
Defensive Impact of Foliar Applied Potassium Nitrate on Growth Linked with Improved Physiological and Antioxidative Activities in Sunflower (Helianthus annuus L.) Hybrids Grown under Salinity Stress


 , ,
, ,  , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Details
2.2. Biochemical, Mineral, and Antioxidant Enzyme Activity Measurement
2.3. Growth Attributes
2.4. Statistical Analysis
3. Results
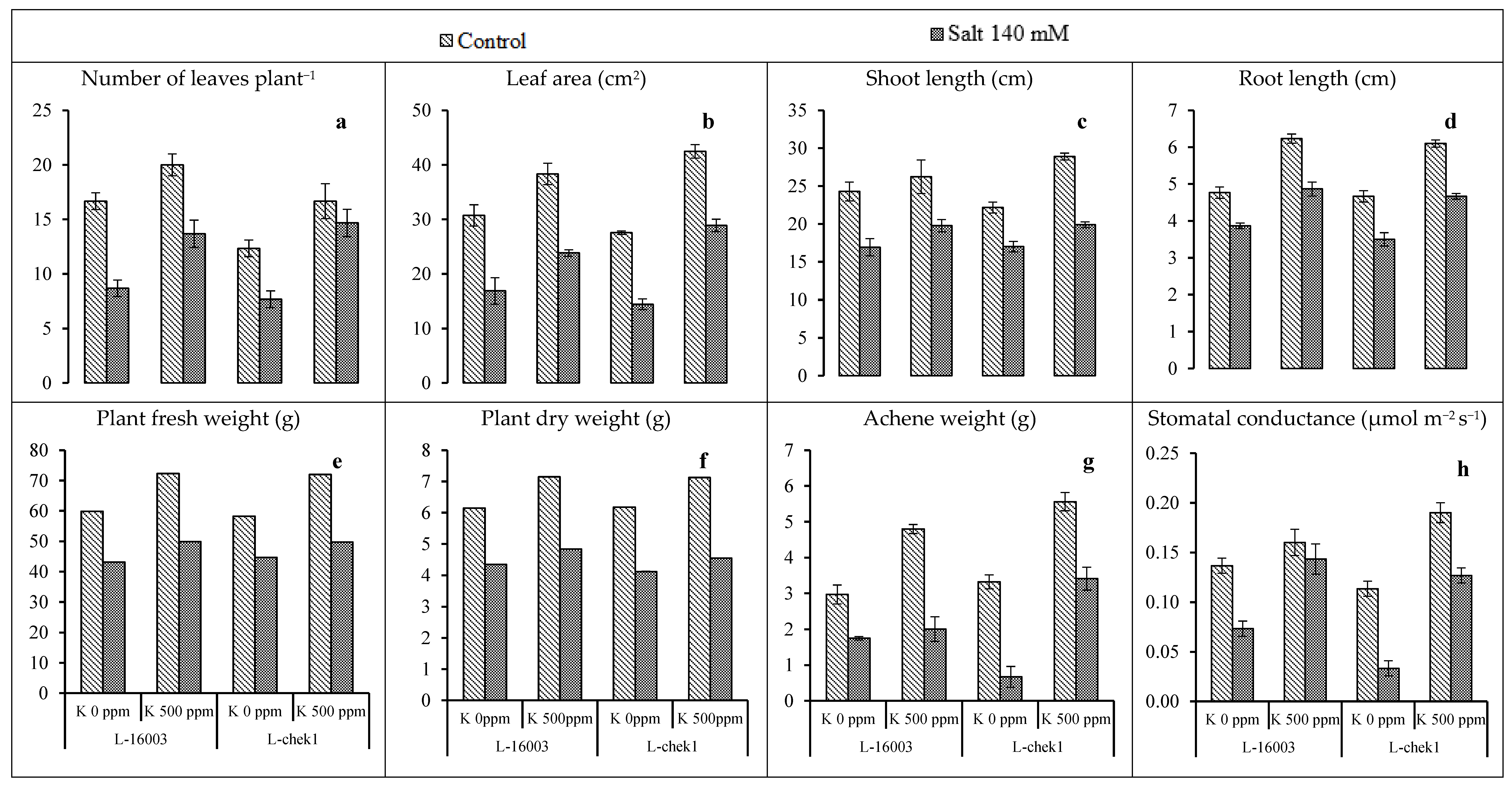
3.1. Growth Attributes
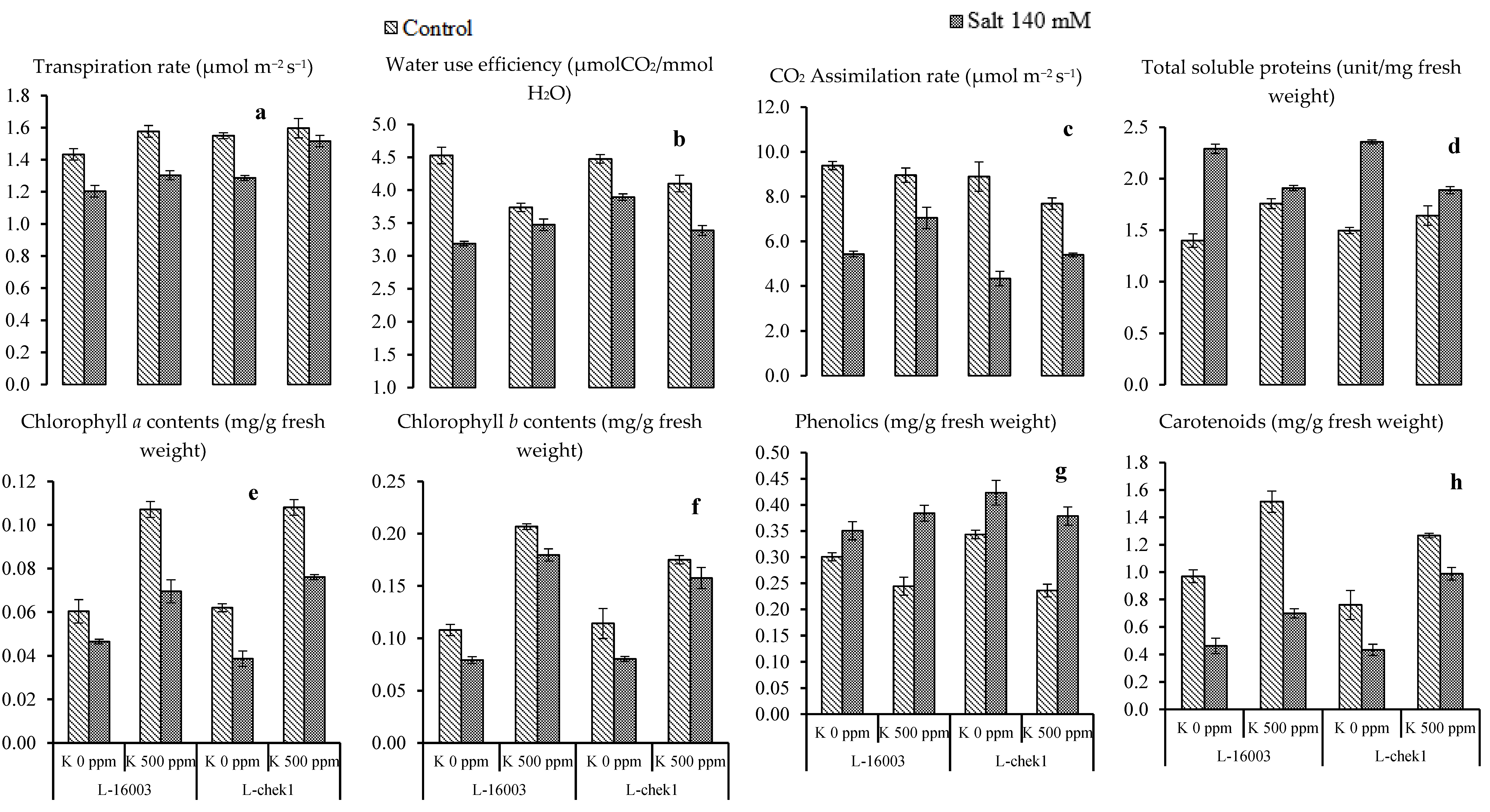
3.2. Biochemical and Physiological Attributes
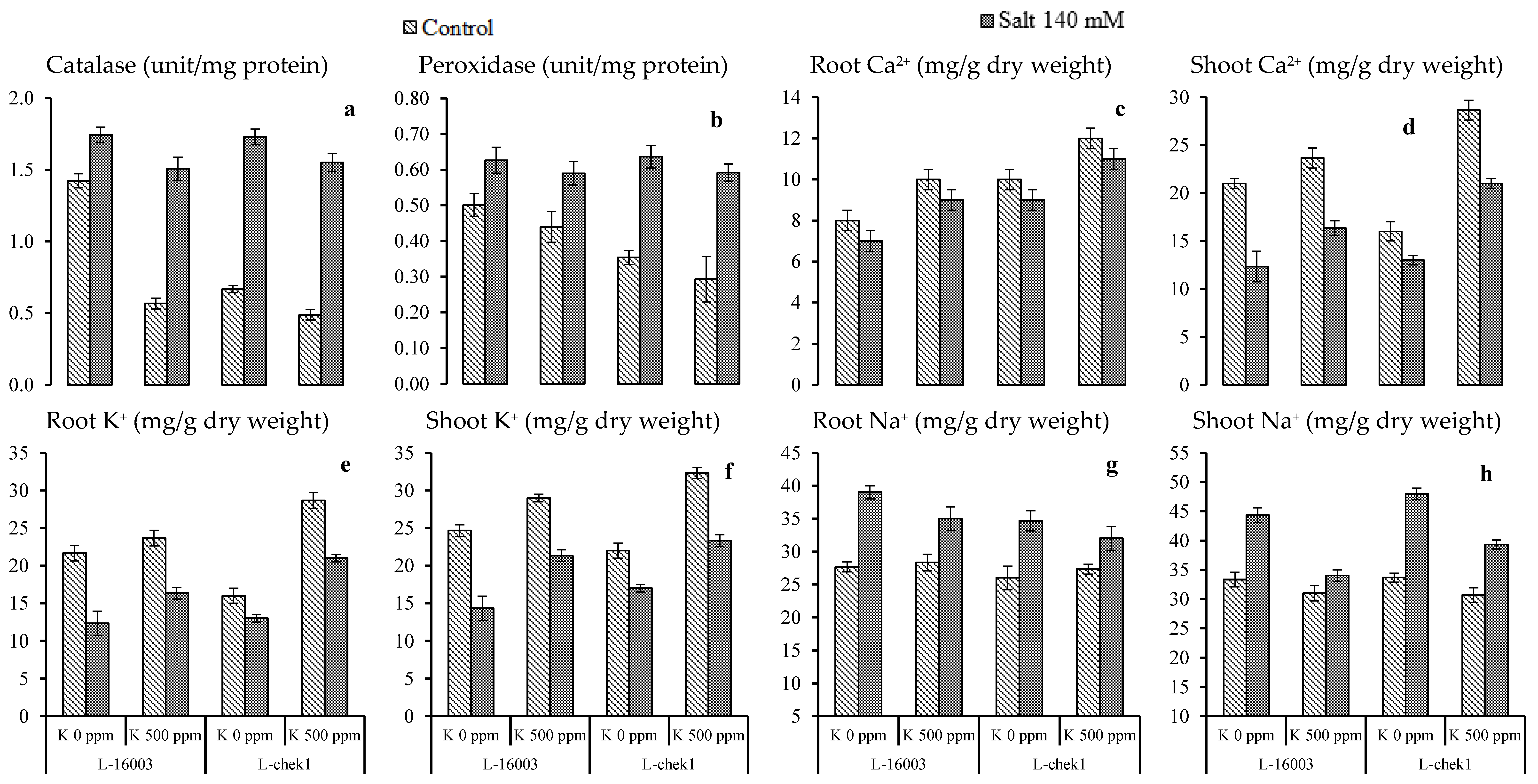
3.3. Antioxidant and Enzymatic Activities
3.4. Mineral Homeostasis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paul, P.L.C.; Bell, R.W.; Barrett-Lennard, E.G. Variation in the yield of sunflower (Helianthus annuus L.) due to differing tillage systems is associated with variation in solute potential of the soil solution in a salt-affected coastal region of the Ganges Delta. Soil Tillage Res. 2020, 197, 104489. [Google Scholar] [CrossRef]
- Hafeez, M.B.; Raza, A.; Zahra, N.; Shaukat, K.; Akram, M.Z.; Iqbal, S.; Basra, S.M.A. Gene regulation in halophytes in conferring salt tolerance. In Handbook of Bioremediation; Elsevier: Amsterdam, The Netherlands, 2021; pp. 341–370. [Google Scholar]
- Taher, M.; Beyaz, R.; Javani, M. Morphological and biochemical changes in response to salinity in sunflower (Helianthus annus L.) cultivars. Ital. J. Agron. 2018, 11, 141–147. [Google Scholar] [CrossRef]
