
Implication of COPB2 Expression on Cutaneous Squamous Cell Carcinoma Pathogenesis

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
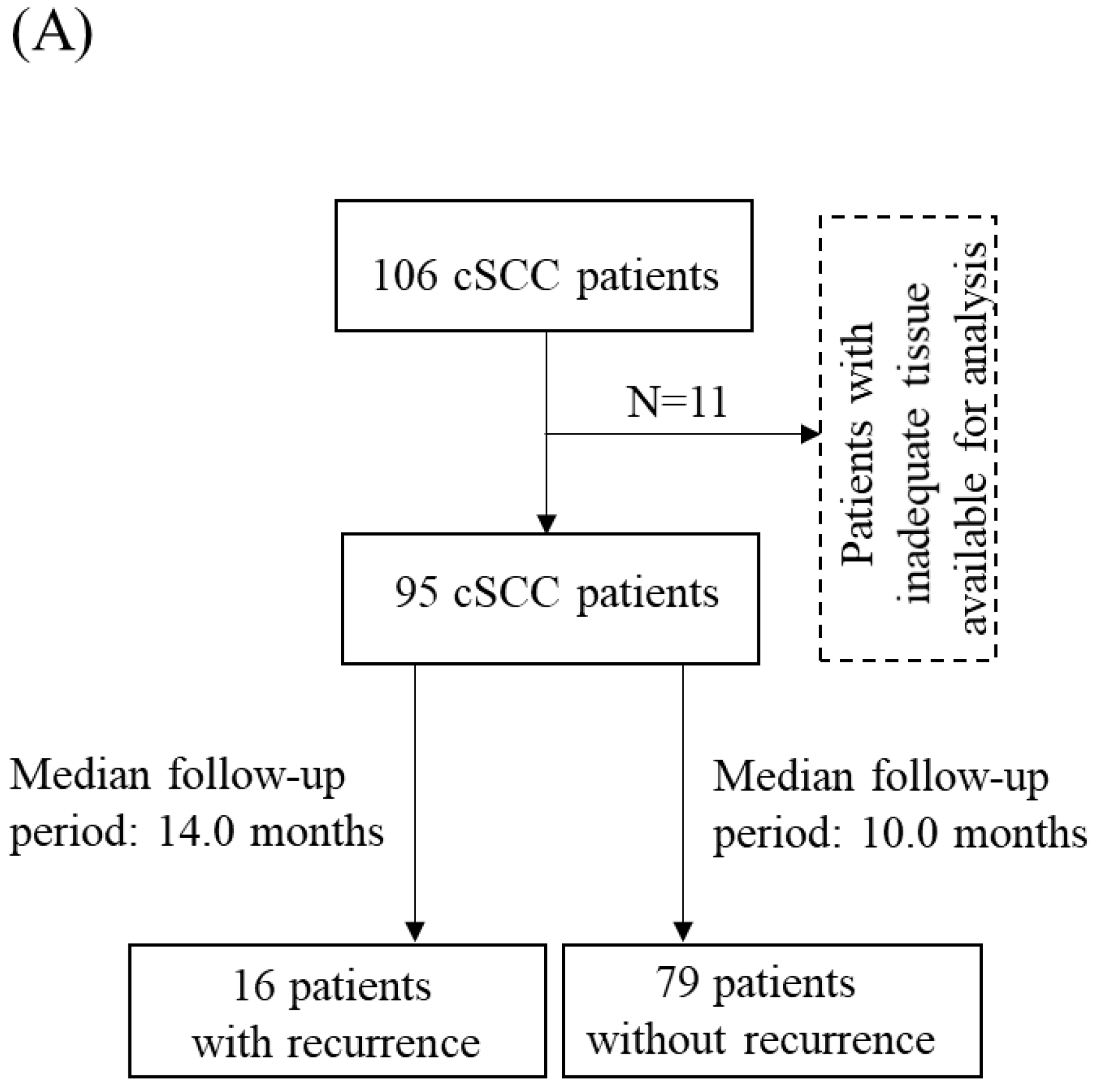
2.1. Clinical Samples
2.2. Establishment of COPB2 Knockdown cSCC Cells In Vitro
2.3. Trypan Blue Assay
2.4. Matrigel Invasion Assay
2.5. Cell Apoptosis Assay
2.6. In Vivo Xenograft Mouse Model
2.7. Immunochemical Staining
2.8. Statistical Analysis
3. Results
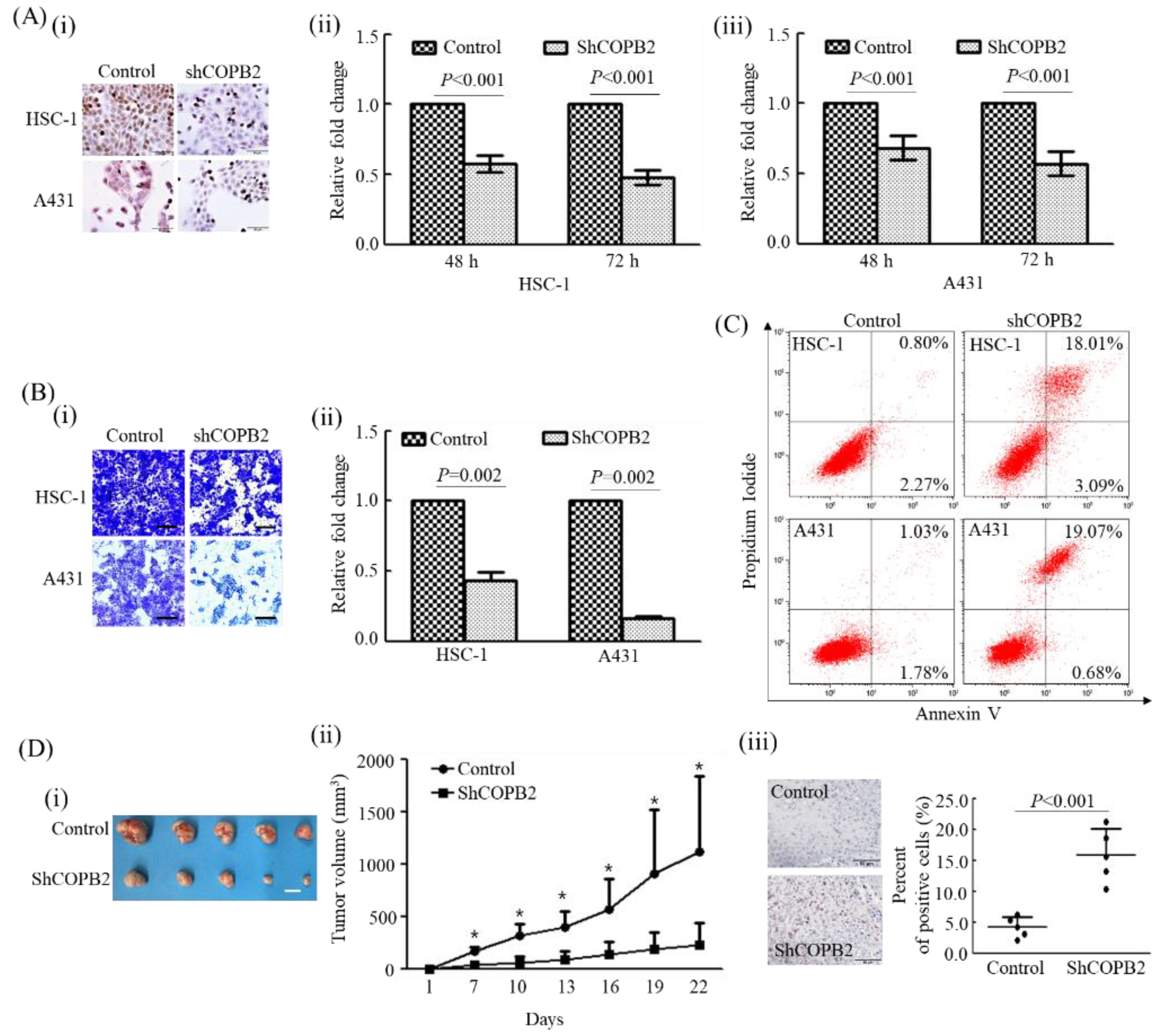
3.1. COPB2 Knockdown Largely Influenced the Biological Behavior of cSCC Cells In Vitro
3.2. COPB2 Knockdown Attenuated Tumorigenic Activity In Vivo
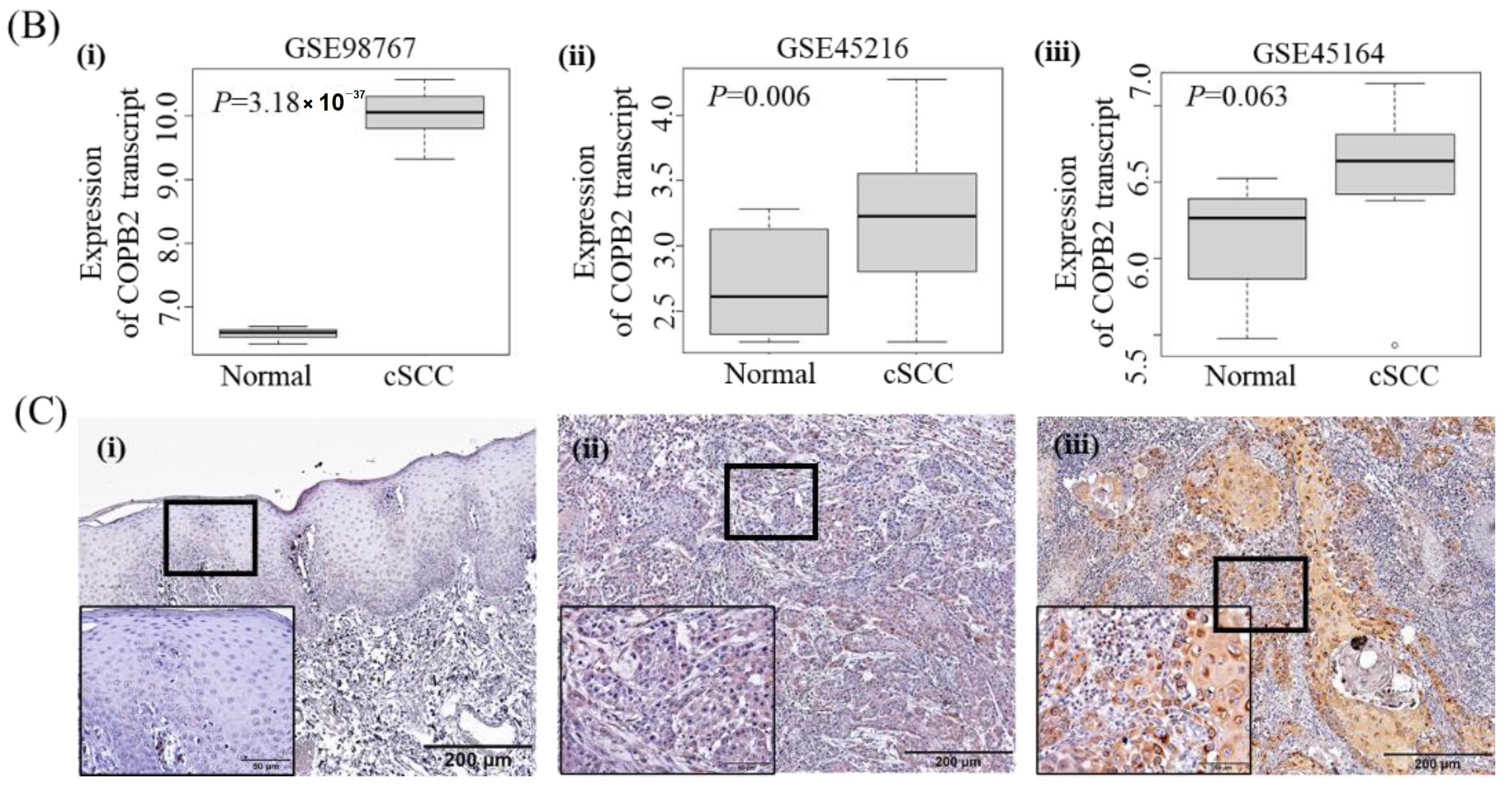
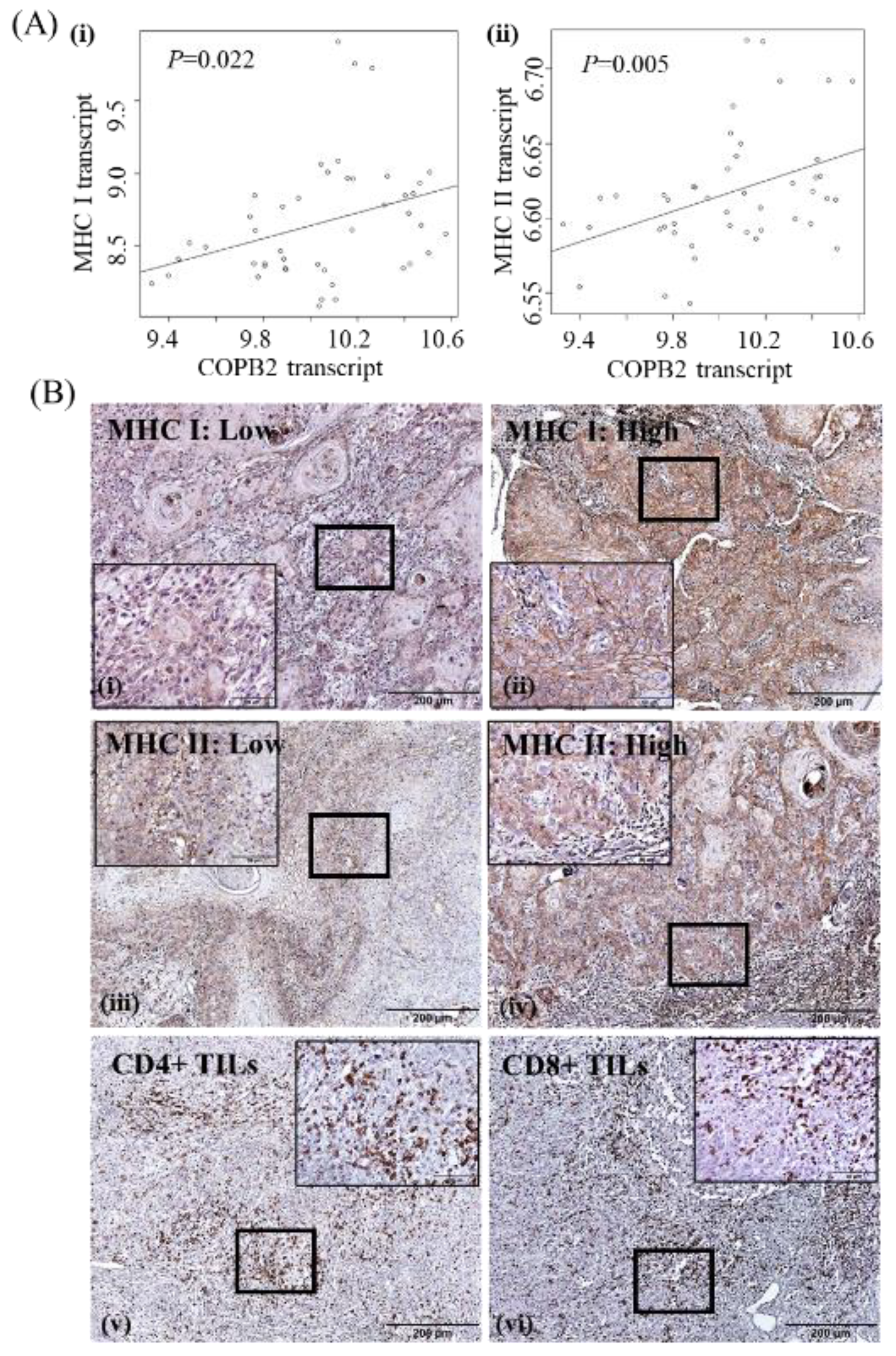
3.3. Clinicopathologic Significance of COPB2 Expression in cSCC
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Y.; Chai, Z.; Wang, M.; Jin, Y.; Yang, A.; Li, M. Copb2 suppresses cell proliferation and induces cell cycle arrest in human colon cancer by regulating cell cycle-related proteins. Exp. Ther. Med. 2018, 15, 777–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pu, X.; Wang, J.; Li, W.; Fan, W.; Wang, L.; Mao, Y.; Yang, S.; Liu, S.; Xu, J.; Lv, Z.; et al. Copb2 promotes cell proliferation and tumorigenesis through up-regulating yap1 expression in lung adenocarcinoma cells. Biomed. Pharmacother. 2018, 103, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Mi, Y.; Yu, M.; Zhang, L.; Sun, C.; Wei, B.; Ding, W.; Zhu, Y.; Tang, J.; Xia, G.; Zhu, L. Copb2 is upregulated in prostate cancer and regulates pc-3 cell proliferation, cell cycle, and apoptosis. Arch. Med. Res. 2016, 47, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.S.; Liu, C.H.; Liu, Z.; Zhu, C.L.; Huang, Q. Downregulation of copb2 by rnai inhibits growth of human cholangiocellular carcinoma cells. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 985–992. [Google Scholar]
