
Vascular Damage in the Aorta of Wild-Type Mice Exposed to Ionizing Radiation: Sparing and Enhancing Effects of Dose Protraction

 ,
,  , , , , , , and
, , , , , , and 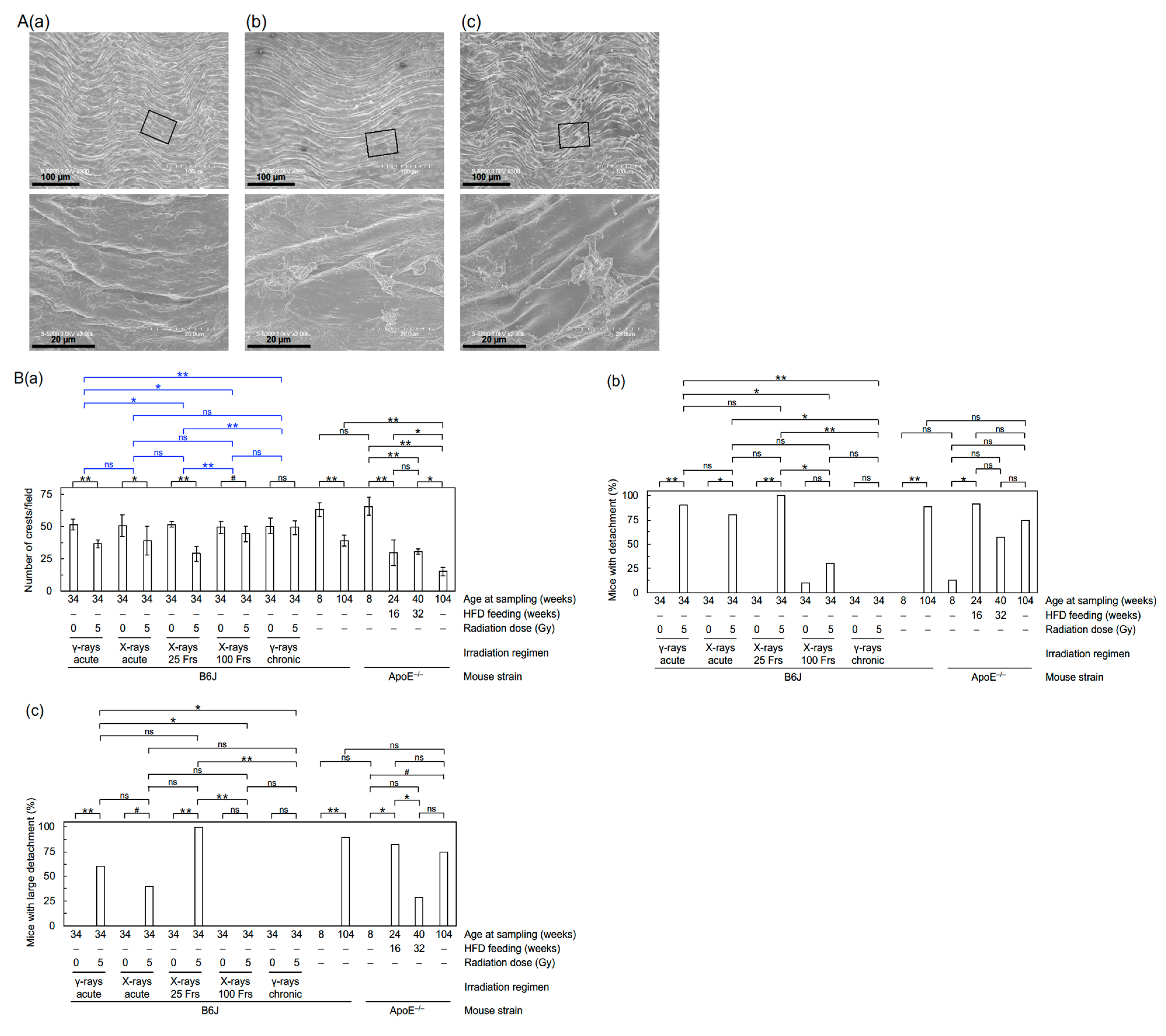{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice and Shipping
2.2. Irradiation and Sampling
2.3. Echocardiography
2.4. FE-SEM
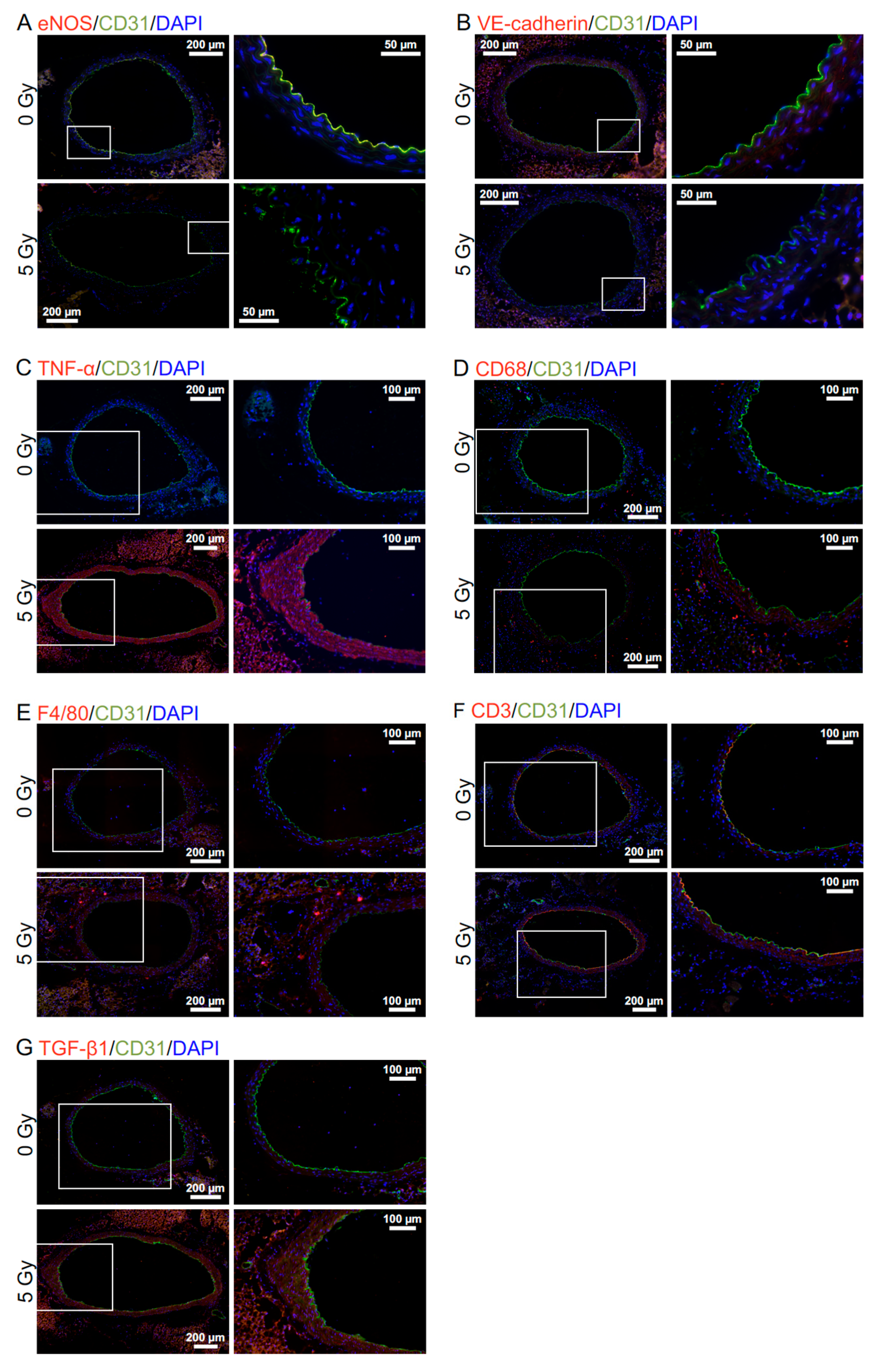
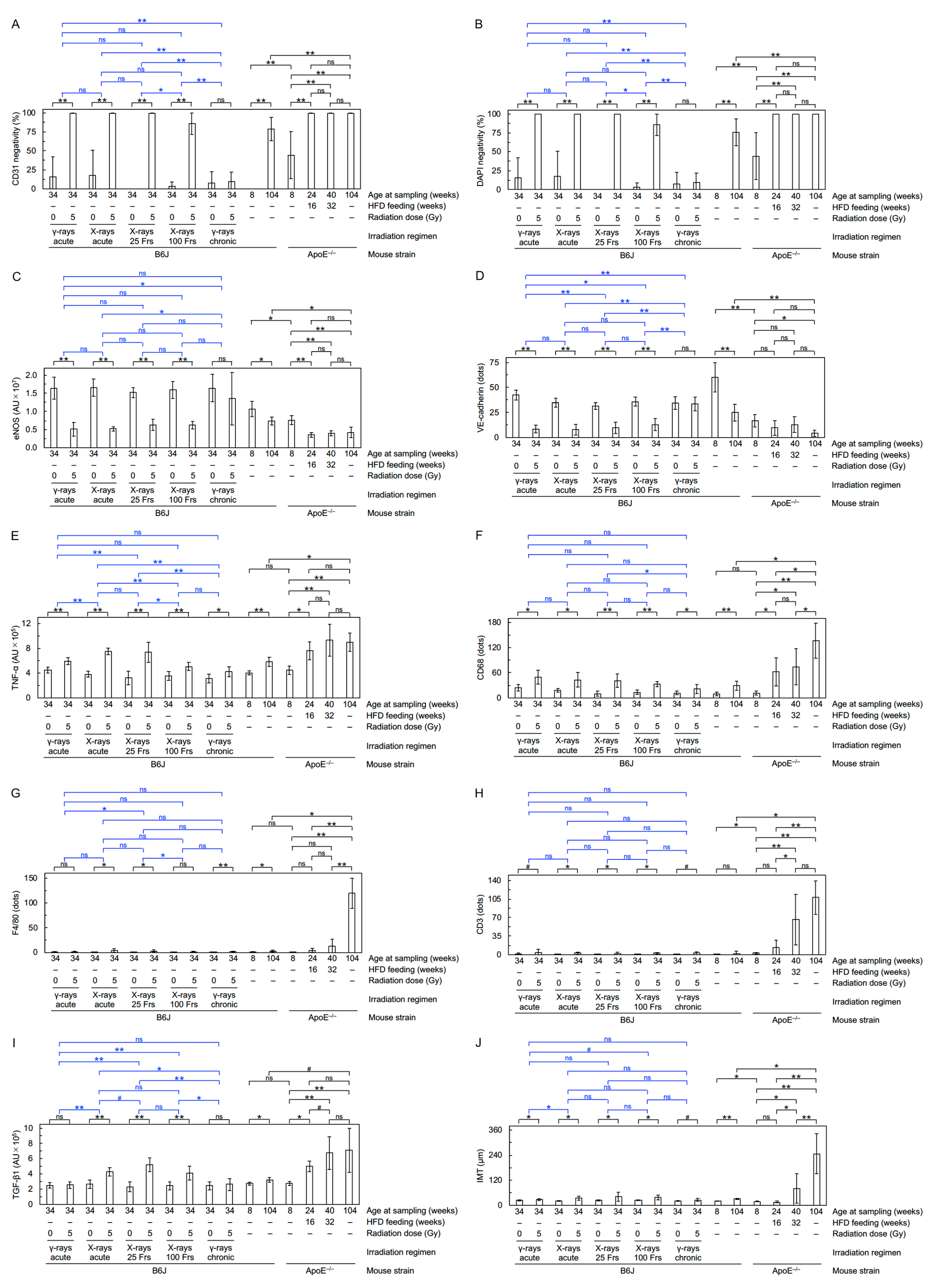
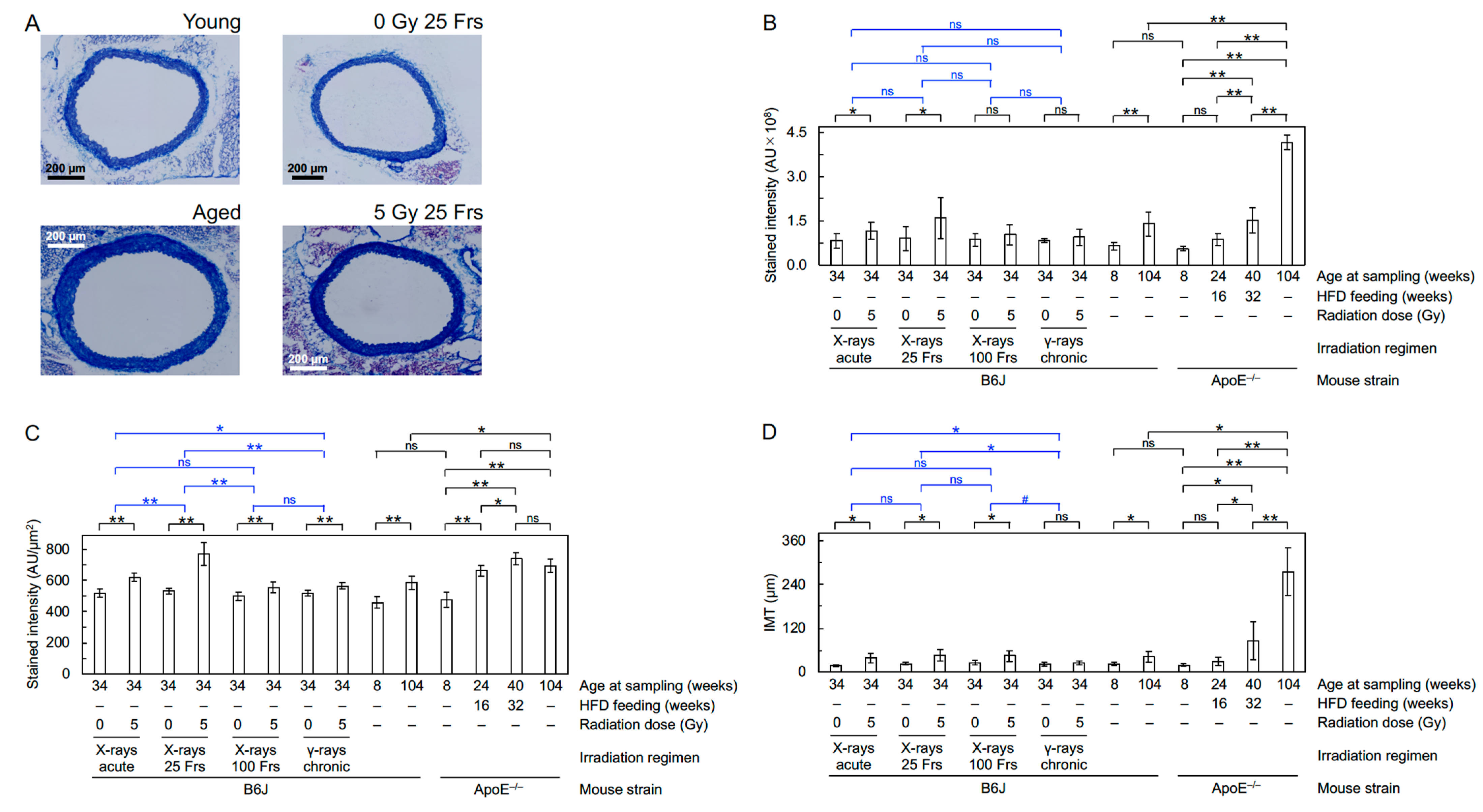
2.5. Immunofluorescence and Histochemistry
2.6. Statistical Analysis
3. Results
3.1. Radiation
3.2. Aging and ApoE Deficiency
4. Discussion
4.1. Radiation
4.1.1. Higher Biological Effectiveness of Acute X-rays vis-à-vis Acute γ-rays
