
Exercise Counteracts the Deleterious Effects of Cancer Cachexia


1
Cachexia Research Laboratory, Exercise Science Research Center, Department of Health, Human Performance and Recreation, University of Arkansas, Fayetteville, AR 72701, USA
2
Molecular Muscle Mass Regulation Laboratory, Exercise Science Research Center, Department of Health, Human Performance and Recreation, University of Arkansas, Fayetteville, AR 72701, USA
3
Exercise Muscle Biology Laboratory, Exercise Science Research Center, Department of Health, Human Performance and Recreation, University of Arkansas, Fayetteville, AR 72701, USA
*
Author to whom correspondence should be addressed.
Cancers 2022, 14(10), 2512; https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14102512
Submission received: 17 April 2022
/
Revised: 12 May 2022
/
Accepted: 17 May 2022
/
Published: 19 May 2022
(This article belongs to the Special Issue Cancer Cachexia: Molecular Insights and Clinical Implications)
Abstract
:Simple Summary
This review provides an overview of the effects of exercise training on the major mechanisms related to cancer cachexia (CC). The review also discusses how cancer comorbidities can influence the ability of patients/animals with cancer to perform exercise training and what precautions should be taken when they exercise. The contribution of other factors, such as exercise modality and biological sex, to exercise effectiveness in ameliorating CC are also elaborated in the final sections. We provide meticulous evidence for how advantageous exercise training can be in patients/animals with CC at molecular and cellular levels. Finally, we emphasise what factors should be considered to optimise and personalise an exercise training program in CC.
Abstract
Cancer cachexia (CC) is a multifactorial syndrome characterised by unintentional loss of body weight and muscle mass in patients with cancer. The major hallmarks associated with CC development and progression include imbalanced protein turnover, inflammatory signalling, mitochondrial dysfunction and satellite cell dysregulation. So far, there is no effective treatment to counteract muscle wasting in patients with CC. Exercise training has been proposed as a potential therapeutic approach for CC. This review provides an overview of the effects of exercise training in CC-related mechanisms as well as how factors such as cancer comorbidities, exercise modality and biological sex can influence exercise effectiveness in CC. Evidence in mice and humans suggests exercise training combats all of the hallmarks of CC. Several exercise modalities induce beneficial adaptations in patients/animals with CC, but concurrent resistance and endurance training is considered the optimal type of exercise. In the case of cancer patients presenting comorbidities, exercise training should be performed only under specific guidelines and precautions to avoid adverse effects. Observational comparison of studies in CC using different biological sex shows exercise-induced adaptations are similar between male and female patients/animals with cancer, but further studies are needed to confirm this.
1. Introduction
Cancer cachexia (CC) is a devastating, multifactorial and so far irreversible syndrome characterised mainly by loss of skeletal muscle mass and body fat [1]. CC affects up to 80% of cancer patients and is responsible for 20–30% of cancer deaths [1,2]. The stages of CC include: (i) precachexia, clinically defined as ≤5% weight loss without lifestyle modification over the past 6 months; (ii) cachexia, defined as >5% involuntary weight loss over the past six months or a body mass index (BMI) < 20 kg/m²; and (iii) refractory cachexia, characterised by an irreversible rapid weight loss accompanied by a life expectancy < three months [3].
The major known mechanisms contributing to the development and progression of CC include imbalanced protein turnover, inflammatory signalling, mitochondrial degeneration and muscle stem cell (satellite cell) dysregulation [4,5]. Specifically, CC is characterised by increased muscle protein degradation combined with decreased muscle protein synthesis [5]. Activation of protein degradation signalling through either the ubiquitin proteasome system (UPS) or the lysosomal proteasome system (LPS) is well described in different preclinical and clinical models [5,6,7,8,9,10]. Chronic inflammation characterised by increased levels of inflammatory factors in skeletal muscle such as tumour necrosis factor α (TNF-α) and IL-6 is also a major mechanism of CC in both patients and mouse models [11,12,13]. Furthermore, mitochondrial degeneration including network degeneration, elevated levels of reactive oxygen species (ROS) production/emission, decreased mitochondrial quality and reduced aerobic metabolism [5,14,15] are considered hallmarks of CC. Finally, skeletal muscle satellite cells, as well as their regulators known as myogenic regulatory factors, may play a role in mechanisms that underlie CC [16]. Satellite cell differentiation is inhibited in CC and muscle fibres that are damaged during CC may undergo degeneration [4]. The contributions of dysregulated protein turnover, increased chronic inflammation, mitochondrial dysfunction and myogenic disruption to the development and progression of CC are complex and intertwined. Future therapeutic interventions for CC should be focused on combating this constellation of dysfunction.
Nutritional interventions are insufficient to fully reverse the loss of body weight induced by CC [17,18]. To date, there is no effective treatment to counteract wasting in patients with CC. Exercise training is considered a therapeutic approach for many chronic diseases. Generally, exercise can improve cardiorespiratory and neuromuscular systems and promote psychological wellbeing [19,20,21]. The positive effects of exercise are accompanied by beneficial adaptations at a molecular level [22]. Specifically, exercise training increases the gene expression of skeletal muscle myogenic factors (e.g., Pax7, MyoD, Myogenin etc.) [23] and activates molecular pathways that regulate skeletal muscle mitochondrial biogenesis (e.g., elevated levels of PGC1-α) [24]. Furthermore, systematic exercise training reduces chronic circulating inflammatory IL-6 levels, especially in individuals with a non-active lifestyle [25].
Exercise training has been proposed as a non-pharmacological therapeutic approach for CC [26,27,28]. The potential of exercise training to decrease cancer-induced inflammation and oxidative stress [29], as well as mitigate cancer-induced suppression of anabolic signalling and protein synthesis [30] can be considered an efficient “tool” to attenuate skeletal muscle abnormalities observed in CC. Preclinical studies highlight the benefits of exercise training in animals with different types of cancer (e.g., ApcMin/+ and C26 colorectal cancer mice [31,32], breast cancer mice [33,34,35,36], SENCAR skin cancer mice [37]) and rat models such as Walker 256 carcinosarcoma [38,39,40]). Irrespective of the model, exercise training further reduces tumour incidence, tumour multiplicity and tumour growth [41], as well as attenuates the progression of CC [31,32]. A major unavoidable limitation of pre-clinical studies in CC mouse models is the short timeline (three to four weeks) the mice have after the inoculation of cancer cells. For this reason, the majority of pre-clinical studies begin the exercise intervention before or just after the inoculation of cancer cells to succeed exercise-induced adaptations [42]. This timepoint is undoubtedly earlier than the development and progression of CC (around three to four weeks after cancer cells inoculation). For this reason, most pre-clinical studies are designed with prevention of cachexia as the primary goal leaving a relative dearth in data regarding reversal of existent cachexia.
Previous clinical studies in patients with cancer from early to advanced stages of disease have concluded that exercise training, independent of modality (i.e., resistance, endurance or concurrent training), has beneficial effects on skeletal muscle health, physical performance, quality of life and psychological health [27,28,43,44,45,46]. Exercise training can be also advantageous as a pre-operative intervention in cancer patients [47,48,49]. Specifically, exercise not only improves the physical capacity and function of cancer patients [47,48], but also ameliorates postoperative recovery [47] and may have a protective direct effect on tumour growth through altering the gene expression of immunity and inflammation “key” factors [49]. Given the heterogeneity observed in cancer patients (i.e., different cancer diagnoses, cancer stages and symptoms) in exercise clinical trials, further works are necessary to evaluate exercise training as a monotherapy or combinatorial therapy throughout all phases of cancer, including the phases of CC development and progression [42,50,51]. To this point, ongoing clinical trials with exercise-based interventions in cancer patients are presented in the new Supplementary Table S1.
Unfortunately, exercise training is not always feasible in humans or animals with cancer due to cancer- and chemotherapy-induced fatigue, anaemia, cardiac dysfunction and other comorbidities [52,53,54]. For instance, mild endurance training in C26 colorectal cancer mice, a CC mouse model that suffers additionally from anaemia, does not prevent body weight and muscle loss, and even worsened their condition [53,55]. In cases of comorbidities along with cancer, exercise training should be performed only under specific guidelines, precautions and supervision by specialists. The exercise workload including exercise intensity, duration, frequency and modality should be meticulously assessed in each case individually. Further preclinical studies using a translatable exercise framework that can account for the comorbidities accompanying cancer are needed to optimise exercise training programs to avoid any potential exercise-induced adverse effect. Although all forms of exercise (resistance, endurance and concurrent training) induce beneficial adaptations in CC-induced skeletal muscle damage [30,31,32,56,57], concurrent resistance and endurance training may surpass resistance training per se or endurance training per se. Such an approach improves both anaerobic and aerobic skeletal muscle metabolism simultaneously [58,59,60], both of which are compromised with CC.
Biological sex differences have been observed in CC development and progression, with females more resistant to the cancer-induced skeletal muscle decline [15,61]. However, no study has directly investigated if exercise-induced beneficial adaptations in skeletal muscle of patients or mice with CC differ between females and males. Studies in healthy individuals have shown females and males respond differently to exercise stimuli [62,63]. When observationally comparing studies using the same CC mouse model but different biological sex, exercise-induced adaptations are similar in females and males [57,64]. Further investigation is needed to confirm these observations.
The primary purpose of this review is to highlight the effects of exercise training on molecular and physiological mechanisms related to CC, namely protein turnover, inflammatory signalling, mitochondrial function and satellite cell-related process. Importantly, we emphasise the contraindications of exercise in some cancer conditions with comorbidities. Secondly, we describe the effects and importance of different exercise modalities. Based on the current literature, we propose the most advantageous exercise modality for cancer patients. Finally, we aim to specify the differences between females and males in response to exercise training during CC development and progression.
2. Effects of Exercise Training on Cancer Cachexia (CC) Mechanisms
2.1. Protein Turnover in Response to Exercise
One of the major hallmark features of CC is the net loss of skeletal muscle protein due to the imbalance between protein breakdown and protein synthesis [5]. Preclinical studies in Lewis Lung Carcinoma (LLC) male [7] and female [15] mice, as well as in a colon cancer cachectic mouse model, ApcMin/+ mice [6] revealed a combination of a loss of protein synthesis and increased markers of protein breakdown. Specifically, protein fractional synthesis rate (FSR) decreases around 40%, while muscle protein ubiquitination and protein levels of ubiquitin-proteasome marker FOXO1 increase around 50% in male LLC mice 4 weeks after tumour implantation [7]. Similar to males, cachectic female LLC mice also present reduced muscle protein FSR in gastrocnemius, whereas the mRNA levels of Ubiquitin C, Atrogin-1 and Murf-1 are elevated [15]. In accordance with the results in LLC mice [7,15], myofibrillar protein synthesis as well as mRNA content of IGF-1 and phosphorylation of the mTOR signalling cascade decrease during CC, while protein degradation (ATP dependent and independent) increases during CC progression in ApcMin/+ mice [6]. For further information about the advantages and disadvantages of preclinical CC mouse models, the reader is directed to a recent review highlighting this topic [5].
Exercise training increases protein synthesis in both healthy and diseased populations [65,66,67], while decelerating the high protein degradation induced by chronic diseases [68,69]. Improvements in protein synthesis have also been observed in different cancer mouse models after performing exercise training [30,70,71]. Specifically, endurance-type wheel running (60 min in 5–6.5 m/min, 5 days/week, 11 weeks) leads to enhanced phosphorylation of mTOR, a critical regulator of skeletal muscle protein synthesis [72], in female C26-bearing mice [71]. Repeated eccentric contractions (10 tetanic contractions in 100 Hz, 6–12 V × 6 sets, 4 times/week for 2 weeks) of the tibialis anterior (TA), considered a representative model of resistance exercise training for animal studies, improves protein synthesis and mTORC1 signalling and attenuates atrophy of oxidative and glycolytic muscle fibres in male ApcMin/+ mice [30]. Activation of protein synthesis and mTORC1 signalling is also induced by a single bout of eccentric contractions in male ApcMin/+ mice. Acute activation of mTORC1 occurs on the condition that the electrical stimulation that elicits the eccentric contractions is not low-frequency (i.e., 10 Hz, 5 V) [73] but is sufficient to induce tetanic contractions (i.e., 100 Hz, 6–12 V) [70]. Performing either single [64] or multiple bouts of tetanic eccentric contractions [57] in female [64] and male [57] ApcMin/+ mice not only activates protein synthesis signalling but also suppresses muscle catabolic signalling. Protein synthetic responses occur by reducing the protein expression of cellular energy-sensing enzyme 5′-adenosine monophosphate-activated protein kinase (AMPK), which is an established mTOR inhibitor and chronically activated in severely cachectic muscle [74].
Beyond anabolic signalling, AMPK is known to activate catabolic signalling, in addition to its canonical functions in promoting energy metabolism and mitochondrial adaptation [75]. Herein, inhibition of AMPK is also observed after 8-week moderate-intensity treadmill running (1 h at 18 m/min, 5% grade, 6 days/week) in male ApcMin/+ mice [31]. Furthermore, voluntary wheel running (for ~2.5 weeks) counteracts cancer-induced protein degradation through suppressing both the induction of ubiquitin ligases (Atrogin1, also known as Fbxo32; and Murf1, also known as Trim63) and the protein expression of autophagic markers (LC3bII/LC3bI ratio and p62/Gapdh ratio) in female C26 tumour-bearing mice [32]. Similarly, decreased LC3bII/LC3bI ratio is also observed by either endurance training (15–45 min wheel running at 5–11 m/minor, 3 days/week, 2.5 weeks) [76] or concurrent resistance (inclined ladder climbing with gradually increased resistance load) and aerobic (25-min wheel running in 5–9 m/min speed) training (4 days/week, 5.5 weeks) [58] in male C26 tumour-bearing mice. Interestingly, similar exercise adaptations in skeletal muscle protein turnover are observed when tumour-bearing mice are treated with chemotherapy agents [77]. Specifically, moderate exercise training on a treadmill (40–60 min at 60% of maximum speed, 5 days/week, 3 weeks) reduces the gene expression of catabolic markers (Fbxo32, Trim63 and Myostatin) related to atrophy and increases it in LLC mice treated with doxorubicin chemotherapy drug [77].
Similarly, a clinical trial reports that 10-week concurrent resistance (knee extension, leg press, lateral pull-down, chest-press, back extension and sit-ups) and endurance (ergometer cycling) training in cancer patients treated with chemotherapy can prevent cancer- and chemotherapy-induced disruptions in molecular signalling cascades associated with the ubiquitin proteasome system (e.g., Atrogin-1 and Murf-1) and protein synthesis (e.g., mTOR) [78]. As a result, exercise training seems to be a good strategy to combat unbalanced protein turnover induced by cancer and chemotherapy in preclinical models and clinical applications (Figure 1 and Table 1); however, further clinical trials are needed to establish this empirically.