- An, C.; Li, H.; Zhang, X.; Wang, J.; Qiang, Y.; Ye, X.; Li, Q.; Guan, Q.; Zhou, Y. Silencing of copb2 inhibits the proliferation of gastric cancer cells and induces apoptosis via suppression of the rtk signaling pathway. Int. J. Oncol. 2019, 54, 1195–1208. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, G.; Li, M.; Du, J.; Wang, M. Copb2 gene silencing inhibits colorectal cancer cell proliferation and induces apoptosis via the jnk/c-jun signaling pathway. PLoS ONE 2020, 15, e0240106. [Google Scholar] [CrossRef]
- Chen, H.L.; Li, J.J.; Jiang, F.; Shi, W.J.; Chang, G.Y. Microrna-4461 derived from bone marrow mesenchymal stem cell exosomes inhibits tumorigenesis by downregulating copb2 expression in colorectal cancer. Biosci. Biotechnol. Biochem. 2020, 84, 338–346. [Google Scholar] [CrossRef]
- Wang, X.; Shi, J.; Niu, Z.; Wang, J.; Zhang, W. Mir-216a-3p regulates the proliferation, apoptosis, migration, and invasion of lung cancer cells via targeting copb2. Biosci. Biotechnol. Biochem. 2020, 84, 2014–2027. [Google Scholar] [CrossRef]
- Pu, X.; Jiang, H.; Li, W.; Xu, L.; Wang, L.; Shu, Y. Upregulation of the coatomer protein complex subunit beta 2 (copb2) gene targets microrna-335-3p in nci-h1975 lung adenocarcinoma cells to promote cell proliferation and migration. Med. Sci. Monit. 2020, 26, e918382. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, X.; Huang, X.; Li, X.D.; Cheng, K.; Yu, H.; Zhou, Y.J.; Lv, P.; Jiang, X.B. High expression of copb2 predicts adverse outcomes: A potential therapeutic target for glioma. CNS Neurosci. Ther. 2020, 26, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, X.; Li, G.; He, J.; Lu, Z.; Yang, Y.; Jiang, Y.; Jiang, L.; Li, F.; Liu, J. Copb2: A novel prognostic biomarker that affects progression of hcc. Biomed. Res. Int. 2021, 2021, 6648078. [Google Scholar] [CrossRef] [PubMed]
