1. Introduction
The introgression of novel genes for
Striga resistance from the wild relative of maize,
Zea diploperennis L., into the background of cultivated maize is a resourceful approach for genetic and physiological studies [
1,
2,
3]. Moreover, it has tremendous potential in breeding that could be utilized to develop novel cultivars and for broadening the genetic base of tropical maize breeding populations. However, a thorough understanding of the genetic diversity in the maize inbred lines and assessment of their reactions to
Striga are essential for systematic exploitation, to provide the capacity to meet changing environments and market requirements [
4]. Furthermore, the information on the genetic diversity and relatedness within a germplasm collection could be an invaluable aid in deciding the best breeding strategies to be employed [
5]. Additionally, the combination of pedigree information and genetic distance (GD) estimates could be useful for assigning inbred lines to distinct heterotic groups that could help to prevent crosses between closely related lines [
6].
Deoxyribonucleic acid (DNA) markers are invaluable in determining the level of genetic diversity present within genetic materials because these markers are not influenced by the different processes of plant physiology or the environmental conditions [
7]. In the past, genetic diversity in maize has been assessed using several types of DNA markers including restriction fragment length polymorphism (RFLP), randomly amplified polymorphism DNA (RAPD), simple sequence repeat (SSR) and single nucleotide polymorphism (SNP) markers. These markers have provided effective genotyping and are not affected by the different processes of plant physiology or the environment [
8,
9,
10,
11]. The high level of polymorphism together with cost effectiveness associated with SNP markers have resulted in this marker system being considered as the “markers of choice” in plant science including maize improvement [
12]. At the International Institute of Tropical Agriculture (IITA), maize breeders have also focused on the genetic diversity of tropical maize inbred lines at the molecular level [
13,
14]. However, such information is rather limited in the extra-early (80–85 days to physiological maturity) and early (90–95 days to physiological maturity) maturing inbred lines compared to the medium-late maturing (100–120 days to physiological maturity) maize inbreds in the IITA’s maize improvement program [
15].
Besides high grain yield, IITA’s early and extra-early maize program is focusing on breeding for combined resistance to
Striga and drought stress. Although direct selection for grain yield alone is not reliable due to the complex genetic nature of the trait, low heritability of grain yield under stress and the significant genotype by environment interaction (GEI), particularly under different stress conditions, make selection a daunting task for breeders. Thus, plant breeders do not only continuously try to identify reliable secondary traits that have significant effects on economic yield, but also make efforts to develop multiple trait base indices for selection for improved stress tolerance [
3,
15,
16]. Recently, we have also developed multiple trait base indices integrating grain yield and other important secondary traits in early and extra-early maize groups under different abiotic stresses [
17], but their inclusion in the breeding program of the extra- early maize needs to be further verified.
The (IITA) maize program has used resistance genes from diverse germplasm sources, including temperate and tropical materials identified following several years of extensive testing in the savanna of West and Central Africa (WCA). However, the
Striga resistance genes have not been as effective as desirable in the control of
Striga, because they allow the flowering and seed production of
Striga plants, thereby increasing the
Striga seed bank in the soil. Consequently, there has been a search for novel genes for resistance to
Striga hermonthica in the wild relative of maize,
Zea diploperennis [
3,
18]. Several early maturing inbred lines have been developed containing the novel
Striga resistance genes from
Z. diploperennis in IITA. However, the combining ability of the inbreds have not been determined. Development of hybrid varieties, their promotion and adoption are promising strategies for an appreciable increase in maize production and to transform agriculture in WCA. Several seed companies have sprung up in the sub-region during the past two decades, setting the stage for commercial hybrid seed production. However, there are no commercial early maturing hybrids with high levels of
Striga resistance available to these companies to produce for the farmers in the savanna, which is a
Striga hotspot [
15].
Information on the general and specific combining abilities of inbred lines is crucial in identifying productive hybrids for commercial hybrid production without making all possible crosses among the parental inbred lines. A major challenge presently confronting the IITA maize improvement program is to test the numerous inbred lines, developed from several source germplasms in hybrid combinations, to identify and promote commercialization of productive hybrids with high levels of Striga resistance genes in WCA. However, limited information is available on the combining ability, repeatability, heritability and performance of the early maturing white maize inbreds in hybrid combinations under Striga infestation and non-infested conditions. In the present study, we characterize a set of early maturing maize inbred lines selected for resistance to Striga from different source populations, including those containing genes for Striga resistance from the wild maize, Zea diploperennis, and assess genetic diversity at the molecular level using SNP markers. Additionally, we determine the combining abilities of the maize inbreds under Striga-infested environments and identify the most productive and stable hybrids under Striga-infested and non-infested conditions. Lastly, we examine the performance of the inbreds in hybrid combinations, heritability and repeatability, and investigate the associations among measured traits under Striga infestation and non-infested conditions using sequential path analysis.