2.2. Inflammatory Signaling in Response to Exercise
Chronic inflammation accompanied by elevated circulating inflammatory cytokines is another major hallmark mechanism of CC pathogenesis [11]. Physical activity is associated with lower odds of having elevated inflammation levels (e.g., elevated levels of C-reactive protein—CRP) [79,80,81]. In preclinical studies, there is evidence that exercise has an anti-inflammatory effect by reducing TNF-α expression [82,83,84]. The mechanism underlying the anti-inflammatory response after performing exercise training is related to the major role of the cytokine IL-6 [81]. Acute exercise can increase circulating levels of IL-6 [85], which induces the production of the anti-inflammatory cytokines IL-1ra and IL-10 [85,86]. Exercise also inhibits the production of the inflammatory cytokine, TNF-α [81,87,88]. Acute elevation of IL-6 after exercise stimulus is therefore considered a beneficial response that could promote protein synthesis and advantageous skeletal muscle adaptations [89]. On the other hand, long-lasting elevated systemic IL-6 levels activate catabolic signalling pathways associated with muscle wasting [89]. Long-term exercise training decreases circulating IL-6 levels, especially in individuals following a sedentary lifestyle [25]. Also, an inverse correlation between the amount of weekly physical activity and the level of plasma IL-6 in the resting state occurs in healthy males [90]. In addition, aged lifelong exercisers with an aerobic training history of ~50 years not only present with lower serum IL-6 levels compared to old healthy non-exercisers in the resting state, but also have similar post-exercise (4 h after an acute resistance training session) gene expression of skeletal muscle anti-inflammatory markers compared to young exercisers [91].
The anti-inflammatory benefits of exercise training have been described in preclinical and clinical cancer studies [33,92,93,94,95]. Specifically, exercise training by treadmill (60 min at 20 m/min, 5% grade, 6 days/week, 20 weeks) reduces the plasma concentration of two major inflammatory markers, MCP-1 and IL-6, in female breast-tumour bearing mice (C3(1)SV40Tag mouse model) [33]. This inflammatory reduction is associated with the deceleration of breast tumour progression observed in C3(1)SV40Tag mice after the 20-week exercise training program [33]. Treadmill running (15–30 min at 15–20 m/min, 3 days/week, 6 weeks) also reduces TNF-α concentration in the plasma and TNF-α gene expression in the colon of mice with azoxymethane (AOM)-induced colon cancer [92]. Similarly, voluntary wheel running for 4 weeks in breast cancer mice (PyMT) decreases not only circulating but intramuscular levels of TNF-α protein, as well as reduces the intramuscular mRNA content of several TNF-α target genes (Traf2, IκB-α, Ank1 and NFκB1) [36]. Furthermore, a treadmill running program (25–40 min at 14–20 m/min, 5 days/week, 8–14 weeks), starting either before or after injecting estrogen-dependent MC4L2 cancer cells in female mice, reduces breast tumour volume and the levels of IL-6 in the tumour, on condition that the mice continue exercising after the onset of tumorigenesis [93]. In addition, the increased protein concentration of IL-6 observed in tumour of LLC mice treated with doxorubicin chemotherapy is mitigated by 3-weeks of moderate exercise training on a treadmill (40–60 min at 60% of maximum speed, 5 days/week) [77].
Surprisingly, a preclinical study in ApcMin/+ colorectal cancer mice showed that treadmill running (1 h at 18 m/min speed and 5% grade, 6 days/week, 8 weeks) does not suppress systemic IL-6 overexpression induced by electroporating IL-6 plasmid in quadriceps. This result is surprising since an exercise training program effectively prevents the IL-6-induced decrease in body weight of male ApcMin/+ mice [31]. Although exercise training improves phenotypic characteristics in male ApcMin/+ mice, such as lean mass and motor function, 5-week voluntary wheel running does not decrease either tumour burden or the elevated circulating IL-6 levels [96].
When data from two randomised controlled exercise training trials in breast cancer patients were pooled and analysed together, the findings showed that exercise training (12–18 weeks of resistance or concurrent training) does not decrease the cancer/chemotherapy-induced levels of the inflammatory markers IL-6 and IL-6/IL-1ra ratio [97]. However, it seems that exercise intensity plays a crucial role on the inflammatory profile of patients with cancer during and after cancer treatment (chemotherapy and/or radiotherapy and/or endocrine therapy) [98]. Specifically, performing high-intensity concurrent resistance and aerobic training during cancer treatment results in a lesser increase of the plasma inflammatory markers CRP and TNF-α immediately after completion of cancer treatment, compared to performing low-intensity concurrent resistance and aerobic training [98]. Also, many clinical trials have concluded that exercise training promotes an anti-inflammatory profile in cancer patients undergoing chemotherapy or radiation therapy [94,99,100]. Specifically, combination of resistance training with high-intensity aerobic training (60-min sessions, 2 days/week for 16 weeks) [100] or resistance training per se (60-min sessions, 2 days/week, 12 weeks) [99] mitigates [100] and even counteracts [99] cancer-mediated increased levels of IL-6. In conclusion, exercise training suppresses the CC-induced elevation of inflammatory markers in different CC mouse models except for ApcMin/+ mice, as well as in humans, but only on condition that exercise intensity is sufficient (Figure 1 and Table 1). Although concurrent resistance and endurance training is considered the optimal exercise mode in cancer populations, further investigation is needed to understand what intensity and duration of exercise training are ideal to promote an anti-inflammatory profile in cancer patients.
2.3. Mitochondrial Function and Health in Response to Exercise
As observed in many pathologies, mitochondrial function and health declines with CC. Mitochondrial dysfunction includes signalling pathways and mechanisms of mitochondrial biogenesis, fusion and fission [14,101,102], production of reactive oxygen species (ROS) [14,103,104], as well as respiratory function [14,105], all of which influence CC development and progression [5]. Results from our laboratory show that mitochondrial degeneration precedes muscle wasting in tumour-bearing mice [14]. Specifically, increased mitochondrial ROS production (one week after cancer cell implantation), degeneration of the mitochondrial network (two weeks after implantation) and impaired respiratory function (three weeks after implantation) were observed prior to muscle mass loss (4 weeks after implantation) in LLC male mice [14]. These results accompanied by previous findings [101,102] suggest mitochondrial degeneration as a potential key promoter of CC and, as such, mitochondrial quality as a potential therapeutic target for the treatment of CC.
It is well-established that exercise training improves mitochondrial health inducing robust adaptations in both mitochondrial content and quality [106,107]. Specifically, exercise training promotes oxidative phosphorylation and respiratory capacity, promotes mitochondrial biogenesis and decreases ROS emission [106,107,108]. Mitochondrial function and health are also improved with exercise training in patients and rodents with cancer [55,56,58,109,110]. Specifically, concurrent resistance (inclined ladder climbing with gradually increased resistance load) and aerobic (25-min wheel running in 5–9 m/min speed) training (4 days/week, 5.5 weeks) in male C26 tumour-bearing mice may preserve mitochondrial function as indicated by preventing the reduction in muscle succinate dehydrogenase (SDH) activity induced by tumour growth [58]. Although it was not statistically significant, a trend toward increased PGC-1α levels was also observed in the C26 mice that performed concurrent training [58]. Interestingly, significant increases in the protein and gene expression levels of PGC-1α, the master regulator of mitochondrial biogenesis [111], have been noted in C26 and LLC tumour-bearing mice when the exercise intervention (45 min running on treadmill at 14 m/min, 5 days/week for 2–8 weeks) is combined with erythropoietin (EPO) treatment for preventing anaemia [55]. Anaemia is identified in 40% to 80% of patients who have cancer [112] as well as in different cancer mouse models [113] and impairs oxidative metabolism due to hypoxia [114]. A recent study found that 4-week voluntary wheel running effectively counteracted mitochondrial dysfunction preventing muscle mass loss in C26 mice, without compromising running activity [109]; this occurred even though an EPO treatment was not administered. Specifically, the voluntary wheel running attenuated CC-induced mitochondrial abnormalities in tumour-bearing C26 mice attenuating: (i) decreased expression levels of mitochondrial-related proteins (OXPHOS subunit proteins and PGC-1α); (ii) decreased mitochondrial enzyme activity (citrate synthase and cytochrome c oxidase); (iii) decreased expression levels of mitochondrial fusion and fission proteins (Mfn2 and Drp1); (iv) increased mitochondrial oxidative stress; and (v) increased appearance of damaged mitochondria with disrupted cristae structure [109]. In addition, five-week voluntary wheel running increases the protein expression of mitochondrial complex II and IV in male ApcMin/+ mice [96].
The beneficial effect of exercise training on mitochondrial health and function in CC mice has been confirmed by a clinical trial in women with breast cancer during chemotherapy [56]. A 16-week training program (2 days/week), including either resistance training combined with high-intensity interval training or aerobic training combined with high-intensity interval training, increased mitochondrial content in the skeletal muscle of cancer patients, as indicated by the elevated levels of citrate synthase activity [56]. However, compared to controls (cancer patients treated with usual care), increased levels of the electron transport chain proteins (complex I, II and IV) were observed only in patients that performed concurrent aerobic training and high-intensity interval training, but not in patients that performed concurrent resistance and high-intensity interval training [56]. Therefore, the training volume as well as the aerobic nature of the exercise stimulus are two major factors to successfully preserve mitochondrial function in cancer patients undergoing chemotherapy. In conclusion, exercise training improves mitochondrial health and function in skeletal muscle of mice and patients with cancer (Figure 1 and Table 1), with aerobic training being the optimal exercise stimulus that favours mitochondrial adaptations.
2.4. Satellite Cells and Myogenic Regulatory Factors in Response to Exercise
Skeletal muscle satellite cells, as well as their regulators known as myogenic regulatory factors (MRFs; e.g., Myf5, MyoD, myogenin and MRF4) and Pax7 play a role in mechanisms underlying CC [16]. Satellite cells (SC), along with other progenitor populations, become activated and enter a regenerative program in response to cancer-induced skeletal muscle membrane damage in both cancer patients and tumour-bearing mouse models [4]. Muscle membrane damage is reportedly mediated by excessive systemic inflammation from tumours. Due to Pax7 overexpression, myogenic cells are unable to fuse and help alleviate muscle fibre damage [4]. Inaba et al. [115] concluded that the proliferation and differentiation abilities of muscle stem cells derived from the C26 tumour-bearing mice are sustained in vitro. However, inefficient regeneration process in skeletal muscle in C26 tumour-bearing mice, as determined by immunohistological analysis (eMyHC staining), is observed in vivo, potentially due to the cancer-induced decrease of neutrophils, macrophages, and mesenchymal progenitors [115]. While speculative, impaired satellite cell function during CC could also potentially mediate muscle atrophy due to dysregulation of fusion-independent communication to muscle fibres and/or interstitial cells [116,117,118,119]. In addition, chronic doxorubicin (DOX) administration, a highly effective chemotherapeutic agent for treating various types of cancer, reduces SC density in the soleus of ovariectomised female rats [120]. Considering that cancer per se affects satellite cell behaviour, the combination of both the disease and chemotherapies may induce SC dysfunction and exacerbate CC [121].
Exercise training can promote hypertrophy and satellite cell contribution to muscle fibres through SC activation, proliferation and differentiation [122], as well as fusion-independent mechanisms [116,117,118,119]. Long-term [123] or short-term [124,125] exercise training increases the number of SCs in both healthy and diseased populations. Preclinical studies in CC mice also confirm the beneficial effects of exercise training on SC function [126]. Specifically, voluntary wheel running for 19 days normalises cancer-induced dysregulated protein expression of Pax7 to control levels, as well as restores muscle mass by increasing the size of glycolytic muscle fibres in C26 mice [126]. Resistance exercise (ladder climbing with resistance loading increased 10% bi-weekly, 3 repetitions × 5 sets, 3 days/week, 11 weeks) potentially activates myogenic cell activity in C26 mice, as indicated by increased mRNA levels of myogenin [71]. Conversely, aerobic training (60 min wheel running at 5–7 m/min speed, 5 days/week, 11 weeks) does not significantly increase the levels of any myogenic regulatory factor [71].
A clinical trial in female cancer patients treated with chemotherapy concluded the number of SCs does not change in either Type I or Type II muscle fibres after a 10-week concurrent resistance and aerobic training program [78]. However, cautious interpretation of these findings is needed since methodological limitations, including the small heterogeneous sample size of cancer patients, the different chemotherapies used for each patient and the lack of non-exercise control group, are present in this study [78]. Specifically, as a non-exercise control group is not included in this study [78], no change in SC content after exercise training could be potentially interpreted as preservation of SCs, which is a beneficial response. In agreement with the aforementioned results, resistance training (3 days/week for 9–16 weeks) does not change the SC content in either Type I or Type II muscle fibres in patients with prostate cancer undergoing androgen deprivation therapy [127] or in patients with germ cell cancer undergoing cisplatin-based chemotherapy [128]. Similarly to the exercised cancer patients, the SC content in either Type I or Type II muscle fibres does not change in the non-exercise cancer patients (control group) of both clinical trials [127,128]. However, it is worth mentioning that even if the SC content is unchanged, that does not mean the function of the SCs is unchanged as well. Nevertheless, the function of SCs, including SC activation, proliferation and differentiation, was not assessed in the aforementioned studies. In addition, to interpret exercise-induced responses, exercise workload including intensity, duration and frequency, should be considered. It is conceivable the exercise training programs of published trials [78,127,128] may not be sufficiently rigorous to induce significant changes at least at a cellular level.
In contrast to single exercise mode studies, concurrent resistance and high-intensity interval training by cycle ergometry (2 days/week for 16 weeks) increases the SC number per fibre in women with breast cancer during chemotherapy [56]. Also, the increase in SC number is significantly associated with the increase in muscle fibre CSA and muscle strength [56]. Surprisingly, in the same clinical trial no improvement of SC content was found in breast cancer patients either in the group performing 16-week concurrent aerobic and high-intensity interval training or in the usual care group (control group) [56]. Therefore, the improvement of SC content after concurrent resistance and high-intensity interval training in the presence of cancer may reflect an activation of the skeletal muscle repair process to counteract CC [56]. Exercise modality seems to play a critical role on triggering skeletal muscle SC activation. In conclusion, exercise may improve SC function in mice with cancer, but there is no clinical trial which has assessed intrinsic SC function in patients with cancer after performing exercise training. Clinical trials report an increase in the number of SCs only when patients with cancer follow concurrent resistance and high-intensity interval training by cycle ergometry (Figure 1 and Table 1). To better understand if exercise training can ameliorate satellite cell dysfunction in cancer, future studies are needed to assess SC content and SC function, as two distinct factors, that both contribute to the CC development and progression. Further attention should also be paid to the muscle microenvironment (e.g., extracellular matrix composition, inflammatory milieu, behaviour of the muscle fibre, etc.) during CC, as the satellite cell niche strongly influences satellite cell behaviour [129]. In addition, given that SCs not only contribute to muscle repair and growth but also communicate throughout muscle and specifically fibrogenic cells [116,117,118,119], it would be interesting for future studies to investigate if exercise training could decrease cancer-induced fibrosis [130] through SC-mediated mechanisms.
