Tef (Eragrostis tef Trotter) Responses to Nitrogen Fertigation under Semi-Arid Mediterranean Climate



Abstract
:1. Introduction
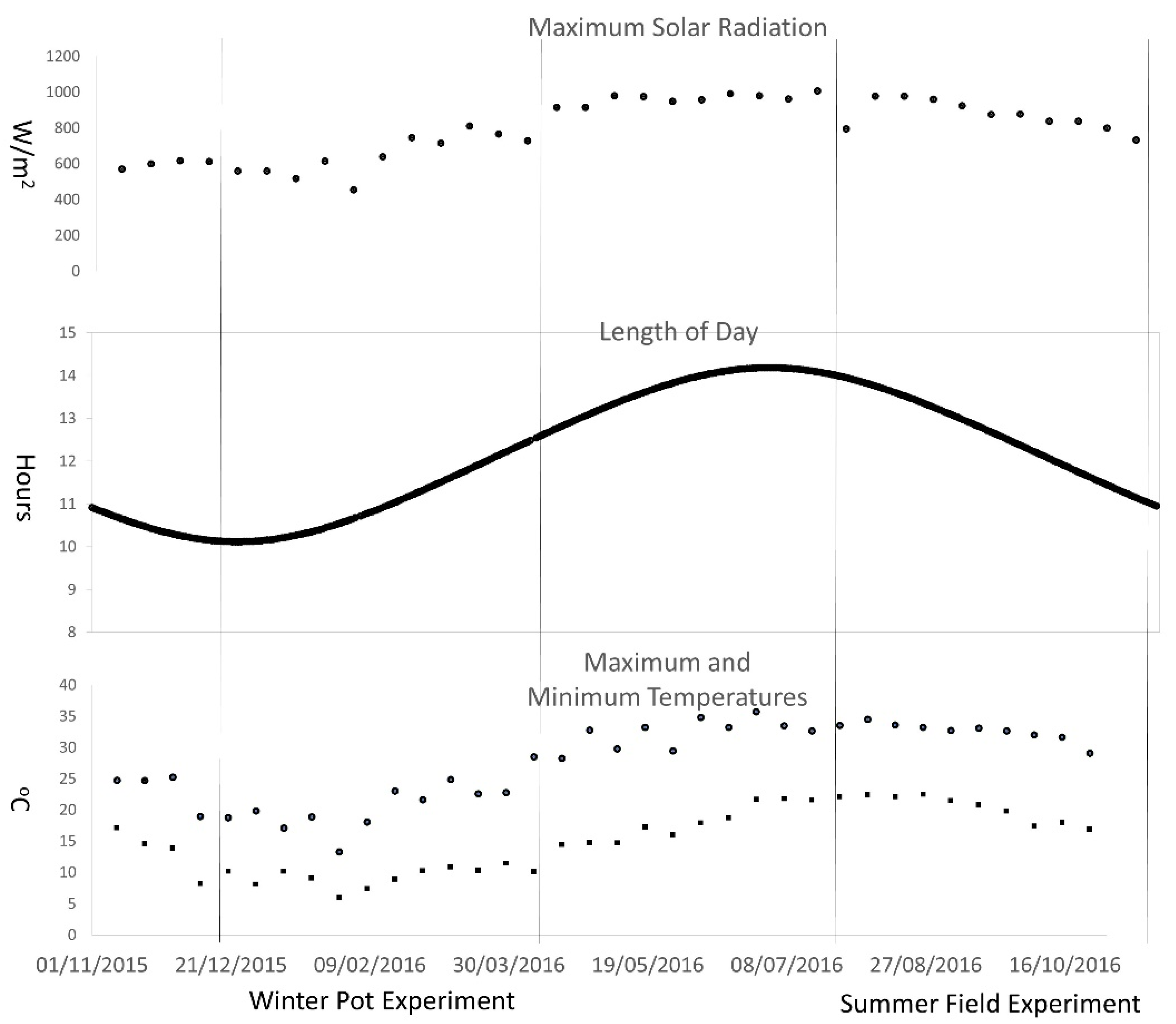
2. Materials and Methods
2.1. Plant Materials and Experimental Design
2.2. Data Collection
2.3. Statistical Analyses
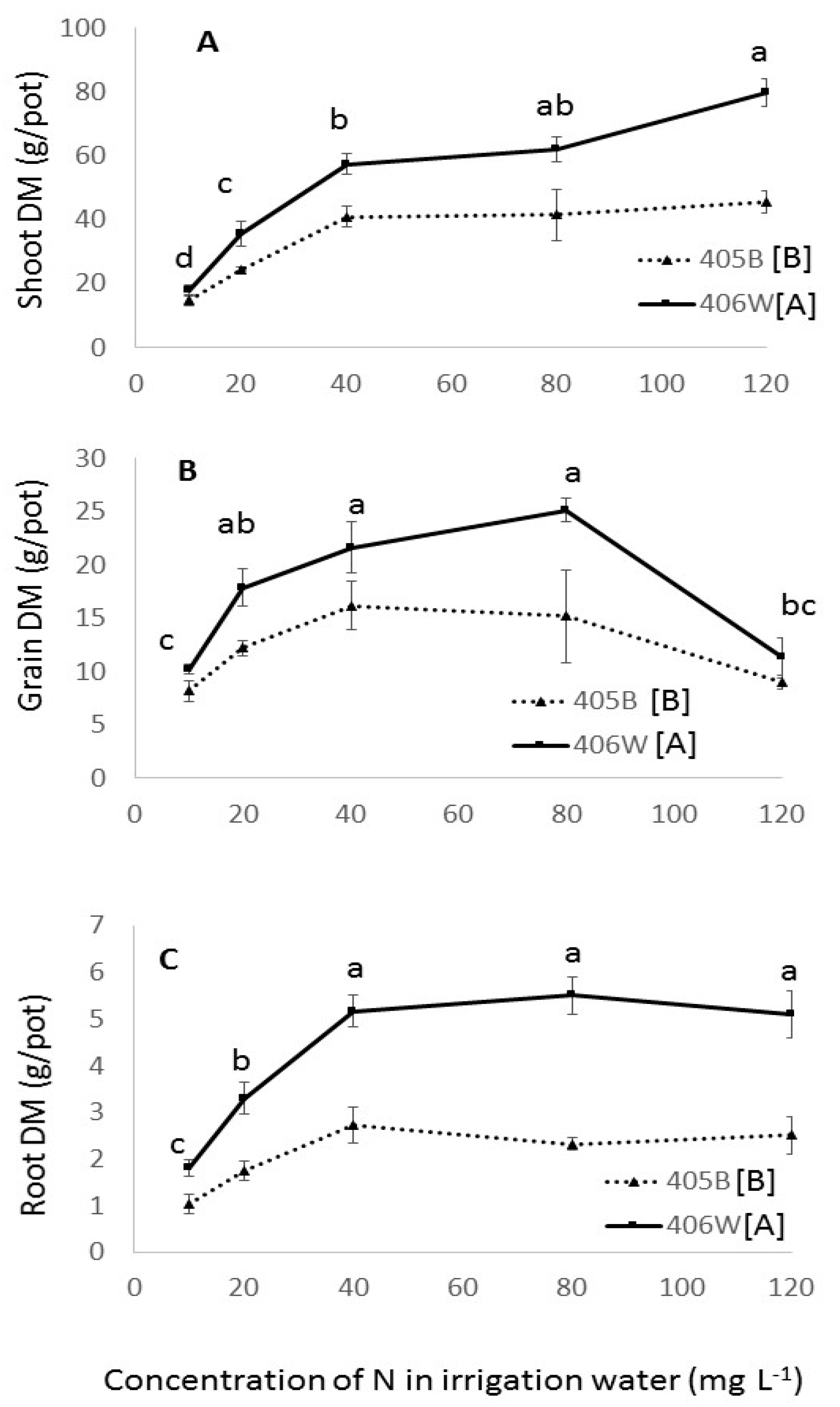
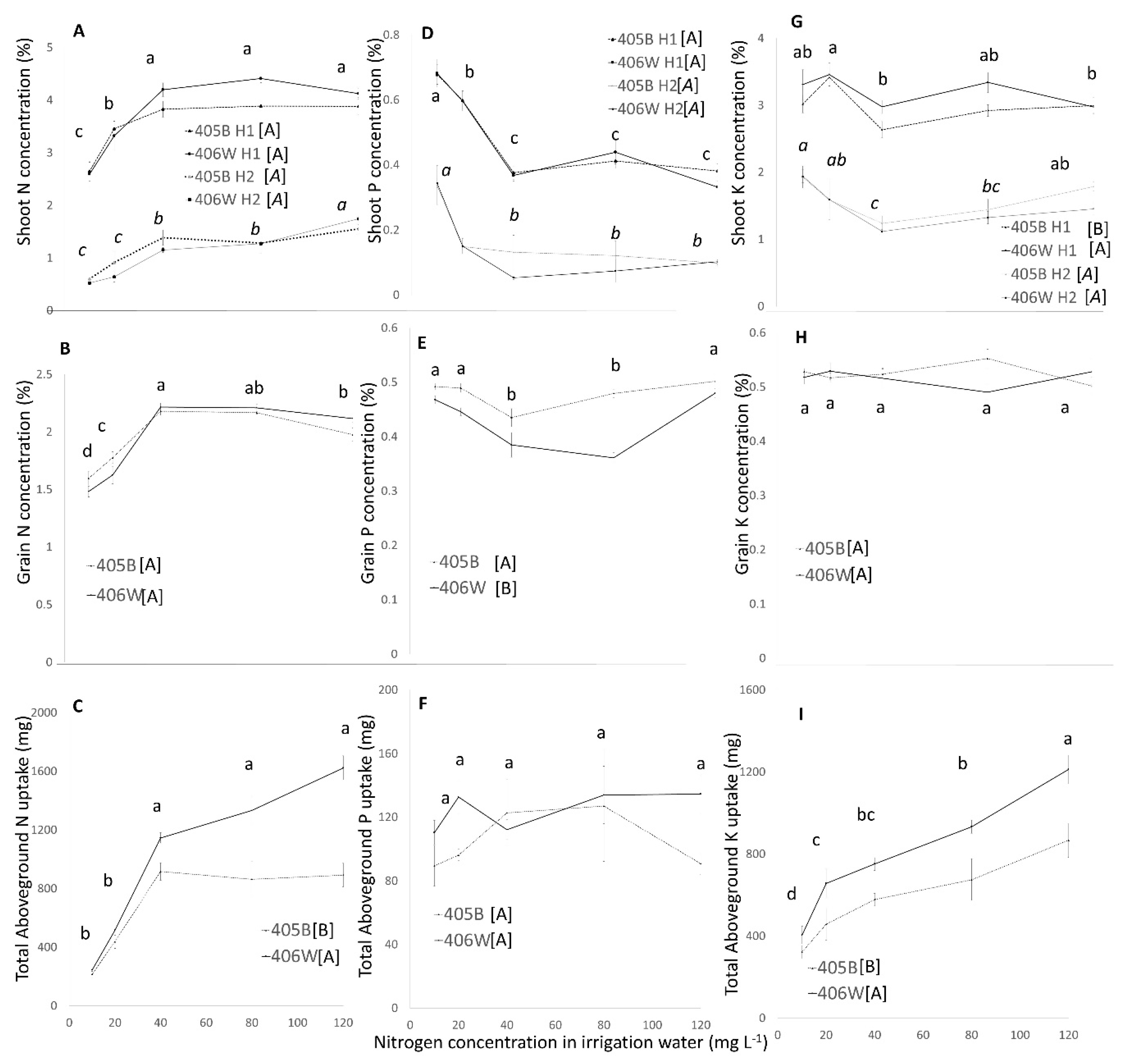
3. Results
3.1. Pot Experiment
3.2. Field Experiment
4. Discussion
4.1. Pot Experiment
4.2. Field Experiment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Roussis, I.; Folina, A.; Kakabouki, I.; Kouneli, V.; Karidogianni, S.; Chroni, M.; Bilalis, D. Effect of organic and inorganic fertilization on yield and yield components of teff [Eragrostis tef (Zucc.) Trotter] cultivated under Mediterranean semi-arid conditions. Sci. Pap. Ser. A Agron. 2019, 62, 138–144. [Google Scholar]
- El-Alfy, T.S.; Ezzat, S.M.; Sleem, A.A. Chemical and biological study of the seeds of Eragrostis tef (Zucc.) Trotter. Nat. Prod. Res. 2012, 26, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F. Chemical composition and food uses of teff (Eragrostis tef). Food Chem. 2018, 239, 402–415. [Google Scholar] [CrossRef] [PubMed]
- Tatham, A.S.; Fido, R.J.; Moore, C.M.; Kasarda, D.D.; Kuzmicky, D.D.; Keen, J.N.; Shewry, P.R. Characterisation of the major prolamins of tef (Eragrostis tef) and finger millet (Eleusine coracana). J. Cereal Sci. 1996, 24, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Spaenij-Dekking, L.; Kooy-Winkelaar, Y.; Koning, F. The Ethiopian cereal tef in celiac disease. N. Engl. J. Med. 2005, 353, 1748–1749. [Google Scholar] [CrossRef] [PubMed]
- Mengesha, M.H. Chemical composition of teff (Eragrostis tef) compared with that of wheat, barley and grain sorghum. Econ. Bot. 1966, 20, 268–273. [Google Scholar] [CrossRef]
- Gebremariam, M.M.; Zarnkow, M.; Becker, T. Teff (Eragrostis tef) as a raw material for malting, brewing and manufacturing of gluten-free foods and beverages: A review. Food Sci. Technol. 2012, 51, 2881–2895. [Google Scholar] [CrossRef] [Green Version]
- Tietel, Z.; Simhon, E.; Gashu, K.; Ananth, D.A.; Schwartz, B.; Saranga, Y.; Yermiyahu, U. Nitrogen availability and genotype affect major nutritional quality parameters of tef grain grown under irrigation. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Ketema, S. Eragrostis tef (Zucc.) Trotter. Promoting the Conservation and Use of Underutilized and Neglected Crops; Bioversity International Publication: Rome, Italy, 1997; Volume 12, pp. 1–52. [Google Scholar]