4. Discussion
The genotypic mean squares of the inbred lines and derived hybrids were highly significant for all the measured traits under
Striga infestation, indicating that substantial genetic variation existed among the inbred lines, which should facilitate selection for
Striga resistance and increased grain yield and other measured traits under the research conditions. Moreover, the significant environment mean squares observed under
Striga infestation indicated that the test locations were unique in discriminating among the inbred lines, and that testing of the inbred lines in a wide array of locations over years will be required to identify the most stable lines for hybrid production [
21]. The significant genotype by environment interaction (GEI) mean squares for
Striga damage at eight and 10 WAP and number of emerged
Striga plants at 10 WAP indicated that the inbred lines varied in their responses to infestation at the different locations, and that such variations in genotypic response could be due to the presence of different biotypes of
S. hermonthica at the experimental locations. This provided a justification for evaluating the hybrids across the two distinct environments to identify those with consistent performance across the environments. This finding corroborates the results of [
15,
21]. The average yield reduction of the hybrids under
Striga infestation was 44% relative to performance under non-infested conditions. This is consistent with the results of [
42,
43] who reported yield reduction of 42 and 44%, respectively, under
Striga infestation. However, it is lower than the 53.7% reported by [
44], 68% by [
45], 65% by [
46] and 55% by [
15]. It is; however, higher than the yield reduction of 23% reported by [
25]. The yield reduction of 44% suggested that the intensity of infestation in this study was high enough to allow the identification of hybrids that possessed genes for
Striga resistance/tolerance.
Despite the high severity of infestation in this study, the novel
Striga resistance genes from the wild relatives of maize,
Z. diploperennis, in the genotypes allowed them to suppress the emergence of the
Striga plants and produced high yields. In
Striga research, resistance to
Striga refers to the capability of the host plant to induce the germination of
Striga seeds but prevent the parasite from attaching to the roots of the maize plants or kills the attached parasitic plants. Under
Striga infestation, the resistant genotype supports significantly fewer
Striga plants and produces a greater yield than the susceptible genotype. Contrarily, a
Striga tolerant genotype supports as many
Striga plants as the sensitive or susceptible genotype but produces more dry matter and shows fewer damage symptoms.
S. hermonthica damage in maize is used as the indicator of tolerance, while emerged
Striga plants is the indicator of resistance. Identification of maize genotypes that combine outstanding levels of resistance and tolerance is a promising breeding strategy and has been recommended for
Striga resistance breeding in several studies [
47,
48,
49]. The increased in grain yield of the hybrids under
Striga infestation was accompanied by reduced number of emerged
Striga plants as well as reduced
Striga damage. The moderately high heritability (> 60%) obtained for days to 50% silking,
Striga damage, number of emerged
Striga plants and ear aspect under
Striga infestation; grain yield, days to silking, plant and ear aspects under non-infested conditions indicated that the traits could be easily transmitted from the parental lines to their offspring. The significant GCA-male, GCA-female and SCA for grain yield and other agronomic traits except for ASI under
Striga infestation suggested that the performance of the inbreds differed when used as either male or female parents in hybrid combinations. The preponderance of GCA variances over SCA for grain yield, and most measured traits under
Striga infestation and non-infested conditions, implied that additive gene action largely controlled the inheritance of these traits. The implication is that selected inbred lines could be intercrossed to form heterotic populations, which could be improved through recurrent selection methods, such as the S
1 family and the full-sib family selection schemes. Inbred lines tolerant to
Striga infestation with high GCA effects could then be extracted from improved cycles of selection of derived populations for hybrid development. The high GCA over SCA mean squares for
Striga damage and number of emerged
Striga plants under
Striga infestation indicated that additive gene action was more important in controlling both host plant damage and number of emerged
Striga plants. The results of this study are, in part, contradictory to the findings of [
47,
48], who showed that non-additive gene action was more important than additive gene action in controlling the inheritance of host plant damage, while additive gene action was more important in governing the inheritance of the number of emerged
Striga plants under
Striga infestation. Furthermore, the findings of this study are contradictory to the results of [
49,
50,
51], who reported that additive gene action controlled
Striga damage, while non-additive gene action modulated the number of emerged
Striga plants. The discrepancy in the results of the present study and those of the earlier researchers may be attributed to the differences in germplasm sources or the severity of the
Striga infestation.
