
Chitosan-Decorated Copper Oxide Nanocomposite: Investigation of Its Antifungal Activity against Tomato Gray Mold Caused by Botrytis cinerea


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials Used in This Study
2.2. Isolation and Molecular Characterization of B. cinerea
2.3. Preparation and Characterization of CH@CuO NPs
2.4. Effect of CH@CuO NPs on the Mycelial Growth of B. cinerea
2.5. Effect of CH@CuO NPs on the Germination of Conidia and Sclerotia of B. cinerea
2.6. Effect of CH@CuO NPs on the Plasma Membrane Integrity of B. cinerea Conidia
2.7. Effect of CH@CuO NPs on the Mycelial Electrolyte Leakage of B. cinerea
2.8. Effect of Heating on the Stability of CH@CuO NPs against B. cinerea
2.9. Effect of CH@CuO NPs on the Gray Mold Disease
2.9.1. Detached Tomato Leaves Experiment
2.9.2. Ripe Tomato Fruit Experiment
2.10. Greenhouse Experiments
Effect of CH@CuO NPs on Tomato Gray Mold
2.11. Data Analysis
3. Results
3.1. Characterization of B. cinerea
3.2. Synthesis and Characterization of CH@CuO NPs
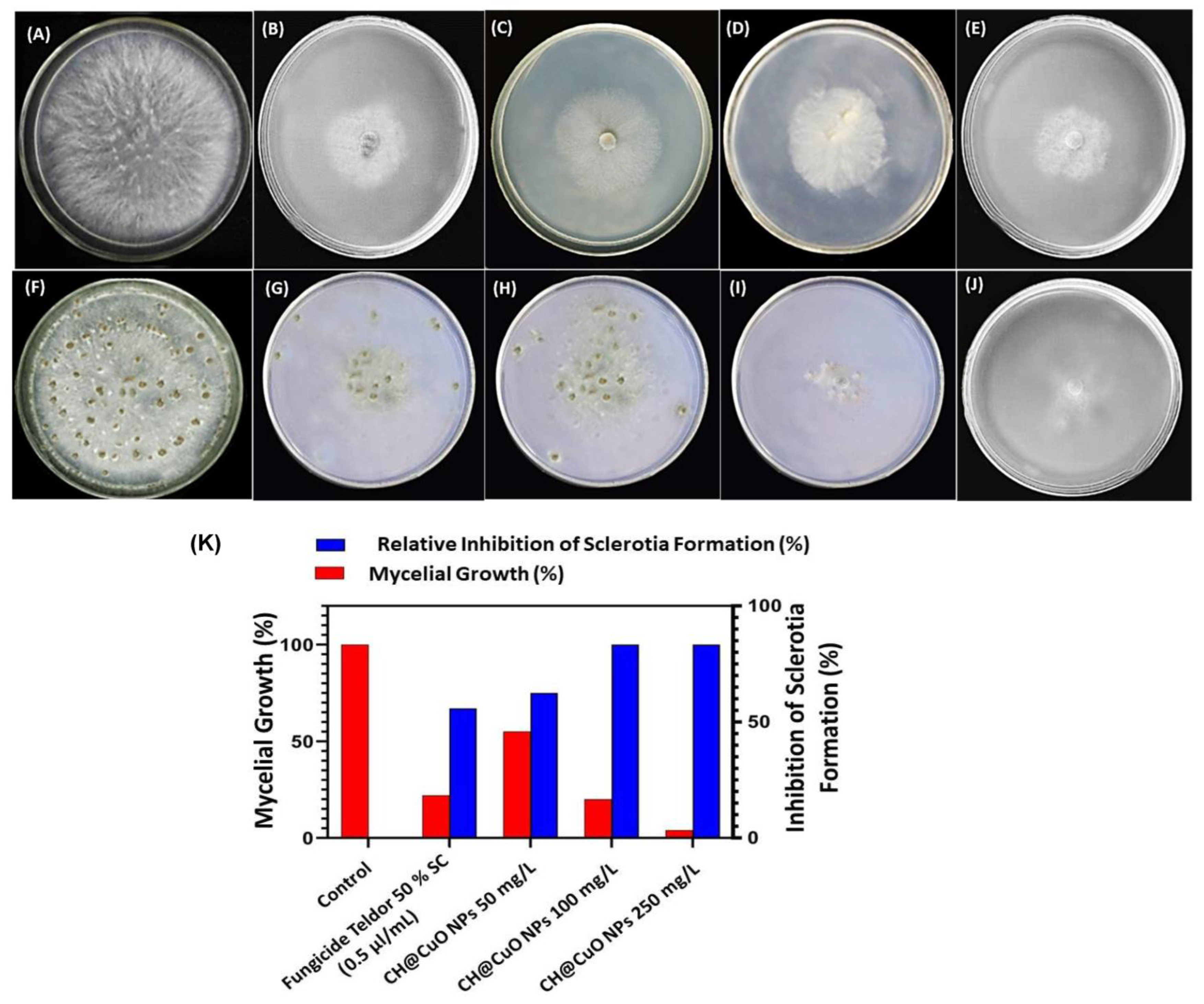
3.3. Antifungal Activity of CH@CuO NPs against Mycelial Growth and Sclerotia Formation of B. cinerea
3.4. CH@CuO NPs Inhibited Conidia Germination of B. cinerea
3.5. Effect of CH@CuO NPs on B. cinerea’s Mycelial Electrolyte Leakage
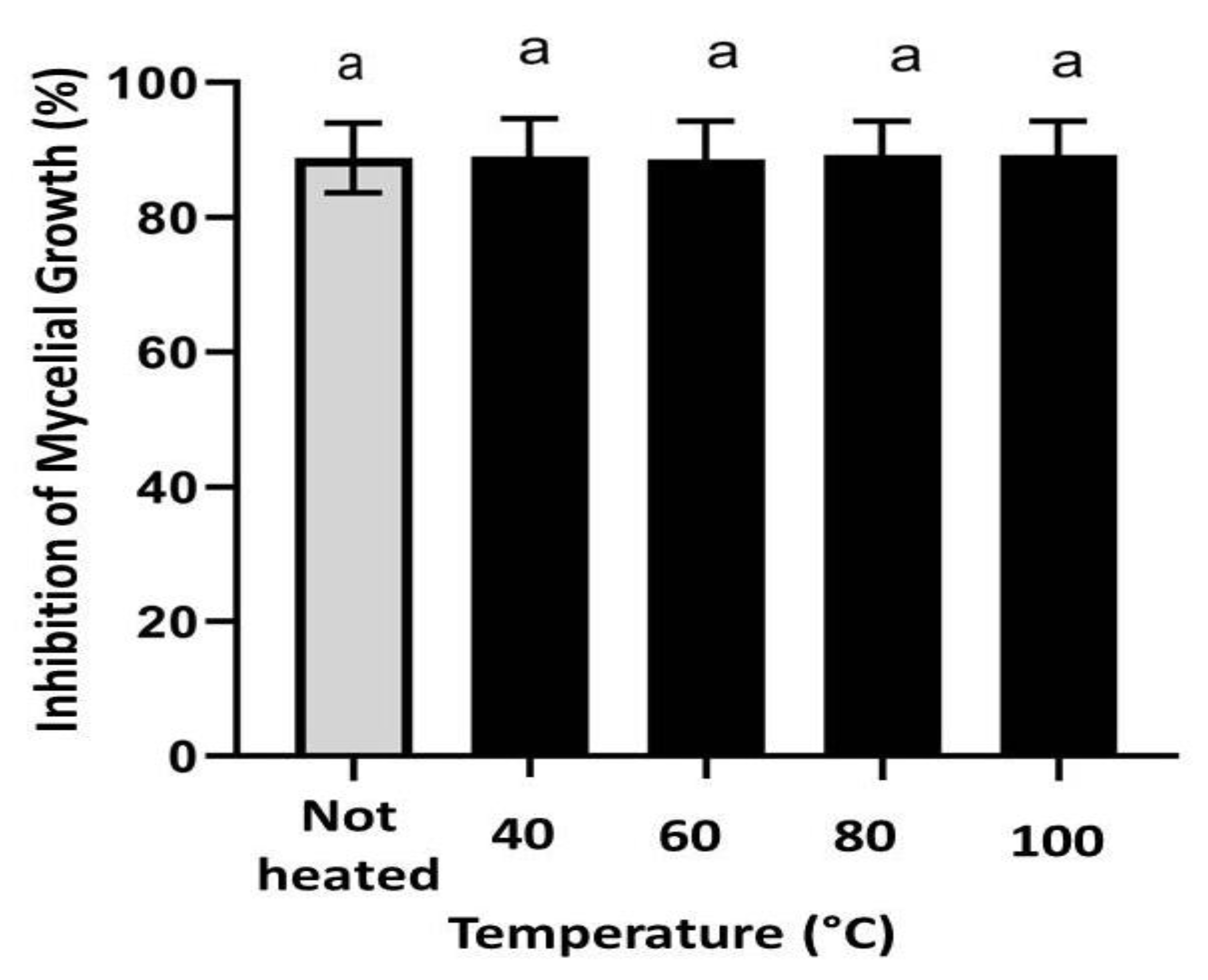
3.6. Antifungal Activity of CH@CuO NPs Retains Its Activity at High Temperatures
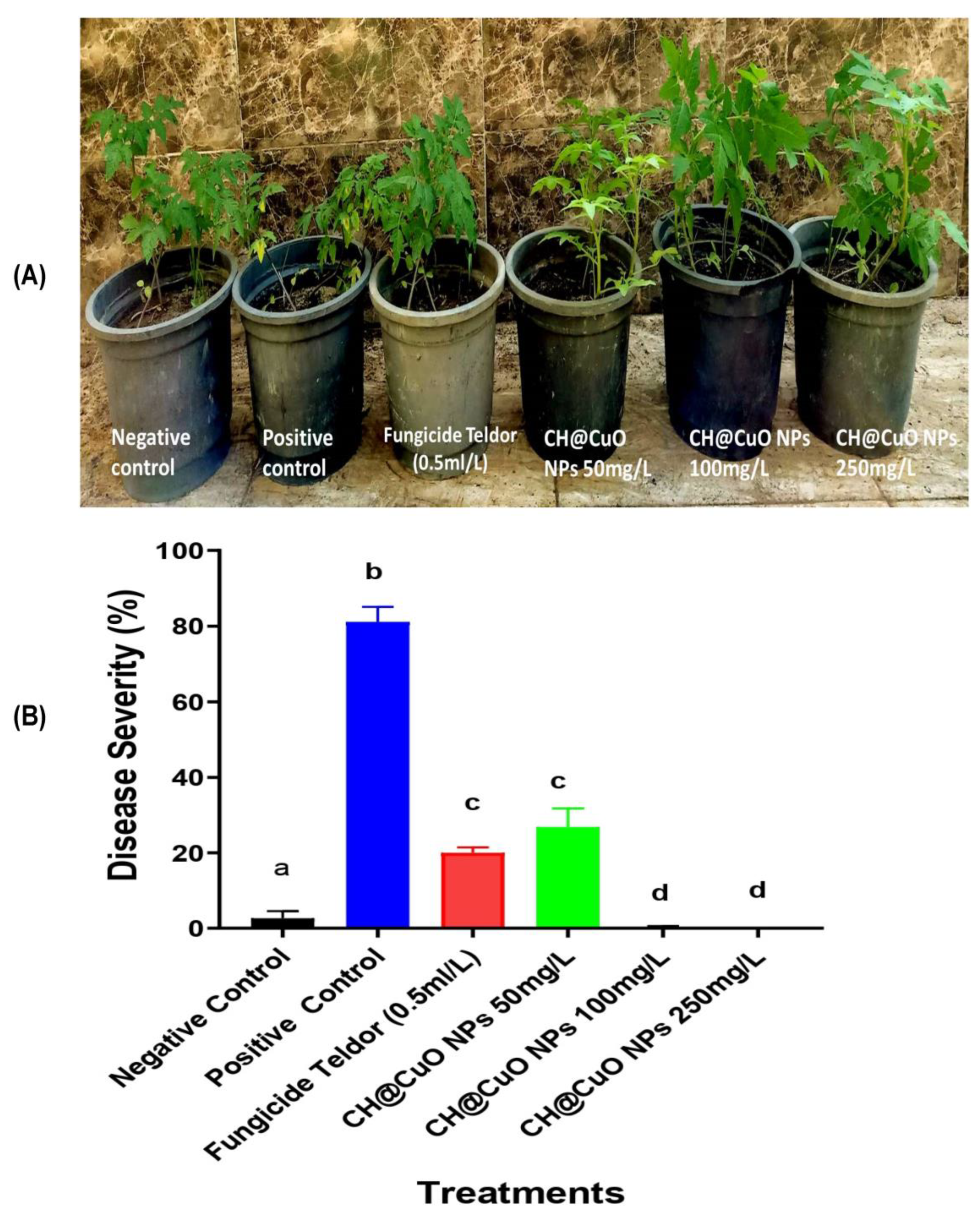
3.7. Fungicidal Activity of CH@CuO NPs in Reducing/Controlling Gray Mold Disease on Detached Tomato Leaves and Their Fruits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heuvelink, E.; Dorais, M. Crop Growth and Yield. Crop Prod. Sci. Hortic. 2005, 13, 85. [Google Scholar]
- Agrios, G.N. Plant Pathology, 5th ed.; Elsevier: Amsterdam, The Netherlands; Academic Press: London, UK, 2005; pp. 398–401. [Google Scholar]
- Elad, Y.; Williamson, B.; Tudzynski, P.; Delen, N. Botrytis spp. and Diseases They Cause in Agricultural Aystems—An Introduction. In Botrytis: Biology, Pathology and Control; Elad, Y., Williamson, B., Tudzynski, P., Delen, N., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 1–8. [Google Scholar]
- Elad, Y.; Pertot, I.; Cotes Prado, A.M.; Stewart, A. Plant Hosts of Botrytis Spp. In Botrytis—the Fungus, the Pathogen and Its Management in Agricultural Systems; Fillinger, S., Elad, Y., Eds.; Springer: Cham, Switzerland, 2016; pp. 413–486. [Google Scholar]