- Gelaw, A.M.; Qureshi, A.S. Tef (Eragrostis tef): A Superfood Grain from Ethiopia with Great Potential as an Alternative Crop for Marginal Environments. In Emerging Research in Alternative Crops; Springer: Cham, Switzerland, 2020; pp. 265–278. [Google Scholar]
- Efrem, B. Performance of some cereal crops under drained Vertisols in the Ethiopian highlands. In Advances in Vertisols management in Ethiopian Highlands; Paulos, D., Asgelil, D., Asfaw, Z., Gezahegn, A., Abebe, K., Eds.; Ethiopian Institute of Agricultural Research: Addis Ababa, Ethiopia, 2001; pp. 151–157. [Google Scholar]
- Chanyalew, S.; Ferede, S.; Damte, T.; Fikre, T.; Genet, Y.; Kebede, W.; Tolossa, K.; Zehirun, T.; Assefa, K. Significance and prospects of an orphan crop tef. Planta 2019, 250, 753–767. [Google Scholar] [CrossRef] [Green Version]
- Lee, H. Teff, a rising global crop: Current status of teff production and value chain. Open Agric. J. 2018, 12, 185–193. [Google Scholar] [CrossRef]
- Teklu, Y.; Tefera, H. Genetic improvement in grain yield potential and associated agronomic traits of tef (Eragrostis tef). Euphytica 2005, 141, 247–254. [Google Scholar] [CrossRef]
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Møller, I.S.; White, P. Functions of macronutrients. In Marschner’s Mineral Nutrition of Higher Plants; Academic Press: Cambridge, CA, USA, 2012; pp. 135–189. [Google Scholar]
- IFDC. Assessment of Fertilizer Consumption and Use by Crop in Ethiopia. 2015. Available online: https://africafertilizer.org/wp-content/uploads/2017/05/FUBC-Ethiopia-Final-Report-2016.pdf (accessed on 23 September 2020).
- Gebretsadik, H.; Haile, M.; Yamoah, C.F. Tillage frequency, soil compaction and N-fertilizer rate effects on yield of teff (Eragrostis tef (Zucc) Trotter) in central zone of Tigray, Northern Ethiopia. Momona Ethiop. J. Sci. 2009, 1. [Google Scholar] [CrossRef] [Green Version]
- Assefa, K.; Erenso, T.F.; Banja, T.H. Phosphorus critical level and optimum nitrogen rate determination on teff for sustainable soil fertility management and economical teff production at Lume Area of Oromia region, Ethiopia. J. Agric. Biol. Healthc. 2017, 7, 2224–3208. [Google Scholar]
- Wato, T. Effects of Nitrogen Fertilizer Rate and Inter-row Spacing on Yield and Yield Components of Teff [Eragrostis teff (Zucc.) Trotter] in Limo District, Southern Ethiopia. Int. J. Plant Soil Sci. 2019, 31, 1–12. [Google Scholar] [CrossRef]