In this study, the percentage contributions of GCA-male and GCA-female effects did not significantly vary for grain yield and other traits under
Striga-infested and non-infested conditions, implying that maternal or cytoplasmic genes did not have any influence on the measured traits. This finding is contrary to the results of [
43], who reported maternal effects for days to silking and paternal effects for EPP in early maturing maize hybrids under
Striga-infested environments.
Inbred lines TZdEI 268, TZdEI 352 and TZdEI 173 had positive and significant GCA (GCA-male and/or GCA-female) effects for grain yield under Striga infestation, indicating that the inbred lines may have contributed to higher grain yield in their hybrids under Striga infestation. Contrarily, TZdEI 260, TZdEI 396, TZdEI 479 and TZdEI 173 had positive and significant GCA effects for grain yield under non-infested conditions and are expected to contribute higher grain yield to their hybrids. The inbred lines with positive and significant GCA effects for grain yields could be used as parents to form a synthetic population that could be improved for Striga resistance based on the heterotic orientations of the inbred lines. Subsequently, new inbred lines with improved levels of Striga resistance could be extracted from the improved populations.
The selection index for Striga resistance/tolerance which integrated grain yield, ears per plant, Striga damage and number of emerged Striga plants revealed that only 50% of the inbred lines had positive base indices and; therefore, had good levels of resistance/tolerance to Striga. Using the base indices, TZdEI 173 × TZdEI 352 showed good performance under Striga infestation This result is interesting and encouraging, as Striga is a menace in the savannas of WCA, which is the maize belt of the region, and farmers in the sub-region are abandoning their heavily infested fields due to S. hermonthica. The superior yielding hybrids identified in the present study should be evaluated extensively in contrasting environments in multi-location trials to confirm the superior performance and commercialized in sub-Saharan Africa (SSA).