4.1.2. Enhancing Dose Protraction Effect following X-rays in 25 Fractions vis-à-vis Acute X-rays and Acute γ-rays
4.1.3. Sparing Dose Protraction Effect following X-rays in 100 Fractions and Chronic γ-rays vis-à-vis Acute X-rays and Acute γ-rays
4.1.4. Limitations
4.2. Aging and ApoE Deficiency
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Little, M.P. Radiation and Circulatory Disease. Mutat. Res. 2016, 770, 299–318. [Google Scholar] [CrossRef] [Green Version]
- Little, M.P.; Azizova, T.V.; Hamada, N. Low- and Moderate-Dose Non-Cancer Effects of Ionizing Radiation in Directly Exposed Individuals, Especially Circulatory and Ocular Diseases: A Review of the Epidemiology. Int. J. Radiat. Biol. 2021, 97, 782–803. [Google Scholar] [CrossRef]
- Tapio, S.; Little, M.P.; Kaiser, J.C.; Impens, N.; Hamada, N.; Georgakilas, A.G.; Simar, D.; Salomaa, S. Ionizing Radiation-Induced Circulatory and Metabolic Diseases. Environ. Int. 2021, 146, 106235. [Google Scholar] [CrossRef] [PubMed]
- ICRP. ICRP Statement on Tissue Reactions/Early and Late Effects of Radiation in Normal Tissues and Organs–Threshold Doses for Tissue Reactions in a Radiation Protection Context: ICRP Publication 118. Ann. ICRP 2012, 41, 1–322. Available online: https://0-journals-sagepub-com.brum.beds.ac.uk/doi/pdf/10.1177/ANIB_41_1-2 (accessed on 28 September 2021). [CrossRef] [PubMed]
- Hamada, N.; Fujimichi, Y.; Iwasaki, T.; Fujii, N.; Furuhashi, M.; Kubo, E.; Minamino, T.; Nomura, T.; Sato, H. Emerging Issues in Radiogenic Cataracts and Cardiovascular Disease. J. Radiat. Res. 2014, 55, 831–846. [Google Scholar] [CrossRef] [PubMed]