- Girma, K.; Reinert, M.; Ali, M.S.; Sutradhar, A.; Mosali, J. Nitrogen and phosphorus requirements of teff grown under dryland production system. Crop. Manag. 2012, 11, 1–14. [Google Scholar] [CrossRef]
- O’Donovan, J.T.; Turkington, T.K.; Edney, M.J.; Clayton, G.W.; McKenzie, R.H.; Juskiw, P.E.; Lafond, G.P.; Grant, C.A.; Brandt, S.; Harker, K.N.; et al. Seeding rate, nitrogen rate, and cultivar effects on malting barley production. Agron. J. 2011, 103, 709–716. [Google Scholar]
- Zhang, M.; Wang, H.; Yi, Y.; Ding, J.; Zhu, M.; Li, C.; Guo, W.; Feng, C.; Zhu, X. Effect of nitrogen levels and nitrogen ratios on lodging resistance and yield potential of winter wheat (Triticum aestivum L.). PLoS ONE 2017, 12, e0187543. [Google Scholar] [CrossRef]
- Pan, J.; Zhao, J.; Liu, Y.; Huang, N.; Tian, K.; Shah, F.; Liang, K.; Zhang, X.; Liu, B. Optimized nitrogen management enhances lodging resistance of rice and its morpho-anatomical, mechanical, and molecular mechanisms. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Kanter, D.R. Nitrogen pollution: A key building block for addressing climate change. Clim. Chang. 2018, 147, 11–21. [Google Scholar] [CrossRef]
- Kidanu, S.; Tanner, D.G.; Mamo, T. Effect of nitrogen fertiliser applied to tef on the yield and N response of succeeding tef and durum wheat on a highland vertisol. Afr. Crop. Sci. J. 1999, 7. [Google Scholar] [CrossRef] [Green Version]
- Habtegebrial, K.; Singh, B.R. Effects of timing of nitrogen and sulphur fertilizers on yield, nitrogen, and sulphur contents of Tef (Eragrostis tef (Zucc.) Trotter). Nutr. Cycl. Agroecosyst. 2006, 75, 213–222. [Google Scholar] [CrossRef]
- Dereje, G.; Alemu, D.; Adisu, T.; Anbessa, B. Response of yield and yield components of Tef [Eragrostis tef (Zucc.) Trotter] to optimum rates of nitrogen and phosphorus fertilizer rate application in Assosa Zone, Benishangul Gumuz Region. Ethiop. J. Agric. Sci. 2018, 28, 81–94. [Google Scholar]
- Szmidt, R.A.K.; Hall, D.A.; Hitchon, G.M. Development of perlite culture systems for the production of greenhouse tomatoes. In Symposium on Horticultural Substrates and Their Analysis; Acta Horticulturae: Brussels, Belgium, 1987; Volume 221, pp. 371–378. [Google Scholar]
- Zhang, Y.; Wang, J.; Gong, S.; Xu, D.; Sui, J. Nitrogen fertigation effect on photosynthesis, grain yield and water use efficiency of winter wheat. Agric. Water Manag. 2017, 179, 277–287. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulos, I. Nitrogen fertigation of trickle-irrigated potato. Nutr. Cycl. Agroecosyst. 1988, 16, 157–167. [Google Scholar] [CrossRef]