- Balkwill, F.R.; Capasso, M.; Hagemann, T. The tumor microenvironment at a glance. J. Cell Sci. 2012, 125, 5591–5596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruno, A.; Ferlazzo, G.; Albini, A.; Noonan, D.M. A think tank of tink/tanks: Tumor-infiltrating/tumor-associated natural killer cells in tumor progression and angiogenesis. J. Natl. Cancer Inst. 2014, 106, dju200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruno, A.; Pagani, A.; Magnani, E.; Rossi, T.; Noonan, D.M.; Cantelmo, A.R.; Albini, A. Inflammatory angiogenesis and the tumor microenvironment as targets for cancer therapy and prevention. Cancer Treat. Res. 2014, 159, 401–426. [Google Scholar]
- Jass, J.R. Lymphocytic infiltration and survival in rectal cancer. J. Clin. Pathol. 1986, 39, 585–589. [Google Scholar] [CrossRef]
- Clark, W.H., Jr.; Elder, D.E.; Guerry, D.t.; Braitman, L.E.; Trock, B.J.; Schultz, D.; Synnestvedt, M.; Halpern, A.C. Model predicting survival in stage i melanoma based on tumor progression. J. Natl. Cancer Inst. 1989, 81, 1893–1904. [Google Scholar] [CrossRef]
- Lomas, A.; Leonardi-Bee, J.; Bath-Hextall, F. A systematic review of worldwide incidence of nonmelanoma skin cancer. Br. J. Dermatol. 2012, 166, 1069–1080. [Google Scholar] [CrossRef]
- Stang, A.; Khil, L.; Kajuter, H.; Pandeya, N.; Schmults, C.D.; Ruiz, E.S.; Karia, P.S.; Green, A.C. Incidence and mortality for cutaneous squamous cell carcinoma: Comparison across three continents. J. Eur. Acad. Dermatol. Venereol. 2019, 33, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.; Kim, J.; Zheng, Z.; Kim, S.K.; Chung, K.Y.; Roh, M.R. Risk factors for recurrence in cutaneous squamous cell carcinoma after mohs micrographic surgery: A retrospective review of 237 asian patients. J. Dermatol. 2020, 47, 72–77. [Google Scholar] [CrossRef]