- Alsaeedi, A.; El-Ramady, H.; Alshaal, T.; El-Garawany, M.; Elhawat, N.; Al-Otaibi, A. Silica nanoparticles boost growth and productivity of cucumber under water deficit and salinity stresses by balancing nutrients uptake. Plant Physiol. Biochem. 2019, 139, 1–10. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. The Response of Salinity Stress Induced A. tricolor to Growth, Anatomy, Physiology, Non-Enzymatic and Enzymatic Antioxidants. Front. Plant Sci. 2020, 11, 559876. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Catalase, superoxide dismutase and ascorbate-glutathione cycle enzymes confer drought tolerance of Amaranthus tricolor. Sci. Rep. 2018, 8, 16496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, U.; Oba, S. Drought Stress Effects on Growth, ROS Markers, Compatible Solutes, Phenolics, Flavonoids, and Antioxidant Activity in Amaranthus tricolor. Appl. Biochem. Biotechnol. 2018, 186, 999–1016. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S. Salinity stress enhances color parameters, bioactive leaf pigments, vitamins, polyphenols, flavonoids and antioxidant activity in selected Amaranthus leafy vegetables. J. Sci. Food Agric. 2019, 99, 2275–2284. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Response of nutrients, minerals, antioxidant leaf pigments, vitamins, polyphenol, flavonoid and antioxidant activity in selected vegetable amaranth under four soil water content. Food Chem. 2018, 252, 72–83. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Drought stress enhances nutritional and bioactive compounds, phenolic acids and antioxidant capacity of Amaranthus leafy vegetable. BMC Plant Biol. 2018, 18, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, U.; Islam, M.T.; Oba, S. Salinity stress accelerates nutrients, dietary fiber, minerals, phytochemicals and antioxidant activity in Amaranthus tricolor leaves. PLoS ONE 2018, 13, e0206388. [Google Scholar] [CrossRef] [Green Version]
- Hurtado, A.C.; Chiconato, D.A.; de Mello, P.R. Silicon attenuates sodium toxicity by improving nutritional efficiency in sorghum and sunflower plants. Plant. Physiol. Biochem. 2019, 142, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, A.A.; Abd Elhamid, E.M.; Sadak, M.S. Comparative study for the effect of arginine and sodium nitroprusside on sunflower plants grown under salinity stress conditions. Bull. Natl. Res. Centre 2019, 43, 1–18. [Google Scholar] [CrossRef]
- Jan, A.U.; Hadi, F.; Akbar, F. Role of potassium, zinc and gibberellic acid in increasing drought stress tolerance in sunflower (helianthus annuus l.). Pak. J. Bot. 2019, 51, 809–815. [Google Scholar] [CrossRef]
- Raza, M.A.S.; Saleem, M.F.; Khan, I.H. Amelioration in growth and physiological efficiency of sunflower (Helianthus annuus L.) under drought by potassium application. Commun. Soil Sci. Plant Anal. 2018, 49, 2291–2300. [Google Scholar] [CrossRef]
- Zaman, B.U.; Ali, A.; Arshadullah, M. Growth Response of Sunflower to Potassium Sulphate Application in Saline-sodic Soil. Asian J. Res. Agric. For. 2018, 1, 1–7. [Google Scholar]