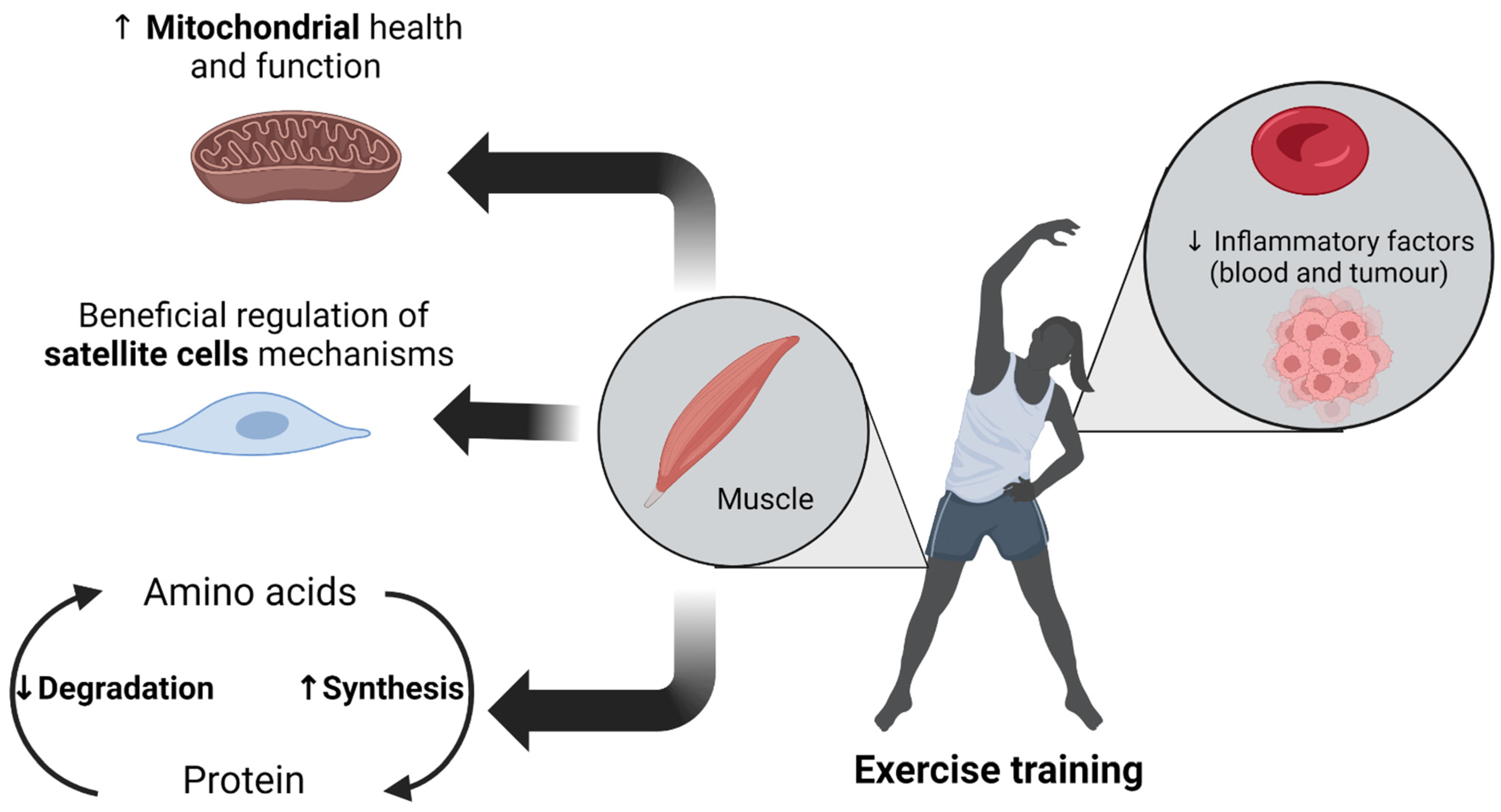
Figure 1.
Beneficial effects of exercise training on counteracting the deleterious mechanisms of cancer cachexia (CC).
Figure 1.
Beneficial effects of exercise training on counteracting the deleterious mechanisms of cancer cachexia (CC).


{kind=link}
{kind=link}
Table 1.
Effects of exercise training on cancer cachexia (CC) mechanisms.
| Protein Turnover | Inflammatory Signalling | Mitochondrial Function | Satellite Cells (SC) and Myogenic Regulatory Factors |
|---|---|---|---|
| ↑ Protein synthesis [30,70,71,78] ↓ Protein degradation: | ↓ Inflammatory markers: IL-6, TNF-α [33,36,92,93,94,99,100] | ↑ Mitochondrial health and function: | ↔ SC content [78,127,128] with resistance or endurance training ↑ SC content with concurrent resistance and high-intensity interval training [56] Beneficial regulation of myogenic regulatory factors: |
Statistically significant increase (↑), decrease (↓) in mice/patients with cancer compared to non-exercising tumour-bearing mice or usual care treated patients with cancer or pre-training baseline results of the exercised patients with cancer. No changes (↔).
3. Contraindications to Exercise in Cancer
Co-existent health problems usually accompany cancer, complicating the way exercise prescription should be applied to cancer patients. Comorbidities are present in around 70% of cancer patients and 35% of these individuals have more than two comorbid conditions. The frequency of comorbidities may decrease or increase based on the type of cancer, age, gender, lifestyle and socioeconomic status of cancer patients [131,132]. The most common comorbidities associated with cancer are anaemia, cardiac dysfunction, chronic fatigue, hypertension, diabetes, asthma and arthritis [53,133,134]. Exercise is considered an unquestionably beneficial intervention to mitigate not only CC but also impairments caused by different comorbidities. However, in case of cancer being accompanied by a comorbidity, components of exercise training workload (i.e., duration, intensity, weekly frequency, rest between sets) should be carefully selected and adjusted based on every individual’s condition, so that any potential exercise-induced adverse effect can be avoided. If a comorbidity compromises the health condition of cancer patient, exercise training should not be performed until addressing the symptoms and/or causes of comorbidity.
As an example, avoidance of exercise should be recommended in anaemic cancer patients until their haemoglobin reaches normal levels [53,135]. Anaemia (haemoglobin level < 12 g/dL) occurs in approximately 40–80% of cancer patients depending on cancer stage and type [112] and may contribute to muscle wasting of cachectic cancer patients [136]. Anaemia is also a major side effect of chemotherapy [137], which may further worsen the already low haemoglobin levels induced by cancer. As mentioned before, in C26 tumour-bearing mice, in which CC is associated with anaemia, treadmill running (45 min at 14 m/min speed, 5 days/week, 8 weeks) does not improve skeletal muscle wasting, but even worsens it when the exercise training is performed for only 2 weeks [55]. However, when haemoglobin levels are corrected through EPO administration, treadmill running (45 min at 14 m/min speed, 5 days/week, 2 weeks) is able to counteract both the oxidative myofiber atrophy and the shift from oxidative to glycolytic fibre type, potentially through stimulating PGC-1α expression [55]. Moreover, in LLC mice where anaemia is more severe compared to C26 mice, exercise training per se (45 min treadmill running at 14 m/min speed, 5 days/week, 4 weeks) can prevent muscle strength loss. A combination of exercise training with EPO administration can additionally promote muscle oxidative capacity and intracellular ATP content, alleviate muscle wasting, and prevent the onset of mitochondrial ultrastructural alterations [55]. These results [55] are meaningful to highlight the major role of preserving haemoglobin concentration close to normal levels to prevent CC through exercise training.
The administration of EPO in cancer patients with anaemia revolutionised the treatment of this complication around 20 years ago with impressive results from a haematological perspective while simultaneously avoiding the adverse effects of blood transfusions in anaemic patients with cancer [138,139]. However, later scientific evidence shows that EPO treatment in cancer patients mediates a variety of serious adverse effects, including survival decrease [140], acceleration of tumour progression [141] and an increase in the incidence of thromboembolic events [142]. For this reason, utilisation of EPO in cancer patients has been restricted, especially among patients treated with a curative intent and in patients with advanced tumours but long-term survival expectations [143]. Iron administration (oral or intravenous) is now considered a promising alternative to normalise haemoglobin levels in anaemic cancer patients [144,145].
Heart function is also impaired in tumour-bearing mice and rats (C26 carcinoma tumour-bearing mice [146,147,148], LLC tumour-bearing mice [149], breast tumour-bearing mice [150] and liver tumour-bearing rats [151]). Also, a series of epidemiological studies conclude that there is a comorbidity of cancer and cardiovascular diseases [152,153]. Many cancer therapies (e.g., anthracyclines, HER-2-targeted agents, mitotic inhibitors, immune modulators, radiation etc.) induce cardiac toxicity and lead to long-term and most likely permanent myocardial injury [154,155]. Considering that a remarkable percentage of cancer patients additionally suffer from heart failure [156,157], further precautions should be taken when exercise is performed in these populations. Any potential adverse effect of exercise in cancer patients with a comorbidity of heart failure could be prevented by following the exercise training guidelines specific to patients with heart failure [158]. Also, in case of immunosuppressed or bone marrow transplanted cancer patients, exercise training should be avoided in public gyms until their white blood cell count returns to normal levels [135]. In addition, due to the additional energetic demands caused by the tumour, especially when cachexia has developed, appropriate nutritional support is required for patients with cancer perform exercise training [159].
Exercise training, even if it is high intensity, is considered safe and effective in cancer patients either shortly after completion of primary cancer treatment [160] or during cancer treatment [161], but on condition that no medical contraindications are present [161]. In case of cancer patients presenting comorbidities, exercise training should be performed only under specific guidelines, precautions and supervision by specialists. Future preclinical exercise studies should consider the limitations of human exercise oncology studies related to cancer- and treatment-induced complications; and design exercise experiments in preclinical models addressing simultaneously other comorbidities which may present [162].
4. Effects of Different Exercise Modalities (Resistance, Endurance and Concurrent Training) in Cancer Cachexia (CC)
Even though exercise training is considered a therapeutic and preventive approach for many diseases, special attention should be paid to exercise modality based on the clinical goal of the exercise intervention. Exercise modality plays a major role on specifying exercise-induced physiological adaptations and stimulating particular biological systems (e.g., cardiorespiratory, neuromuscular). Specifically, in healthy populations resistance training promotes muscle hypertrophy and muscle strength/power increase [20]. Endurance training mainly induces cardiovascular and respiratory adaptations improving respiratory and heart function [19], but also increases skeletal muscle capillary density [163,164], oxidative capacity [165,166] and mitochondria content and function [164,166]. Paradoxically, endurance training can also produce skeletal muscle hypertrophy but mainly in sedentary individuals or populations with muscle wasting, such as the elderly [167]. Concurrent training can stimulate both aerobic and anaerobic metabolic pathways, combining the beneficial adaptations induced by both resistance and endurance training [168]. Interestingly, concurrent training can be also more hypertrophic in untrained/sedentary populations compared to athletes or well-trained populations [169].
In preclinical cancer studies, as elaborated in aforementioned sections, exercise training protocols consisting of resistance training per se [30,57,71], endurance training per se [31,32,33,71,76,77,92,96,109,126] or concurrent resistance and aerobic training [58] induce beneficial adaptations in tumour-bearing mice (C26, LLC, ApcMin/+, AOM, breast-tumour bearing mice). In addition, regardless of modality, exercise training positively affects the quality of life of cancer patients [170,171]. Resistance training improves muscle strength of the upper and lower body, increases lean body mass, decreases percentage of body fat and counteracts cancer-induced increases in inflammatory factors without eliciting adverse effects in cancer patients [99,172,173]. Endurance training is also a safe and efficient method to improve aerobic capacity in cancer patients through preserving skeletal muscle mitochondrial function [56], promoting antioxidant defence system [174] and reducing fatigue symptomatology [175]. Although the majority of clinical trials emphasise the beneficial role of exercise training in cancer patients, given that specific precautions are taken (e.g., addressing comorbidities [161], individualising exercise programs [159], covering additional energetic demands [159]); a recent meta-analysis highlighted the inconsistency of data in clinical trials about the effectiveness of exercise in patients with cancer [50]. These inconsistencies can be explained by limitations (i.e., heterogeneity of participants in cancer diagnosis, cancer stages and symptoms) of the clinical trials [50].
A great number of clinical trials, as well as the guidelines by American College of Sports Medicine [59], propose a combination of both resistance and endurance training in cancer patients, since concurrent training can induce wide-ranging physical adaptations promoting both aerobic (VO2max and resistance to fatigue) and anaerobic (muscle strength and function) components simultaneously without any adverse effect in cancer patients [60,176,177,178]. The phenotypic improvements of concurrent training in patients with cancer have been confirmed at the molecular level as well. Based on previous human studies, concurrent resistance and endurance training prevents cancer- and chemotherapy-induced disruptions in protein degradation and protein synthesis [78], decreases circulating inflammatory markers [98,100] and increases mitochondrial and SC content [56]. From a molecular perspective, the advantage of concurrent training in cancer patients may be related to the determinant role of mitochondria in CC. Specifically, given that cancer-induced mitochondrial impairments may lead to decrements in protein synthesis and elevated protein degradation [179], then endurance training may maximise the effects of resistance training by normalising the cancer-induced unbalanced protein turnover through improving mitochondrial health and function.
Considering the findings of exercise studies in both animals and humans with cancer, concurrent resistance and endurance training could be considered an optimal exercise modality to successfully elicit improvements in total muscle health/quality. The characteristics of concurrent training may be also crucial to produce the intended results in improving or preserving skeletal muscle mass in patients with CC. Based on literature in healthy populations, to maximise the hypertrophic effects of concurrent training, endurance and resistance elements should be separated 6–24 h [169] and if this is not feasible, the resistance exercise bout should precede the endurance exercise bout within the same session [180,181]. The endurance element should have an interval form including high-intensity bouts, ideally cycling [168,169,180,181]. More preclinical studies in tumour-bearing mice are needed to clarify the molecular and physiological mechanisms contributing to the anaerobic and aerobic improvements induced by concurrent resistance and endurance training in patients and animals with cancer. A novel, very promising and well representative model of human concurrent training, called “PoWeR”, has been recently introduced to mouse studies [182,183,184]. Briefly, “PoWeR” training consists of voluntary wheel running with progressively increased loading (from 2 g to 6 g of weight) and induces robust cardiac adaptations and skeletal muscle hypertrophy [182,183,184]. Generally, voluntary wheel running is considered a viable preclinical exercise framework that can account for the fatiguing aspects of cancer and/or chemotherapy, closing the gap between preclinical and clinical oncology studies [162]. Therefore, future preclinical studies could use the PoWeR training as a translatable murine model of concurrent training in tumour-bearing mice. An alternative to concurrent training for preclinical studies with vulnerable tumour-bearing mice could be the combination of voluntary wheel running with a novel voluntary weightlifting model for mice, which elicits squat-like activities against adjustable load during feeding [185].
To conclude, concurrent resistance and endurance training is suggested as the most beneficial modality of exercise for patients with cancer. Nevertheless, further investigation using a translatable concurrent exercise training in cancer mouse models is needed to better understand the physiological and molecular responses in cancer cachectic skeletal muscle after performing concurrent training.