- Vanitha, K.; Mohandass, S. Ass Drip fertigation could improve source-sink relationship of aerobic rice (Oryza sativa L.). Afr. J. Agric. Res. 2014, 9, 294–301. [Google Scholar]
- Demirbas, A.; Akpinar, C.; Coskan, A.; Karakoy, T. Effects of different fertigation levels on maize yield and nutrient uptake under semi-arid Mediterranean conditions. Infrastruktura i Ekologia Terenów Wiejskich 2017, 727–736. [Google Scholar] [CrossRef]
- Halpern, M.; Bar-Tal, A.; Lugassi, N.; Egbaria, A.; Granot, D.; Yermiyahu, U. The role of nitrogen in photosynthetic acclimation to elevated [CO2] in tomatoes. Plant Soil 2018, 434, 397–411. [Google Scholar] [CrossRef]
- García Lozano, M.; Escobar, I.; Berenguer, J.J. Green-pepper fertigation in soilless culture. In International Symposium on Soilless Culture and Hydroponics; Acta Horticulturae: Brussels, Belgium, 2004; Volume 697, pp. 543–547. [Google Scholar]
- Silber, A.; Bruner, M.; Kenig, E.; Reshef, G.; Zohar, H.; Posalski, I.; Yehezkel, H.; Shmuel, D.; Cohen, S.; Dinar, M.; et al. High fertigation frequency and phosphorus level: Effects on summer-grown bell pepper growth and blossom-end rot incidence. Plant Soil 2005, 270, 135–146. [Google Scholar] [CrossRef]
- Omondi, J.O.; Lazarovitch, N.; Rachmilevitch, S.; Boahen, S.; Ntawuruhunga, P.; Sokolowski, E.; Yermiyahu, U. Nutrient use efficiency and harvest index of cassava decline as fertigation solution concentration increases. J. Plant Nutr. Soil Sci. 2018, 181, 644–654. [Google Scholar] [CrossRef]
- Ben-Zeev, S.; Bimro, J.; Barak, V.; Saranga, Y. Phenotypic diversity and heritability in Eragrostis tef under irrigated Mediterranean conditions. Isr. J. Plant Sci. 2018, 65, 222–231. [Google Scholar] [CrossRef]
- Yihun, Y.M.; Haile, A.M.; Schultz, B.; Erkossa, T. Crop Water Productivity of Irrigated Teff in a Water Stressed Region. Water Resour. Manag. 2013, 27, 3115–3125. [Google Scholar] [CrossRef]
- Assefa, K.; Cannarozzi, G.; Girma, D.; Kamies, R.; Chanyalew, S.; Plaza-Wüthrich, S.; Biosch, R.; Rindisbacher, A.; Rafudeen, S.; Tadele, Z. Genetic diversity in tef [Eragrostis tef (Zucc.) Trotter]. Front. Plant Sci. 2015, 6, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedane, G.M.; Saukuru, A.M.; George, D.L.; Gupta, M.L. Evaluation of teff (Eragrostis tef [Zucc.] Trotter) lines for agronomic traits in Australia. Aust. J. Crop Sci. 2015, 9, 242. [Google Scholar]