- Wang, J.; Xia, X.-M.; Wang, H.-Y.; Li, P.-P.; Wang, K.-Y. Inhibitory Effect of Lactoferrin against Gray Mould on Tomato Plants Caused by Botrytis cinerea and Possible Mechanisms of Action. Int. J. Food Microbiol. 2013, 161, 151–157. [Google Scholar] [CrossRef]
- Hua, L.; Yong, C.; Zhanquan, Z.; Boqiang, L.; Guozheng, Q.; Shiping, T. Pathogenic Mechanisms and Control Strategies of Botrytis cinerea Causing Post-Harvest Decay in Fruits and Vegetables. Food Qual. Saf. 2018, 2, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Ciliberti, N.; Fermaud, M.; Roudet, J.; Rossi, V. Environmental Conditions Affect Botrytis cinerea Infection of Mature Grape Berries More than the Strain or Transposon Genotype. Phytopathology 2015, 105, 1090–1096. [Google Scholar] [CrossRef] [Green Version]
- Elmer, P.A.G.; Michailides, T.J. Epidemiology of Botrytis cinerea in Orchard and Vine Crops. In Botrytis: Biology, Pathology and Control; Elad, Y., Williamson, B., Tudzynski, P., Delen, N., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 243–272. [Google Scholar]
- Edder, P.; Ortelli, D.; Viret, O.; Cognard, E.; De Montmollin, A.; Zali, O. Control Strategies against Grey Mould (Botrytis cinerea Pers.: Fr) and Corresponding Fungicide Residues in Grapes and Wines. Food Addit. Contam. 2009, 26, 719–725. [Google Scholar] [CrossRef]
- Buonaurio, R.; Moretti, C.; Bertona, A.; Scarponi, L. Investigations on the Systemic Acquired Resistance Induced by Acibenzolar-S-Methyl in Tomato Plants Against Pseudomonas syringae pv. Tomato. In Pseudomonas syringae and Related Pathogens; Iacobellis, N.S., Collmer, A., Hutcheson, S.W., Mansfield, J.W., Morris, C.E., Murillo, J., Schaad, N.W., Stead, D.E., Surico, G., Ullrich, M.S., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 475–482. [Google Scholar]
- Kretschmer, M.; Leroch, M.; Mosbach, A.; Walker, A.-S.; Fillinger, S.; Mernke, D.; Schoonbeek, H.-J.; Pradier, J.-M.; Leroux, P.; De Waard, M.A. Fungicide-Driven Evolution and Molecular Basis of Multidrug Resistance in Field Populations of the Grey Mould Fungus Botrytis cinerea. PLoS Pathog. 2009, 5, e1000696. [Google Scholar] [CrossRef]
- Myresiotis, C.K.; Karaoglanidis, G.S.; Tzavella-Klonari, K. Resistance of Botrytis Cinerea Isolates from Vegetable Crops to Anilinopyrimidine, Phenylpyrrole, Hydroxyanilide, Benzimidazole, and Dicarboximide Fungicides. Plant Dis. 2007, 91, 407–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosbach, A.; Edel, D.; Farmer, A.D.; Widdison, S.; Barchietto, T.; Dietrich, R.A.; Corran, A.; Scalliet, G. Anilinopyrimidine Resistance in Botrytis Cinerea Is Linked to Mitochondrial Function. Front. Microbiol. 2017, 8, 2361. [Google Scholar] [CrossRef] [PubMed]
