Assessing the Potential of Extra-Early Maturing Landraces for Improving Tolerance to Drought, Heat, and Both Combined Stresses in Maize

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Agronomic Management
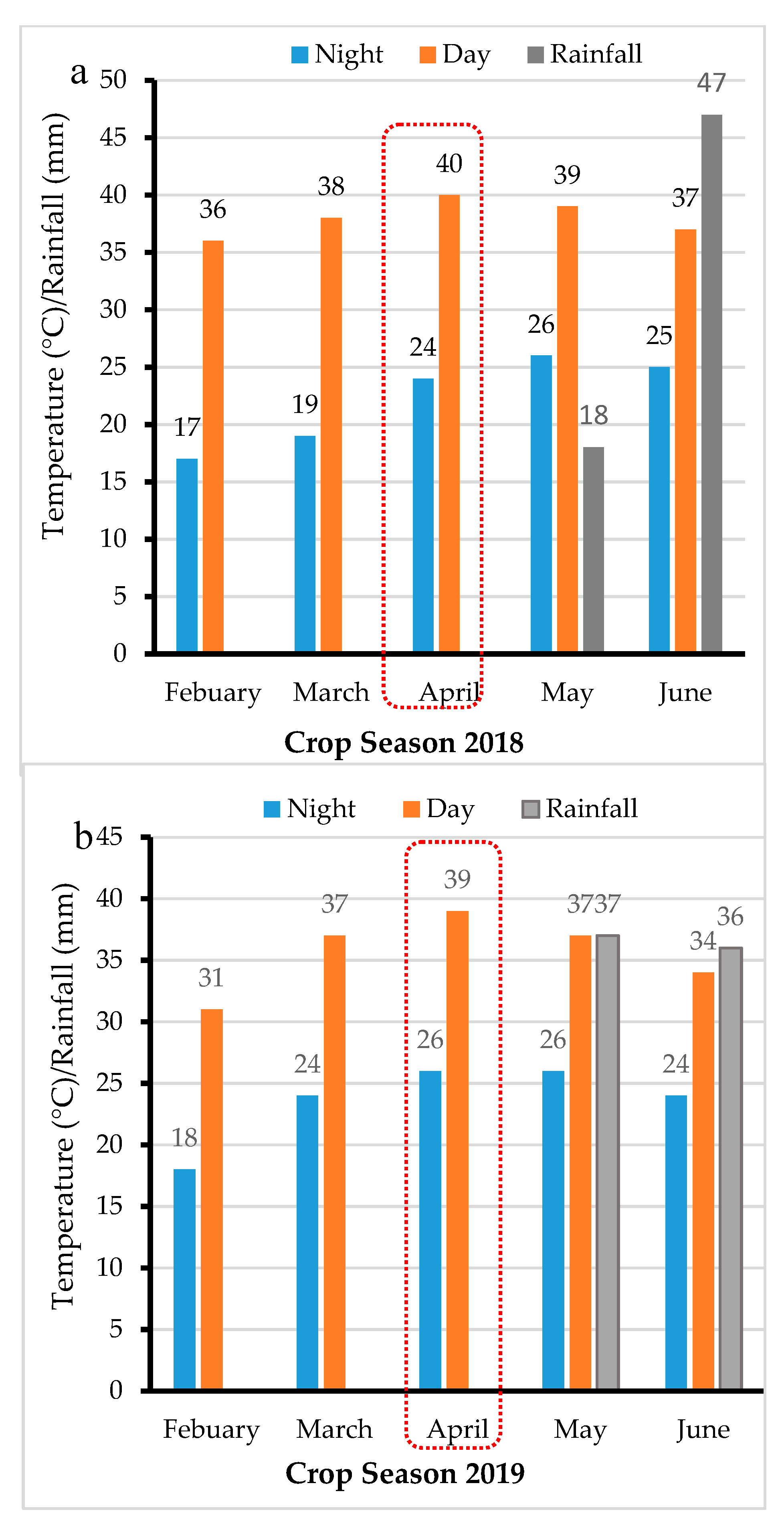
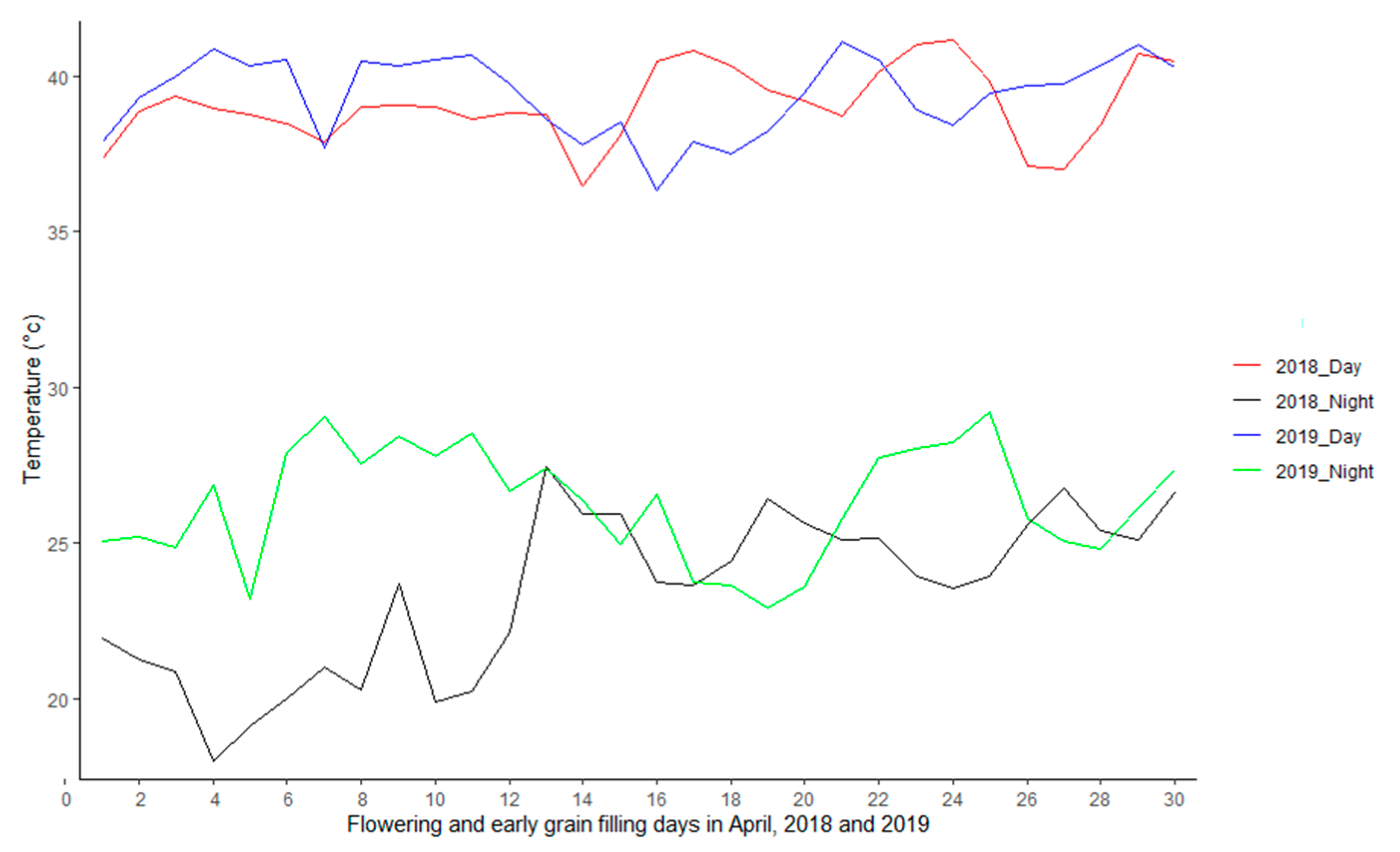
2.2.1. Drought Stress Trials
2.2.2. Heat and Combined Drought and Heat Stress Trials
2.3. Trait Measurement
2.4. Statistical Analysis
3. Results
3.1. Analysis of Variance and Broad-Sense Heritability
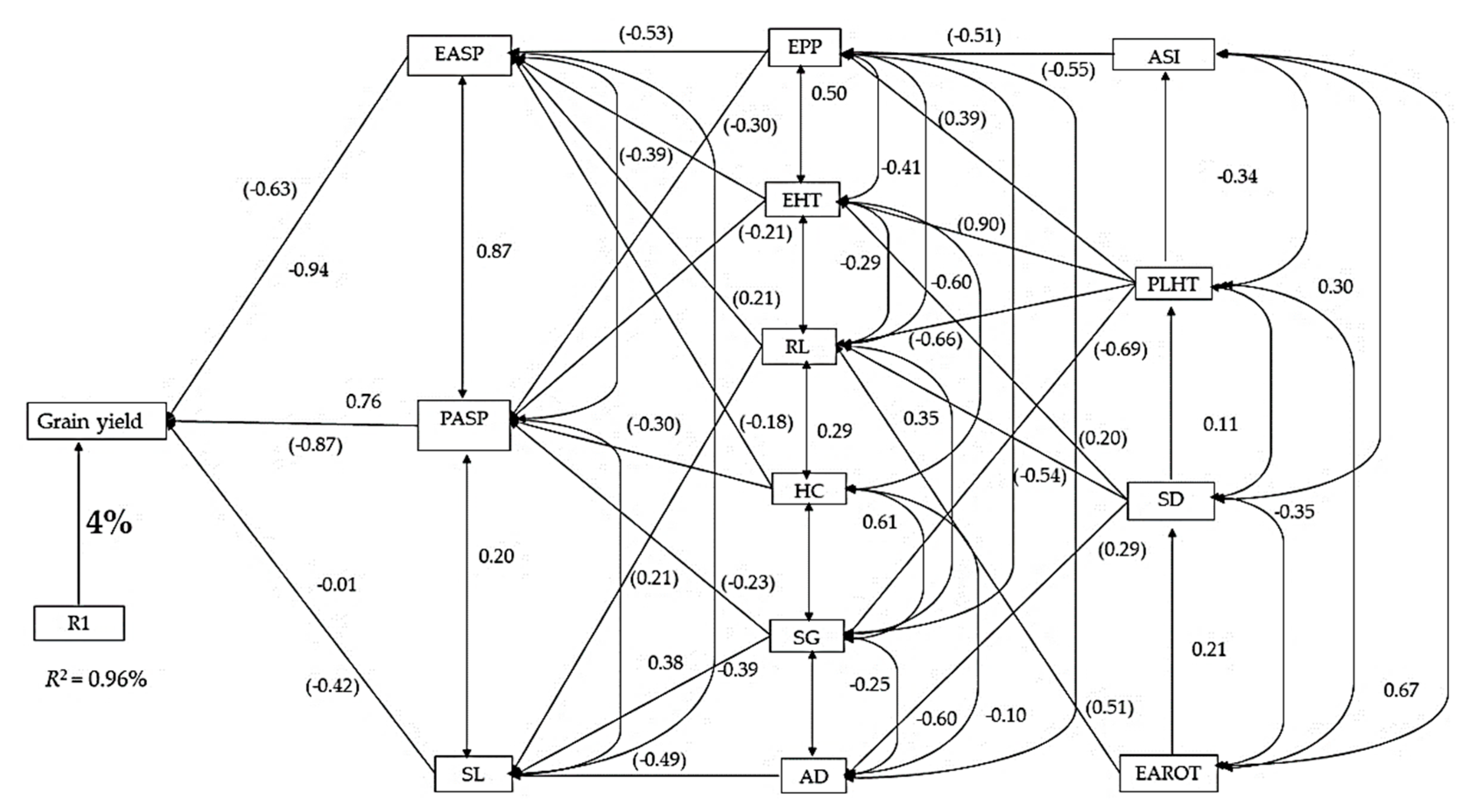
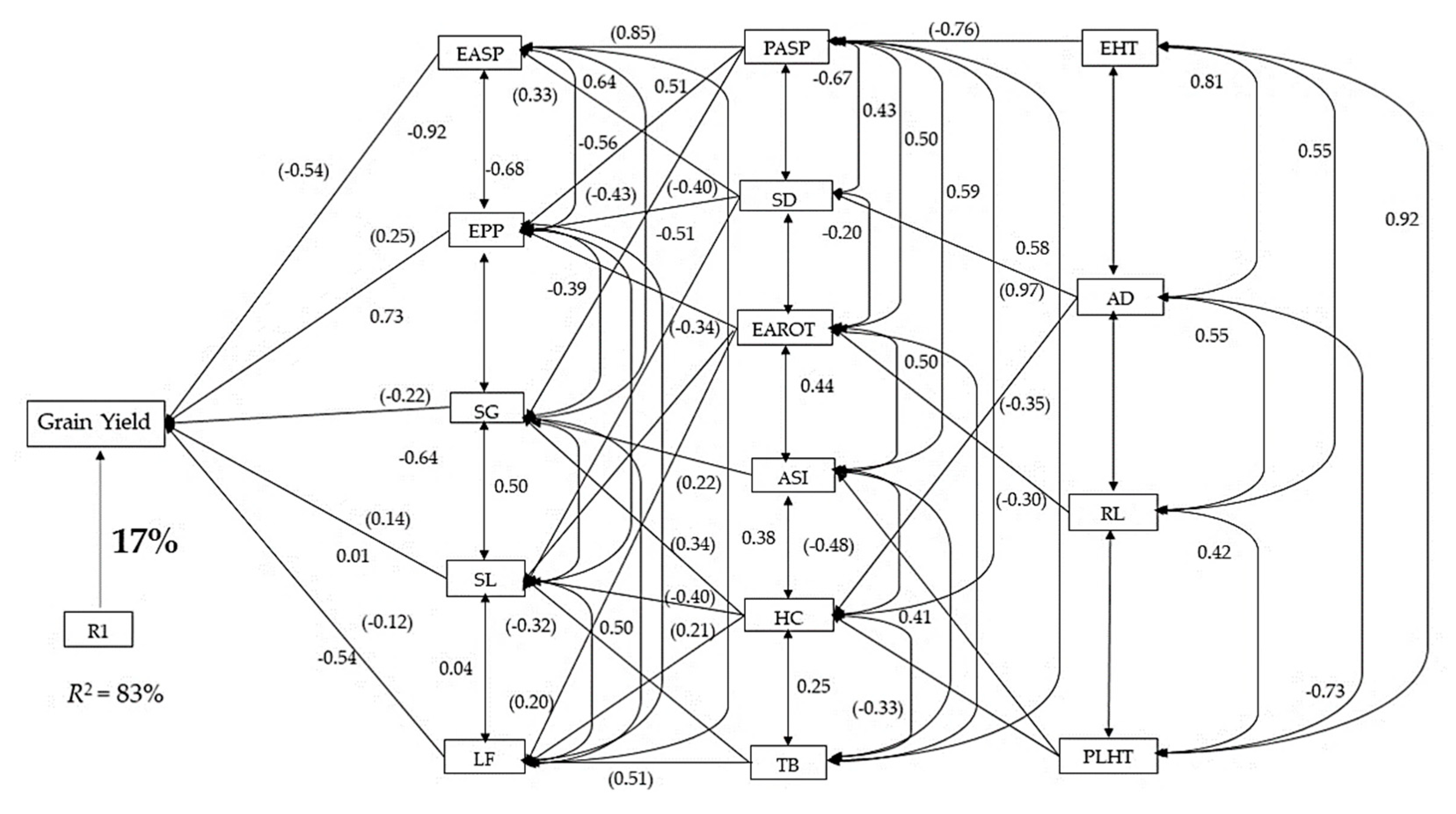
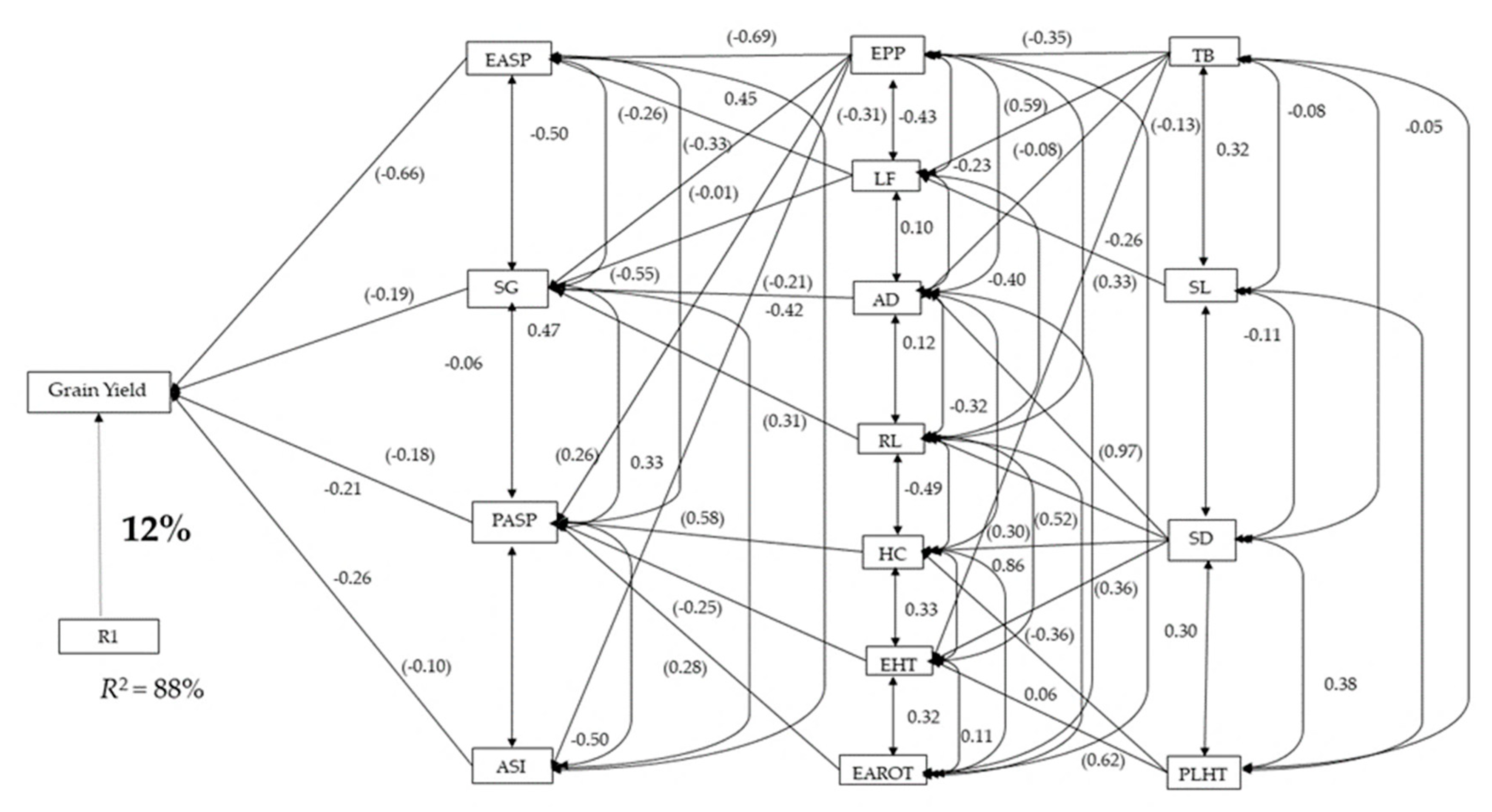
3.2. Genetic Correlations and Sequential Regression Analysis
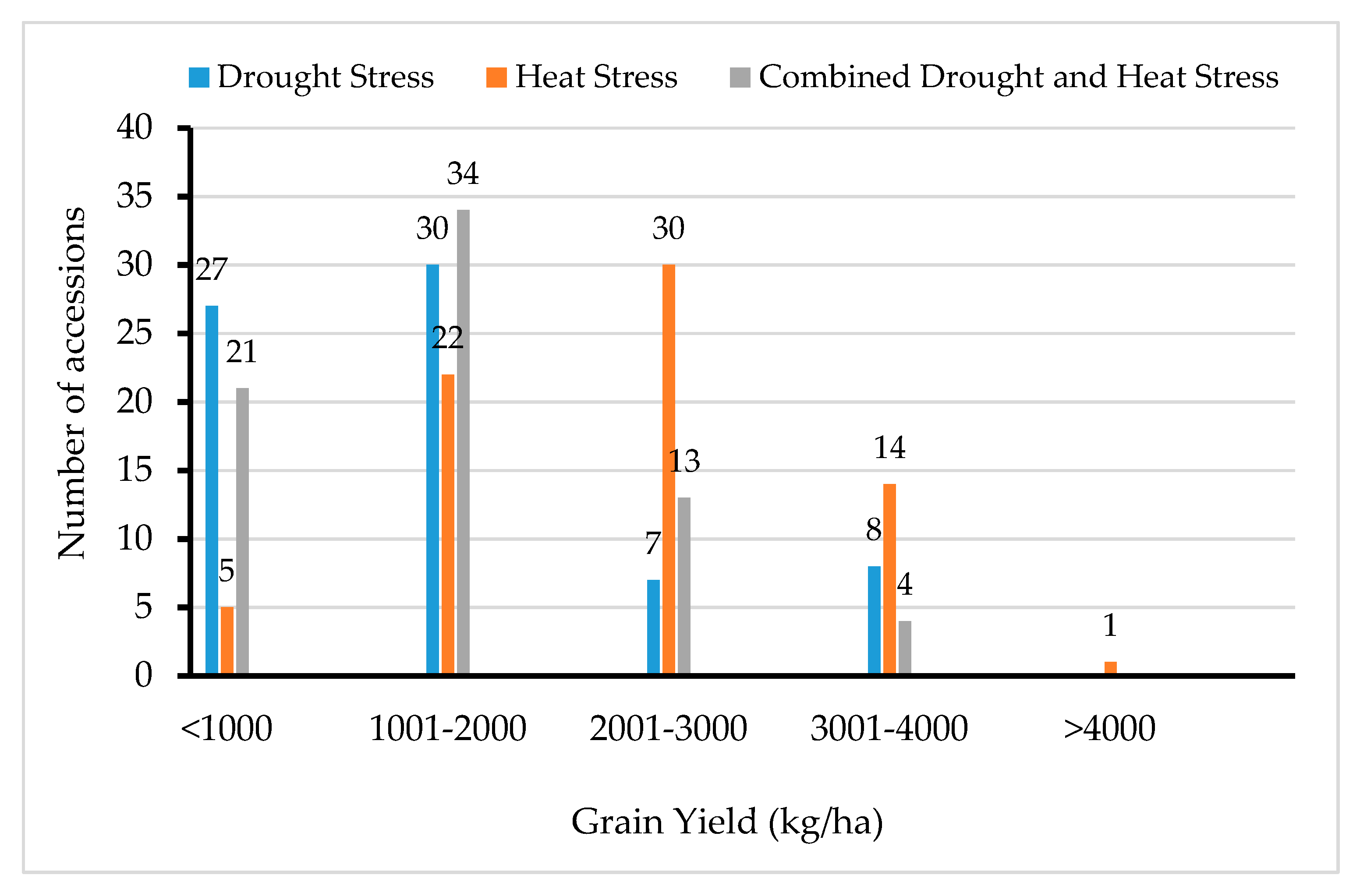
3.3. Performance of Accessions under the Contrasting Environment
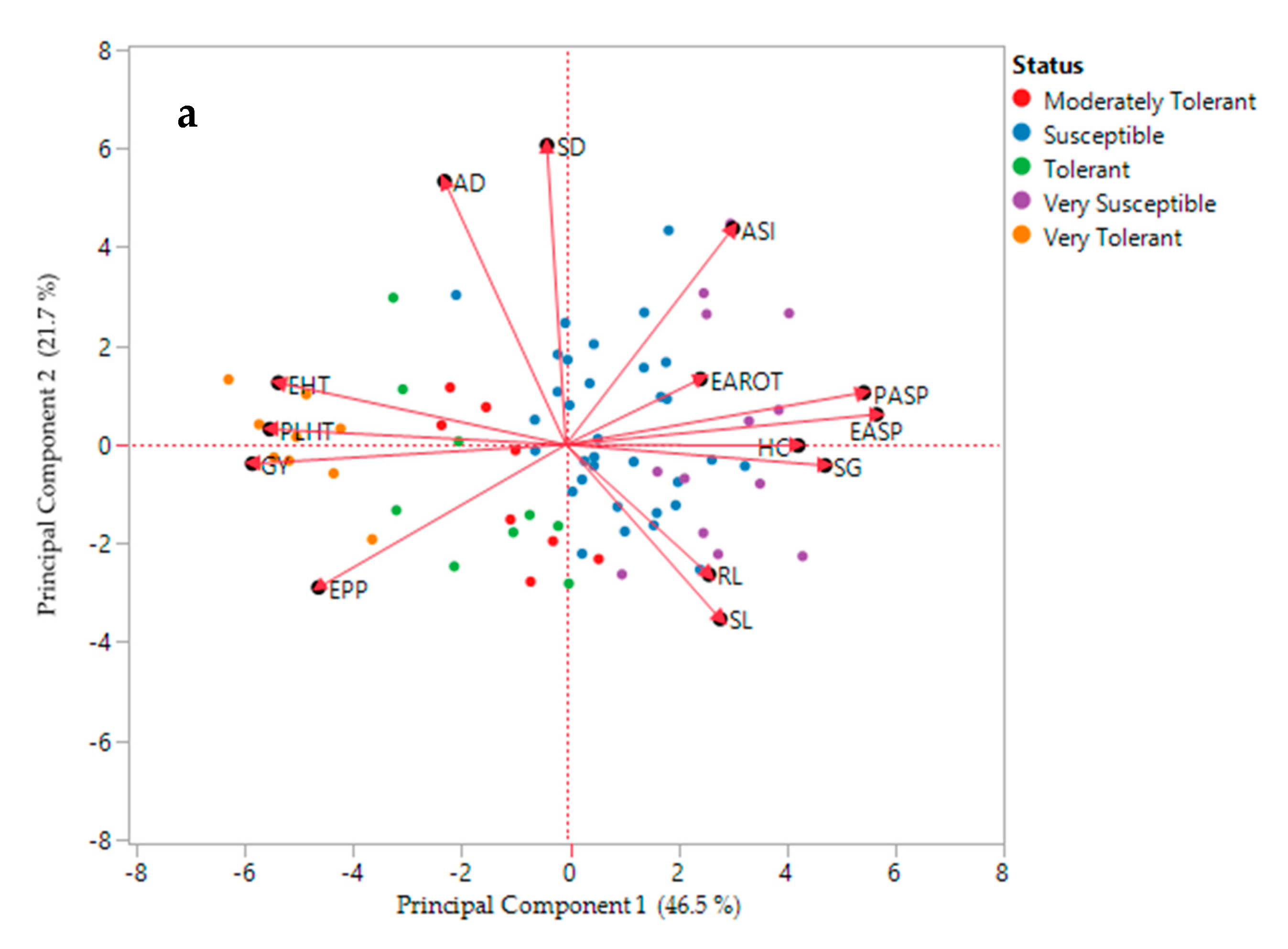
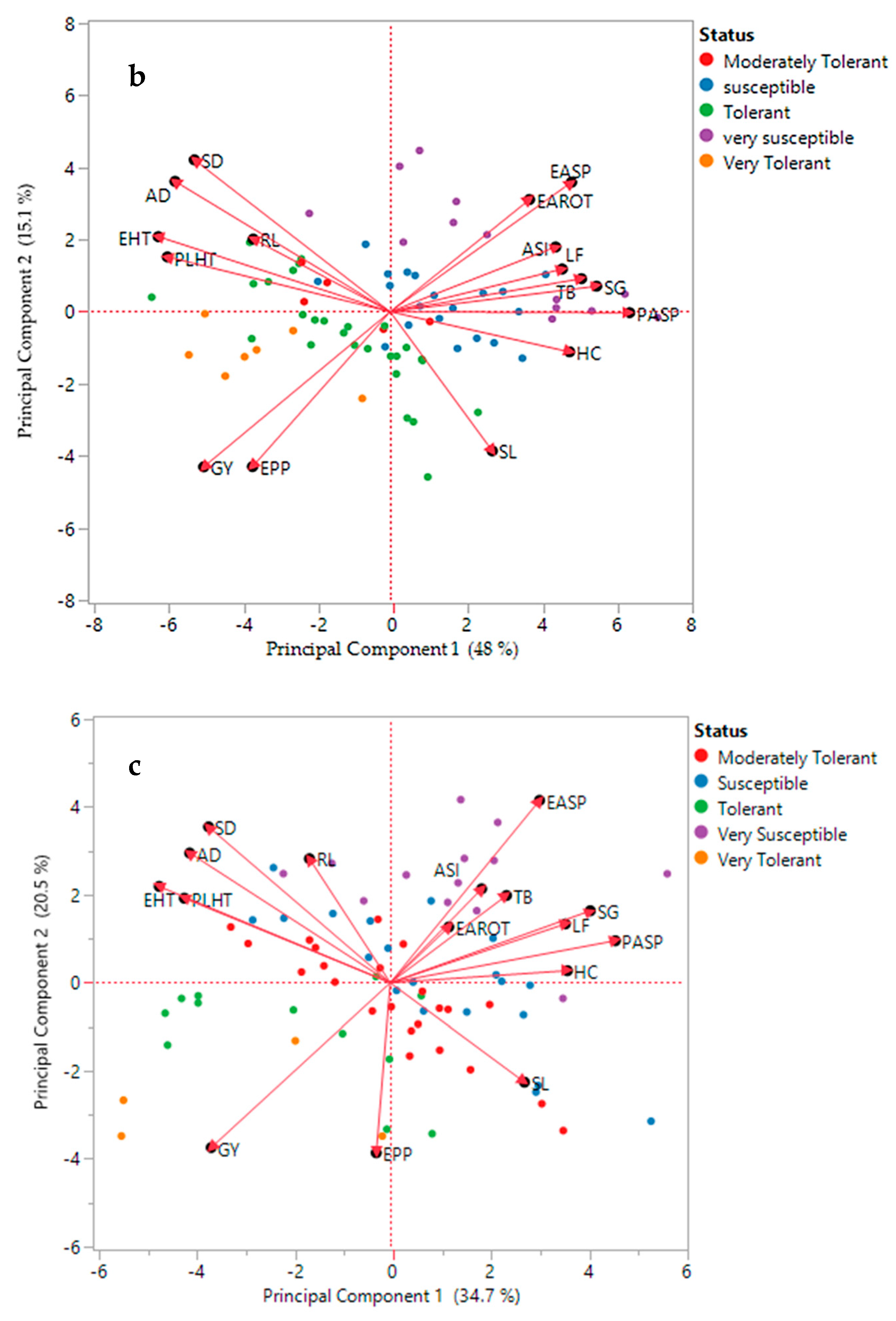
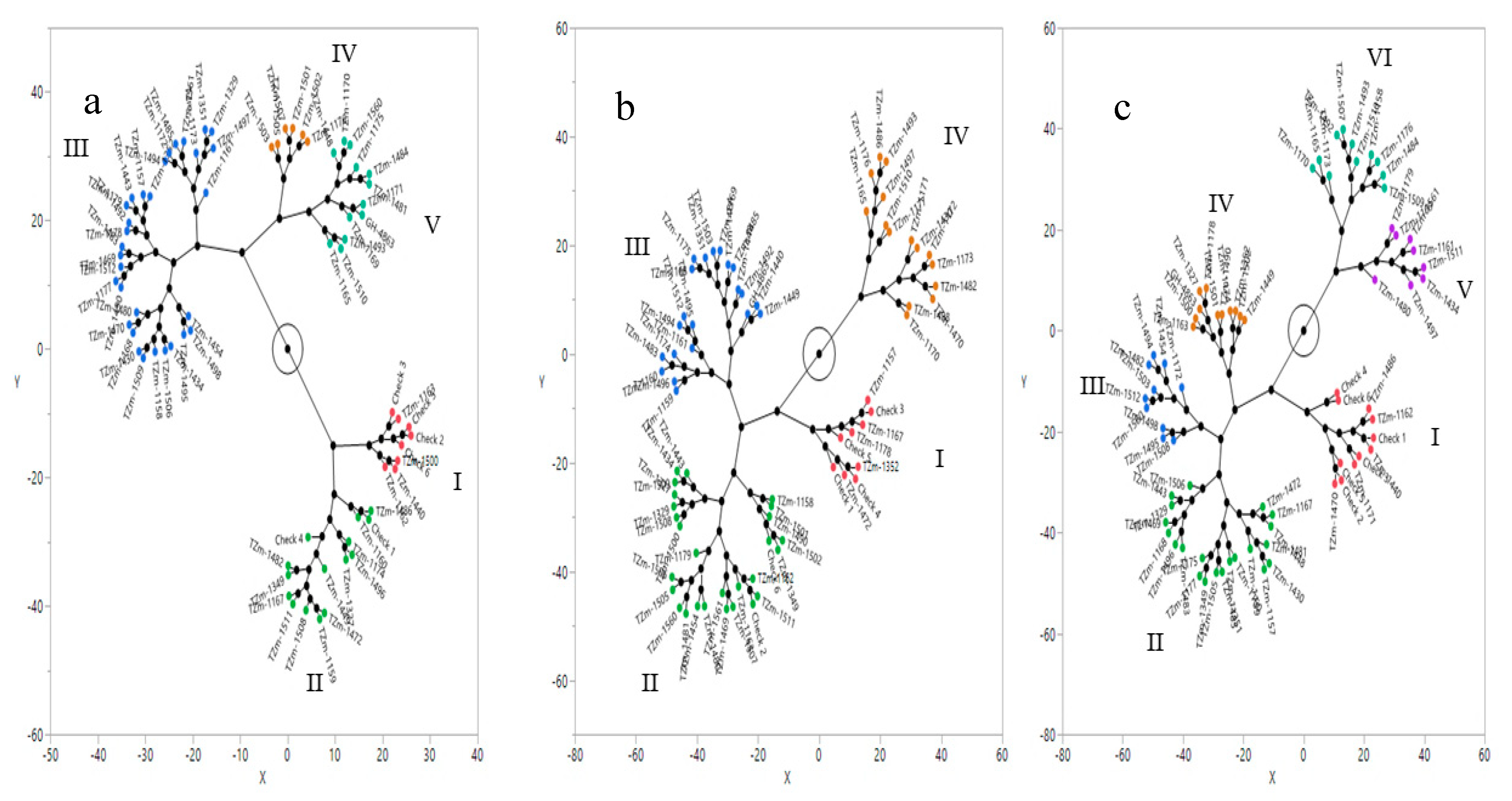
3.4. Principal Component Biplot and Cluster Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McCann, J.C. Maize and Grace: Africa’s Encounter with a New World Crop, 1500–2000; Harvard University Press: Cambridge, MA, USA, 2005. [Google Scholar]
- Warburton, M.L.; Reif, J.C.; Frisch, M.; Bohn, M.; Bedoya, C.; Xia, X.C.; Crossa, J.; Franco, J.; Hoisington, D.; Pixley, K.; et al. Genetic diversity in CIMMYT non-temperate maize germplasm: Landraces, open pollinated varieties, and inbred lines. Crop Sci. 2008, 48, 617–624. [Google Scholar] [CrossRef] [Green Version]
- Abate, T.; Fisher, M.; Abdoulaye, T.; Kassie, G.T.; Luduka, R.; Marenya, P.; Asnake, W. Characteristics of maize cultivars in Africa: How modern are they and how many do smallholder farmers grow? Agric. Food Secur. 2017, 6, 30. [Google Scholar] [CrossRef] [Green Version]
