T.M. Comparison of the distribution of D-1 and D-2 dopamine receptors in the rat brain. J. Chem. Neuroanat. 1989, 2, 119–137. [Google Scholar] [PubMed]
- Guthrie, K.M.; Pullara, J.M.; Marshall, J.F.; Leon, M. Olfactory deprivation increases dopamine D2 receptor density in the rat olfactory bulb. Synapse 1991, 8, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Boyson, S.J.; McGonigle, P.; Molinoff, P.B. Quantitative autoradiographic localization of the D1 and D2 subtypes of dopamine receptors in rat brain. J. Neurosci. 1986, 6, 3177–3188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huisman, E.; Uylings, H.B.M.; Hoogland, P.V. A 100% increase of dopaminergic cells in the olfactory bulb may explain hyposmia in parkinson’s disease. Mov. Disord. 2004, 19, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Berkowicz, D.A.; Trombley, P.Q. Dopaminergic modulation at the olfactory nerve synapse. Brain Res. 2000, 855, 90–99. [Google Scholar] [CrossRef]
- Ennis, M.; Zimmer, L.A.; Shipley, M.T. Olfactory nerve stimulation activates rat mitral cells via NMDA and non-NMDA receptors in vitro. Neuroreport 1996, 7, 989–992. [Google Scholar] [CrossRef] [PubMed]
- Huisman, E.; Uylings, H.B.M.; Hoogland, P.V. Gender-related changes in increase of dopaminergic neurons in the olfactory bulb of Parkinson’s disease patients. Mov. Disord. 2008, 23, 1407–1413. [Google Scholar] [CrossRef]
- Schober, A. Classic toxin-induced animal models of Parkinson’s disease: 6-OHDA and MPTP. Cell Tissue Res. 2004, 318, 215–224. [Google Scholar] [CrossRef]
- Ghatpande, A.S.; Gelperin, A. Presynaptic muscarinic receptors enhance glutamate release at the mitral/tufted to granule cell dendrodendritic synapse in the rat main olfactory bulb. J. Neurophysiol. 2009, 101, 2052–2061. [Google Scholar] [CrossRef] [Green Version]
- Pressler, R.T.; Inoue, T.; Strowbridge, B.W. Muscarinic receptor activation modulates granule cell excitability and potentiates inhibition onto mitral cells in the rat olfactory bulb. J. Neurosci. 2007, 27, 10969–10981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nai, Q.; Dong, H.W.; Hayar, A.; Linster, C.; Ennis, M. Noradrenergic regulation of GABAergic inhibition of main olfactory bulb mitral cells varies as a function of concentration and receptor subtype. J. Neurophysiol. 2009, 101, 2472–2484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nai, Q.; Dong, H.W.; Linster, C.; Ennis, M. Activation of α1 and α2 noradrenergic receptors exert opposing effects on excitability of main olfactory bulb granule cells. Neuroscience 2010, 169, 882–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petzold, G.C.; Hagiwara, A.; Murthy, V.N. Serotonergic modulation of odor input to the mammalian olfactory bulb. Nat. Neurosci. 2009, 12, 784–791. [Google Scholar] [CrossRef] [PubMed]
- Mundiñano, I.C.; Caballero, M.C.; Ordóñez, C.; Hernandez, M.; DiCaudo, C.; Marcilla, I.; Erro, M.E.; Tuñon, M.T.; Luquin, M.R. Increased dopaminergic cells and protein aggregates in the olfactory bulb of patients with neurodegenerative disorders. Acta Neuropathol. 2011, 122, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Sui, Y.; Horne, M.K.; Stanić, D. Reduced proliferation in the adult mouse subventricular zone increases survival of olfactory bulb interneurons. PLoS ONE 2012, 7, e31549. [Google Scholar] [CrossRef]