- Adnan, M.; Hamada, M.S.; Li, G.Q.; Luo, C.X. Detection and Molecular Characterization of Resistance to the Dicarboximide and Benzamide Fungicides in Botrytis Cinerea from Tomato in Hubei Province, China. Plant Dis. 2018, 102, 1299–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismail, A.M. Efficacy of Copper Oxide and Magnesium Oxide Nanoparticles on Controlling Black Scurf Disease on Potato. Egypt. J. Phytopathol. 2021, 49, 116–130. [Google Scholar] [CrossRef]
- Dimkpa, C.O.; McLean, J.E.; Britt, D.W.; Anderson, A.J. Antifungal Activity of ZnO Nanoparticles and Their Interactive Effect with a Biocontrol Bacterium on Growth Antagonism of the Plant Pathogen Fusarium Graminearum. Biometals 2013, 26, 913–924. [Google Scholar] [CrossRef]
- Ismail, A.M.; El-Gawad, A.; Mona, E. Antifungal Activity of MgO and ZnO Nanoparticles against Powdery Mildew of Pepper under Greenhouse Conditions. Egypt. J. Agric. Res. 2021, 99, 421–434. [Google Scholar]
- Krishna, V.; Pumprueg, S.; Lee, S.-H.; Zhao, J.; Sigmund, W.; Koopman, B.; Moudgil, B.M. Photocatalytic Disinfection with Titanium Dioxide Coated Multi-Wall Carbon Nanotubes. Process Saf. Environ. Prot. 2005, 83, 393–397. [Google Scholar] [CrossRef]
- El-Gazzar, N.; Ismail, A.M. The Potential Use of Titanium, Silver and Selenium Nanoparticles in Controlling Leaf Blight of Tomato Caused by Alternaria alternata. Biocatal. Agric. Biotechnol. 2020, 27, 101708. [Google Scholar] [CrossRef]
- El-Gazzar, N.; Almaary, K.; Ismail, A.; Polizzi, G. Influence of Funneliformis Mosseae Enhanced with Titanium Dioxide Nanoparticles (TiO2NPs) on Phaseolus vulgaris L. under Salinity Stress. PLoS ONE 2020, 15, e0235355. [Google Scholar] [CrossRef]
- El-Ganainy, S.M.; El-Bakery, A.M.; Hafez, H.M.; Ismail, A.M.; El-Abdeen, A.Z.; Ata, A.A.E.; Elraheem, O.A.Y.A.; El Kady, Y.M.Y.; Hamouda, A.F.; El-Beltagi, H.S. Humic Acid-Coated Fe3O4 Nanoparticles Confer Resistance to Acremonium Wilt Disease and Improve Physiological and Morphological Attributes of Grain Sorghum. Polymers (Basel) 2022, 14, 3099. [Google Scholar] [CrossRef]
- Mosa, M.A.; Youssef, K. Topical Delivery of Host Induced RNAi Silencing by Layered Double Hydroxide Nanosheets: An Efficient Tool to Decipher Pathogenicity Gene Function of Fusarium Crown and Root Rot in Tomato. Physiol. Mol. Plant Pathol. 2021, 115, 101684. [Google Scholar] [CrossRef]
- Mosa, M.A.; El-Abeid, S.E.; Khalifa, M.M.A.; Elsharouny, T.H.; El-Baz, S.M.; Ahmed, A.Y. Smart PH Responsive System Based on Hybrid Mesoporous Silica Nanoparticles for Delivery of Fungicide to Control Fusarium Crown and Root Rot in Tomato. J. Plant Pathol. 2022, 104, 979–992. [Google Scholar] [CrossRef]