- Edmeades, G.O.; Trevisan, W.; Prasanna, B.M.; Campos, H. Tropical maize (Zea mays L.). In Genetic Improvement of Tropical Crops; Campos, H., Caligari, P.D.S., Eds.; Springer: Cham, Switzerland, 2017; pp. 57–109. [Google Scholar]
- Wu, X.; Wang, A.; Guo, X.; Liu, P.; Zhu, Y.; Li, X.; Chen, Z. Genetic characterization of maize germplasm derived from Suwan population and temperate resources. Hereditas 2019, 156, 2. [Google Scholar] [CrossRef]
- Singh, S.; Vikram, P.; Sehgal, D.; Burgueño, J.; Sharma, A.; Singh, S.K.; Sansaloni, C.P.; Joynson, R.; Brabbs, T.; Ortiz, C.; et al. Harnessing genetic potential of wheat germplasm banks through impact-oriented-prebreeding for future food and nutritional security. Sci. Rep. 2018, 8, 12527. [Google Scholar] [CrossRef] [Green Version]
- Masuka, B.P.; van Biljon, A.; Cairns, J.E.; Das, B.; Labuschagne, M.; MacRobert, J.; Olsen, M. Genetic diversity among selected elite CIMMYT maize hybrids in East and Southern Africa. Crop Sci. 2017, 57, 2395–2404. [Google Scholar] [CrossRef] [Green Version]
- Edenhofer, O. (Ed.) Climate Change 2014: Mitigation of Climate Change; Cambridge University Press: Cambridge, UK, 2015; p. 3. [Google Scholar]
- Muller, C.; Cramer, W.; Hare, W.L.; Lotze-Campen, H. Climate change risks for African agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 4313–4315. [Google Scholar] [CrossRef] [Green Version]
- Kurukulasuriya, P.; Mendelsohn, R.; Hassan, R.; Benhin, J.; Deressa, T.; Diop, M.; Eid, H.M.; Fosu, K.Y.; Gbetibouo, G.; Jain, S.; et al. Will African agriculture survive climate change? World Bank Econ. Rev. 2006, 20, 367–388. [Google Scholar] [CrossRef] [Green Version]