- Mariotti, F.; Tomé, D.; Mirand, P.P. Converting nitrogen into protein—Beyond 6.25 and Jones’ factors. Crit. Rev. Food Sci. Nutr. 2008, 48, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Giday, O.; Gibrekidan, H.; Berhe, T. Response of Teff (Eragrostis tef) to Different Rates of Slow Release and Conventional Urea Fertilizers in Vertisols of Southern Tigray, Ethiopia. Adv. Plants Agric. Res. 2014, 1. [Google Scholar] [CrossRef] [Green Version]
- Dagnaw, H.B. Influence of Nitrogen Fertilizer Rates and Varieties on Grain yield, Grain Nutrition and Injera Sensory Quality of Tef [Eragrostis tef (Zucc.) Trotter] Varieties. Master’s Thesis, Adis Ababa University, Addis Ababa, Ethiopia, 2018. [Google Scholar]
- Paff, K.; Asseng, S. A review of tef physiology for developing a tef crop model. Eur. J. Agron. 2018, 94, 54–66. [Google Scholar] [CrossRef]
- Halse, N.J.; Greenwood, E.A.N.; Lapins, P.; Boundy, C.A.P. An analysis of the effects of nitrogen deficiency on the growth and yield of a Western Australian wheat crop. Aust. J. Agric. Res. 1969, 20, 987–998. [Google Scholar] [CrossRef]
- Van Oosterom, E.J.; Borrell, A.K.; Chapman, S.C.; Broad, I.J.; Hammer, G.L. Functional dynamics of the nitrogen balance of sorghum: I. N demand of vegetative plant parts. Field Crop. Res. 2010, 115, 19–28. [Google Scholar] [CrossRef]
- Massignam, A.M.; Chapman, S.C.; Hammer, G.L.; Fukai, S. Physiological determinants of maize and sunflower grain yield as affected by nitrogen supply. Field Crop. Res. 2009, 113, 256–267. [Google Scholar] [CrossRef]
- Gungula, D.T.; Kling, J.G.; Togun, A.O. CERES-Maize predictions of maize phenology under nitrogen-stressed conditions in Nigeria. Agron. J. 2003, 95, 892–899. [Google Scholar] [CrossRef]
- Birch, C.J.; Long, K.E. Effect of nitrogen on the growth, yield and grain protein content of barley (Hordeum vulgare). Aust. J. Exp. Agric. 1990, 30, 237–242. [Google Scholar] [CrossRef]
- Williams, R.L.; Angus, J.F. Deep floodwater protects high-nitrogen rice crops from low-temperature damage. Aust. J. Exp. Agric. 1994, 34, 927–932. [Google Scholar] [CrossRef]
- Hall, A.J.; Savin, R.; Slafer, G.A. Is time to flowering in wheat and barley influenced by nitrogen? A critical appraisal of recent published reports. Eur. J. Agron. 2014, 54, 40–46. [Google Scholar] [CrossRef]
- Gashu, K. Response of teff (Eragrostis Tef (Zucc.) Trotter) to Nitrogen, Phosphorus, and Potassium Fertilization. Master’s Thesis, H. Smith Fuculty of Agriculture, Food, and Environment of the Hebrew University of Jerusalem, Jerusalem, Israel, 2017. [Google Scholar]