The average mid- and high-parent heterosis for grain yield observed under Striga infestation indicated that the hybrids produced more grain yield than their inbred parents. Negative mid- and high-parent heterosis values obtained for days to 50% anthesis and silking indicated that the hybrids flowered earlier than their corresponding inbred parents under Striga-infested environments. Additionally, the positive mid- and high-parent heterosis observed for Striga damage and number of emerged Striga plants at eight and 10 WAP indicated that the hybrids suffered severe Striga damage and allowed the emergence of more Striga plants than their susceptible parental lines.
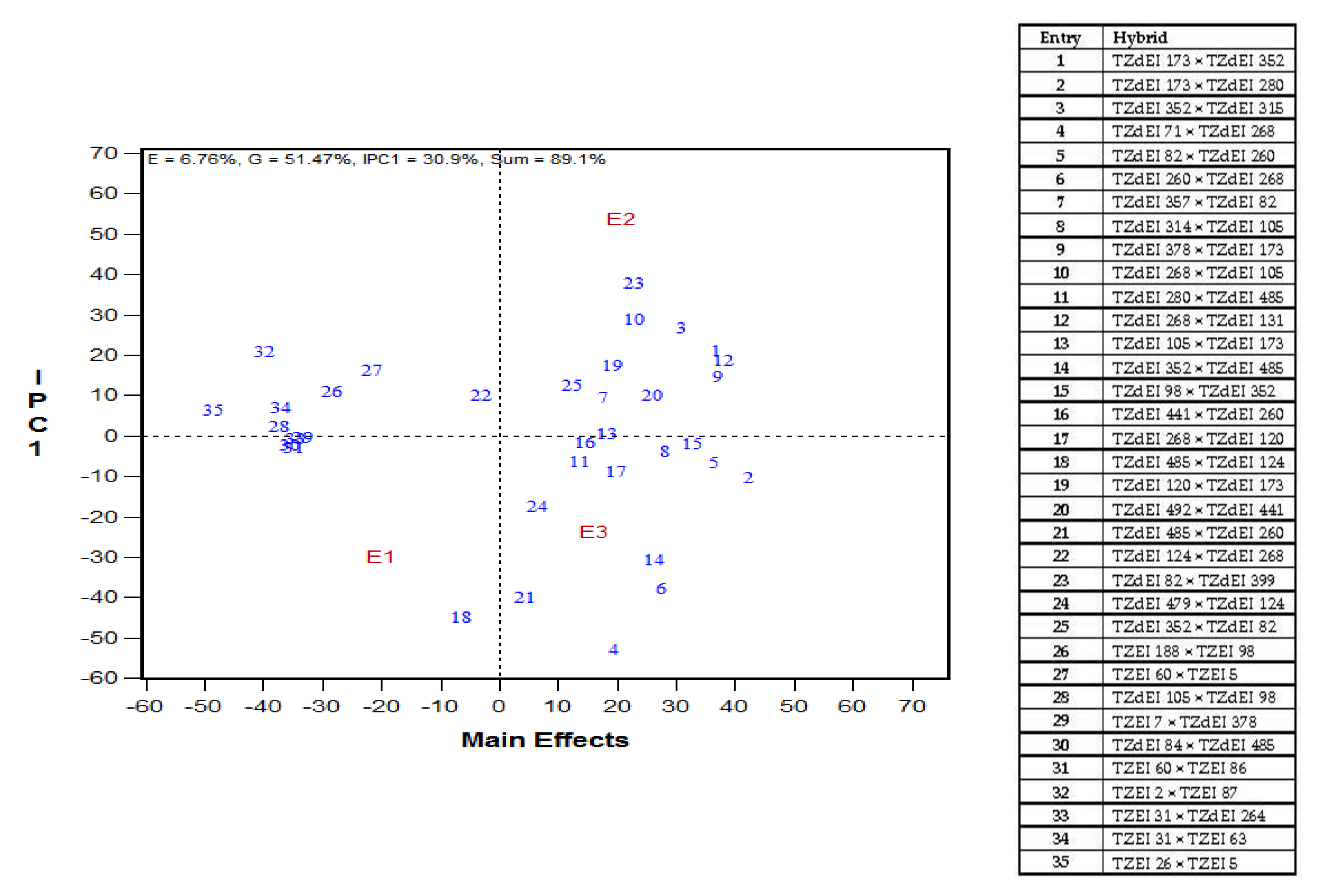
It is striking to note that five hybrids (TZdEI 173 × TZdEI 280, TZdEI 82 × TZdEI 260, TZdEI 98 × TZdEI 352, TZdEI 441 × TZdEI 260, TZdEI 492 × TZdEI 441) were identified by AMMI biplots as high yielding and stable under Striga infestation. These hybrids should be extensively tested in on-farm trials in SSA to confirm the consistency in performance, and vigorously promoted for commercialization, to contribute to food security and improved livelihoods of resource poor farmers in the sub-region.
Although maize is native of the tropical region, its global germplasm naturally forms two major groups—temperate and tropical (including subtropical)—which is further sub-grouped based on either trait or region in the world. In maize, several researchers have investigated marker-based diversity focusing on specific germplasm with limited sample sizes, including tropical and subtropical lines [
52,
53]. These studies have shown that there is much more diversity in tropical lines, giving huge opportunities for continuous long-term genetic gain. The average polymorphic information content (PIC) value of 0.10, in the present set of maize inbreds revealed by SNP markers, was lower than what was reported in previous studies of maize [
53,
54,
55]. The differences in the results of the various authors may be attributed to the composition of germplasm, population size and type and number of molecular markers examined.