- Cairns, J.E.; Hellin, J.; Sonder, K.; Araus, J.L.; MacRobert, J.F.; Thierfelder, C.; Prasanna, B.M. Adapting maize production to climate change in Sub-Saharan Africa. Food Secur. 2012, 5, 345–360. [Google Scholar] [CrossRef] [Green Version]
- Knox, J.; Hess, T.; Daccache, A.; Wheeler, T. Climate change impacts on crop productivity in Africa and South Asia. Environ. Res. 2012, 7, 034032. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B. Improvement of Early and Extra-Early Maize for Combined Tolerance to Drought and Heat Stress in Sub-Saharan Africa. In Advances in Genetic Enhancement of Early and Extra-Early Maize for Sub-Saharan Africa; Springer: Cham, Switzerland, 2017; pp. 311–358. [Google Scholar]
- Cicchino, M.; Rattalino-Edreria, J.I.; Uribelarrea, M.; Otegui, M.E. Heat stress in field-grown maize: Response of physiological determinants of grain yield. Crop Sci. 2011, 50, 1438–1448. [Google Scholar] [CrossRef]
- Zaidi, P.H.; Zaman-Allah, M.; Trachsel, S.; Seetharam, K.; Cairns, J.E.; Vinayan, M.T. Phenotyping for Abiotic Stress Tolerance in Maize Heat Stress: A Field Manual; CIMMYT: Mexico City, Mexico, 2016. [Google Scholar]
- Araus, J.L.; Serret, M.D.; Edmeades, G.O. Phenotyping maize for adaptation to drought. Front. Physiol. 2012, 3, 305. [Google Scholar] [CrossRef] [Green Version]
- NeSmith, D.S.; Ritchie, J.T. Effects of soil water-deficits during tassel emergence on development and yield components of maize (Zea mays L.). Field Crop Res. 1992, 28, 251–256. [Google Scholar] [CrossRef]
- Meseka, S.; Menkir, A.; Bossey, B.; Mengesha, W. Performance Assessment of Drought Tolerant Maize Hybrids under Combined Drought and Heat Stress. Agronomy 2018, 8, 274. [Google Scholar] [CrossRef] [Green Version]