- Chiu, W.H.; Carlsson, T.; Depboylu, C.; Höglinger, G.U.; Oertel, W.H.; Ries, V. Selegiline normalizes, while l-DOPA sustains the increased number of dopamine neurons in the olfactory bulb in a 6-OHDA mouse model of Parkinson’s disease. Neuropharmacology 2014, 79, 212–221. [Google Scholar] [CrossRef]
- Wenning, G.K.; Donnemiller, E.; Granata, R.; Riccabona, G.; Poewe, W. 123I-β-CIT and 123I-IBZM-SPECT scanning in levodopa-naive Parkinson’s disease. Mov. Disord. 1998, 13, 438–445. [Google Scholar] [CrossRef]
- Ichise, M.; Kim, Y.J.; Ballinger, J.R.; Vines, D.; Erami, S.S.; Tanaka, F.; Lang, A.E. SPECT imaging of pre- and postsynaptic dopaminergic alterations in L- dopa-untreated PD. Neurology 1999, 52, 1206–1214. [Google Scholar] [CrossRef]
- Cremer, J.N.; Amunts, K.; Graw, J.; Piel, M.; Rösch, F.; Zilles, K. Neurotransmitter receptor density changes in Pitx3ak mice—A model relevant to parkinson’s disease. Neuroscience 2015, 285, 11–23. [Google Scholar] [CrossRef]
- Cremer, J.N.N.; Amunts, K.; Schleicher, A.; Palomero-Gallagher, N.; Piel, M.; Rösch, F.; Zilles, K. Changes in the expression of neurotransmitter receptors in Parkin and DJ-1 knockout mice—A quantitative multireceptor study. Neuroscience 2015, 311, 539–551. [Google Scholar] [CrossRef] [PubMed]
- Hawlitschka, A.; Wree, A. Experimental intrastriatal applications of botulinum neurotoxin-A: A review. Int. J. Mol. Sci. 2018, 19, 1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wedekind, F.; Oskamp, A.; Lang, M.; Hawlitschka, A.; Zilles, K.; Wree, A.; Bauer, A. Intrastriatal administration of botulinum neurotoxin A normalizes striatal D2R binding and reduces striatal D1R binding in male hemiparkinsonian rats. J. Neurosci. Res. 2018, 96, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Palomero-Gallagher, N.; Schleicher, A.A.; Zilles, K.; Löscher, W. The circling ci2 rat mutant revisited: Receptor architecture of the motor cortex. Neuroscience 2010, 170, 542–550. [Google Scholar] [CrossRef]
- Palomero-Gallagher, N.; Schleicher, A.; Lindemann, S.; Lessenich, A.; Zilles, K.; Löscher, W. Receptor fingerprinting the circling ci2 rat mutant: Insights into brain asymmetry and motor control. Exp. Neurol. 2008, 210, 624–637. [Google Scholar] [CrossRef]
- Topic, B.; Willuhn, I.; Palomero-Gallagher, N.; Zilles, K.; Huston, J.P.; Hasenöhrl, R.U. Impaired maze performance in aged rats is accompanied by increased density of NMDA, and 5-HT1A, and α-adrenoceptor binding in hippocampus. Hippocampus 2007, 17, 68–77. [Google Scholar] [CrossRef]
- Marinelli, S.; Vacca, V.; Ricordy, R.; Uggenti, C.; Tata, A.M.; Luvisetto, S.; Pavone, F. The Analgesic Effect on Neuropathic Pain of Retrogradely Transported botulinum Neurotoxin A Involves Schwann Cells and Astrocytes. PLoS ONE 2012, 7, e47977. [Google Scholar] [CrossRef] [Green Version]
- Luvisetto, S. Botulinum neurotoxins in central nervous system: An overview from animal models to human therapy. Toxins 2021, 13, 751. [Google Scholar] [CrossRef]