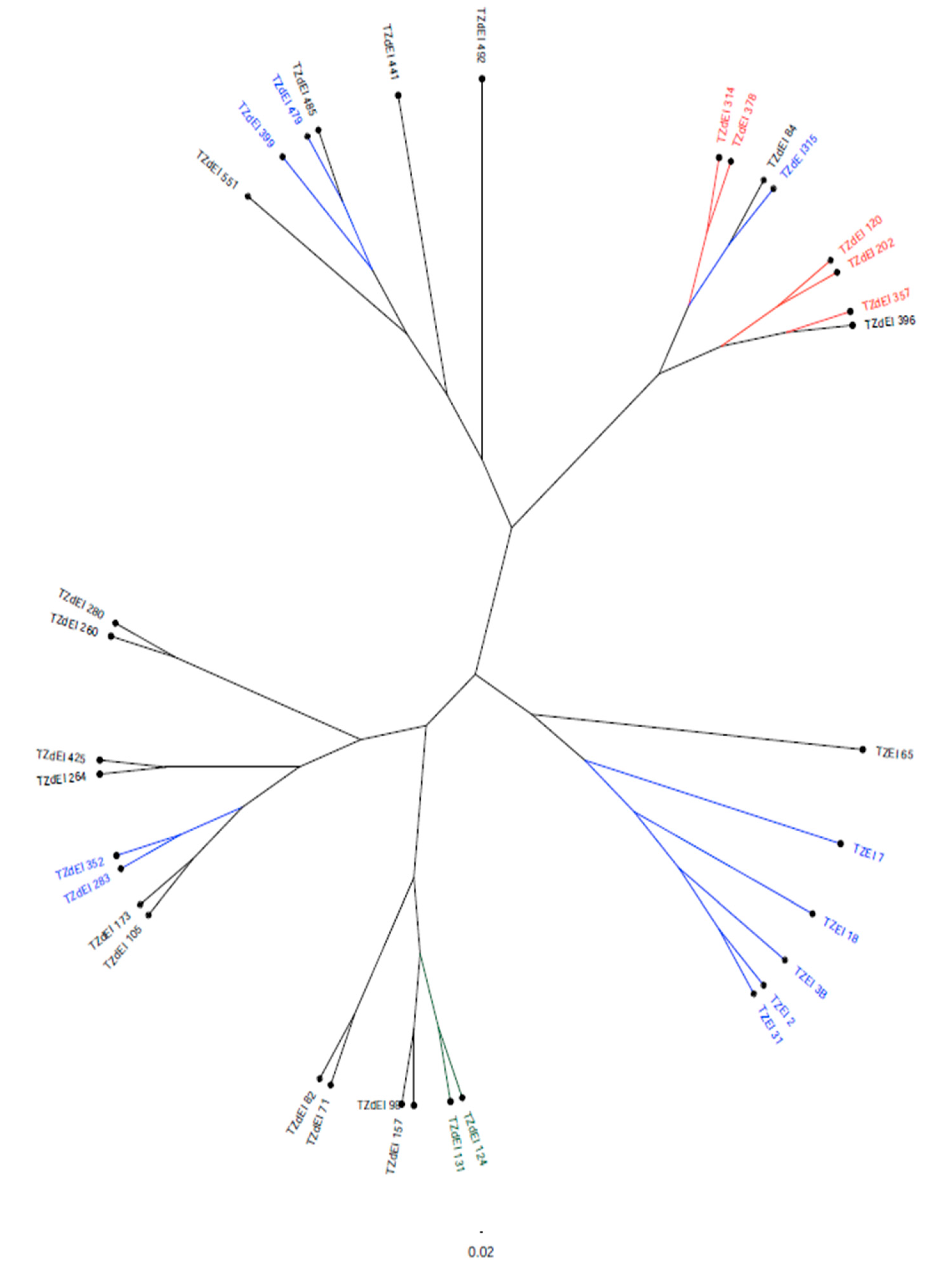
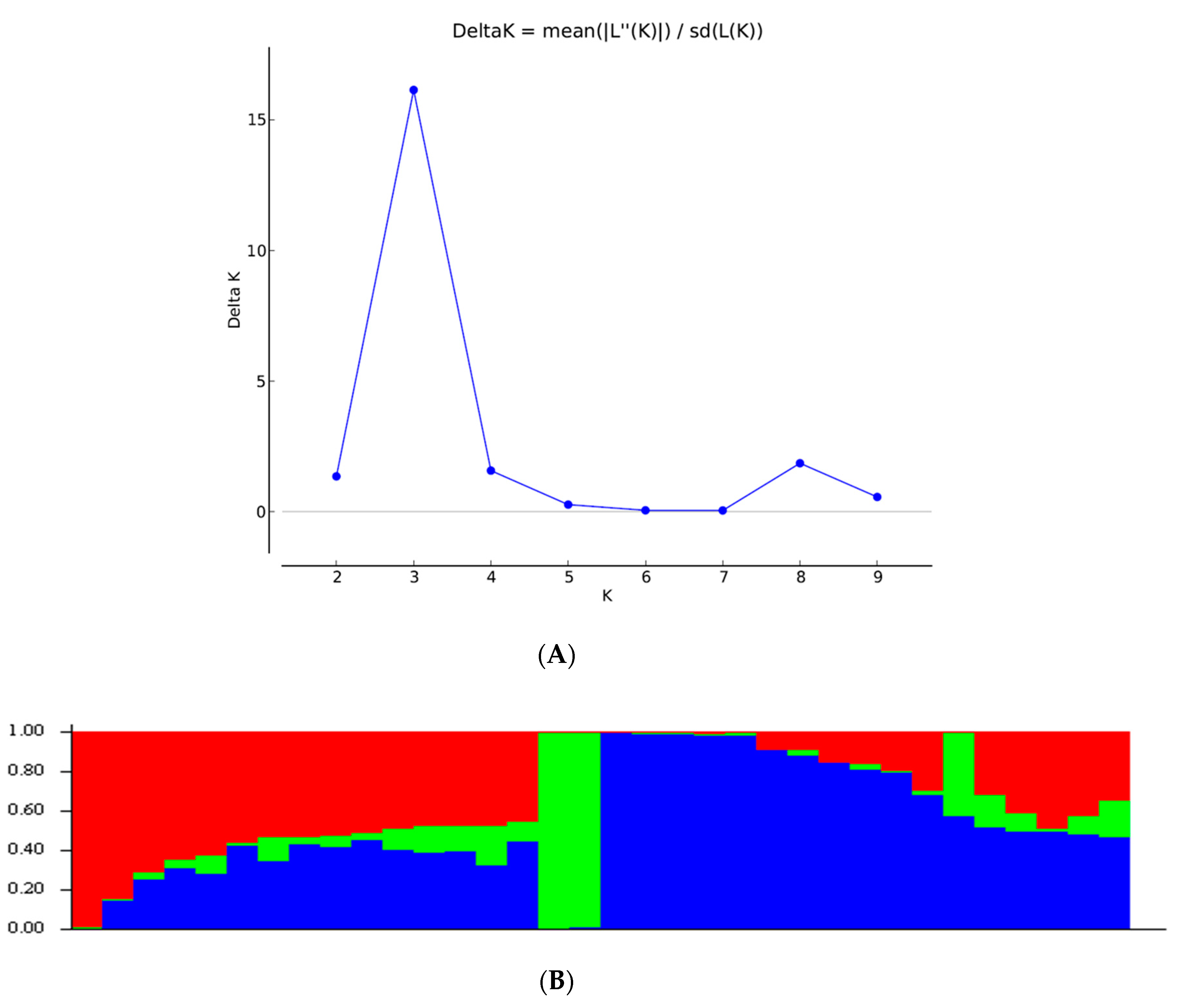
The SNP markers clearly grouped all the inbred lines into three clusters. The six TZEI lines without introgression from
Z. diploperennis clustered closely together (C-I), whereas the remaining inbred lines having genes introgressed from the wild relative of maize,
Z. diploperennis (designated as TZdEI), were grouped into two different clusters (C-II and C-III), each corresponding to two sub-clusters. Similarly, the STRUCTURE analysis differentiated the population into three sub-population groups. However, a large proportion of the inbred lines (50%) appeared as admixture at a probability >70%, as these inbred lines did not fit in any of the sub-population groups. Such large admixture is expected since these inbred lines were selected under
Striga-infested conditions that may possibly favor the retention of higher proportion of introgression from the wild relatives of maize,
Z. diploperennis L, for
Striga resistance into the background of these inbred lines. For example, all the members of cluster I (C-I) also belonged to sub-population III (blue colour), except TZEI 65, which was considered as admixture genotype by STRUCTURE analysis. The second cluster (C-II) was subdivided in two sub-clusters (C-IIa and C-IIb), but the sub-cluster C-IIb, which contained most of lines susceptible to
Striga stress, also contained all the five inbreds belonging to sub-population I (red colour) based on STRUCTURE analysis. The third cluster (C-III) contained 14 individuals, the majority being in the admixture sub-population, but the two inbred lines forming sub-population II (green color) via STRUCTURE analysis were clustered together in sub-cluster C-IIIa, representing resistance to
Striga infestation. This is consistent with the results of the previous study of [
53] with CIMMYT maize inbred lines, which suggested that it was much more difficult to find a clear clustering based on the traits for which lines were bred. Because of their similar origin, they are not genetically distinct.