- Nelimor, C.; Badu-Apraku, B.; Tetteh, A.Y.; N’guetta, A.S.P. Assessment of Genetic Diversity for Drought, Heat and Combined Drought and Heat Stress Tolerance in Early Maturing Maize Landraces. Plants 2019, 8, 518. [Google Scholar] [CrossRef] [Green Version]
- Bassu, S.; Brisson, N.; Durand, J.L.; Boote, K.; Lizaso, J.; Jones, J.W.; Basso, B. How do various maize crop models vary in their responses to climate change factors? Glob. Chang. Biol. 2014, 20, 2301–2320. [Google Scholar] [CrossRef]
- Wardlaw, I.F. Interaction between drought and chronic high temperature during kernel filling in wheat in a controlled environment. Ann. Bot. 2002, 90, 469–476. [Google Scholar] [CrossRef] [Green Version]
- Shah, N.H.; Paulsen, G.M. Interaction of drought and high temperature on photosynthesis and grain-filling of wheat. Plant Soil 2003, 257, 219–226. [Google Scholar] [CrossRef]
- Prasad, P.V.V.; Pisipati, S.R.; Momicilovic, I.; Ristic, Z. Independent and combined effects of high temperature and drought stress during grain filling on plant yield and chloroplast EF-Tu expression in spring wheat. J. Agron. Crop Sci. 2011, 197, 430–441. [Google Scholar] [CrossRef]
- Pradhan, G.P.; Prasad, P.V.V.; Fritz, A.K.; Kirkham, M.B.; Gil, B.S. Effects of drought and high temperature stress on synthetic hexaploid wheat. Funct. Plant Biol. 2012, 39, 190–198. [Google Scholar] [CrossRef]
- Heiniger, R.W. The Impact of Early Drought on Corn Yield; North Carolina State University: Raleigh, NC, USA, 2001. [Google Scholar]
- Dwivedi, S.L.; Ceccarelli, S.; Blair, M.W.; Upadhyaya, H.D.; Are, A.K.; Ortiz, R. Landrace germplasm for improving yield and abiotic stress adaptation. Trends Plant Sci. 2016, 21, 31–42. [Google Scholar] [CrossRef]