- Antonucci, F.; Rossi, C.; Gianfranceschi, L.; Rossetto, O.; Caleo, M. Long-distance retrograde effects of botulinum neurotoxin A. J. Neurosci. 2008, 28, 3689–3696. [Google Scholar] [CrossRef]
- Restani, L.; Antonucci, F.; Gianfranceschi, L.; Rossi, C.; Rossetto, O.; Caleo, M. Evidence for Anterograde Transport and Transcytosis of Botulinum Neurotoxin A (BoNT/A). J. Neurosci. 2011, 31, 15650–15659. [Google Scholar] [CrossRef]
- Koizumi, H.; Goto, S.; Okita, S.; Morigaki, R.; Akaike, N.; Torii, Y.; Harakawa, T.; Ginnaga, A.; Kaji, R. Spinal central effects of peripherally applied botulinum neurotoxin A in comparison between its subtypes A1 and A2. Front. Neurol. 2014, 5, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 7th ed.; Elsevier Academic Press: Amsterdam, The Netherlands; London, UK, 2015; ISBN 9780123919496. [Google Scholar]
- Ungerstedt, U.; Arbuthnott, G.W. Quantitative recording of rotational behavior in rats after 6-hydroxy-dopamine lesions of the nigrostriatal dopamine system. Brain Res. 1970, 24, 485–493. [Google Scholar] [CrossRef]
- Schwarting, R.K.W.; Huston, J.P. The unilateral 6-hydroxydopamine lesion model in behavioral brain research. Analysis of functional deficits, recovery and treatments. Prog. Neurobiol. 1996, 50, 275–331. [Google Scholar] [CrossRef]
- Schackel, S.; Pauly, M.-C.C.; Piroth, T.; Nikkhah, G.; Döbrössy, M.D. Donor age dependent graft development and recovery in a rat model of Huntington’s disease: Histological and behavioral analysis. Behav. Brain Res. 2013, 256, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Schiffer, W.K.; Mirrione, M.M.; Biegon, A.; Alexoff, D.L.; Patel, V.; Dewey, S.L. Serial microPET measures of the metabolic reaction to a microdialysis probe implant. J. Neurosci. Methods 2006, 155, 272–284. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, O.; Eipert, P. NeuroVIISAS: Approaching multiscale simulation of the rat connectome. Neuroinformatics 2012, 10, 243–267. [Google Scholar] [CrossRef]
- Schmitt, O.; Eipert, P.; Schwanke, S.; Lessmann, F.; Meinhardt, J.; Beier, J.; Kadir, K.; Karnitzki, A.; Sellner, L.; Klünker, A.C.; et al. Connectome verification: Inter-rater and connection reliability of tract-tracing-based intrinsic hypothalamic connectivity. Brief. Bioinform. 2019, 20, 1944–1955. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | PET/CT 1 | PET/CT 2 | PET/CT 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| BPnd Left | BPnd Right | [%] | BPnd Left | BPnd Right | [%] | BPnd Left | BPnd Right | [%] | |
| Sham + Sham | 3.34 | 3.27 | −1.98 | 2.39 | 2.53 | 5.81 | n/a | n/a | n/a |
| Sham + Sham | 2.2 | 2.29 | 4.11 | 2.77 | 2.83 | 2.08 | 3 | 3.16 | 5.28 |
| Sham + Sham | 2.73 | 2.79 | 2.03 | 2.55 | 2.52 | −1.14 | n/a | n/a | n/a |
| Sham + Sham | 3.33 | 3.46 | 3.87 | 3.12 | 3.13 | 0.24 | 2.78 | 2.67 | −4.18 |
| Sham + Sham | 3.01 | 2.97 | −1.45 | 3.3 | 3.17 | −3.95 | 3.42 | 3.35 | −2.2 |
| Sham + Sham | n/a | n/a | n/a | 3.38 | 3.29 | −2.71 | 3.24 | 3.26 | 0.65 |
| Sham + Sham | n/a | n/a | n/a | 2.72 | 2.78 | 2 | 3.1 | 3.07 | −0.98 |
| Sham + Sham | n/a | n/a | n/a | 3.07 | 3.1 | 1.06 | 3.14 | 3.28 | 4.51 |