- Cordero, T.; Mohamed, M.A.; Lopez-Moya, J.-J.; Daròs, J.-A. A Recombinant Potato Virus y Infectious Clone Tagged with the Rosea1 Visual Marker (Pvy-Ros1) Facilitates the Analysis of Viral Infectivity and Allows the Production of Large Amounts of Anthocyanins in Plants. Front. Microbiol. 2017, 8, 611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Abeid, S.E.; Ahmed, Y.; Daròs, J.-A.; Mohamed, M.A. Reduced Graphene Oxide Nanosheet-Decorated Copper Oxide Nanoparticles: A Potent Antifungal Nanocomposite against Fusarium Root Rot and Wilt Diseases of Tomato and Pepper Plants. Nanomaterials 2020, 10, 1001. [Google Scholar] [CrossRef] [PubMed]
- Youssef, K.; de Oliveira, A.G.; Tischer, C.A.; Hussain, I.; Roberto, S.R. Synergistic Effect of a Novel Chitosan/Silica Nanocomposites-Based Formulation against Gray Mold of Table Grapes and Its Possible Mode of Action. Int. J. Biol. Macromol. 2019, 141, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, S.; Patra, P.; Mitra, S.; Dey, K.K.; Jain, S.; Sarkar, S.; Roy, S.; Palit, P.; Goswami, A. Manganese Nanoparticles: Impact on Non-Nodulated Plant as a Potent Enhancer in Nitrogen Metabolism and Toxicity Study Both In Vivo and In Vitro. J. Agric. Food Chem. 2014, 62, 8777–8785. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, R.C.; Kumaraswamy, R.V.; Kumari, S.; Sharma, S.S.; Pal, A.; Raliya, R.; Biswas, P.; Saharan, V. Cu-Chitosan Nanoparticle Boost Defense Responses and Plant Growth in Maize (Zea mays L.). Sci. Rep. 2017, 7, 9754. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, K.R.; Cowley, S.; Martinez, F.O.; Shaw, M.; Minger, S.L.; James, W. Homogeneous Monocytes and Macrophages from Human Embryonic Stem Cells Following Coculture-Free Differentiation in M-CSF and IL-3. Exp. Hematol. 2008, 36, 1167–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubina, M.S.; Vasil’kov, A.Y.; Naumkin, A.V.; Shtykova, E.V.; Abramchuk, S.S.; Alghuthaymi, M.A.; Abd-Elsalam, K.A. Synthesis and Characterization of Chitosan–Copper Nanocomposites and Their Fungicidal Activity against Two Sclerotia-Forming Plant Pathogenic Fungi. J. Nanostructure Chem. 2017, 7, 249–258. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, X.; Han, X.; Du, Y. Antifungal Activity of Oligochitosan against Phytophthora capsici and Other Plant Pathogenic Fungi in Vitro. Pestic. Biochem. Physiol. 2007, 87, 220–228. [Google Scholar] [CrossRef]