- Dean, N.R.; Sweeny, L.; Magnuson, J.S.; Carroll, W.R.; Robinson, D.; Desmond, R.A.; Rosenthal, E.L. Outcomes of recurrent head and neck cutaneous squamous cell carcinoma. J. Skin Cancer 2011, 2011, 972497. [Google Scholar] [CrossRef]
- Cohen, D.N.; Lawson, S.K.; Shaver, A.C.; Du, L.; Nguyen, H.P.; He, Q.; Johnson, D.B.; Lumbang, W.A.; Moody, B.R.; Prescott, J.L.; et al. Contribution of beta-hpv infection and uv damage to rapid-onset cutaneous squamous cell carcinoma during braf-inhibition therapy. Clin. Cancer Res. 2015, 21, 2624–2634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Que, S.K.T.; Zwald, F.O.; Schmults, C.D. Cutaneous squamous cell carcinoma: Incidence, risk factors, diagnosis, and staging. J. Am. Acad. Dermatol. 2018, 78, 237–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunstfeld, R.; Wickenhauser, G.; Michaelis, U.; Teifel, M.; Umek, W.; Naujoks, K.; Wolff, K.; Petzelbauer, P. Paclitaxel encapsulated in cationic liposomes diminishes tumor angiogenesis and melanoma growth in a “humanized” scid mouse model. J. Investig. Dermatol. 2003, 120, 476–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zheng, Z.; Shin, Y.K.; Kim, K.Y.; Rha, S.Y.; Noh, S.H.; Chung, H.C.; Jeung, H.C. Angiogenic factor thymidine phosphorylase associates with angiogenesis and lymphangiogenesis in the intestinal-type gastric cancer. Pathology 2014, 46, 316–324. [Google Scholar] [CrossRef]
- Bhandari, A.; Zheng, C.; Sindan, N.; Sindan, N.; Quan, R.; Xia, E.; Thapa, Y.; Tamang, D.; Wang, O.; Ye, X.; et al. Copb2 is up-regulated in breast cancer and plays a vital role in the metastasis via n-cadherin and vimentin. J. Cell Mol. Med. 2019, 23, 5235–5245. [Google Scholar] [CrossRef] [Green Version]