- Hamada, N.; Fujimichi, Y. Classification of Radiation Effects for Dose Limitation Purposes: History, Current Situation and Future Prospects. J. Radiat. Res. 2014, 55, 629–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamada, N.; Kawano, K.I.; Yusoff, F.M.; Furukawa, K.; Nakashima, A.; Maeda, M.; Yasuda, H.; Maruhashi, T.; Higashi, Y. Ionizing Irradiation Induces Vascular Damage in the Aorta of Wild-Type Mice. Cancers 2020, 12, 3030. [Google Scholar] [CrossRef] [PubMed]
- McWilliam, A.; Khalifa, J.; Vasquez Osorio, E.; Banfill, K.; Abravan, A.; Faivre-Finn, C.; van Herk, M. Novel Methodology to Investigate the Effect of Radiation Dose to Heart Substructures on Overall Survival. Int. J. Radiat. Oncol. Biol. Phys. 2020, 108, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Schröder, C.; Stiefel, I.; Tanadini-Lang, S.; Pytko, I.; Vu, E.; Guckenberger, M.; Andratschke, N. Re-Irradiation in the Thorax—An Analysis of Efficacy and Safety Based on Accumulated EQD2 Doses. Radiother Oncol. 2020, 152, 56–62. [Google Scholar] [CrossRef] [PubMed]
- NCRP. Implications of Recent Epidemiologic Studies for the Linear-Nonthreshold Model and Radiation Protection. NCRP Commentary No. 27; 2018; pp. 1–181. Available online: https://ncrponline.org/shop/commentaries/commentary-no-27-implications-of-recent-epidemiologic-studies-for-the-linear-nonthreshold-model-and-radiation-protection-2018/ (accessed on 25 October 2021).