- Muzzarelli, R.A.A.; Muzzarelli, C.; Tarsi, R.; Miliani, M.; Gabbanelli, F.; Cartolari, M. Fungistatic Activity of Modified Chitosans against Saprolegnia parasitica. Biomacromolecules 2001, 2, 165–169. [Google Scholar] [CrossRef]
- El Hadrami, A.; Adam, L.R.; El Hadrami, I.; Daayf, F. Chitosan in Plant Protection. Mar. Drugs 2010, 8, 968–987. [Google Scholar] [CrossRef]
- Liu, H.; Bao, J.; Du, Y.; Zhou, X.; Kennedy, J.F. Effect of Ultrasonic Treatment on the Biochemphysical Properties of Chitosan. Carbohydr. Polym. 2006, 64, 553–559. [Google Scholar] [CrossRef]
- Kruk, T.; Szczepanowicz, K.; Stefańska, J.; Socha, R.P.; Warszyński, P. Synthesis and Antimicrobial Activity of Monodisperse Copper Nanoparticles. Colloids Surf. B 2015, 128, 17–22. [Google Scholar] [CrossRef]
- Kanhed, P.; Birla, S.; Gaikwad, S.; Gade, A.; Seabra, A.B.; Rubilar, O.; Duran, N.; Rai, M. In Vitro Antifungal Efficacy of Copper Nanoparticles against Selected Crop Pathogenic Fungi. Mater. Lett. 2014, 115, 13–17. [Google Scholar] [CrossRef]
- Saharan, V.; Mehrotra, A.; Khatik, R.; Rawal, P.; Sharma, S.S.; Pal, A. Synthesis of Chitosan Based Nanoparticles and Their In Vitro Evaluation against Phytopathogenic Fungi. Int. J. Biol. Macromol. 2013, 62, 677–683. [Google Scholar] [CrossRef]
- Saharan, V.; Sharma, G.; Yadav, M.; Choudhary, M.K.; Sharma, S.S.; Pal, A.; Raliya, R.; Biswas, P. Synthesis and In Vitro Antifungal Efficacy of Cu–Chitosan Nanoparticles against Pathogenic Fungi of Tomato. Int. J. Biol. Macromol. 2015, 75, 346–353. [Google Scholar] [CrossRef]
- Saharan, V.; Kumaraswamy, R.V.; Choudhary, R.C.; Kumari, S.; Pal, A.; Raliya, R.; Biswas, P. Cu-Chitosan Nanoparticle Mediated Sustainable Approach to Enhance Seedling Growth in Maize by Mobilizing Reserved Food. J. Agric. Food Chem. 2016, 64, 6148–6155. [Google Scholar] [CrossRef]
- Arjunan, N.; Singaravelu, C.M.; Kulanthaivel, J.; Kandasamy, J. A Potential Photocatalytic, Antimicrobial and Anticancer Activity of Chitosan-Copper Nanocomposite. Int. J. Biol. Macromol. 2017, 104, 1774–1782. [Google Scholar] [CrossRef] [PubMed]
- Nelson, P.E.; Toussoun, T.A.; Marasas, W.F.O. Fusarium Species: An Illustrated Manual for Identification; Pennsylvania State University Press: University Park, PA, USA, 1983. [Google Scholar]
- Hirano, Y.; Arie, T. PCR-Based Differentiation of Fusarium oxysporum f. sp. lycopersici and Radicis-Lycopersici and Races of F. oxysporum f. sp. lycopersici. J. Gen. Plant Pathol. 2006, 72, 273–283. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J.; Nnis, M.A.; Gelfand, D.H.; Sninsky, J.J. PCR Protocols—A Guide to Methods and Applications; The University of Michigan: Ann Arbor, MI, USA; Academic Press: Cambridge, UK, 1990. [Google Scholar]