- Slaughter, D.P.; Southwick, H.W.; Smejkal, W. Field cancerization in oral stratified squamous epithelium; clinical implications of multicentric origin. Cancer 1953, 6, 963–968. [Google Scholar] [CrossRef]
- Wang, Q.; Zhou, D.; Wu, F.; Liang, Q.; He, Q.; Peng, M.; Yao, T.; Hu, Y.; Qian, B.; Tang, J.; et al. Immune microenvironment signatures as biomarkers to predict early recurrence of stage ia-b lung cancer. Front. Oncol. 2021, 11, 680287. [Google Scholar] [CrossRef]
- Takeshita, T.; Yan, L.; Asaoka, M.; Rashid, O.; Takabe, K. Late recurrence of breast cancer is associated with pro-cancerous immune microenvironment in the primary tumor. Sci. Rep. 2019, 9, 16942. [Google Scholar] [CrossRef] [Green Version]
- Watermann, C.; Pasternack, H.; Idel, C.; Ribbat-Idel, J.; Bragelmann, J.; Kuppler, P.; Offermann, A.; Jonigk, D.; Kuhnel, M.P.; Schrock, A.; et al. Recurrent hnscc harbor an immunosuppressive tumor immune microenvironment suggesting successful tumor immune evasion. Clin. Cancer Res. 2021, 27, 632–644. [Google Scholar] [CrossRef]
- Holzel, M.; Bovier, A.; Tuting, T. Plasticity of tumour and immune cells: A source of heterogeneity and a cause for therapy resistance? Nat. Rev. Cancer 2013, 13, 365–376. [Google Scholar] [CrossRef]
- Ahn, S.Y.; Kim, N.H.; Lee, K.; Cha, Y.H.; Yang, J.H.; Cha, S.Y.; Cho, E.S.; Lee, Y.; Cha, J.S.; Cho, H.S.; et al. Niclosamide is a potential therapeutic for familial adenomatosis polyposis by disrupting axin-gsk3 interaction. Oncotarget 2017, 8, 31842–31855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakabayashi, O.; Yamazaki, K.; Oizumi, S.; Hommura, F.; Kinoshita, I.; Ogura, S.; Dosaka-Akita, H.; Nishimura, M. Cd4+ t cells in cancer stroma, not cd8+ t cells in cancer cell nests, are associated with favorable prognosis in human non-small cell lung cancers. Cancer Sci. 2003, 94, 1003–1009. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Qiu, S.J.; Fan, J.; Zhou, J.; Wang, X.Y.; Xiao, Y.S.; Xu, Y.; Li, Y.W.; Tang, Z.Y. Intratumoral balance of regulatory and cytotoxic t cells is associated with prognosis of hepatocellular carcinoma after resection. J. Clin. Oncol. 2007, 25, 2586–2593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bretz, A.C.; Parnitzke, U.; Kronthaler, K.; Dreker, T.; Bartz, R.; Hermann, F.; Ammendola, A.; Wulff, T.; Hamm, S. Domatinostat favors the immunotherapy response by modulating the tumor immune microenvironment (time). J. Immunother. Cancer 2019, 7, 294. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinicopathological Variables | No. Patients (%) |
|---|---|
| Total cases of normal skin tissue | 5 |
| Median age, years | 43 (32–89) |
| Gender | |
| Male | 2 (40) |
| Female | 3 (60) |
| Location | |
| Head and neck | 1 (20) |
| Acral | 1 (20) |
| Extremity | 2 (40) |
| Trunk | 1 (20) |
| Total cases of cSCC patients | 95 |
| Age, years | |
| Median age (range) | 76 (30–98) |
| ≤60 | 12 (12.6) |
| >60 | 83 (87.4) |
| Gender | |
| Male | 41 (43.2) |
| Female | 54 (56.8) |
| Location | |
| Head and neck | 79 (83.2) |
| Acral | 12 (12.6) |
| Extremity | 2 (2.1) |
| Trunk | 2 (2.1) |
| Tumor size, cm | |
| ≤2cm | 62 (65.3) |
| >2cm | 33 (34.7) |
| Histologic grade | |
| WD | 54 (56.8) |
| MD | 32 (33.7) |
| PD | 9 (9.5) |
| Recurrence | |
| Yes | 16 (16.8) |
| No | 79 (83.2) |
| Variables | Total, n (%) | COPB2 | p Value | |
|---|---|---|---|---|
| Low | High | |||
| Age | ||||
| ≤60 | 12 (12.6) | 7 (58.3) | 5 (41.7) | |
| >60 | 83 (87.4) | 66 (79.5) | 17 (20.5) | 0.141 |
| Gender | ||||
| Male | 41 (43.2) | 29 (70.7) | 12 (29.3) | |
| Female | 54 (56.8) | 44 (81.5) | 10 (18.5) | 0.232 |