5. Biological Sex Differences in Response to Exercise in Cancer Cachexia (CC)
Both clinical and preclinical studies in the CC field suggest biological sex differences affect muscle wasting development and progression in cancer patients and cancer mouse models [5,61,186]. Specifically, it is well established that male cancer patients present a higher prevalence of cachexia, more severe muscle mass and weight loss and greater reduction of muscle strength compared to female cancer patients [61]. In addition, animal studies have suggested the cellular and molecular mechanisms contributing to CC, as well as the pace of CC development and progression, are different between male and female cachectic mice [15,61,187]. Specifically, male ApcMin/+ mice are sensitive to inflammation (IL-6)-mediated cachexia [6], while female ApcMin/+ mice undergo cachexia progression IL-6-independently [187]. Similarly, female LLC tumour-bearing mice present multiple protections in both metabolic and contractile skeletal muscle function in the early stages of tumour development [15], while male LLC tumour-bearing mice present many metabolic perturbations before the onset of cachexia [7,14]. Therefore, female tumour-bearing mice may have a stronger defence system protecting them from tumour-induced muscle degeneration [15].
Generally speaking, female muscles are more fatigue-resistant and appear to exhibit enhanced mitochondrial quality compared to male muscles [61,62]. However, males exhibit greater SC content [188] and increased mRNA expression of MyoD and myogenin [189], two factors associated with SC proliferation and differentiation and potentially skeletal muscle hypertrophic and regenerative potential. Considering the biological sex dissimilarities observed in the skeletal muscle, different skeletal muscle adaptations in response to a metabolic stimulus, such as exercise training, may be induced in males and females. Indeed, a previous human study based on young and healthy adults concluded that males present greater muscle protein synthesis and mitochondrial biogenesis than females in response to 3-weeks of sprint interval training [190]. However, when low-to-moderate continuous endurance training (cycling at 60% of VO2max, 5 days/week, 7 weeks) is performed, no difference is observed in skeletal muscle enzyme oxidative potential between males and females [191]. Furthermore, acute high-intensity resistance training combined with post-exercise whey protein ingestion increases myofibrillar protein synthesis and p70S6K1 phosphorylation similarly in males and females, although the postexercise testosterone responses in females are 45-fold lower than males [192]. Conversely, compared to male, female skeletal muscle is more resistant to fatigue for a task of the same relative intensity [62,193,194]. Females present a lower degree of contractile impairment and faster rate of recovery following exercise with intermittent isometric knee extensions [194].
To our knowledge, no study has directly investigated biological sex differences in response to exercise in cancer patients or tumour-bearing mice. However, an indirect and observational comparison between preclinical studies with different biological sex suggests exercise-induced adaptations in skeletal muscle are similar in female and male tumour-bearing mice [57,64]. Specifically, a representative resistance training for animals involving lower limb contractions through high-frequency electric stimulation for 2-weeks, increases weight and size of Type IIa and IIb fibres in the TA muscle, improves myofibrillar protein synthesis, as well as attenuates cachexia-induced AMPK activity and reduction of skeletal muscle oxidative capacity similarly in ApcMin/+ female [64] and male ApcMin/+ mice [57]. In addition, 2-week resistance training in the form of high-frequency electric stimulation increases EDL weight and protein content in female C26 mice [195]. Although no study has investigated the effects of resistance training per se in male C26 mice, previous studies have shown that even endurance training per se or concurrent endurance and resistance training can induce beneficial skeletal muscle adaptations in male C26 mice preventing cancer-induced muscle wasting and loss of muscle strength [58,76]. Also, in both male [76] and female [32] C26 mice physical activity via wheel running suppresses the protein expression of the autophagic markers LC3B and p62.
The beneficial effects of exercise training in cancer patients appear similar between males and females [196]. Specifically, clinical studies that recruited either female [197,198] or male [199,200] cancer patients suggest exercise is an efficient therapeutic intervention for preserving muscle wasting and muscle strength loss. However, similarly with preclinical cancer studies, no clinical study has directly investigated biological sex differences in exercise-induced adaptations in cancer patients. Future preclinical exercise studies in CC mouse models should be focused on assessing potential biological sex differences in skeletal muscle adaptations in response to exercise training. Discovering potential differences in exercise-induced skeletal muscle adaptations between male and female tumour-bearing mice could guide personalised exercise training protocols based on biological sex, thereby optimising the effects of exercise training in both male and female cancer patients.
6. Conclusions
There is now a large body of evidence suggesting exercise training as a beneficial therapeutic approach for decelerating the development and progression of cancer-induced atrophy. Preclinical and clinical studies have shown that exercise training, regardless of modality, beneficially regulates the molecular and cellular mechanisms that contribute to CC pathology. Protein turnover, inflammatory signalling, mitochondrial function and satellite cell number can be advantageously controlled by exercise training in animals and patients with cancer. Given the majority of cancer patients present at least one comorbidity, exercise training could elicit adverse events in these vulnerable individuals. For this reason, meticulous precautions (e.g., low-intensity exercise training protocols under constant supervision by specialists) should be taken while exercise is performed. In many instances, a complete address of comorbidities (e.g., haemoglobin back or close to the normal levels) may be essential before cancer patients begin an exercise training program. Fatigue of cancer patients related to either comorbidities (e.g., anaemia) or radiation therapies/chemotherapies should be also taken into account for the exercise training program prescription to avoid exercise adverse effects in these vulnerable patients.
Based on findings from preclinical and clinicals studies, as well as the guidelines of American College of Sports Medicine for cancer patients, concurrent resistance and endurance training is considered the most advantageous exercise modality for cancer patients. Concurrent training simultaneously improves elements of both aerobic and anaerobic metabolism. Nevertheless, further investigation is needed to understand the molecular mechanisms activated through concurrent training and contributing to improving skeletal muscle health of patients with CC. A novel voluntary wheel running protocol for mice, called PoWeR training [182,183,184], could be a representative exercise mode of concurrent resistance and endurance training. Generally, voluntary wheel running in cancer mice is considered a translatable exercise mode since it accounts for the fatiguing aspects induced by comorbidities or side effects of cancer treatments. Consequently, future preclinical studies using the PoWeR exercise training protocol in cancer mice could explain many aspects of mechanisms underlying the deceleration of cancer-induced atrophy after performing a concurrent training protocol, which is appropriate even if comorbidities are presented along with cancer.
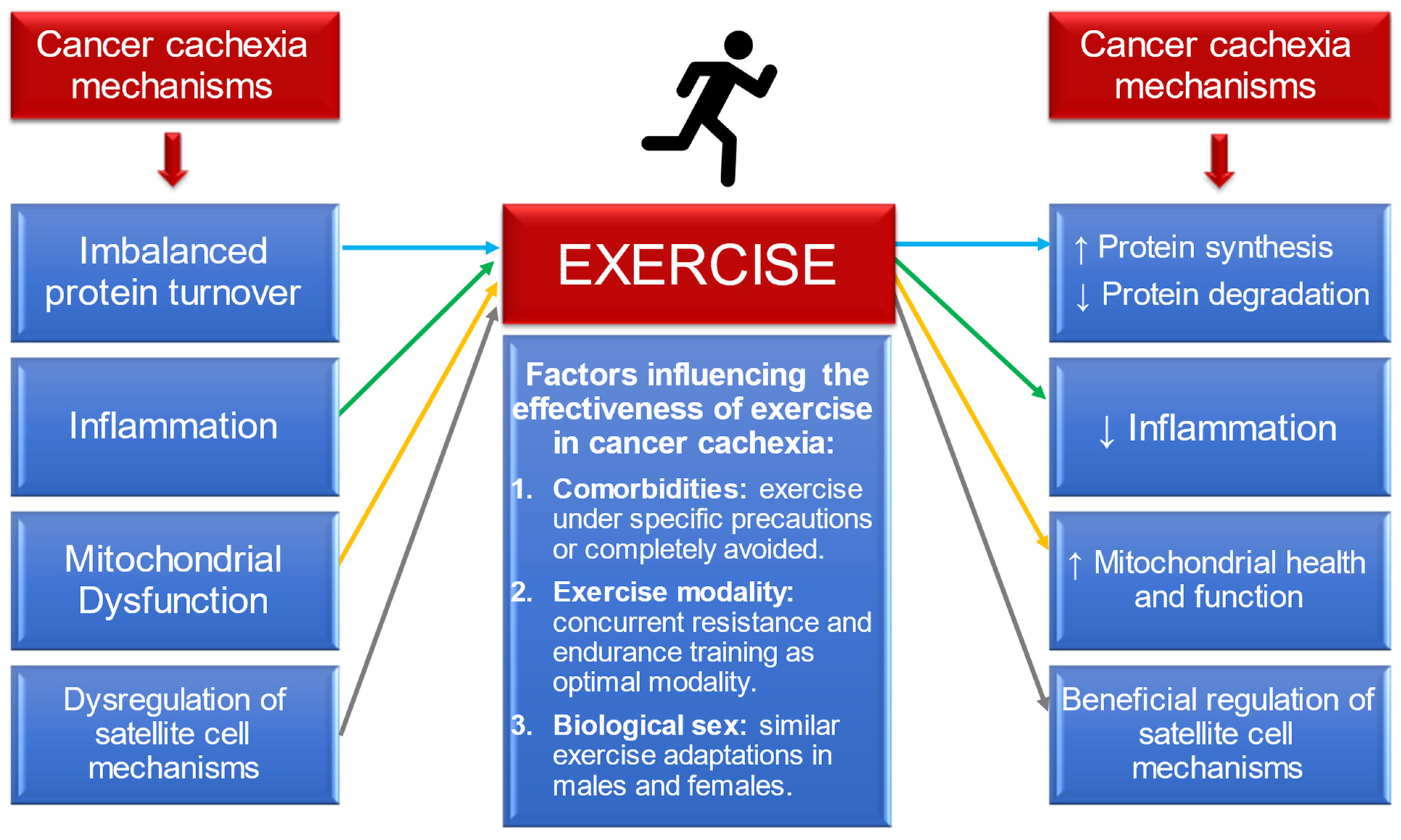
Finally, when comparing observationally the beneficial exercise-induced adaptations between male and female patients or mice with cancer, both biological sexes seem to respond similarly to exercise stimuli. However, neither preclinical nor clinical studies in the cancer field have investigated directly the potential biological sex differences on the skeletal muscle in response to exercise training in the tumour-bearing state. Given that dissimilarities are observed in CC development and progression between females and males, future investigation is needed to understand if the biological sex in cancer patients plays a crucial role on the exercise-induced adaptations. The Figure 2 summarises all of the aforementioned conclusions of this review.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/cancers14102512/s1, Table S1: Ongoing clinical trials with exercise-based interventions in cancer patients (source: https://clinicaltrials.gov).
Author Contributions
S.T. and N.P.G. conceived and designed research; S.T. drafted manuscript; S.T., K.A.M., T.A.W. and N.P.G. edited and revised manuscript; S.T., K.A.M., T.A.W. and N.P.G. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Institute of Arthritis and Musculoskeletal and Skin Diseases of the National Institutes of Health Award No. R01AR075794-01A1/AR/NIAMS (to N.P.G).