| Sham + Sham | n/a | n/a | n/a | 2.9 | 3.15 | 8.89 | n/a | n/a | n/a |
| |Mean| ± SEM | 2.93 ± 0.26 | 2.96 ± 0.29 | 1.32 ± 1.66 | 2.91 ± 0.204 | 2.94 ± 0.21 | 1.36 ± 1.24 | 3.11 ± 0.24 | 3.13 ± 0.26 | 0.51 ± 1.51 |
| 6-OHDA + Sham | 2.5 | 2.44 | −2.22 | 2.61 | 2.54 | −2.68 | 2.5 | 2.56 | 2.15 |
| 6-OHDA + Sham | 3.11 | 3.11 | −0.02 | n/a | n/a | n/a | 4.07 | 3.98 | −2.22 |
| 6-OHDA + Sham | 2.8 | 2.79 | −0.44 | 2.59 | 2.69 | 3.74 | 3.13 | 3.04 | −3.05 |
| 6-OHDA + Sham | 2.93 | 3.08 | 5.3 | 3.22 | 3.42 | 6.24 | 3.66 | 3.85 | 5.33 |
| 6-OHDA + Sham | 2.06 | 2.06 | 0.37 | n/a | n/a | n/a | n/a | n/a | n/a |
| 6-OHDA + Sham | 3.45 | 3.73 | 8.31 | n/a | n/a | n/a | n/a | n/a | n/a |
| 6-OHDA + Sham | n/a | n/a | n/a | 2.4 | 2.56 | 6.67 | 3.43 | 3.53 | 2.8 |
| 6-OHDA + Sham | 2.61 | 2.65 | 1.48 | 2.25 | 2.28 | 0.98 | 3.89 | 3.85 | −1.01 |
| 6-OHDA + Sham | n/a | n/a | n/a | 2.9 | 2.82 | −2.8 | 3.36 | 3.39 | 0.9 |
| |Mean| ± SEM | 2.78 ± 0.22 | 2.84 ± 0.24 | 1.83 ± 1.40 | 2.67 ± 0.24 | 2.72 ± 0.26 | 2.03 ± 1.51 | 3.44 ± 0.22 | 3.46 ± 0.24 | 0.70 ± 1.40 |
| 6-OHDA+BoNT | 2.2 | 2.45 | 11.62 | n/a | n/a | n/a | 2.21 | 2.49 | 12.66 |
| 6-OHDA+BoNT | 1.86 | 1.88 | 0.88 | 3.02 | 3.17 | 4.87 | 1.32 | 1.39 | 5.12 |
| 6-OHDA+BoNT | 3.33 | 3.66 | 10 | 3.2 | 3.44 | 7.7 | n/a | n/a | n/a |
| 6-OHDA+BoNT | 2.22 | 2.52 | 13.67 | 3.16 | 3.2 | 1.54 | 3.34 | 3.56 | 6.75 |
| 6-OHDA+BoNT | 1.87 | 2.11 | 12.84 | n/a | n/a | n/a | 2.79 | 3.07 | 10.18 |
| 6-OHDA+BoNT | 3.58 | 3.81 | 6.26 | 3.07 | 3.32 | 8 | 3.6 | 3.74 | 4.01 |
| 6-OHDA+BoNT | 2.73 | 2.76 | 1.42 | 2.77 | 2.96 | 6.94 | 2.86 | 3.14 | 9.65 |
| 6-OHDA+BoNT | 3.61 | 3.91 | 8.25 | 3.69 | 4 | 8.49 | 4.58 | 4.97 | 8.44 |
| 6-OHDA+BoNT | 3.24 | 3.63 | 11.96 | 3.44 | 3.74 | 8.68 | 4.4 | 4.89 | 11.28 |
| 6-OHDA+BoNT | n/a | n/a | n/a | 2.68 | 2.99 | 11.71 | 4.22 | 4.75 | 12.71 |
| |Mean| ± SEM | 2.74 ± 0.20 | 2.98 ± 0.21 | 8.54 ± 1.24 | 3.13 ± 0.21 | 3.36 ± 0.23 | 7.24 ± 1.31 | 3.26 ± 0.20 | 3.56 ± 0.21 | 8.98 ± 1.24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alberts, T.; Antipova, V.; Holzmann, C.; Hawlitschka, A.; Schmitt, O.; Kurth, J.; Stenzel, J.; Lindner, T.; Krause, B.J.; Wree, A.; et al. Olfactory Bulb D2/D3 Receptor Availability after Intrastriatal Botulinum Neurotoxin-A Injection in a Unilateral 6-OHDA Rat Model of Parkinson’s Disease. Toxins 2022, 14, 94. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14020094
Alberts T, Antipova V, Holzmann C, Hawlitschka A, Schmitt O, Kurth J, Stenzel J, Lindner T, Krause BJ, Wree A, et al. Olfactory Bulb D2/D3 Receptor Availability after Intrastriatal Botulinum Neurotoxin-A Injection in a Unilateral 6-OHDA Rat Model of Parkinson’s Disease. Toxins. 2022; 14(2):94. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14020094
Chicago/Turabian StyleAlberts, Teresa, Veronica Antipova, Carsten Holzmann, Alexander Hawlitschka, Oliver Schmitt, Jens Kurth, Jan Stenzel, Tobias Lindner, Bernd J. Krause, Andreas Wree, and et al. 2022. "Olfactory Bulb D2/D3 Receptor Availability after Intrastriatal Botulinum Neurotoxin-A Injection in a Unilateral 6-OHDA Rat Model of Parkinson’s Disease" Toxins 14, no. 2: 94. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14020094