- Azizova, T.V.; Bannikova, M.V.; Briks, K.V.; Grigoryeva, E.S.; Hamada, N. Incidence Risks for Subtypes of Heart Diseases in a Russian Cohort of Mayak Production Association Nuclear Workers. PLoS ONE 2021, 16. in press. [Google Scholar]
- Lowe, D.; Raj, K. Premature Aging Induced by Radiation Exhibits Pro-Atherosclerotic Effects Mediated by Epigenetic Activation of CD44 Expression. Aging Cell 2014, 13, 900–910. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Tong, F.; Qian, C.; Zhang, R.; Dong, J.; Wu, G.; Hu, Y. NEMO Modulates Radiation-Induced Endothelial Senescence of Human Umbilical Veins Through NF-κB Signal Pathway. Radiat. Res. 2015, 183, 82–93. [Google Scholar] [CrossRef]
- Shimizu, I.; Minamino, T. Cellular Senescence in Arterial Diseases. J. Lipid Atheroscler. 2020, 9, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Ho, D.; Vatner, D.E.; Vatner, S.F. Echocardiography in Mice. Curr. Protoc. Mouse Biol. 2011, 1, 71–83. [Google Scholar] [CrossRef]
- Kabacik, S.; Raj, K. Ionising Radiation Increases Permeability of Endothelium Through ADAM10-Mediated Cleavage of VE-Cadherin. Oncotarget 2017, 8, 82049–82063. [Google Scholar] [CrossRef] [Green Version]
- Kouam, P.N.; Rezniczek, G.A.; Adamietz, I.A.; Bühler, H. Ionizing Radiation Increases the Endothelial Permeability and the Transendothelial Migration of Tumor Cells Through ADAM10-Activation and Subsequent Degradation of VE-Cadherin. BMC Cancer 2019, 19, 958. [Google Scholar] [CrossRef] [PubMed]
- Angelini, D.J.; Hyun, S.-W.; Grigoryev, D.N.; Garg, P.; Gong, P.; Singh, I.S.; Passaniti, A.; Hasday, J.D.; Goldblum, S.E. TNF-α Increases Tyrosine Phosphorylation of Vascular Endothelial Cadherin and Opens the Paracellular Pathway Through fyn Activation in Human Lung Endothelia. Am. J. Physiol. Cell Mol. Physiol. 2006, 291, L1232–L1245. [Google Scholar] [CrossRef] [PubMed]
- Monceau, V.; Meziani, L.; Strup-Perrot, C.; Morel, E.; Schmidt, M.; Haagen, J.; Escoubet, B.; Dörr, W.; Vozenin, M.C. Enhanced Sensitivity to Low Dose Irradiation of ApoE−/− Mice Mediated by Early Pro-Inflammatory Profile and Delayed Activation of the TGFβ1 Cascade Involved in Fibrogenesis. PLoS ONE 2013, 8, e57052. [Google Scholar] [CrossRef] [Green Version]
- NCRP. Evaluation of the Relative Effectiveness of Low-Energy Photons and Electrons in Inducing Cancer in Humans. NCRP Report No. 181; 2018; pp. 1–269. Available online: https://ncrponline.org/shop/reports/report-no-181-evaluation-of-the-relative-effectiveness-of-low-energy-photons-and-electrons-in-inducing-cancer-in-humans-2018/ (accessed on 25 October 2021).