Acknowledgments
Figure 1 was generated using BioRender.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Argiles, J.M.; Busquets, S.; Stemmler, B.; Lopez-Soriano, F.J. Cancer cachexia: Understanding the molecular basis. Nat. Rev. Cancer 2014, 14, 754–762. [Google Scholar] [CrossRef]
- Hovell, M.F.; Black, D.R. Minimal intervention and arthritis treatment: Implications for patient and physician compliance. Arthritis Care Res. 1989, 2, S65–S70. [Google Scholar] [CrossRef] [PubMed]
- Fearon, K.; Strasser, F.; Anker, S.D.; Bosaeus, I.; Bruera, E.; Fainsinger, R.L.; Jatoi, A.; Loprinzi, C.; MacDonald, N.; Mantovani, G.; et al. Definition and classification of cancer cachexia: An international consensus. Lancet Oncol. 2011, 12, 489–495. [Google Scholar] [CrossRef]
- He, W.A.; Berardi, E.; Cardillo, V.M.; Acharyya, S.; Aulino, P.; Thomas-Ahner, J.; Wang, J.; Bloomston, M.; Muscarella, P.; Nau, P.; et al. NF-kappaB-mediated Pax7 dysregulation in the muscle microenvironment promotes cancer cachexia. J. Clin. Investig. 2013, 123, 4821–4835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa-Caldwell, M.E.; Fix, D.K.; Washington, T.A.; Greene, N.P. Muscle alterations in the development and progression of cancer-induced muscle atrophy: A review. J. Appl. Physiol. 2020, 128, 25–41. [Google Scholar] [CrossRef] [PubMed]
- White, J.P.; Baynes, J.W.; Welle, S.L.; Kostek, M.C.; Matesic, L.E.; Sato, S.; Carson, J.A. The regulation of skeletal muscle protein turnover during the progression of cancer cachexia in the Apc(Min/+) mouse. PLoS ONE 2011, 6, e24650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.L.; Lee, D.E.; Rosa-Caldwell, M.E.; Brown, L.A.; Perry, R.A.; Haynie, W.S.; Huseman, K.; Sataranatarajan, K.; Van Remmen, H.; Washington, T.A.; et al. Protein imbalance in the development of skeletal muscle wasting in tumour-bearing mice. J. Cachexia Sarcopenia Muscle 2018, 9, 987–1002. [Google Scholar] [CrossRef]
- Baracos, V.E. Regulation of skeletal-muscle-protein turnover in cancer-associated cachexia. Nutrition 2000, 16, 1015–1018. [Google Scholar] [CrossRef]
- DeJong, C.H.; Busquets, S.; Moses, A.G.; Schrauwen, P.; Ross, J.A.; Argiles, J.M.; Fearon, K.C. Systemic inflammation correlates with increased expression of skeletal muscle ubiquitin but not uncoupling proteins in cancer cachexia. Oncol. Rep. 2005, 14, 257–263. [Google Scholar]
- Williams, J.P.; Phillips, B.E.; Smith, K.; Atherton, P.J.; Rankin, D.; Selby, A.L.; Liptrot, S.; Lund, J.; Larvin, M.; Rennie, M.J. Effect of tumor burden and subsequent surgical resection on skeletal muscle mass and protein turnover in colorectal cancer patients. Am. J. Clin. Nutr. 2012, 96, 1064–1070. [Google Scholar] [CrossRef] [Green Version]
- Onesti, J.K.; Guttridge, D.C. Inflammation based regulation of cancer cachexia. BioMed Res. Int. 2014, 2014, 168407. [Google Scholar] [CrossRef] [PubMed]
- Narsale, A.A.; Carson, J.A. Role of interleukin-6 in cachexia: Therapeutic implications. Curr. Opin. Support. Palliat. Care 2014, 8, 321–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catalano, M.G.; Fortunati, N.; Arena, K.; Costelli, P.; Aragno, M.; Danni, O.; Boccuzzi, G. Selective up-regulation of tumor necrosis factor receptor I in tumor-bearing rats with cancer-related cachexia. Int. J. Oncol. 2003, 23, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.L.; Rosa-Caldwell, M.E.; Lee, D.E.; Blackwell, T.A.; Brown, L.A.; Perry, R.A.; Haynie, W.S.; Hardee, J.P.; Carson, J.A.; Wiggs, M.P.; et al. Mitochondrial degeneration precedes the development of muscle atrophy in progression of cancer cachexia in tumour-bearing mice. J. Cachexia Sarcopenia Muscle 2017, 8, 926–938. [Google Scholar] [CrossRef]
- Lim, S.; Deaver, J.W.; Rosa-Caldwell, M.E.; Haynie, W.S.; Morena da Silva, F.; Cabrera, A.R.; Schrems, E.R.; Saling, L.W.; Jansen, L.T.; Dunlap, K.R.; et al. Development of metabolic and contractile alterations in development of cancer cachexia in female tumor-bearing mice. J. Appl. Physiol. 2022, 132, 58–72. [Google Scholar] [CrossRef]
- Bossola, M.; Marzetti, E.; Rosa, F.; Pacelli, F. Skeletal muscle regeneration in cancer cachexia. Clin. Exp. Pharmacol. Physiol. 2016, 43, 522–527. [Google Scholar] [CrossRef] [Green Version]
- Argiles, J.M.; Lopez-Soriano, F.J.; Stemmler, B.; Busquets, S. Therapeutic strategies against cancer cachexia. Eur. J. Transl. Myol. 2019, 29, 7960. [Google Scholar] [CrossRef] [Green Version]
- Bourdel-Marchasson, I.; Blanc-Bisson, C.; Doussau, A.; Germain, C.; Blanc, J.F.; Dauba, J.; Lahmar, C.; Terrebonne, E.; Lecaille, C.; Ceccaldi, J.; et al. Nutritional advice in older patients at risk of malnutrition during treatment for chemotherapy: A two-year randomized controlled trial. PLoS ONE 2014, 9, e108687. [Google Scholar] [CrossRef]
- Lin, X.; Zhang, X.; Guo, J.; Roberts, C.K.; McKenzie, S.; Wu, W.C.; Liu, S.; Song, Y. Effects of Exercise Training on Cardiorespiratory Fitness and Biomarkers of Cardiometabolic Health: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. J. Am. Heart Assoc. 2015, 4, e002014. [Google Scholar] [CrossRef] [Green Version]
- Ahtiainen, J.P.; Pakarinen, A.; Alen, M.; Kraemer, W.J.; Hakkinen, K. Muscle hypertrophy, hormonal adaptations and strength development during strength training in strength-trained and untrained men. Eur. J. Appl. Physiol. 2003, 89, 555–563. [Google Scholar] [CrossRef]
- Scully, D.; Kremer, J.; Meade, M.M.; Graham, R.; Dudgeon, K. Physical exercise and psychological well being: A critical review. Br. J. Sports Med. 1998, 32, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Farup, J.; Madaro, L.; Puri, P.L.; Mikkelsen, U.R. Interactions between muscle stem cells, mesenchymal-derived cells and immune cells in muscle homeostasis, regeneration and disease. Cell Death Dis. 2015, 6, e1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaczmarek, A.; Kaczmarek, M.; Cialowicz, M.; Clemente, F.M.; Wolanski, P.; Badicu, G.; Murawska-Cialowicz, E. The Role of Satellite Cells in Skeletal Muscle Regeneration-The Effect of Exercise and Age. Biology 2021, 10, 1056. [Google Scholar] [CrossRef] [PubMed]
- Mesquita, P.H.C.; Vann, C.G.; Phillips, S.M.; McKendry, J.; Young, K.C.; Kavazis, A.N.; Roberts, M.D. Skeletal Muscle Ribosome and Mitochondrial Biogenesis in Response to Different Exercise Training Modalities. Front. Physiol. 2021, 12, 725866. [Google Scholar] [CrossRef]
- Thompson, D.; Markovitch, D.; Betts, J.A.; Mazzatti, D.; Turner, J.; Tyrrell, R.M. Time course of changes in inflammatory markers during a 6-mo exercise intervention in sedentary middle-aged men: A randomized-controlled trial. J. Appl. Physiol. 2010, 108, 769–779. [Google Scholar] [CrossRef] [Green Version]
- Courneya, K.S.; Mackey, J.R.; Jones, L.W. Coping with cancer: Can exercise help? Phys. Sportsmed. 2000, 28, 49–73. [Google Scholar] [CrossRef]
- Lira, F.S.; Neto, J.C.; Seelaender, M. Exercise training as treatment in cancer cachexia. Appl. Physiol. Nutr. Metab. 2014, 39, 679–686. [Google Scholar] [CrossRef]
- Rosa-Neto, J.C.; Silveira, L.S. Endurance Exercise Mitigates Immunometabolic Adipose Tissue Disturbances in Cancer and Obesity. Int. J. Mol. Sci. 2020, 21, 9745. [Google Scholar] [CrossRef]
- Murphy, E.A.; Enos, R.T.; Velazquez, K.T. Influence of Exercise on Inflammation in Cancer: Direct Effect or Innocent Bystander? Exerc. Sport Sci. Rev. 2015, 43, 134–142. [Google Scholar] [CrossRef]
- Hardee, J.P.; Fix, D.K.; Koh, H.J.; Wang, X.; Goldsmith, E.C.; Carson, J.A. Repeated eccentric contractions positively regulate muscle oxidative metabolism and protein synthesis during cancer cachexia in mice. J. Appl. Physiol. 2020, 128, 1666–1676. [Google Scholar] [CrossRef]
- Puppa, M.J.; White, J.P.; Velazquez, K.T.; Baltgalvis, K.A.; Sato, S.; Baynes, J.W.; Carson, J.A. The effect of exercise on IL-6-induced cachexia in the Apc (Min/+) mouse. J. Cachexia Sarcopenia Muscle 2012, 3, 117–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pigna, E.; Berardi, E.; Aulino, P.; Rizzuto, E.; Zampieri, S.; Carraro, U.; Kern, H.; Merigliano, S.; Gruppo, M.; Mericskay, M.; et al. Aerobic Exercise and Pharmacological Treatments Counteract Cachexia by Modulating Autophagy in Colon Cancer. Sci. Rep. 2016, 6, 26991. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.A.; Davis, J.M.; Barrilleaux, T.L.; McClellan, J.L.; Steiner, J.L.; Carmichael, M.D.; Pena, M.M.; Hebert, J.R.; Green, J.E. Benefits of exercise training on breast cancer progression and inflammation in C3(1)SV40Tag mice. Cytokine 2011, 55, 274–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wennerberg, E.; Lhuillier, C.; Rybstein, M.D.; Dannenberg, K.; Rudqvist, N.P.; Koelwyn, G.J.; Jones, L.W.; Demaria, S. Exercise reduces immune suppression and breast cancer progression in a preclinical model. Oncotarget 2020, 11, 452–461. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.K.; Kim, Y.; Park, S.; Kim, E.; Kim, Y.; Kim, Y.; Kim, J.H. Effects of Steady Low-Intensity Exercise on High-Fat Diet Stimulated Breast Cancer Progression Via the Alteration of Macrophage Polarization. Integr. Cancer Ther. 2020, 19, 1534735420949678. [Google Scholar] [CrossRef]
- Mader, T.; Chaillou, T.; Alves, E.S.; Jude, B.; Cheng, A.J.; Kenne, E.; Mijwel, S.; Kurzejamska, E.; Vincent, C.T.; Rundqvist, H.; et al. Exercise reduces intramuscular stress and counteracts muscle weakness in mice with breast cancer. J. Cachexia Sarcopenia Muscle 2022, 13, 1151–1163. [Google Scholar] [CrossRef]
- Yu, M.; King, B.; Ewert, E.; Su, X.; Mardiyati, N.; Zhao, Z.; Wang, W. Exercise Activates p53 and Negatively Regulates IGF-1 Pathway in Epidermis within a Skin Cancer Model. PLoS ONE 2016, 11, e0160939. [Google Scholar] [CrossRef] [Green Version]
- Lira, F.S.; Yamashita, A.; Carnevali, L.C., Jr.; Goncalves, D.C.; Lima, W.P.; Rosa, J.C.; Caperuto, E.C.; Rosa, L.F.; Seelaender, M. Exercise training reduces PGE2 levels and induces recovery from steatosis in tumor-bearing rats. Horm. Metab. Res. 2010, 42, 944–949. [Google Scholar] [CrossRef] [Green Version]
- Donatto, F.F.; Neves, R.X.; Rosa, F.O.; Camargo, R.G.; Ribeiro, H.; Matos-Neto, E.M.; Seelaender, M. Resistance exercise modulates lipid plasma profile and cytokine content in the adipose tissue of tumour-bearing rats. Cytokine 2013, 61, 426–432. [Google Scholar] [CrossRef] [Green Version]
- Lira, F.S.; Tavares, F.L.; Yamashita, A.S.; Koyama, C.H.; Alves, M.J.; Caperuto, E.C.; Batista, M.L., Jr.; Seelaender, M. Effect of endurance training upon lipid metabolism in the liver of cachectic tumour-bearing rats. Cell. Biochem. Funct. 2008, 26, 701–708. [Google Scholar] [CrossRef]
- Pedersen, L.; Christensen, J.F.; Hojman, P. Effects of exercise on tumor physiology and metabolism. Cancer J. 2015, 21, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Niels, T.; Tomanek, A.; Freitag, N.; Schumann, M. Can Exercise Counteract Cancer Cachexia? A Systematic Literature Review and Meta-Analysis. Integr. Cancer Ther. 2020, 19, 1534735420940414. [Google Scholar] [CrossRef] [PubMed]
- Vermaete, N.; Wolter, P.; Verhoef, G.; Gosselink, R. Physical activity, physical fitness and the effect of exercise training interventions in lymphoma patients: A systematic review. Ann. Hematol. 2013, 92, 1007–1021. [Google Scholar] [CrossRef] [PubMed]
- Samuel, S.R.; Maiya, A.G.; Fernandes, D.J.; Guddattu, V.; Saxena, P.U.P.; Kurian, J.R.; Lin, P.J.; Mustian, K.M. Effectiveness of exercise-based rehabilitation on functional capacity and quality of life in head and neck cancer patients receiving chemo-radiotherapy. Support. Care Cancer 2019, 27, 3913–3920. [Google Scholar] [CrossRef]
- Fisher, H.M.; Jacobs, J.M.; Taub, C.J.; Lechner, S.C.; Lewis, J.E.; Carver, C.S.; Blomberg, B.B.; Antoni, M.H. How changes in physical activity relate to fatigue interference, mood, and quality of life during treatment for non-metastatic breast cancer. Gen. Hosp. Psychiatry 2017, 49, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Wochner, R.; Clauss, D.; Nattenmuller, J.; Tjaden, C.; Bruckner, T.; Kauczor, H.U.; Hackert, T.; Wiskemann, J.; Steindorf, K. Impact of progressive resistance training on CT quantified muscle and adipose tissue compartments in pancreatic cancer patients. PLoS ONE 2020, 15, e0242785. [Google Scholar] [CrossRef]
- Vermillion, S.A.; James, A.; Dorrell, R.D.; Brubaker, P.; Mihalko, S.L.; Hill, A.R.; Clark, C.J. Preoperative exercise therapy for gastrointestinal cancer patients: A systematic review. Syst. Rev. 2018, 7, 103. [Google Scholar] [CrossRef]
- Singh, F.; Newton, R.U.; Galvao, D.A.; Spry, N.; Baker, M.K. A systematic review of pre-surgical exercise intervention studies with cancer patients. Surg. Oncol. 2013, 22, 92–104. [Google Scholar] [CrossRef]
- Ligibel, J.A.; Dillon, D.; Giobbie-Hurder, A.; McTiernan, A.; Frank, E.; Cornwell, M.; Pun, M.; Campbell, N.; Dowling, R.J.O.; Chang, M.C.; et al. Impact of a Pre-Operative Exercise Intervention on Breast Cancer Proliferation and Gene Expression: Results from the Pre-Operative Health and Body (PreHAB) Study. Clin. Cancer Res. 2019, 25, 5398–5406. [Google Scholar] [CrossRef] [Green Version]
- Grande, A.J.; Silva, V.; Sawaris Neto, L.; Teixeira Basmage, J.P.; Peccin, M.S.; Maddocks, M. Exercise for cancer cachexia in adults. Cochrane Database Syst. Rev. 2021, 3, CD010804. [Google Scholar] [CrossRef]