- Mulugeta, D.; Mamo, T.; Sokolowski, E.; Nachmansohn, J. Potash fertilization of teff and wheat in the highlands of Ethiopia. Electron. Int. Fertil. Corresp. 2017, 48, 12–19. [Google Scholar]
- Demiss, M.; Beyene, S.; Kidanu, S. Biomass accumulation and potassium concentrations in tissue of Teff (Eragrostis tef Zucc. Trotter) at three growth stages in Vertisols and Nitisols of the Central Highlands of Ethiopia. Afr. J. Agric. Res. 2019, 146, 345–353. [Google Scholar]
- EthioSIS (Ethiopia Soil Information System). Soil Fertility Status and Fertilizer Recommendation Atlas for Tigray Regional State; Agricultural Transformation Agency: Ethiopia, Addis Ababa, 2014. [Google Scholar]
- Misskire, Y.; Mamo, T.; Taddesse, A.M.; Yermiyahu, U. The effect of potassium on yield, nutrient uptake and efficiency of teff (Eragrostis tef Zucc. Trotter) on vertisols of North Western Ethiopian Highlands. J. Plant Nutr. 2019, 42, 307–322. [Google Scholar] [CrossRef]
- Albornoz, F. Crop responses to nitrogen overfertilization: A review. Sci. Hortic. 2016, 205, 79–83. [Google Scholar] [CrossRef]
- Tesfahunegn, G.B. Response of Yield and Yield Components of Tef [Eragrostis tef (Zucc.) Trotter] to Tillage, Nutrient, and Weed Management Practices in Dura Area, Northern Ethiopia. Int. Sch. Res. Not. 2014, 2014, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Tesfay, T.; Gebresamuel, G. Agronomic and economic evaluations of compound fertilizer applications under different planting methods and seed rates of tef [Eragrostis tef (Zucc.) Trotter] in Northern Ethiopia. J. Drylands 2016, 6, 409–422. [Google Scholar]
- Gebrehawariyat, F.M.; Haile, W.; Mamo, T.; Zipori, I.; Sokolowski, E. Response of teff [Eragrostis tef (Zucc.) Trotter] to potassium fertilizer application in four districts of North Shewa, Ethiopia. Electron. Int. Fertil. Corresp. 2018, 53, 3–15. [Google Scholar]
- Ben-Zeev, S.; Rabinovitz, O.; Orlov-Levin, V.; Chen, A.; Graff, N.; Goldwasser, Y.; Saranga, Y. Less Is More: Lower Sowing Rate of Irrigated Tef (Eragrostis tef) Alters Plant Morphology and Reduces Lodging. Agronomy 2020, 10, 570. [Google Scholar] [CrossRef]
- Van Delden, S.H. On Seed Physiology, Biomechanics and Plant Phenology in Eragrostis tef. Ph.D., Dissertation, Wageningen University, Wageningen, The Netherlands. 2011. Available online: https://library.wur.nl/WebQuery/wurpubs/fulltext/169451 (accessed on 5 November 2020).