- Hamada, N.; Schettino, G.; Kashino, G.; Vaid, M.; Suzuki, K.; Kodama, S.; Vojnovic, B.; Folkard, M.; Watanabe, M.; Michael, B.D.; et al. Histone H2AX Phosphorylation in Normal Human Cells Irradiated with Focused Ultrasoft X Rays: Evidence for Chromatin Movement During Repair. Radiat. Res. 2006, 166, 31–38. [Google Scholar] [CrossRef]
- Schmid, E.; Regulla, D.; Kramer, H.M.; Harder, D. The Effect of 29 kV X Rays on the Dose Response of Chromosome Aberrations in Human Lymphocytes. Radiat. Res. 2002, 158, 771–777. [Google Scholar] [CrossRef] [Green Version]
- Baumann, M.; Liertz, C.; Baisch, H.; Wiegel, T.; Lorenzen, J.; Arps, H. Impact of Overall Treatment Time of Fractionated Irradiation on Local Control of Human FaDu Squamous Cell Carcinoma in Nude Mice. Radiother. Oncol. 1994, 32, 137–143. [Google Scholar] [CrossRef]
- Seol, M.-A.; Jung, U.; Eom, H.S.; Kim, S.-H.; Park, H.-R.; Jo, S.-K. Prolonged Expression of Senescence Markers in Mice Exposed to Gamma-Irradiation. J. Vet. Sci. 2012, 13, 331–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervelli, T.; Panetta, D.; Navarra, T.; Andreassi, M.G.; Basta, G.; Galli, A.; Salvadori, P.A.; Picano, E.; Del Turco, S. Effects of Single and Fractionated Low-Dose Irradiation on Vascular Endothelial Cells. Atherosclerosis 2014, 235, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Zablotska, L.B.; Little, M.P.; Cornett, R.J. Potential Increased Risk of Ischemic Heart Disease Mortality with Significant Dose Fractionation in the Canadian Fluoroscopy Cohort Study. Am. J. Epidemiol. 2014, 179, 120–131. [Google Scholar] [CrossRef] [Green Version]
- Little, M.P.; Pawel, D.; Misumi, M.; Hamada, N.; Cullings, H.M.; Wakeford, R.; Ozasa, K. Lifetime Mortality Risk from Cancer and Circulatory Disease Predicted from the Japanese Atomic Bomb Survivor Life Span Study Data Taking Account of Dose Measurement Error. Radiat. Res. 2020, 194, 259–276. [Google Scholar] [CrossRef]
- Hoel, D.G.; Carnes, B.A. Cardiovascular Effects of Fission Neutron or 60Co γ Exposure in the B6CF1 Mouse. Int. J. Radiat. Biol. 2017, 93, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Rühm, W.; Woloschak, G.E.; Shore, R.E.; Azizova, T.V.; Grosche, B.; Niwa, O.; Akiba, S.; Ono, T.; Suzuki, K.; Iwasaki, T.; et al. Dose and Dose-Rate Effects of Ionizing Radiation: A Discussion in the Light of Radiological Protection. Radiat. Environ. Biophys. 2015, 54, 379–401. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.L.; Lee, J.W.; Daniel, A.R.; Holbrook, M.; Hasapis, S.; Wright, A.O.; Brownstein, J.; Da Silva Campos, L.; Ma, Y.; Mao, L.; et al. Characterization of Cardiovascular Injury in Mice Following Partial-Heart Irradiation with Clinically Relevant Dose and Fractionation. Radiother. Oncol. 2021, 157, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Foray, N.; Bourguignon, M.; Hamada, N. Individual Response to Ionizing Radiation. Mutat. Res. Rev. Mutat. Res. 2016, 770, 369–386. [Google Scholar] [CrossRef]
- Mancuso, M.; Pasquali, E.; Braga-Tanaka, I., 3rd; Tanaka, S.; Pannicelli, A.; Giardullo, P.; Pazzaglia, S.; Tapio, S.; Atkinson, M.J.; Saran, A. Acceleration of Atherogenesis in ApoE−/− Mice Exposed to Acute or Low-Dose-Rate Ionizing Radiation. Oncotarget 2015, 6, 31263–31271. [Google Scholar] [CrossRef]