- Wada, S.; Takagi, D.; Miyake, C. Responses of the photosynthetic electron transport reactions stimulate the oxidation of the reaction center chlorophyll of photosystem I, p700, under drought and high temperatures in rice. Int. J. Mol. Sci. 2019, 20, 2068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shehzad, M.A.; Fahim, N.; Fiaz, A. Protective effect of potassium and chitosan supply on growth, physiological processes and antioxidative machinery in sunflower (Helianthus annuus L.) under drought stress. Ecotox. Environ. Saf. 2020, 187, 1–11. [Google Scholar] [CrossRef]
- Cui, Y.N.; Xia, Z.R.; Ma, Q. The synergistic effects of sodium and potassium on the xerophyte Apocynum venetum in response to drought stress. Plant Physiol. Biochem. 2019, 135, 489–498. [Google Scholar] [CrossRef]
- Zamani, S.; Naderi, M.R.; Soleymani, A. Sunflower (Helianthus annuus L.) biochemical properties and seed components affected by potassium fertilization under drought conditions. Ecotox. Environ. Saf. 2020, 190, 11–17. [Google Scholar] [CrossRef]
- F.A.O. Statistical Year Book; Food and Agriculture Organization of United Nations: Rome, Italy, 2013. [Google Scholar]
- Aishwarya, S.; Anisha, V. Nutritional composition of sunflower seeds flour and nutritive value of products prepared by incorporating sunflower seeds flour. Int. J. Pharm. Res. Allied Sci. 2014, 3, 45–49. [Google Scholar]
- Economic Survey of Pakistan, 2020–2021. Available online: https://www.finance.gov.pk/survey/chapters_21/02-Agriculture.pdf (accessed on 11 September 2021).