- Lemaire, G.; Jeuffroy, M.H.; Gastal, F. Diagnosis tool for plant and crop N status in vegetative stage: Theory and practices for crop N management. Eur. J. Agron. 2008, 28, 614–624. [Google Scholar] [CrossRef]
- Justes, E.; Mary, B.; Meynard, J.M.; Machet, J.M.; Thelier-Huché, L. Determination of a Critical Nitrogen Dilution Curve for Winter Wheat Crops. Ann. Bot. 1994, 74, 397–407. [Google Scholar] [CrossRef]
- Colnenne, C.; Meynard, J.M.; Reau, R.; Justes, E.; Merrien, A. Determination of a Critical Nitrogen Dilution Curve for Winter Oilseed Rape. Ann. Bot. 1998, 81, 311–317. [Google Scholar] [CrossRef] [Green Version]
- Ata-Ul-Karim, S.T.; Liu, X.; Lu, Z.; Zheng, H.; Cao, W.; Zhu, Y. Estimation of nitrogen fertilizer requirement for rice crop using critical nitrogen dilution curve. Field Crop. Res. 2017, 201, 32–40. [Google Scholar] [CrossRef]
- Monostori, I.; Árendás, T.; Hoffman, B.; Galiba, G.; Gierczik, K.; Szira, F.; Vágújfalvi, A. Relationship between SPAD value and grain yield can be affected by cultivar, environment and soil nitrogen content in wheat. Euphytica 2016, 211, 103–112. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Chemical Properties | ||||||||
|---|---|---|---|---|---|---|---|---|
| Depth (cm) | EC (dS/m) | pH | Water Content | K (mg/L) | Cl (mg/L) | P (mg/L) | NO3 (mg/L) | NH4 (mg/kg soil) |
| 0–30 | 1.16 ± 0.09 | 7.9 ± 0.06 | 56 ± 1.8 | 38.8 ± 7.1 | 90.2 ± 20.8 | 1.1 ± 0.21 | 0.2 ± 0.04 | 3.0 ± 0.39 |
| 30–60 | 0.75 ± 0.07 | 8.2 ± 0.05 | 55 ± 1.0 | 8.7 ± 2.2 | 74.2 ± 11.6 | 0.7 ± 0.09 | 0.3 ± 0.18 | 0.9 ± 0.11 |
| 60–90 | 0.91 ± 0.22 | 8.2 ± 0.09 | 53 ± 1.8 | 3.7 ± 0.7 | 146.4 ± 70.0 | 0.4 ± 0.07 | 0.3 ± 0.03 | 0.7 ± 0.06 |
| Nitrogen Treatment (mg L−1) | Height (cm) | Panicle Length (cm) | Days to Flowering | Harvest Index | SPAD | # of Tillers | Lodging Index | |
|---|---|---|---|---|---|---|---|---|
| Genotype 405B | 10 | 38.6 | 36 | 49.8 | 0.35 | 28.6 | 6.1 | 1.2 |
| 20 | 43.5 | 38 | 48.4 | 0.3 | 32.2 | 7.3 | 1.3 | |
| 40 | 45.9 | 40.6 | 47 | 0.29 | 40.6 | 10.2 | 1.9 | |
| 80 | 44.7 | 40.4 | 46 | 0.27 | 40.9 | 9.9 | 2.7 | |
| 120 | 45.6 | 37.6 | 44 | 0.16 | 41.2 | 12.8 | 2.1 | |
| Genotype 406W | 10 | 55.3 | 31 | 57 | 0.36 | 29.2 | 2.7 | 1.5 |
| 20 | 58.7 | 37.2 | 55.2 | 0.33 | 30.5 | 4.2 | 1.6 | |
| 40 | 60 | 37.8 | 53.8 | 0.28 | 38.1 | 5.5 | 1.9 | |
| 80 | 57 | 40.6 | 52.6 | 0.28 | 38.7 | 5.4 | 2.4 | |
| 120 | 60.7 | 38.8 | 52.2 | 0.13 | 38.9 | 7.8 | 2.1 | |