| Lesion site | ||||
| Head and neck | 79 (83.2) | 60 (75.9) | 19 (24.1) | 1.000 |
| Acral | 12 (12.6) | 9 (75.0) | 3 (25.0) | |
| Extremity | 2 (2.1) | 2 (100) | 0(0) | |
| Trunk | 2 (2.1) | 2 (100) | 0(0) | |
| Tumor size, cm | ||||
| ≤2 | 62 (65.3) | 46 (74.2) | 16 (25.8) | 0.454 |
| >2 | 33 (34.7) | 27 (81.8) | 6 (18.2) | |
| Histologic grade | ||||
| WD | 54 (56.8) | 40 (74.1) | 14 (25.9) | 0.872 |
| MD | 32 (33.7) | 26 (81.3) | 6 (18.8) | |
| PD | 9 (9.5) | 7 (77.8) | 2 (22.2) | |
| MHC I | ||||
| Low | 78 (82.1) | 63 (80.8) | 15 (19.2) | 0.063 |
| High | 17 (17.9) | 10 (58.8) | 7 (41.2) | |
| MHC II | ||||
| Low | 68 (71.6) | 56 (82.4) | 12 (17.6) | 0.059 |
| High | 27 (28.4) | 17 (63.0) | 10 (37.0) | |
| Intra CD4+TIL | ||||
| Low | 68 (71.6) | 59 (86.8) | 9 (13.2) | 0.001 |
| High | 27 (28.4) | 14 (51.9) | 13 (48.1) | |
| Peri CD4+TIL | ||||
| Low | 51 (53.7) | 44 (86.3) | 7 (13.7) | 0.018 |
| High | 44 (46.3) | 29 (65.9) | 15 (34.1) | |
| Intra CD8+TIL | ||||
| Low | 61 (64.2) | 53 (86.9) | 8 (13.1) | 0.004 |
| High | 34 (35.8) | 20 (58.8) | 14 (41.2) | |
| Peri CD8+TIL | ||||
| Low | 52 (54.7) | 46 (88.5) | 6 (11.5) | 0.006 |
| High | 43 (45.3) | 27 (62.8) | 16 (37.2) | |
| Variables | Univariate | p | Multivariate | p |
|---|---|---|---|---|
| Odds Ratio (95% CI) | Odds Ratio (95% CI) | |||
| Age | 0.991 (0.953–1.031) | 0.644 | 0.994 (0.924–1.069) | 0.873 |
| Gender | ||||
| Male | 1 | 1 | ||
| Female | 1.770 (0.622–5.037) | 0.285 | 2.800 (0.415–18.887) | 0.290 |
| Lesion site | ||||
| Head and neck | 1 | 1 | ||
| Acral | 1.287 (0.356–4.652) | 0.701 | 3.349 (0.382–29.374) | 0.275 |
| Extremity | 0.000 (0.000–0.000) | 0.995 | 0.000 (0.000–0.000) | 0.998 |
| Trunk | 0.000 (0.000–0.000) | 0.986 | 0.000 (0.000–0.000) | 0.983 |
| Tumor size, cm | ||||
| ≤2 | 1 | 1 | ||