- Çolak, A.; Bicici, M. PCR Detection of Fusarium oxysporum f.sp. Radicis-Lycopersici and Races of F. oxysporum f. sp. lycopersici of Tomato in Protected Tomato-Growing Areas of the Eastern Mediterranean Region of Turkey. Turkish J. Agric. For. 2013, 37, 457–467. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Oh, J.-W.; Chun, S.C.; Chandrasekaran, M. Preparation and in Vitro Characterization of Chitosan Nanoparticles and Their Broad-Spectrum Antifungal Action Compared to Antibacterial Activities against Phytopathogens of Tomato. Agronomy 2019, 9, 21. [Google Scholar] [CrossRef] [Green Version]
- Tomaszewska, E.; Soliwoda, K.; Kadziola, K.; Tkacz-Szczesna, B.; Celichowski, G.; Cichomski, M.; Szmaja, W.; Grobelny, J. Detection Limits of DLS and UV-Vis Spectroscopy in Characterization of Polydisperse Nanoparticles Colloids. J. Nanomater. 2013, 2013, 60. [Google Scholar] [CrossRef] [Green Version]
- Boukaew, S.; Prasertsan, P.; Troulet, C.; Bardin, M. Biological Control of Tomato Gray Mold Caused by Botrytis cinerea by Using Streptomyces spp. BioControl 2017, 62, 793–803. [Google Scholar] [CrossRef]
- Vicedo, B.; Leyva, M.d.l.O.; Flors, V.; Finiti, I.; Amo, G.d.; Walters, D.; Real, M.D.; García-Agustín, P.; González-Bosch, C. Control of the Phytopathogen Botrytis cinerea Using Adipic Acid Monoethyl Ester. Arch. Microbiol. 2006, 184, 316–326. [Google Scholar] [CrossRef]
- Yang, F.; Abdelnabby, H.; Xiao, Y. A Mutant of the Nematophagous Fungus Paecilomyces Lilacinus (Thom) is a Novel Biocontrol Agent for Sclerotinia sclerotiorum. Microb. Pathog. 2015, 89, 169–176. [Google Scholar] [CrossRef]
- Abd-El-Khair, H.; El-Gamal Nadia, G. Effects of Aqueous Extracts of Some Plant Species against Fusarium solani and Rhizoctonia solani in Phaseolus vulgaris Plants. Arch. Phytopathol. Plant Prot. 2011, 44, 1–16. [Google Scholar] [CrossRef]
- Wang, S.; Ruan, C.; Yi, L.; Deng, L.; Yao, S.; Zeng, K. Biocontrol Ability and Action Mechanism of Metschnikowia Citriensis against Geotrichum citri-aurantii Causing Sour Rot of Postharvest Citrus Fruit. Food Microbiol. 2020, 87, 103375. [Google Scholar] [CrossRef]