| Tukey for N treatments | 10 | 46.9 B | 33.5 B | 53.4 A | 0.35 A | 28.9 C | 4.4 D | 1.4 C |
| 20 | 51.1 A | 37.6 AB | 51.8 AB | 0.32 A | 31.4 B | 5.7 C | 1.4 C | |
| 40 | 52.9 A | 39.2 A | 50.4 BC | 0.28 A | 39.3 A | 7.8 B | 1.9 BC | |
| 80 | 50.8 A | 40.5 A | 49.3 C | 0.28 A | 39.9 A | 7.6 B | 2.6 A | |
| 120 | 53.1 A | 38.2 A | 48.1 C | 0.15 B | 30.1 A | 10.3 A | 2.1 AB | |
| Tukey for genotype | 405B | 47.0 b | 38.5 a | 47.0 b | 0.28 a | 36.7 a | 9.3 a | 1.8 a |
| 406W | 54.1 a | 37.1a | 54.1 a | 0.27 a | 35.1 b | 5.1 b | 1.9 a | |
| N × genotype | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. |
| Nitrogen Treatment (mg L−1) | Height (cm) | Panicle Length (cm) | Days to Flowering | Harvest Index | SPAD | # of Tillers | |
|---|---|---|---|---|---|---|---|
| Genotype 405B | 0 | 100.9 | 29.4 | 41 | 0.08 | 31.7 | 8.2 |
| 30 | 95.8 | 29.8 | 38.6 | 0.09 | 34.9 | 11.6 | |
| 60 | 100.7 | 33.1 | 37.4 | 0.1 | 37.3 | 13.8 | |
| 120 | 99.2 | 28.7 | 36.6 | 0.09 | 36.4 | 16.4 | |
| Genotype 406W | 0 | 89.9 | 26.7 | 59.2 | 0.04 | 33.5 | 5.4 |
| 30 | 104.4 | 25.1 | 55 | 0.05 | 33.4 | 6.8 | |
| 60 | 94.6 | 34.6 | 56.4 | 0.05 | 37.3 | 11.2 | |
| 120 | 95.4 | 25.8 | 55 | 0.03 | 40.5 | 13 | |
| Tukey’s for N treatments | 0 | 95.4 A | 28.1 A | 50.1 A | 0.06 A | 31.6 C | 6.8 A |
| 30 | 100.1 A | 27.5 A | 46.8 A | 0.07 A | 34.1BC | 9.2 A | |
| 60 | 97.7 A | 33.9 A | 46.9 A | 0.08 A | 37.3 AB | 12.5 A | |
| 120 | 97.3 A | 27.3 A | 45.8 A | 0.06 A | 38.4 A | 14.7 A | |
| Tukey’s for genotype | 405B | 99.1 a | 30.3 a | 38.4 b | 0.09 a | 35.1 a | 12.5 a |
| 406W | 96.1 a | 28.1 a | 56.4 a | 0.04 b | 35.7 a | 9.1 a | |
| N × genotype | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gashu, K.; Halpern, M.; Zipori, I.; Bustan, A.; Saranga, Y.; Yermiyahu, U. Tef (Eragrostis tef Trotter) Responses to Nitrogen Fertigation under Semi-Arid Mediterranean Climate. Agronomy 2020, 10, 1870. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10121870
Gashu K, Halpern M, Zipori I, Bustan A, Saranga Y, Yermiyahu U. Tef (Eragrostis tef Trotter) Responses to Nitrogen Fertigation under Semi-Arid Mediterranean Climate. Agronomy. 2020; 10(12):1870. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10121870
Chicago/Turabian StyleGashu, Kelem, Moshe Halpern, Isaac Zipori, Amnon Bustan, Yehoshua Saranga, and Uri Yermiyahu. 2020. "Tef (Eragrostis tef Trotter) Responses to Nitrogen Fertigation under Semi-Arid Mediterranean Climate" Agronomy 10, no. 12: 1870. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10121870