- Leal, L.G.; Lopes, M.A.; Peres, S.B.; Batista, M.L., Jr. Exercise Training as Therapeutic Approach in Cancer Cachexia: A Review of Potential Anti-inflammatory Effect on Muscle Wasting. Front. Physiol. 2020, 11, 570170. [Google Scholar] [CrossRef] [PubMed]
- Aversa, Z.; Costelli, P.; Muscaritoli, M. Cancer-induced muscle wasting: Latest findings in prevention and treatment. Ther. Adv. Med. Oncol. 2017, 9, 369–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argiles, J.M.; Busquets, S.; Lopez-Soriano, F.J.; Costelli, P.; Penna, F. Are there any benefits of exercise training in cancer cachexia? J. Cachexia Sarcopenia Muscle 2012, 3, 73–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brownstein, C.G.; Twomey, R.; Temesi, J.; Medysky, M.E.; Culos-Reed, S.N.; Millet, G.Y. Mechanisms of Neuromuscular Fatigability in People with Cancer-related Fatigue. Med. Sci. Sports Exerc. 2022. ahead of print. [Google Scholar] [CrossRef]
- Pin, F.; Busquets, S.; Toledo, M.; Camperi, A.; Lopez-Soriano, F.J.; Costelli, P.; Argiles, J.M.; Penna, F. Combination of exercise training and erythropoietin prevents cancer-induced muscle alterations. Oncotarget 2015, 6, 43202–43215. [Google Scholar] [CrossRef]
- Mijwel, S.; Cardinale, D.A.; Norrbom, J.; Chapman, M.; Ivarsson, N.; Wengstrom, Y.; Sundberg, C.J.; Rundqvist, H. Exercise training during chemotherapy preserves skeletal muscle fiber area, capillarization, and mitochondrial content in patients with breast cancer. FASEB J. 2018, 32, 5495–5505. [Google Scholar] [CrossRef] [Green Version]
- Hardee, J.P.; Mangum, J.E.; Gao, S.; Sato, S.; Hetzler, K.L.; Puppa, M.J.; Fix, D.K.; Carson, J.A. Eccentric contraction-induced myofiber growth in tumor-bearing mice. J. Appl. Physiol. 2016, 120, 29–37. [Google Scholar] [CrossRef]
- Ranjbar, K.; Ballaro, R.; Bover, Q.; Pin, F.; Beltra, M.; Penna, F.; Costelli, P. Combined Exercise Training Positively Affects Muscle Wasting in Tumor-Bearing Mice. Med. Sci. Sports Exerc. 2019, 51, 1387–1395. [Google Scholar] [CrossRef]
- Wolin, K.Y.; Schwartz, A.L.; Matthews, C.E.; Courneya, K.S.; Schmitz, K.H. Implementing the exercise guidelines for cancer survivors. J. Support. Oncol. 2012, 10, 171–177. [Google Scholar] [CrossRef] [Green Version]
- De Luca, V.; Minganti, C.; Borrione, P.; Grazioli, E.; Cerulli, C.; Guerra, E.; Bonifacino, A.; Parisi, A. Effects of concurrent aerobic and strength training on breast cancer survivors: A pilot study. Public Health 2016, 136, 126–132. [Google Scholar] [CrossRef]
- Zhong, X.; Zimmers, T.A. Sex Differences in Cancer Cachexia. Curr. Osteoporos. Rep. 2020, 18, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Ansdell, P.; Thomas, K.; Hicks, K.M.; Hunter, S.K.; Howatson, G.; Goodall, S. Physiological sex differences affect the integrative response to exercise: Acute and chronic implications. Exp. Physiol. 2020, 105, 2007–2021. [Google Scholar] [CrossRef] [PubMed]
- Sewright, K.A.; Hubal, M.J.; Kearns, A.; Holbrook, M.T.; Clarkson, P.M. Sex differences in response to maximal eccentric exercise. Med. Sci. Sports Exerc. 2008, 40, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Gao, S.; Puppa, M.J.; Kostek, M.C.; Wilson, L.B.; Carson, J.A. High-Frequency Stimulation on Skeletal Muscle Maintenance in Female Cachectic Mice. Med. Sci. Sports Exerc. 2019, 51, 1828–1837. [Google Scholar] [CrossRef]
- Takigawa, K.; Matsuda, R.; Uchitomi, R.; Onishi, T.; Hatazawa, Y.; Kamei, Y. Effects of long-term physical exercise on skeletal muscles in senescence-accelerated mice (SAMP8). Biosci. Biotechnol. Biochem. 2019, 83, 518–524. [Google Scholar] [CrossRef]
- Dreyer, H.C.; Fujita, S.; Cadenas, J.G.; Chinkes, D.L.; Volpi, E.; Rasmussen, B.B. Resistance exercise increases AMPK activity and reduces 4E-BP1 phosphorylation and protein synthesis in human skeletal muscle. J. Physiol. 2006, 576, 613–624. [Google Scholar] [CrossRef]
- Ferrando, A.A.; Tipton, K.D.; Bamman, M.M.; Wolfe, R.R. Resistance exercise maintains skeletal muscle protein synthesis during bed rest. J. Appl. Physiol. 1997, 82, 807–810. [Google Scholar] [CrossRef] [Green Version]
- Borges, I.B.P.; de Oliveira, D.S.; Marie, S.K.N.; Lenario, A.M.; Oba-Shinjo, S.M.; Shinjo, S.K. Exercise Training Attenuates Ubiquitin-Proteasome Pathway and Increases the Genes Related to Autophagy on the Skeletal Muscle of Patients With Inflammatory Myopathies. J. Clin. Rheumatol. 2021, 27, S224–S231. [Google Scholar] [CrossRef]
- Cunha, T.F.; Bacurau, A.V.; Moreira, J.B.; Paixao, N.A.; Campos, J.C.; Ferreira, J.C.; Leal, M.L.; Negrao, C.E.; Moriscot, A.S.; Wisloff, U.; et al. Exercise training prevents oxidative stress and ubiquitin-proteasome system overactivity and reverse skeletal muscle atrophy in heart failure. PLoS ONE 2012, 7, e41701. [Google Scholar] [CrossRef]
- Hardee, J.P.; Counts, B.R.; Gao, S.; VanderVeen, B.N.; Fix, D.K.; Koh, H.J.; Carson, J.A. Inflammatory signalling regulates eccentric contraction-induced protein synthesis in cachectic skeletal muscle. J. Cachexia Sarcopenia Muscle 2018, 9, 369–383. [Google Scholar] [CrossRef]
- Khamoui, A.V.; Park, B.S.; Kim, D.H.; Yeh, M.C.; Oh, S.L.; Elam, M.L.; Jo, E.; Arjmandi, B.H.; Salazar, G.; Grant, S.C.; et al. Aerobic and resistance training dependent skeletal muscle plasticity in the colon-26 murine model of cancer cachexia. Metabolism 2016, 65, 685–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, M.S. mTOR as a Key Regulator in Maintaining Skeletal Muscle Mass. Front. Physiol. 2017, 8, 788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puppa, M.J.; Murphy, E.A.; Fayad, R.; Hand, G.A.; Carson, J.A. Cachectic skeletal muscle response to a novel bout of low-frequency stimulation. J. Appl. Physiol. 2014, 116, 1078–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, J.P.; Puppa, M.J.; Gao, S.; Sato, S.; Welle, S.L.; Carson, J.A. Muscle mTORC1 suppression by IL-6 during cancer cachexia: A role for AMPK. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E1042–E1052. [Google Scholar] [CrossRef] [PubMed]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of metabolism and mitochondrial homeostasis. Nat. Rev. Mol. Cell Biol. 2018, 19, 121–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballaro, R.; Penna, F.; Pin, F.; Gomez-Cabrera, M.C.; Vina, J.; Costelli, P. Moderate Exercise Improves Experimental Cancer Cachexia by Modulating the Redox Homeostasis. Cancers 2019, 11, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves de Lima, E., Jr.; Teixeira, A.A.S.; Biondo, L.A.; Diniz, T.A.; Silveira, L.S.; Coletti, D.; Busquets Rius, S.; Rosa Neto, J.C. Exercise Reduces the Resumption of Tumor Growth and Proteolytic Pathways in the Skeletal Muscle of Mice Following Chemotherapy. Cancers 2020, 12, 3466. [Google Scholar] [CrossRef]
- Moller, A.B.; Lonbro, S.; Farup, J.; Voss, T.S.; Rittig, N.; Wang, J.; Hojris, I.; Mikkelsen, U.R.; Jessen, N. Molecular and cellular adaptations to exercise training in skeletal muscle from cancer patients treated with chemotherapy. J. Cancer Res. Clin. Oncol. 2019, 145, 1449–1460. [Google Scholar] [CrossRef]
- Reuben, D.B.; Judd-Hamilton, L.; Harris, T.B.; Seeman, T.E.; MacArthur Studies of Successful Aging. The associations between physical activity and inflammatory markers in high-functioning older persons: MacArthur Studies of Successful Aging. J. Am. Geriatr. Soc. 2003, 51, 1125–1130. [Google Scholar] [CrossRef]
- Abramson, J.L.; Vaccarino, V. Relationship between physical activity and inflammation among apparently healthy middle-aged and older US adults. Arch. Intern. Med. 2002, 162, 1286–1292. [Google Scholar] [CrossRef] [Green Version]
- Petersen, A.M.; Pedersen, B.K. The anti-inflammatory effect of exercise. J. Appl. Physiol. 2005, 98, 1154–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman-Goetz, L.; Spagnuolo, P.A.; Guan, J. Repeated exercise in mice alters expression of IL-10 and TNF-alpha in intestinal lymphocytes. Brain Behav. Immun. 2008, 22, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Hoffman-Goetz, L.; Pervaiz, N.; Guan, J. Voluntary exercise training in mice increases the expression of antioxidant enzymes and decreases the expression of TNF-alpha in intestinal lymphocytes. Brain Behav. Immun. 2009, 23, 498–506. [Google Scholar] [CrossRef]
- Tanaka, Y.; Kawanishi, N.; Shiva, D.; Tsutsumi, N.; Uchida, M.; Kitamura, H.; Kato, Y.; Yano, H. Exhaustive exercise reduces tumor necrosis factor-alpha production in response to lipopolysaccharide in mice. Neuroimmunomodulation 2010, 17, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Cerqueira, E.; Marinho, D.A.; Neiva, H.P.; Lourenco, O. Inflammatory Effects of High and Moderate Intensity Exercise-A Systematic Review. Front. Physiol. 2019, 10, 1550. [Google Scholar] [CrossRef] [PubMed]
- Steensberg, A.; Fischer, C.P.; Keller, C.; Moller, K.; Pedersen, B.K. IL-6 enhances plasma IL-1ra, IL-10, and cortisol in humans. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E433–E437. [Google Scholar] [CrossRef] [PubMed]
- Starkie, R.; Ostrowski, S.R.; Jauffred, S.; Febbraio, M.; Pedersen, B.K. Exercise and IL-6 infusion inhibit endotoxin-induced TNF-alpha production in humans. FASEB J. 2003, 17, 884–886. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Steensberg, A.; Fischer, C.; Keller, C.; Keller, P.; Plomgaard, P.; Febbraio, M.; Saltin, B. Searching for the exercise factor: Is IL-6 a candidate? J. Muscle Res. Cell Motil. 2003, 24, 113–119. [Google Scholar] [CrossRef]
- Munoz-Canoves, P.; Scheele, C.; Pedersen, B.K.; Serrano, A.L. Interleukin-6 myokine signaling in skeletal muscle: A double-edged sword? FEBS J. 2013, 280, 4131–4148. [Google Scholar] [CrossRef]
- FitzGerald, L.; Macey, P.M.; Brecht, M.L. Pathways to interleukin-6 in healthy males and serious leisure male athletes: Physical activity, body composition and age. PLoS ONE 2012, 7, e40513. [Google Scholar] [CrossRef] [Green Version]
- Lavin, K.M.; Perkins, R.K.; Jemiolo, B.; Raue, U.; Trappe, S.W.; Trappe, T.A. Effects of aging and lifelong aerobic exercise on basal and exercise-induced inflammation. J. Appl. Physiol. 2020, 128, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Aoi, W.; Naito, Y.; Takagi, T.; Kokura, S.; Mizushima, K.; Takanami, Y.; Kawai, Y.; Tanimura, Y.; Hung, L.P.; Koyama, R.; et al. Regular exercise reduces colon tumorigenesis associated with suppression of iNOS. Biochem. Biophys. Res. Commun. 2010, 399, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Shalamzari, S.A.; Agha-Alinejad, H.; Alizadeh, S.; Shahbazi, S.; Khatib, Z.K.; Kazemi, A.; Saei, M.A.; Minayi, N. The effect of exercise training on the level of tissue IL-6 and vascular endothelial growth factor in breast cancer bearing mice. Iran. J. Basic Med. Sci. 2014, 17, 231–258. [Google Scholar] [PubMed]
- Kleckner, I.R.; Kamen, C.; Cole, C.; Fung, C.; Heckler, C.E.; Guido, J.J.; Culakova, E.; Onitilo, A.A.; Conlin, A.; Kuebler, J.P.; et al. Effects of exercise on inflammation in patients receiving chemotherapy: A nationwide NCORP randomized clinical trial. Support. Care Cancer 2019, 27, 4615–4625. [Google Scholar] [CrossRef] [PubMed]
- Daou, H.N. Exercise as an anti-inflammatory therapy for cancer cachexia: A focus on interleukin-6 regulation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2020, 318, R296–R310. [Google Scholar] [CrossRef]
- Vanderveen, B.N.; Fix, D.K.; Counts, B.R.; Carson, J.A. The Effect of Wheel Exercise on Functional Indices of Cachexia in Tumor-bearing Mice. Med. Sci. Sports Exerc. 2020, 52, 2320–2330. [Google Scholar] [CrossRef]
- van Vulpen, J.K.; Schmidt, M.E.; Velthuis, M.J.; Wiskemann, J.; Schneeweiss, A.; Vermeulen, R.C.H.; Habermann, N.; Ulrich, C.M.; Peeters, P.H.M.; van der Wall, E.; et al. Effects of physical exercise on markers of inflammation in breast cancer patients during adjuvant chemotherapy. Breast Cancer Res. Treat. 2018, 168, 421–431. [Google Scholar] [CrossRef]
- Schauer, T.; Mazzoni, A.S.; Henriksson, A.; Demmelmaier, I.; Berntsen, S.; Raastad, T.; Nordin, K.; Pedersen, B.K.; Christensen, J.F. Exercise intensity and markers of inflammation during and after (neo-) adjuvant cancer treatment. Endocr. Relat. Cancer 2021, 28, 191–201. [Google Scholar] [CrossRef]
- Schmidt, M.E.; Meynkohn, A.; Habermann, N.; Wiskemann, J.; Oelmann, J.; Hof, H.; Wessels, S.; Klassen, O.; Debus, J.; Potthoff, K.; et al. Resistance Exercise and Inflammation in Breast Cancer Patients Undergoing Adjuvant Radiation Therapy: Mediation Analysis From a Randomized, Controlled Intervention Trial. Int. J. Radiat. Oncol. Biol. Phys. 2016, 94, 329–337. [Google Scholar] [CrossRef]
- Hiensch, A.E.; Mijwel, S.; Bargiela, D.; Wengstrom, Y.; May, A.M.; Rundqvist, H. Inflammation Mediates Exercise Effects on Fatigue in Patients with Breast Cancer. Med. Sci. Sports Exerc. 2021, 53, 496–504. [Google Scholar] [CrossRef]
- White, J.P.; Baltgalvis, K.A.; Puppa, M.J.; Sato, S.; Baynes, J.W.; Carson, J.A. Muscle oxidative capacity during IL-6-dependent cancer cachexia. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R201–R211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, J.P.; Puppa, M.J.; Sato, S.; Gao, S.; Price, R.L.; Baynes, J.W.; Kostek, M.C.; Matesic, L.E.; Carson, J.A. IL-6 regulation on skeletal muscle mitochondrial remodeling during cancer cachexia in the ApcMin/+ mouse. Skelet. Muscle 2012, 2, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukawa, T.; Yan-Jiang, B.C.; Min-Wen, J.C.; Jun-Hao, E.T.; Huang, D.; Qian, C.N.; Ong, P.; Li, Z.; Chen, S.; Mak, S.Y.; et al. Excessive fatty acid oxidation induces muscle atrophy in cancer cachexia. Nat. Med. 2016, 22, 666–671. [Google Scholar] [CrossRef] [PubMed]