| >2 | 1.614 (0.598–4.357) | 0.345 | 4.812 (1.025–22.588) | 0.046 |
| Histologic grade | ||||
| WD | 1 | 1 | ||
| MD | 1.102 (0.322–3.772) | 0.877 | 3.452 (0.543–21.936) | 0.189 |
| PD | 3.130 (0.626–15.646) | 0.165 | 13.576 (1.151–160.104) | 0.038 |
| COPB2 | ||||
| Low | 1 | 1 | ||
| High | 6.734 (2.328–19.478) | <0.001 | 10.905 (1.714–69.372) | 0.011 |
| MHC I | ||||
| Low | 1 | 1 | ||
| High | 3.403 (1.270–9.121) | 0.015 | 3.172 (0.593–16.975) | 0.177 |
| MHC II | ||||
| Low | 1 | 1 | ||
| High | 3.701 (1.335–10.261) | 0.012 | 1.849 (0.289–11.830) | 0.516 |
| Intra CD4+TIL | ||||
| Low | 1 | 1 | ||
| High | 3.611 (1.305–9.993) | 0.013 | 2.473 (0.375––16.325) | 0.347 |
| Peri CD4+TIL | ||||
| Low | 1 | 1 | ||
| High | 0.984 (0.363–2.666) | 0.984 | 0.218 (0.042–1.123) | 0.069 |
| Intra CD8+TIL | ||||
| Low | 1 | 1 | ||
| High | 2.798 (1.007–7.773) | 0.048 | 4.186 (0.851–20.579) | 0.078 |
| Peri CD8+TIL | ||||
| Low | 1 | 1 | ||
| High | 2.017 (0.692–5.875) | 0.199 | 0.489 (0.083–2.875) | 0.429 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, T.; Kim, K.-Y.; Oh, Y.; Jeung, H.C.; Chung, K.Y.; Roh, M.R.; Zhang, X. Implication of COPB2 Expression on Cutaneous Squamous Cell Carcinoma Pathogenesis. Cancers 2022, 14, 2038. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14082038
Chen T, Kim K-Y, Oh Y, Jeung HC, Chung KY, Roh MR, Zhang X. Implication of COPB2 Expression on Cutaneous Squamous Cell Carcinoma Pathogenesis. Cancers. 2022; 14(8):2038. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14082038
Chicago/Turabian StyleChen, Taiqin, Ki-Yeol Kim, Yeongjoo Oh, Hei Cheul Jeung, Kee Yang Chung, Mi Ryung Roh, and Xianglan Zhang. 2022. "Implication of COPB2 Expression on Cutaneous Squamous Cell Carcinoma Pathogenesis" Cancers 14, no. 8: 2038. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14082038