- Gao, P.; Qin, J.; Li, D.; Zhou, S. Inhibitory Effect and Possible Mechanism of a Pseudomonas Strain QBA5 against Gray Mold on Tomato Leaves and Fruits Caused by Botrytis cinerea. PLoS ONE 2018, 13, e0190932. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Yao, S.; Deng, L.; Ming, J.; Zeng, K. Different Mechanisms of Action of Isolated Epiphytic Yeasts against Penicillium digitatum and Penicillium italicum on Citrus Fruit. Postharvest Biol. Technol. 2019, 152, 100–110. [Google Scholar] [CrossRef]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 7th ed.; Iowa State University Press: Ames, IA, USA, 1980. [Google Scholar]
- Morones, J.R.; Elechiguerra, J.L.; Camacho, A.; Holt, K.; Kouri, J.B.; Ramírez, J.T.; Yacaman, M.J. The Bactericidal Effect of Silver Nanoparticles. Nanotechnology 2005, 16, 2346. [Google Scholar] [CrossRef] [Green Version]
- Raffi, M.; Mehrwan, S.; Bhatti, T.M.; Akhter, J.I.; Hameed, A.; Yawar, W. Investigations into the Antibacterial Behavior of Copper Nanoparticles against Escherichia coli. Ann. Microbiol. 2010, 60, 75–80. [Google Scholar] [CrossRef]
- Huang, Y.; Qin, Y.; Zhou, Y.; Niu, H.; Yu, Z.-Z.; Dong, J.-Y. Polypropylene/Graphene Oxide Nanocomposites Prepared by in Situ Ziegler− Natta Polymerization. Chem. Mater. 2010, 22, 4096–4102. [Google Scholar] [CrossRef]
- Chen, J.; Wang, X.; Han, H. A New Function of Graphene Oxide Emerges: Inactivating Phytopathogenic Bacterium Xanthomonas oryzae pv. oryzae. J. Nanopart. Res. 2013, 15, 1658. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, L.; Chen, G. Facile Preparation of Graphene-Copper Nanoparticle Composite by in Situ Chemical Reduction for Electrochemical Sensing of Carbohydrates. Anal. Chem. 2012, 84, 171–178. [Google Scholar] [CrossRef]
- Xu, Y.; Chen, D.; Jiao, X. Fabrication of CuO Pricky Microspheres with Tunable Size by a Simple Solution Route. J. Phys. Chem. B 2005, 109, 13561–13566. [Google Scholar] [CrossRef]
- Bondarenko, O.; Juganson, K.; Ivask, A.; Kasemets, K.; Mortimer, M.; Kahru, A. Toxicity of Ag, CuO and ZnO Nanoparticles to Selected Environmentally Relevant Test Organisms and Mammalian Cells in Vitro: A Critical Review. Arch. Toxicol. 2013, 87, 1181–1200. [Google Scholar] [CrossRef] [Green Version]
- Goyal, P.; Chakraborty, S.; Misra, S.K. Multifunctional Fe3O4-ZnO Nanocomposites for Environmental Remediation Applications. Environ. Nanotechnol. Monit. Manag. 2018, 10, 28–35. [Google Scholar] [CrossRef]
- Dreyer, D.R.; Park, S.; Bielawski, C.W.; Ruoff, R.S. The Chemistry of Graphene Oxide. Chem. Soc. Rev. 2010, 39, 228–240. [Google Scholar] [CrossRef]
- Karthik, R.; Govindasamy, M.; Chen, S.-M.; Chen, T.-W.; Elangovan, A.; Muthuraj, V.; Yu, M.-C. A Facile Graphene Oxide Based Sensor for Electrochemical Detection of Prostate Anti-Cancer (Anti-Testosterone) Drug Flutamide in Biological Samples. Rsc. Adv. 2017, 7, 25702–25709. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, M.; Yukimoto, M. Effects of Fungicides on Channels in the Fungal Membrane. Pestic. Biochem. Physiol. 1993, 47, 171–177. [Google Scholar] [CrossRef]