- Castro-Nava, S.; Ramos-Ortiz, V.H.; Reyes-Méndez, C.A.; Briones-Encinia, C.A.F.; López-Santillán, J.A. Preliminary field screening of maize landrace germplasm from northeastern Mexico under high temperatures. Maydica 2011, 56, 77–82. [Google Scholar]
- Gedil, M.; Menkir, A. An integrated molecular and conventional breeding scheme for enhancing genetic gain in maize in Africa. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Cairns, J.E.; Crossa, J.; Zaidi, P.H.; Grudloyma, P.; Sanchez, C.; Araus, J.L.; Menkir, A. Identification of drought, heat, and combined drought and heat tolerant donors in maize. Crop Sci. 2013, 53, 1335–1346. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.S.; Seetharam, K.; Zaidi, P.H.; Dinesh, A.; Vinayan, M.T.; Nath, U.K. Dissecting heat stress tolerance in tropical maize (Zea mays L.). Field Crop. Res. 2017, 204, 110–119. [Google Scholar] [CrossRef]
- Nelimor, C.; Badu-Apraku, B.; Nguetta, S.P.A.; Tetteh, A.Y.; Garcia-Oliveira, A.L. Phenotypic characterization of maize landraces from Sahel and Coastal West Africa reveals marked diversity and potential for genetic improvement. J. Crop Improv. 2019. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS User’s Guide: Statistics; Version 9.4; SAS Institute Inc.: Cary, NC, USA, 2017. [Google Scholar]
- Falconer, D.S.; Mackay, T.F.C. Introduction to Quantitative Genetics, 4th ed.; Longman: New York, NY, USA, 1996. [Google Scholar]
- Peterson, B.G.; Carl, P.; Boudt, K.; Bennett, R.; Ulrich, J.; Zivot, E.; Lestel, M.; Balkissoon, K.; Wuertz, D.; Peterson, M.B.G. Performance Analytics: Econometric Tools for Performance and Risk Analysis. R Package Version 1. Available online: https://cran.r-project.org/web/packages/PerformanceAnalytics/index.html (accessed on 15 August 2014).