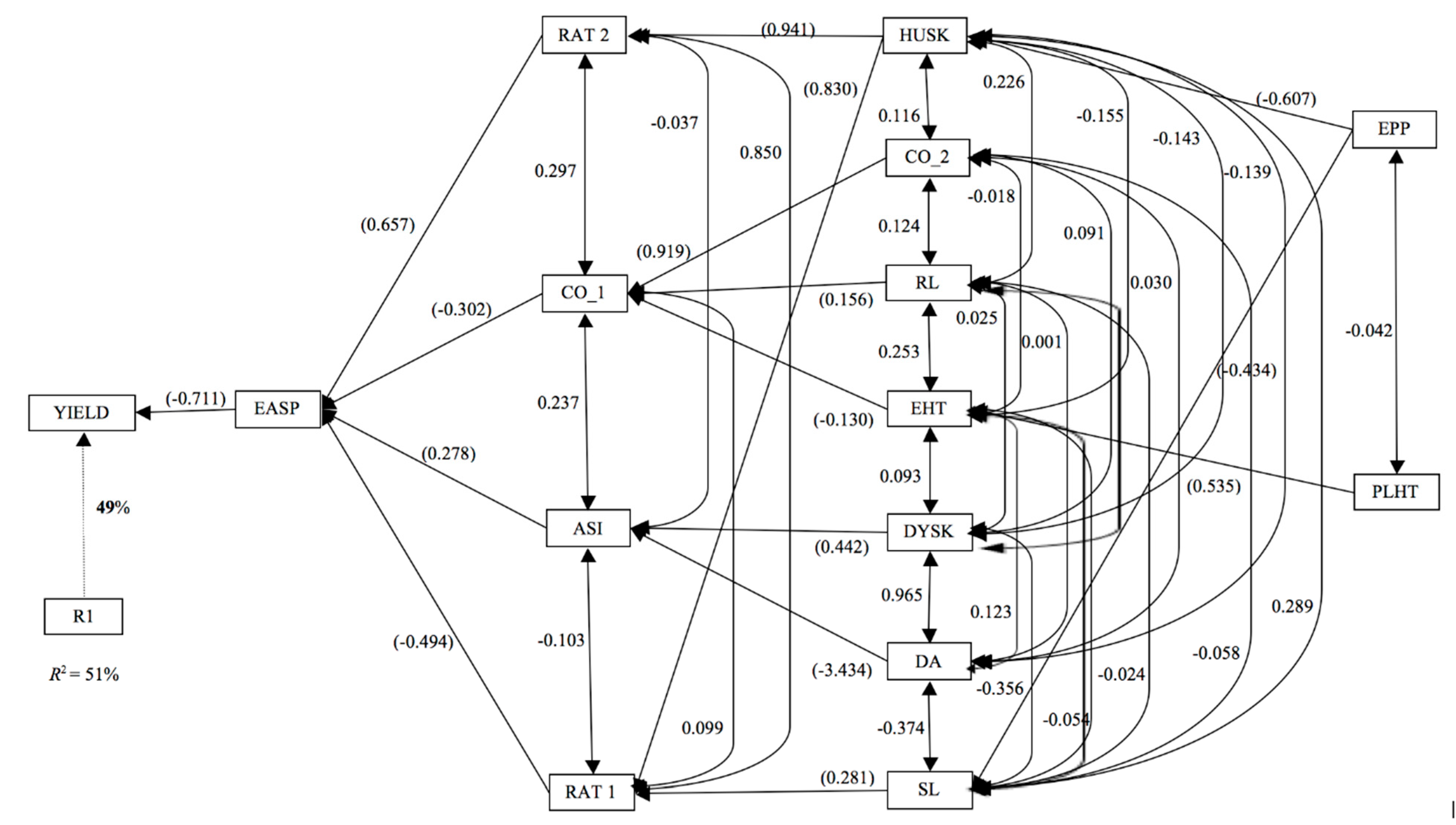
The ear aspect identified as the most important trait contributing to the variation in grain yield under
Striga infestation in the present study is concordant with previous studies suggesting the ear aspect as most reliable secondary trait for selecting
Striga tolerant/resistant maize inbred lines [
31,
33]. For instance,
Striga damage at eight and 10 WAP, number of emerged
Striga plants at eight WAP and ASI were identified as the second-order traits contributing to variation in grain yield under
Striga infestation. Obviously, reliable secondary traits in selecting outstanding
Striga-resistant/tolerant genotypes may vary depending on the nature of the genetic material used, prevailing climatic conditions and location of the experiment [
31,
33]. Finally, the results of the cluster analysis were consistent with the genetic background of the inbred lines and revealed valuable information that might be useful in resolving the heterotic groups of early-maturing inbred lines bred at IITA that are yet to be field-tested in hybrid combinations.


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}