- Yan, X.; Liang, X.; Wang, D. Primary Mode of Action of Novel Fungicide Chesulfamide against Botrytis cinerea. In Proceedings of the Abstracts of Papers of the American Chemical Society; American Chemical Society: Washington, DC, USA, 2014; Volume 248. [Google Scholar]
- Priester, J.H.; Ge, Y.; Mielke, R.E.; Horst, A.M.; Moritz, S.C.; Espinosa, K.; Gelb, J.; Walker, S.L.; Nisbet, R.M.; An, Y.-J. Soybean Susceptibility to Manufactured Nanomaterials with Evidence for Food Quality and Soil Fertility Interruption. Proc. Natl. Acad. Sci. USA 2012, 109, E2451–E2456. [Google Scholar] [CrossRef] [Green Version]
- Elmer, W.H.; White, J.C. The Use of Metallic Oxide Nanoparticles to Enhance Growth of Tomatoes and Eggplants in Disease Infested Soil or Soilless Medium. Environ. Sci. Nano 2016, 3, 1072–1079. [Google Scholar] [CrossRef]
- Singh, R.K.; Ruiz-May, E.; Rajput, V.D.; Minkina, T.; Gómez-Peraza, R.L.; Verma, K.K.; Shekhawat, M.S.; Pinto, C.; Falco, V.; Quiroz-Figueroa, F.R. Viewpoint of Chitosan Application in Grapevine for Abiotic Stress/Disease Management towards More Resilient Viticulture Practices. Agriculture 2022, 12, 1369. [Google Scholar] [CrossRef]
- Mamode Ally, N.; Neetoo, H.; Ranghoo-Sanmukhiya, V.M.; Hardowar, S.; Vally, V.; Gungoosingh-Bunwaree, A.; Maudarbaccus, F.; Coutinho, T.A.; Vojvodić, M.; Bulajić, A. First Report of Botrytis cinerea Causing Gray Mold on Greenhouse-Grown Tomato Plants in Mauritius. Plant Dis. 2021, 105, 2725. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ismail, A.M.; Mosa, M.A.; El-Ganainy, S.M. Chitosan-Decorated Copper Oxide Nanocomposite: Investigation of Its Antifungal Activity against Tomato Gray Mold Caused by Botrytis cinerea. Polymers 2023, 15, 1099. https://0-doi-org.brum.beds.ac.uk/10.3390/polym15051099
Ismail AM, Mosa MA, El-Ganainy SM. Chitosan-Decorated Copper Oxide Nanocomposite: Investigation of Its Antifungal Activity against Tomato Gray Mold Caused by Botrytis cinerea. Polymers. 2023; 15(5):1099. https://0-doi-org.brum.beds.ac.uk/10.3390/polym15051099
Chicago/Turabian StyleIsmail, Ahmed Mahmoud, Mohamed A. Mosa, and Sherif Mohamed El-Ganainy. 2023. "Chitosan-Decorated Copper Oxide Nanocomposite: Investigation of Its Antifungal Activity against Tomato Gray Mold Caused by Botrytis cinerea" Polymers 15, no. 5: 1099. https://0-doi-org.brum.beds.ac.uk/10.3390/polym15051099