- Cooper, M.; DeLacy, I.H.; Basford, K.E. Relationship among analytical methods used to analyse genotypic adaptation in multi-environment trials. In Plant Adaptation and Crop Improvement; Cooper, M., Hammer, G.L., Eds.; Cambridge University Press: Cambridge, UK, 1996; pp. 193–224. [Google Scholar]
- Vargas, M.; Combs, E.; Alvarado, G.; Atlin, G.; Mathews, K.; Crossa, J. META: A suit of SAS programs to analyse multi-environment breeding trials. Agron. J. 2013, 105, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, S.A.; Prasanna, B.M.; Singh, N.N. Sequential path model for determining interrelationships among grain yield and related characters in maize. Crop Sci. 2003, 43, 1690–1697. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Akinwale, R.O.; Oyekunle, M. Efficiency of secondary traits in selecting for improved grain yield in extra-early maize under Striga-infested and Striga-free environments. Plant Breed. 2014, 133, 373–380. [Google Scholar] [CrossRef]
- Talabi, A.O.; Badu-Apraku, B.; Fakorede, M.A.B. Genetic variances and relationship among traits of an early-maturing maize population under drought-stress and low-N environments. Crop Sci. 2016, 57, 681–692. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B.; Talabi, A.O.; Oyekunle, M.; Akaogu, I.C.; Akinwale, R.O.; Annor, B.; Melaku, G.; Fasanmade, Y.; Aderounmu, M. Gene action and heterotic groups of early white quality protein maize inbreds under multiple stress environments. Crop Sci. 2015, 56, 183–199. [Google Scholar] [CrossRef]
- Statistical Package for Social Sciences (SPSS) Inc. SPSS Base 17.0 for Windows User’s Guide; SPSS Inc.: Chicago, IL, USA, 2007. [Google Scholar]
- Bolaños, J.; Edmeades, G.O. Eight cycles of selection for drought tolerance in lowland tropical maize. I. Responses in grain yield, biomass, and radiation utilization. Field Crop. Res. 1993, 31, 233–252. [Google Scholar] [CrossRef]
- Araus, J.L.; Kefauver, S.C.; Zaman-Allah, M.; Olsen, M.S.; Cairns, J.E. Translating high-throughput phenotyping into genetic gain. Trends Plant Sci. 2018, 23, 451–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, B.; Atlin, G.N.; Olsen, M.; Burgueño, J.; Tarekegne, A.; Babu, R.; Ndou, E.N.; Mashingaidze, K.; Moremoholo, L.; Ligeyo, D.; et al. Identification of donors for low-nitrogen stress with maize lethal necrosis (MLN) tolerance for maize breeding in sub-Saharan Africa. Euphytica 2019, 215, 80. [Google Scholar] [CrossRef] [Green Version]
- Gouesnard, B.; Zanetto, B.; Welcker, C. Identification of adaptation traits to drought in collections of maize landraces from southern Europe and temperate regions. Euphytica 2015, 209, 565–584. [Google Scholar] [CrossRef]
- Mahrookashani, A.; Siebert, S.; Hüging, H.; Ewert, F. Independent and combined effects of high temperature and drought stress around anthesis on wheat. J. Agron. Crop Sci. 2017, 203, 453–463. [Google Scholar] [CrossRef]
- Edmeades, G.O.; Bolaños, J.; Chapman, S.C.; Lafiite, H.R.; Bänziger, M. Selection improves drought tolerance in tropical maize populations. I. Gains in biomass, grain yield and harvest index. Crop Sci. 1999, 39, 1306–1315. [Google Scholar] [CrossRef]
- Tandzi, L.N.; Bradley, G.; Mutengwa, C. Morphological responses of maize to drought, heat and combined drought and heat stresses. J. Biol. Sci. 2019, 19, 7–16. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-Stress Conditions (NS) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SV | df | GY | AD | SD | ASI | PLHT | EHT | HC | EPP | PASP | EASP | SG | RL | SL | EAROT |
| Env | 2 | 5,746,960.3** | 1.3 | 108.8* | 133.4** | 83913.4** | 32981.7** | 86.7** | 0.5** | 24.5** | 3.3* | - | - | - | - |
| Rep (Env) | 3 | 169,368.8 | 9.5* | 0.8 | 5.4 | 1485.9* | 1260.4** | 7.6** | 0 | 1.4* | 0 | - | - | - | - |
| Block (Env × Rep) | 42 | 530,887.1* | 3.4 | 8 | 5.2 | 690.1** | 139 | 1.3** | 0 | 0.9* | 1.2* | - | - | - | - |
| Genotype | 71 | 2,466,309.9** | 47.7** | 75.6** | 13.2** | 686.7** | 344.2** | 0.8* | 0.1** | 1.7** | 3.8** | - | - | - | - |
| Env × Genotype Error | 142 171 | 84,207.7 204,531.2 | 5.5** 2.5 | 13.9* 7.9 | 6.8* 5.1 | 180.3 214.4 | 85.9 95 | 0.5 0.4 | 0 0 | 0.4 0.4 | 0.4 0.7 | - - | - - | - - | - - |
| Repeatability | 0.94 | 0.89 | 0.81 | 0.5 | 0.75 | 0.77 | 0.41 | 0.7 | 0.82 | 0.86 | - | - | - | - | |