- Mitchel, R.E.; Hasu, M.; Bugden, M.; Wyatt, H.; Little, M.P.; Gola, A.; Hildebrandt, G.; Priest, N.D.; Whitman, S.C. Low-Dose Radiation Exposure and Atherosclerosis in ApoE−/− Mice. Radiat. Res. 2011, 175, 665–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebrahimian, T.G.; Beugnies, L.; Surette, J.; Priest, N.; Gueguen, Y.; Gloaguen, C.; Benderitter, M.; Jourdain, J.R.; Tack, K. Chronic Exposure to External Low-Dose Gamma Radiation Induces an Increase in Anti-inflammatory and Anti-oxidative Parameters Resulting in Atherosclerotic Plaque Size Reduction in ApoE−/− Mice. Radiat. Res. 2018, 189, 187–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.Y.; Kim, A.; Seong, K.M. Chronic Radiation Exposure Aggravates Atherosclerosis by Stimulating Neutrophil Infiltration. Int. J. Radiat. Biol. 2021, 97, 1270–1281. [Google Scholar] [CrossRef] [PubMed]
- NCRP. Approaches for Integrating Information from Radiation Biology and Epidemiology to Enhance Low-Dose Health Risk Assessment. NCRP Report No. 186; 2020; pp. 1–270. Available online: https://ncrponline.org/shop/reports/report-no-186-approaches-for-integrating-information-from-radiation-biology-and-epidemiology-to-enhance-low-dose-health-risk-assessment-2020/ (accessed on 25 October 2021).
- Chauhan, V.; Hamada, N.; Monceau, V.; Ebrahimian, T.; Adam, N.; Wilkins, R.C.; Soji, S.; Patel, Z.S.; Huff, J.L.; Simonetto, C.; et al. Expert Consultation is Vital for Adverse Outcome Pathway Development: A Case Example of Cardiovascular Effects of Ionizing Radiation. Int. J. Radiat. Biol. 2021, 97, 1516–1525. [Google Scholar] [CrossRef]
- Georgakilas, A.G.; Pavlopoulou, A.; Louka, M.; Nikitaki, Z.; Vorgias, C.E.; Bagos, P.G.; Michalopoulos, I. Emerging Molecular Networks Common In Ionizing Radiation, Immune And Inflammatory Responses By Employing Bioinformatics Approaches. Cancer Lett. 2015, 368, 164–172. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamada, N.; Kawano, K.-i.; Nomura, T.; Furukawa, K.; Yusoff, F.M.; Maruhashi, T.; Maeda, M.; Nakashima, A.; Higashi, Y. Vascular Damage in the Aorta of Wild-Type Mice Exposed to Ionizing Radiation: Sparing and Enhancing Effects of Dose Protraction. Cancers 2021, 13, 5344. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13215344
Hamada N, Kawano K-i, Nomura T, Furukawa K, Yusoff FM, Maruhashi T, Maeda M, Nakashima A, Higashi Y. Vascular Damage in the Aorta of Wild-Type Mice Exposed to Ionizing Radiation: Sparing and Enhancing Effects of Dose Protraction. Cancers. 2021; 13(21):5344. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13215344
Chicago/Turabian StyleHamada, Nobuyuki, Ki-ichiro Kawano, Takaharu Nomura, Kyoji Furukawa, Farina Mohamad Yusoff, Tatsuya Maruhashi, Makoto Maeda, Ayumu Nakashima, and Yukihito Higashi. 2021. "Vascular Damage in the Aorta of Wild-Type Mice Exposed to Ionizing Radiation: Sparing and Enhancing Effects of Dose Protraction" Cancers 13, no. 21: 5344. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13215344