- McLean, J.B.; Moylan, J.S.; Andrade, F.H. Mitochondria dysfunction in lung cancer-induced muscle wasting in C2C12 myotubes. Front. Physiol. 2014, 5, 503. [Google Scholar] [CrossRef] [Green Version]
- Halle, J.L.; Pena, G.S.; Paez, H.G.; Castro, A.J.; Rossiter, H.B.; Visavadiya, N.P.; Whitehurst, M.A.; Khamoui, A.V. Tissue-specific dysregulation of mitochondrial respiratory capacity and coupling control in colon-26 tumor-induced cachexia. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2019, 317, R68–R82. [Google Scholar] [CrossRef]
- Memme, J.M.; Erlich, A.T.; Phukan, G.; Hood, D.A. Exercise and mitochondrial health. J. Physiol. 2021, 599, 803–817. [Google Scholar] [CrossRef]
- Jacobs, R.A.; Lundby, C. Mitochondria express enhanced quality as well as quantity in association with aerobic fitness across recreationally active individuals up to elite athletes. J. Appl. Physiol. 2013, 114, 344–350. [Google Scholar] [CrossRef] [Green Version]
- Porter, C.; Reidy, P.T.; Bhattarai, N.; Sidossis, L.S.; Rasmussen, B.B. Resistance Exercise Training Alters Mitochondrial Function in Human Skeletal Muscle. Med. Sci. Sports Exerc. 2015, 47, 1922–1931. [Google Scholar] [CrossRef] [Green Version]
- Kitaoka, Y.; Miyazaki, M.; Kikuchi, S. Voluntary exercise prevents abnormal muscle mitochondrial morphology in cancer cachexia mice. Physiol. Rep. 2021, 9, e15016. [Google Scholar] [CrossRef]
- Montero-Bullon, J.F.; Melo, T.; Ferreira, R.; Padrao, A.I.; Oliveira, P.A.; Domingues, M.R.M.; Domingues, P. Exercise training counteracts urothelial carcinoma-induced alterations in skeletal muscle mitochondria phospholipidome in an animal model. Sci. Rep. 2019, 9, 13423. [Google Scholar] [CrossRef] [Green Version]
- Halling, J.F.; Pilegaard, H. PGC-1alpha-mediated regulation of mitochondrial function and physiological implications. Appl. Physiol. Nutr. Metab. 2020, 45, 927–936. [Google Scholar] [CrossRef] [PubMed]
- Knight, K.; Wade, S.; Balducci, L. Prevalence and outcomes of anemia in cancer: A systematic review of the literature. Am. J. Med. 2004, 116 (Suppl. 7A), 11S–26S. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Rivera, S.; Shprung, D.; Limbrick, D.; Gabayan, V.; Nemeth, E.; Ganz, T. Mouse models of anemia of cancer. PLoS ONE 2014, 9, e93283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grune, T.; Sommerburg, O.; Siems, W.G. Oxidative stress in anemia. Clin. Nephrol. 2000, 53, S18–S22. [Google Scholar]
- Inaba, S.; Hinohara, A.; Tachibana, M.; Tsujikawa, K.; Fukada, S.I. Muscle regeneration is disrupted by cancer cachexia without loss of muscle stem cell potential. PLoS ONE 2018, 13, e0205467. [Google Scholar] [CrossRef] [Green Version]
- Murach, K.A.; Fry, C.S.; Dupont-Versteegden, E.E.; McCarthy, J.J.; Peterson, C.A. Fusion and beyond: Satellite cell contributions to loading-induced skeletal muscle adaptation. FASEB J. 2021, 35, e21893. [Google Scholar] [CrossRef]
- Murach, K.A.; Peck, B.D.; Policastro, R.A.; Vechetti, I.J.; Van Pelt, D.W.; Dungan, C.M.; Denes, L.T.; Fu, X.; Brightwell, C.R.; Zentner, G.E.; et al. Early satellite cell communication creates a permissive environment for long-term muscle growth. iScience 2021, 24, 102372. [Google Scholar] [CrossRef]
- Murach, K.A.; Vechetti, I.J., Jr.; Van Pelt, D.W.; Crow, S.E.; Dungan, C.M.; Figueiredo, V.C.; Kosmac, K.; Fu, X.; Richards, C.I.; Fry, C.S.; et al. Fusion-Independent Satellite Cell Communication to Muscle Fibers During Load-Induced Hypertrophy. Function 2020, 1, zqaa009. [Google Scholar] [CrossRef]
- Fry, C.S.; Kirby, T.J.; Kosmac, K.; McCarthy, J.J.; Peterson, C.A. Myogenic Progenitor Cells Control Extracellular Matrix Production by Fibroblasts during Skeletal Muscle Hypertrophy. Cell Stem Cell 2017, 20, 56–69. [Google Scholar] [CrossRef] [Green Version]
- D’Lugos, A.C.; Fry, C.S.; Ormsby, J.C.; Sweeney, K.R.; Brightwell, C.R.; Hale, T.M.; Gonzales, R.J.; Angadi, S.S.; Carroll, C.C.; Dickinson, J.M. Chronic doxorubicin administration impacts satellite cell and capillary abundance in a muscle-specific manner. Physiol. Rep. 2019, 7, e14052. [Google Scholar] [CrossRef] [Green Version]
- Mallard, J.; Hucteau, E.; Hureau, T.J.; Pagano, A.F. Skeletal Muscle Deconditioning in Breast Cancer Patients Undergoing Chemotherapy: Current Knowledge and Insights From Other Cancers. Front. Cell Dev. Biol. 2021, 9, 719643. [Google Scholar] [CrossRef] [PubMed]
- Solsona, R.; Pavlin, L.; Bernardi, H.; Sanchez, A.M. Molecular Regulation of Skeletal Muscle Growth and Organelle Biosynthesis: Practical Recommendations for Exercise Training. Int. J. Mol. Sci. 2021, 22, 2741. [Google Scholar] [CrossRef] [PubMed]
- Kadi, F.; Eriksson, A.; Holmner, S.; Butler-Browne, G.S.; Thornell, L.E. Cellular adaptation of the trapezius muscle in strength-trained athletes. Histochem. Cell Biol. 1999, 111, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Verdijk, L.B.; Gleeson, B.G.; Jonkers, R.A.; Meijer, K.; Savelberg, H.H.; Dendale, P.; van Loon, L.J. Skeletal muscle hypertrophy following resistance training is accompanied by a fiber type-specific increase in satellite cell content in elderly men. J. Gerontol. A Biol. Sci. Med. Sci. 2009, 64, 332–339. [Google Scholar] [CrossRef]
- Kadi, F.; Thornell, L.E. Concomitant increases in myonuclear and satellite cell content in female trapezius muscle following strength training. Histochem. Cell Biol. 2000, 113, 99–103. [Google Scholar] [CrossRef]
- Coletti, D.; Aulino, P.; Pigna, E.; Barteri, F.; Moresi, V.; Annibali, D.; Adamo, S.; Berardi, E. Spontaneous Physical Activity Downregulates Pax7 in Cancer Cachexia. Stem Cells Int. 2016, 2016, 6729268. [Google Scholar] [CrossRef] [Green Version]
- Nilsen, T.S.; Thorsen, L.; Fossa, S.D.; Wiig, M.; Kirkegaard, C.; Skovlund, E.; Benestad, H.B.; Raastad, T. Effects of strength training on muscle cellular outcomes in prostate cancer patients on androgen deprivation therapy. Scand. J. Med. Sci. Sports 2016, 26, 1026–1035. [Google Scholar] [CrossRef]
- Christensen, J.F.; Schjerling, P.; Andersen, J.L.; Daugaard, G.; Rorth, M.; Mackey, A.L. Muscle satellite cell content and mRNA signaling in germ cell cancer patients—Effects of chemotherapy and resistance training. Acta Oncol. 2016, 55, 1246–1250. [Google Scholar] [CrossRef] [Green Version]
- van Velthoven, C.T.J.; Rando, T.A. Stem Cell Quiescence: Dynamism, Restraint, and Cellular Idling. Cell Stem Cell 2019, 24, 213–225. [Google Scholar] [CrossRef] [Green Version]
- Judge, S.M.; Nosacka, R.L.; Delitto, D.; Gerber, M.H.; Cameron, M.E.; Trevino, J.G.; Judge, A.R. Skeletal Muscle Fibrosis in Pancreatic Cancer Patients with Respect to Survival. JNCI Cancer Spectr. 2018, 2, pky043. [Google Scholar] [CrossRef]
- Ogle, K.S.; Swanson, G.M.; Woods, N.; Azzouz, F. Cancer and comorbidity: Redefining chronic diseases. Cancer 2000, 88, 653–663. [Google Scholar] [CrossRef]
- Fowler, H.; Belot, A.; Ellis, L.; Maringe, C.; Luque-Fernandez, M.A.; Njagi, E.N.; Navani, N.; Sarfati, D.; Rachet, B. Comorbidity prevalence among cancer patients: A population-based cohort study of four cancers. BMC Cancer 2020, 20, 2. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.W.; Reeve, B.B.; Bellizzi, K.M.; Harlan, L.C.; Klabunde, C.N.; Amsellem, M.; Bierman, A.S.; Hays, R.D. Cancer, comorbidities, and health-related quality of life of older adults. Health Care Financ. Rev. 2008, 29, 41–56. [Google Scholar] [PubMed]
- Natalucci, V.; Virgili, E.; Calcagnoli, F.; Valli, G.; Agostini, D.; Zeppa, S.D.; Barbieri, E.; Emili, R. Cancer Related Anemia: An Integrated Multitarget Approach and Lifestyle Interventions. Nutrients 2021, 13, 482. [Google Scholar] [CrossRef] [PubMed]
- Rajarajeswaran, P.; Vishnupriya, R. Exercise in cancer. Indian J. Med. Paediatr. Oncol. 2009, 30, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.W.; Zhang, Q.; Song, M.M.; Zhang, K.P.; Zhang, X.; Ruan, G.T.; Yang, M.; Ge, Y.Z.; Tang, M.; Li, X.R.; et al. The prognostic effect of hemoglobin on patients with cancer cachexia: A multicenter retrospective cohort study. Support. Care Cancer 2022, 30, 875–885. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, G.M., 3rd; Becker, P.S.; Blinder, M.; Cella, D.; Chanan-Khan, A.; Cleeland, C.; Coccia, P.F.; Djulbegovic, B.; Gilreath, J.A.; Kraut, E.H.; et al. Cancer- and chemotherapy-induced anemia. J. Natl. Compr. Canc. Netw. 2012, 10, 628–653. [Google Scholar] [CrossRef] [Green Version]
- Ferrario, E.; Ferrari, L.; Bidoli, P.; De Candis, D.; Del Vecchio, M.; De Dosso, S.; Buzzoni, R.; Bajetta, E. Treatment of cancer-related anemia with epoetin alfa: A review. Cancer Treat. Rev. 2004, 30, 563–575. [Google Scholar] [CrossRef]
- Littlewood, T.J.; Bajetta, E.; Nortier, J.W.; Vercammen, E.; Rapoport, B.; Epoetin Alfa Study Group. Effects of epoetin alfa on hematologic parameters and quality of life in cancer patients receiving nonplatinum chemotherapy: Results of a randomized, double-blind, placebo-controlled trial. J. Clin. Oncol. 2001, 19, 2865–2874. [Google Scholar] [CrossRef]
- Wright, J.R.; Ung, Y.C.; Julian, J.A.; Pritchard, K.I.; Whelan, T.J.; Smith, C.; Szechtman, B.; Roa, W.; Mulroy, L.; Rudinskas, L.; et al. Randomized, double-blind, placebo-controlled trial of erythropoietin in non-small-cell lung cancer with disease-related anemia. J. Clin. Oncol. 2007, 25, 1027–1032. [Google Scholar] [CrossRef]
- Tankiewicz-Kwedlo, A.; Hermanowicz, J.; Surazynski, A.; Rozkiewicz, D.; Pryczynicz, A.; Domaniewski, T.; Pawlak, K.; Kemona, A.; Pawlak, D. Erythropoietin accelerates tumor growth through increase of erythropoietin receptor (EpoR) as well as by the stimulation of angiogenesis in DLD-1 and Ht-29 xenografts. Mol. Cell. Biochem. 2016, 421, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, C.L.; Silver, S.M.; Djulbegovic, B.; Samaras, A.T.; Blau, C.A.; Gleason, K.J.; Barnato, S.E.; Elverman, K.M.; Courtney, D.M.; McKoy, J.M.; et al. Venous thromboembolism and mortality associated with recombinant erythropoietin and darbepoetin administration for the treatment of cancer-associated anemia. JAMA 2008, 299, 914–924. [Google Scholar] [CrossRef] [PubMed]
- Escobar Alvarez, Y.; de Las Penas Bataller, R.; Perez Altozano, J.; Ros Martinez, S.; Sabino Alvarez, A.; Blasco Cordellat, A.; Brozos Vazquez, E.; Corral Jaime, J.; Garcia Escobar, I.; Beato Zambrano, C. SEOM clinical guidelines for anaemia treatment in cancer patients (2020). Clin. Transl. Oncol. 2021, 23, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Razeq, H.; Saadeh, S.S.; Malhis, R.; Yasser, S.; Abdulelah, H.; Eljaber, R.; Kleib, A.; Ismael, R. Treatment of anemia in cancer patients undergoing chemotherapy with intravenous ferric carboxymaltose without erythropoiesis-stimulating agents. Ther. Adv. Med. Oncol. 2020, 12, 1758835920953292. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, H.; Evstatiev, R.; Kornek, G.; Aapro, M.; Bauernhofer, T.; Buxhofer-Ausch, V.; Fridrik, M.; Geissler, D.; Geissler, K.; Gisslinger, H.; et al. Iron metabolism and iron supplementation in cancer patients. Wien. Klin. Wochenschr. 2015, 127, 907–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, M.; Nishijima, Y.; Asp, M.L.; Stout, M.B.; Reiser, P.J.; Belury, M.A. Cardiac alterations in cancer-induced cachexia in mice. Int. J. Oncol. 2010, 37, 347–353. [Google Scholar] [CrossRef] [Green Version]
- Cosper, P.F.; Leinwand, L.A. Cancer causes cardiac atrophy and autophagy in a sexually dimorphic manner. Cancer Res. 2011, 71, 1710–1720. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Crawford, D.; Hutchinson, K.R.; Youtz, D.J.; Lucchesi, P.A.; Velten, M.; McCarthy, D.O.; Wold, L.E. Myocardial dysfunction in an animal model of cancer cachexia. Life Sci. 2011, 88, 406–410. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.E.; Brown, J.L.; Rosa-Caldwell, M.E.; Perry, R.A.; Brown, L.A.; Haynie, W.S.; Washington, T.A.; Wiggs, M.P.; Rajaram, N.; Greene, N.P. Cancer-induced Cardiac Atrophy Adversely Affects Myocardial Redox State and Mitochondrial Oxidative Characteristics. JCSM Rapid. Commun. 2021, 4, 3–15. [Google Scholar] [CrossRef]
- Mishra, S.; Tamta, A.K.; Sarikhani, M.; Desingu, P.A.; Kizkekra, S.M.; Pandit, A.S.; Kumar, S.; Khan, D.; Raghavan, S.C.; Sundaresan, N.R. Subcutaneous Ehrlich Ascites Carcinoma mice model for studying cancer-induced cardiomyopathy. Sci. Rep. 2018, 8, 5599. [Google Scholar] [CrossRef]
- Springer, J.; Tschirner, A.; Haghikia, A.; von Haehling, S.; Lal, H.; Grzesiak, A.; Kaschina, E.; Palus, S.; Potsch, M.; von Websky, K.; et al. Prevention of liver cancer cachexia-induced cardiac wasting and heart failure. Eur. Heart J. 2014, 35, 932–941. [Google Scholar] [CrossRef] [PubMed]
- Bertero, E.; Canepa, M.; Maack, C.; Ameri, P. Linking Heart Failure to Cancer: Background Evidence and Research Perspectives. Circulation 2018, 138, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Ameri, P.; Canepa, M.; Anker, M.S.; Belenkov, Y.; Bergler-Klein, J.; Cohen-Solal, A.; Farmakis, D.; Lopez-Fernandez, T.; Lainscak, M.; Pudil, R.; et al. Cancer diagnosis in patients with heart failure: Epidemiology, clinical implications and gaps in knowledge. Eur. J. Heart Fail. 2018, 20, 879–887. [Google Scholar] [CrossRef]