| Grand Mean | 2992.38 | 47 | 50 | 3 | 152.72 | 68.81 | 4 | 0.92 | 5 | 5 | - | - | - | - | |
| Drought Stress (DS) | |||||||||||||||
| Env | 1 | 3,976,484 | 75.4** | 130.7** | 19.4** | 10811.4** | 852.3** | 54.2** | 1.6** | 24.3** | 34.9** | 9.4** | 0.9** | 0.5** | - |
| Rep (Env) | 2 | 2,258,791 | 4.9* | 6.5* | 0.5 | 280.5 | 206.4* | 0.9 | 0 | 0.6 | 0.7 | 4.8** | 0.2** | 0 | - |
| Block (Env × Rep) | 28 | 1,039,093 | 3.2* | 3.4* | 0.6 | 402.9** | 177.5** | 0.7 | 0 | 0.5 | 0.5 | 0.94* | 0 | 0 | - |
| Genotype | 71 | 6,056,025.5 ** | 72.4** | 76.4** | 2.6** | 1531.6** | 946.2** | 3.0** | 0.1** | 4.2** | 4.2** | 1.6** | 0* | 0** | - |
| Env × Genotype | 71 | 1,840,190 | 3.9** | 5.2** | 1.5** | 105.2 | 43 | 1.0* | 0.0** | 0.6* | 0.8** | 0.37 | 0 | 0 | - |
| Error | 114 | 1,638,119 | 1.6 | 2 | 0.8 | 109.5 | 60 | 0.7 | 0 | 0.4 | 0.4 | 0.39 | 0 | 0 | - |
| Repeatability | 0.69 | 0.94 | 0.93 | 0.45 | 0.94 | 0.95 | 0.68 | 0.3 | 0.86 | 0.82 | 0.79 | 0.33 | 0.68 | - | |
| Grand Mean | 1523.92 | 46 | 50 | 4 | 116.73 | 54.95 | 4 | 0.68 | 5 | 5 | 4 | 0.1 | 0.22 | - | |
| Reduction (%) | 0.49 | 1 | - | 1 | 0.24 | 0.20 | - | 0.26 | - | - | - | - | - | - | |
| Heat Stress (HS) | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SV | df | GY | AD | SD | ASI | PLHT | EHT | HC | EPP | PASP | EASP | SG | RL | SL | TB | LF | EAROT |
| Env | 1 | 16,037,142.5** | 9.8 | 20.1* | 1.8 | 40110.6** | 23562.6** | 103.9** | 1.8** | 0.2 | 8.7* | 48.3** | 0.2** | 0.1* | 0.2** | 0.0* | - |
| Rep (Env) | 2 | 8,533,488.5** | 11.9 | 11.6 | 1 | 287.9 | 652.8* | 1.7 | 0.6** | 0.8 | 2.9* | 3.5* | 0.1** | 0 | 0.0* | 0 | - |
| Block (Env × Rep) | 28 | 1,970,909.7* | 9.1* | 9.2* | 1.1 | 1363.7** | 501.8** | 1 | 0.1 | 0.6 | 1.3* | 0.7 | 0 | 0 | 0 | 0 | - |
| Genotype | 71 | 2,330,918.4** | 59.9** | 51. 0** | 3.4** | 1181.9** | 716.6** | 1.5* | 0.1** | 2.0** | 2.0** | 1.4** | 0 | 0.0** | 0.0* | 0.0* | - |
| Env × Genotype | 71 | 753,004.1 | 4.5 | 3.2 | 1.3 | 286.3 | 168.4 | 0.9 | 0 | 0.3 | 0.6 | 0.6 | 0.0* | 0 | 0.0* | 0 | - |
| Error | 114 | 841,366 | 4.3 | 4.6 | 1.5 | 336.8 | 135.6 | 0.7 | 0 | 0.4 | 0.7 | 0.5 | 0 | 0 | 0 | 0 | - |
| Repeatability | 0.67 | 0.94 | 0.93 | 0.63 | 0.76 | 0.79 | 0.34 | 0.63 | 0.85 | 0.69 | 0.56 | 0.37 | 0.64 | 0.21 | 0.38 | - | |
| Mean | 2301.39 | 57 | 60 | 3 | 144.12 | 59.65 | 4 | 0.82 | 5 | 5 | 3 | 7.33 | 12.71 | 3.91 | 4.19 | - | |
| Reduction (%) | 0.23 | −10 | −10 | - | 0.06 | 0.13 | - | 0.11 | - | - | - | - | - | - | - | ||
| Combined Drought and Heat Stress (DSHS) | |||||||||||||||||
| Env | 1 | 613,509,9.7* | 1.9 | 0.1 | 2.5 | 155177.3** | 58356.3** | 38.9** | 2.3* | 0.1 | 5.7* | 23.0** | 0.1* | 0 | 0 | 0.1* | - |
| Rep (Env) | 2 | 843,164.7 | 14.4* | 17.2* | 0.3 | 522.3* | 21.3 | 1.4 | 0.1 | 0.5 | 2.9* | 0.4 | 0 | 0 | 0 | 0.1* | - |
| Block (Env × Rep) | 28 | 1,522,778.5* | 7.7* | 13.0** | 2.8* | 913.3** | 263.3** | 2.0** | 0.3 | 0.7 | 2.8** | 0.9 | 0 | 0.0* | 0 | 0.0* | - |
| Genotype | 71 | 2,575,212.4** | 47.3** | 46.6** | 4.1** | 1010.2** | 719.7** | 1.6** | 0.2* | 1.8** | 3.5** | 2.1** | 0.0* | 0.0** | 0.0** | 0.0* | - |
| Env × Genotype | 71 | 4,923,88.2 | 2.3 | 3.1 | 0.7 | 254.1* | 113.4** | 0.6 | 0.2 | 0.3 | 0.8 | 0.6 | 0 | 0 | 0 | 0 | - |
| Error | 114 | 595,224.5 | 3.2 | 4.7 | 1.7 | 161.7 | 46.8 | 0.7 | 0.2 | 0.5 | 0.8 | 0.6 | 0 | 0 | 0 | 0 | - |
| Repeatability | 0.79 | 0.95 | 0.93 | 0.66 | 0.77 | 0.86 | 0.62 | 0.25 | 0.78 | 0.77 | 0.69 | 0.57 | 0.62 | 0.64 | 0.55 | - | |
| Grand Mean | 1258.51 | 56 | 58 | 3 | 142.68 | 58.59 | 4 | 0.68 | 5 | 5 | 4 | 9.75 | 15.71 | 9.94 | 11.97 | - | |
| Reduction (%) | 0.58 | −9 | −8 | - | 0.07 | 0.15 | - | 0.26 | - | - | - | - | - | - | - | - | |
| Trait | NS vs. DS | NS vs. HS | NS vs. DSHS | HS vs. DS | DS vs. DSHS | HS vs. DSHS |
|---|---|---|---|---|---|---|
| Grain yield | 0.66*** | 0.75*** | 0.62*** | 0.48** | 0.60*** | 0.37*** |
| Anthesis days | 0.90*** | −0.05 | 0.87*** | −0.04 | 0.82*** | −0.13 |
| Silking | 0.80*** | −0.07 | 0.84*** | −0.08 | 0.73*** | −0.08 |
| Anthesis-Silking Interval | 0.23* | −0.09 | 0.42*** | −0.13 | 0.13 | −0.28* |
| Plant height | 0.48*** | 0.30* | 0.64*** | 0.18 | 0.29* | 0.23* |
| Ear height | 0.60*** | 0.21 | 0.77*** | 0.24* | 0.14 | 0.55*** |
| Ears per plant | 0.37** | 0.11 | 0.06 | 0.03 | 0.22 | −0.07 |