- Lyon, A.R. Heart failure resulting from cancer treatment: Still serious but an opportunity for prevention. Heart 2019, 105, 6–8. [Google Scholar] [CrossRef] [PubMed]
- Hamo, C.E.; Bloom, M.W. Cancer and Heart Failure: Understanding the Intersection. Card. Fail. Rev. 2017, 3, 66–70. [Google Scholar] [CrossRef] [Green Version]
- Sturgeon, K.M.; Deng, L.; Bluethmann, S.M.; Zhou, S.; Trifiletti, D.M.; Jiang, C.; Kelly, S.P.; Zaorsky, N.G. A population-based study of cardiovascular disease mortality risk in US cancer patients. Eur. Heart J. 2019, 40, 3889–3897. [Google Scholar] [CrossRef] [Green Version]
- Stoltzfus, K.C.; Zhang, Y.; Sturgeon, K.; Sinoway, L.I.; Trifiletti, D.M.; Chinchilli, V.M.; Zaorsky, N.G. Fatal heart disease among cancer patients. Nat. Commun. 2020, 11, 2011. [Google Scholar] [CrossRef]
- Pina, I.L.; Apstein, C.S.; Balady, G.J.; Belardinelli, R.; Chaitman, B.R.; Duscha, B.D.; Fletcher, B.J.; Fleg, J.L.; Myers, J.N.; Sullivan, M.J.; et al. Exercise and heart failure: A statement from the American Heart Association Committee on exercise, rehabilitation, and prevention. Circulation 2003, 107, 1210–1225. [Google Scholar] [CrossRef]
- Allan, J.; Buss, L.A.; Draper, N.; Currie, M.J. Exercise in People With Cancer: A Spotlight on Energy Regulation and Cachexia. Front. Physiol. 2022, 13, 836804. [Google Scholar] [CrossRef]
- Kampshoff, C.S.; Chinapaw, M.J.; Brug, J.; Twisk, J.W.; Schep, G.; Nijziel, M.R.; van Mechelen, W.; Buffart, L.M. Randomized controlled trial of the effects of high intensity and low-to-moderate intensity exercise on physical fitness and fatigue in cancer survivors: Results of the Resistance and Endurance exercise After ChemoTherapy (REACT) study. BMC Med. 2015, 13, 275. [Google Scholar] [CrossRef] [Green Version]
- Henriksson, A.; Johansson, B.; Radu, C.; Berntsen, S.; Igelstrom, H.; Nordin, K. Is it safe to exercise during oncological treatment? A study of adverse events during endurance and resistance training—Data from the Phys-Can study. Acta Oncol. 2021, 60, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Delphan, M.; Delfan, N.; West, D.; Delfan, M. Exercise protocols: The gap between preclinical and clinical exercise oncology studies. Metabol. Open 2022, 13, 100165. [Google Scholar] [CrossRef] [PubMed]
- Denis, C.; Chatard, J.C.; Dormois, D.; Linossier, M.T.; Geyssant, A.; Lacour, J.R. Effects of endurance training on capillary supply of human skeletal muscle on two age groups (20 and 60 years). J. Physiol. 1986, 81, 379–383. [Google Scholar]
- Ingjer, F. Effects of endurance training on muscle fibre ATP-ase activity, capillary supply and mitochondrial content in man. J. Physiol. 1979, 294, 419–432. [Google Scholar] [CrossRef] [PubMed]
- Turner, D.L.; Hoppeler, H.; Claassen, H.; Vock, P.; Kayser, B.; Schena, F.; Ferretti, G. Effects of endurance training on oxidative capacity and structural composition of human arm and leg muscles. Acta Physiol. Scand. 1997, 161, 459–464. [Google Scholar] [CrossRef]
- Hood, M.S.; Little, J.P.; Tarnopolsky, M.A.; Myslik, F.; Gibala, M.J. Low-volume interval training improves muscle oxidative capacity in sedentary adults. Med. Sci. Sports Exerc. 2011, 43, 1849–1856. [Google Scholar] [CrossRef]
- Konopka, A.R.; Harber, M.P. Skeletal muscle hypertrophy after aerobic exercise training. Exerc. Sport Sci. Rev. 2014, 42, 53–61. [Google Scholar] [CrossRef]
- Wilson, J.M.; Marin, P.J.; Rhea, M.R.; Wilson, S.M.; Loenneke, J.P.; Anderson, J.C. Concurrent training: A meta-analysis examining interference of aerobic and resistance exercises. J. Strength Cond. Res. 2012, 26, 2293–2307. [Google Scholar] [CrossRef]
- Murach, K.A.; Bagley, J.R. Skeletal Muscle Hypertrophy with Concurrent Exercise Training: Contrary Evidence for an Interference Effect. Sports Med. 2016, 46, 1029–1039. [Google Scholar] [CrossRef]
- Gerritsen, J.K.; Vincent, A.J. Exercise improves quality of life in patients with cancer: A systematic review and meta-analysis of randomised controlled trials. Br. J. Sports Med. 2016, 50, 796–803. [Google Scholar] [CrossRef]
- Knobf, M.T.; Musanti, R.; Dorward, J. Exercise and quality of life outcomes in patients with cancer. Semin. Oncol. Nurs. 2007, 23, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Strasser, B.; Steindorf, K.; Wiskemann, J.; Ulrich, C.M. Impact of resistance training in cancer survivors: A meta-analysis. Med. Sci. Sports Exerc. 2013, 45, 2080–2090. [Google Scholar] [CrossRef] [PubMed]
- Mavropalias, G.; Sim, M.; Taaffe, D.R.; Galvao, D.A.; Spry, N.; Kraemer, W.J.; Hakkinen, K.; Newton, R.U. Exercise medicine for cancer cachexia: Targeted exercise to counteract mechanisms and treatment side effects. J. Cancer Res. Clin. Oncol. 2022, 148, 1389–1406. [Google Scholar] [CrossRef] [PubMed]
- Aquila, G.; Re Cecconi, A.D.; Brault, J.J.; Corli, O.; Piccirillo, R. Nutraceuticals and Exercise against Muscle Wasting during Cancer Cachexia. Cells 2020, 9, 2536. [Google Scholar] [CrossRef] [PubMed]
- Knols, R.H. Endurance training for cancer patients during and shortly after completion of the anti-cancer treatment. Eur. Rev. Aging Phys. Act. 2013, 10, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Samuel, S.R.; Maiya, G.A.; Babu, A.S.; Vidyasagar, M.S. Effect of exercise training on functional capacity & quality of life in head & neck cancer patients receiving chemoradiotherapy. Indian J. Med. Res. 2013, 137, 515–520. [Google Scholar]
- Adamsen, L.; Quist, M.; Andersen, C.; Moller, T.; Herrstedt, J.; Kronborg, D.; Baadsgaard, M.T.; Vistisen, K.; Midtgaard, J.; Christiansen, B.; et al. Effect of a multimodal high intensity exercise intervention in cancer patients undergoing chemotherapy: Randomised controlled trial. BMJS 2009, 339, b3410. [Google Scholar] [CrossRef] [Green Version]
- Henke, C.C.; Cabri, J.; Fricke, L.; Pankow, W.; Kandilakis, G.; Feyer, P.C.; de Wit, M. Strength and endurance training in the treatment of lung cancer patients in stages IIIA/IIIB/IV. Support. Care Cancer 2014, 22, 95–101. [Google Scholar] [CrossRef]
- Coen, P.M.; Musci, R.V.; Hinkley, J.M.; Miller, B.F. Mitochondria as a Target for Mitigating Sarcopenia. Front. Physiol. 2018, 9, 1883. [Google Scholar] [CrossRef] [Green Version]
- Fyfe, J.J.; Bishop, D.J.; Stepto, N.K. Interference between concurrent resistance and endurance exercise: Molecular bases and the role of individual training variables. Sports Med. 2014, 44, 743–762. [Google Scholar] [CrossRef]
- Tsitkanou, S.; Spengos, K.; Stasinaki, A.N.; Zaras, N.; Bogdanis, G.; Papadimas, G.; Terzis, G. Effects of high-intensity interval cycling performed after resistance training on muscle strength and hypertrophy. Scand. J. Med. Sci. Sports 2017, 27, 1317–1327. [Google Scholar] [CrossRef] [PubMed]
- Murach, K.A.; McCarthy, J.J.; Peterson, C.A.; Dungan, C.M. Making Mice Mighty: Recent advances in translational models of load-induced muscle hypertrophy. J. Appl. Physiol. 2020, 129, 516–521. [Google Scholar] [CrossRef] [PubMed]
- Dungan, C.M.; Murach, K.A.; Frick, K.K.; Jones, S.R.; Crow, S.E.; Englund, D.A.; Vechetti, I.J., Jr.; Figueiredo, V.C.; Levitan, B.M.; Satin, J.; et al. Elevated myonuclear density during skeletal muscle hypertrophy in response to training is reversed during detraining. Am. J. Physiol. Cell Physiol. 2019, 316, C649–C654. [Google Scholar] [CrossRef] [PubMed]
- Murach, K.A.; Dungan, C.M.; Peterson, C.A.; McCarthy, J.J. Muscle Fiber Splitting Is a Physiological Response to Extreme Loading in Animals. Exerc. Sport Sci. Rev. 2019, 47, 108–115. [Google Scholar] [CrossRef]
- Cui, D.; Drake, J.C.; Wilson, R.J.; Shute, R.J.; Lewellen, B.; Zhang, M.; Zhao, H.; Sabik, O.L.; Onengut, S.; Berr, S.S.; et al. A novel voluntary weightlifting model in mice promotes muscle adaptation and insulin sensitivity with simultaneous enhancement of autophagy and mTOR pathway. FASEB J. 2020, 34, 7330–7344. [Google Scholar] [CrossRef] [Green Version]
- Rosa-Caldwell, M.E.; Greene, N.P. Muscle metabolism and atrophy: Let’s talk about sex. Biol. Sex Differ. 2019, 10, 43. [Google Scholar] [CrossRef]
- Hetzler, K.L.; Hardee, J.P.; Puppa, M.J.; Narsale, A.A.; Sato, S.; Davis, J.M.; Carson, J.A. Sex differences in the relationship of IL-6 signaling to cancer cachexia progression. Biochim. Biophys. Acta 2015, 1852, 816–825. [Google Scholar] [CrossRef] [Green Version]
- Neal, A.; Boldrin, L.; Morgan, J.E. The satellite cell in male and female, developing and adult mouse muscle: Distinct stem cells for growth and regeneration. PLoS ONE 2012, 7, e37950. [Google Scholar] [CrossRef] [Green Version]
- Manzano, R.; Toivonen, J.M.; Calvo, A.C.; Miana-Mena, F.J.; Zaragoza, P.; Munoz, M.J.; Montarras, D.; Osta, R. Sex, fiber-type, and age dependent in vitro proliferation of mouse muscle satellite cells. J. Cell. Biochem. 2011, 112, 2825–2836. [Google Scholar] [CrossRef]
- Scalzo, R.L.; Peltonen, G.L.; Binns, S.E.; Shankaran, M.; Giordano, G.R.; Hartley, D.A.; Klochak, A.L.; Lonac, M.C.; Paris, H.L.; Szallar, S.E.; et al. Greater muscle protein synthesis and mitochondrial biogenesis in males compared with females during sprint interval training. FASEB J. 2014, 28, 2705–2714. [Google Scholar] [CrossRef]
- Carter, S.L.; Rennie, C.D.; Hamilton, S.J.; Tarnopolsky, M.A. Changes in skeletal muscle in males and females following endurance training. Can. J. Physiol. Pharmacol. 2001, 79, 386–392. [Google Scholar] [CrossRef] [PubMed]
- West, D.W.; Burd, N.A.; Churchward-Venne, T.A.; Camera, D.M.; Mitchell, C.J.; Baker, S.K.; Hawley, J.A.; Coffey, V.G.; Phillips, S.M. Sex-based comparisons of myofibrillar protein synthesis after resistance exercise in the fed state. J. Appl. Physiol. 2012, 112, 1805–1813. [Google Scholar] [CrossRef] [PubMed]
- Hunter, S.K. Sex differences in fatigability of dynamic contractions. Exp. Physiol. 2016, 101, 250–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansdell, P.; Brownstein, C.G.; Skarabot, J.; Hicks, K.M.; Howatson, G.; Thomas, K.; Hunter, S.K.; Goodall, S. Sex differences in fatigability and recovery relative to the intensity-duration relationship. J. Physiol. 2019, 597, 5577–5595. [Google Scholar] [CrossRef] [PubMed]
- al-Majid, S.; McCarthy, D.O. Resistance exercise training attenuates wasting of the extensor digitorum longus muscle in mice bearing the colon-26 adenocarcinoma. Biol. Res. Nurs. 2001, 2, 155–166. [Google Scholar] [CrossRef]
- Montalvo, R.N.; Counts, B.R.; Carson, J.A. Understanding sex differences in the regulation of cancer-induced muscle wasting. Curr. Opin. Support. Palliat. Care 2018, 12, 394–403. [Google Scholar] [CrossRef]
- Adraskela, K.; Veisaki, E.; Koutsilieris, M.; Philippou, A. Physical Exercise Positively Influences Breast Cancer Evolution. Clin. Breast Cancer 2017, 17, 408–417. [Google Scholar] [CrossRef]
- Battaglini, C.; Bottaro, M.; Dennehy, C.; Rae, L.; Shields, E.; Kirk, D.; Hackney, A.C. The effects of an individualized exercise intervention on body composition in breast cancer patients undergoing treatment. Sao Paulo Med. J. 2007, 125, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Galvao, D.A.; Taaffe, D.R.; Spry, N.; Joseph, D.; Newton, R.U. Combined resistance and aerobic exercise program reverses muscle loss in men undergoing androgen suppression therapy for prostate cancer without bone metastases: A randomized controlled trial. J. Clin. Oncol. 2010, 28, 340–347. [Google Scholar] [CrossRef] [Green Version]
- Andersen, M.F.; Midtgaard, J.; Bjerre, E.D. Do Patients with Prostate Cancer Benefit from Exercise Interventions? A Systematic Review and Meta-Analysis. Int. J. Environ. Res. Public Health 2022, 19, 972. [Google Scholar] [CrossRef]
Figure 2.
Schematic summary of the review: effects of exercise training in the major mechanisms of cancer cachexia (protein turnover, inflammation, mitochondrial function and satellite cell regulation); and the influence of cancer comorbidities, exercise modality and biological sex in the effectiveness of exercise training in cancer cachexia.
Figure 2.
Schematic summary of the review: effects of exercise training in the major mechanisms of cancer cachexia (protein turnover, inflammation, mitochondrial function and satellite cell regulation); and the influence of cancer comorbidities, exercise modality and biological sex in the effectiveness of exercise training in cancer cachexia.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tsitkanou, S.; Murach, K.A.; Washington, T.A.; Greene, N.P. Exercise Counteracts the Deleterious Effects of Cancer Cachexia. Cancers 2022, 14, 2512. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14102512
AMA Style
Tsitkanou S, Murach KA, Washington TA, Greene NP. Exercise Counteracts the Deleterious Effects of Cancer Cachexia. Cancers. 2022; 14(10):2512. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14102512
Chicago/Turabian StyleTsitkanou, Stavroula, Kevin A. Murach, Tyrone A. Washington, and Nicholas P. Greene. 2022. "Exercise Counteracts the Deleterious Effects of Cancer Cachexia" Cancers 14, no. 10: 2512. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14102512
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.