| Ear aspect | 0.54*** | 0.32** | 0.33** | 0.19 | 0.52*** | 0.05 |
| Plant aspect | 0.46*** | 0.14 | 0.63*** | 0.18 | 0.46*** | 0.04 |
| Stay green | - | - | - | 0.08 | 0.38*** | 0.03 |
| Leaf firing | - | - | - | - | - | −0.20 |
| Tassel blast | - | - | - | - | - | −0.10 |
| Trait | NS | DS | HS | DSHS |
|---|---|---|---|---|
| Days to anthesis | ||||
| Grain yield | 0.34 | −0.71 | −0.42 | −0.23 |
| Silking days | 0.95 | 0.99 | 0.98 | 0.97 |
| Anthesis-silking interval | 0.42 | 0.12 | 0.51 | 0.20 |
| Husk cover | −0.48 | −0.71 | −0.92 | −0.63 |
| Plant height | 0.58 | 0.80 | 0.91 | 0.79 |
| Ear height | 0.65 | 0.87 | 0.98 | 0.87 |
| Plant aspect | −0.31 | −0.76 | −0.87 | −0.62 |
| Ear aspect | −0.32 | −0.83 | −0.41 | −0.12 |
| Ears per plant | 0.06 | −0.89 | 0.19 | −0.34 |
| Stay green | - | −0.49 | −0.88 | −0.49 |
| Leaf firing | - | - | −0.79 | −0.26 |
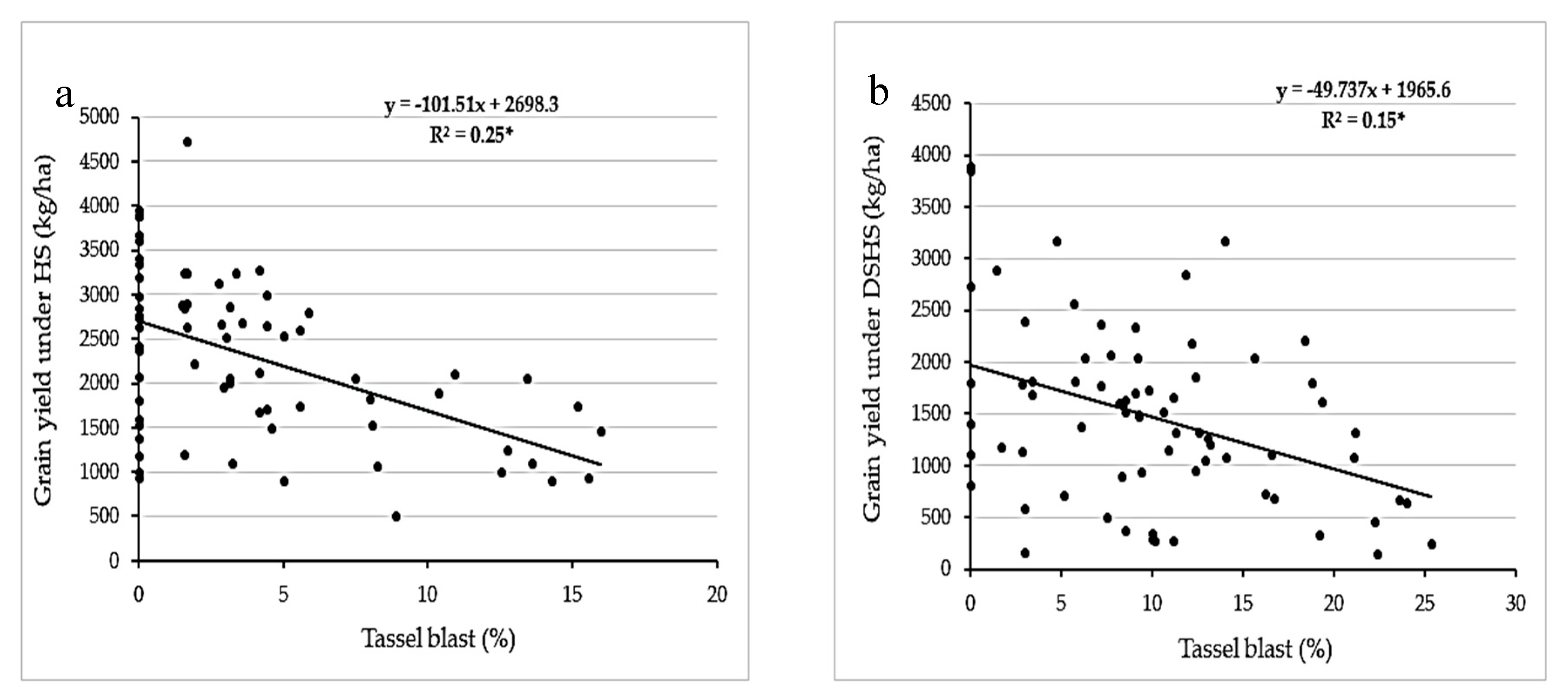
| Tassel blast | - | - | −0.98 | −0.12 |
| Days to Silking | ||||
| Grain yield | 0.14 | -0.65 | -0.31 | -0.14 |
| Anthesis-silking interval | 0.68 | 0.25 | 0.34 | 0.01 |
| Husk cover | −0.24 | −0.64 | −0.82 | −0.60 |
| Plant height | 0.35 | 0.80 | 0.87 | 0.81 |
| Ear height | 0.40 | 0.86 | 0.95 | 0.85 |
| Plant aspect | −0.06 | −0.71 | −0.81 | −0.59 |
| Ear aspect | −0.09 | −0.78 | −0.26 | −0.04 |
| Ears per plant | −0.33 | −0.77 | 0.08 | −0.53 |
| Stay green | −0.28 | - | −0.76 | −0.45 |
| Leaf firing | - | - | −0.79 | −0.20 |
| Tassel blast | - | - | −0.98 | - |
| Drought Stress (DS) | Heat Stress (HS) | Combined Drought and Heat Stress (DSHS) | ||||||
|---|---|---|---|---|---|---|---|---|
| Accession | GY (kg/ha) | BI | Accession | GY (kg/ha) | BI | Accession | GY (kg/ha) | BI |
| Check 3 | 3863 | 13.4 | Check 5 | 4723 | 14.5 | Check 4 | 3899 | 10.9 |
| TZm-1440 | 3287 | 11.4 | TZm-1167 | 3895 | 12.3 | TZm-1486 | 3167 | 8.2 |
| TZm-1163 | 3487 | 11.3 | TZm-1157 | 3614 | 9.1 | TZm-1162 | 3174 | 7.7 |
| TZm-1500 | 3086 | 11.1 | TZm-1178 | 3896 | 8.0 | TZm-1472 | 2334 | 4.8 |
| TZm-1162 | 3256 | 10.5 | TZm-1472 | 3238 | 7.8 | TZm-1171 | 2070 | 4.7 |
| TZm-1486 | 2815 | 9.2 | TZm-1163 | 3673 | 6.4 | TZm-1440 | 2373 | 4.5 |
| TZm-1160 | 2215 | 7.4 | TZm-1158 | 3247 | 6.3 | TZm-1470 | 2391 | 4.2 |
| TZm-1174 | 1886 | 6.5 | TZm-1352 | 2524 | 6.1 | TZm-1160 | 2187 | 3.9 |
| TZm-1349 | 1932 | 5.1 | TZm-1162 | 3137 | 4.7 | TZm-1481 | 2039 | 3.8 |
| TZm-1496 | 1685 | 4.9 | TZm-1179 | 3956 | 4.5 | TZm-1508 | 1797 | 3.1 |
| TZm-1449 | 2404 | 4.9 | TZm-1508 | 3350 | 4.2 | TZm-1483 | 2042 | 2.6 |
| TZm-1508 | 2521 | 4.5 | TZm-1329 | 3415 | 3.9 | TZm-1485 | 1735 | 2.5 |
| TZm-1472 | 2015 | 4.3 | TZm-1443 | 2984 | 3.9 | TZm-1167 | 1700 | 2.5 |
| TZm-1159 | 2026 | 4.3 | TZm-1561 | 3277 | 3.9 | TZm-1496 | 1802 | 2.4 |
| TZm-1511 | 1861 | 4.1 | TZm-1511 | 2793 | 3.9 | TZm-1506 | 2210 | 2.2 |
| TZm-1167 | 1926 | 3.7 | TZm-1454 | 3194 | 3.4 | TZm-1448 | 1853 | 2.0 |
| TZm-1169 | 530 | −8.8 | TZm-1497 | 907 | −9.36 | TZm-1510 | 336 | −6.8 |
| TZm-1493 | 571 | −9.0 | TZm-1493 | 907 | −9.37 | TZm-1509 | 281 | −7.3 |
| GH-4863 | 430 | −9.6 | TZm-1177 | 936 | −9.41 | TZm-1176 | 366 | −7.3 |
| TZm-1165 | 501 | −10.4 | TZm-1170 | 932 | −12.08 | TZm-1480 | 467 | −8.6 |
| TZm-1510 | 543 | −10.6 | TZm-1498 | 501 | −12.29 | TZm-1173 | 152 | −11.3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nelimor, C.; Badu-Apraku, B.; Tetteh, A.Y.; Garcia-Oliveira, A.L.; N’guetta, A.S.-P. Assessing the Potential of Extra-Early Maturing Landraces for Improving Tolerance to Drought, Heat, and Both Combined Stresses in Maize. Agronomy 2020, 10, 318. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10030318
Nelimor C, Badu-Apraku B, Tetteh AY, Garcia-Oliveira AL, N’guetta AS-P. Assessing the Potential of Extra-Early Maturing Landraces for Improving Tolerance to Drought, Heat, and Both Combined Stresses in Maize. Agronomy. 2020; 10(3):318. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10030318
Chicago/Turabian StyleNelimor, Charles, Baffour Badu-Apraku, Antonia Yarney Tetteh, Ana Luísa Garcia-Oliveira, and Assanvo Simon-Pierre N’guetta. 2020. "Assessing the Potential of Extra-Early Maturing Landraces for Improving Tolerance to Drought, Heat, and Both Combined Stresses in Maize" Agronomy 10, no. 3: 318. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10030318