- Wu, H.; Zhang, X.; Giraldo, J.P. It is not all about sodium: Revealing tissue specificity and signalling roles of potassium in plant responses to salt stress. Plant Soil. 2018, 431, 1–17. [Google Scholar] [CrossRef]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Rehman, A.; Hassan, F.; Qamar, R.; Rehman, A.U. Application of plant growth promoters on sugarcane (Saccharum officinarum L.) budchip under subtropical conditions. Asian J. Agric. Biol. 2021, 2, 202003202. [Google Scholar] [CrossRef]
- Makawita, G.I.P.S.; Wickramasinghe, I.; Wijesekara, I. Using brown seaweed as a biofertilizer in the crop management industry and assessing the nutrient upliftment of crops. Asian J. Agric. Biol. 2021. [Google Scholar] [CrossRef]
- Hussain, M.U.; Saleem, M.F.; Hafeez, M.B.; Khan, S.; Hussain, S.; Ahmad, N.; Ramzan, Y.; Nadeem, M. Impact of soil applied humic acid, zinc and boron supplementation on the growth, yield and zinc translocation in winter wheat. Asian J. Agric. Biol. 2021. [Google Scholar] [CrossRef]
- Anwar, Z.; Basharat, Z.; Hafeez, M.B.; Khan, S.; Zahra, N.; Rafique, Z.; Maqsood, M. Biofortification of Maize with Zinc and Iron not only Enhances Crop Growth but also Improves Grain Quality. Asian J. Agric. Biol. 2021. [Google Scholar] [CrossRef]
- Khan, S.; Basit, A.; Hafeez, M.B.; Irshad, S.; Bashir, S.; Bashir, S.; Maqbool, M.M.; Saddiq, M.S.; Hasnain, Z.; Aljuaid, B.S.; et al. Moringa leaf extract improves biochemical attributes, yield and grain quality of rice (Oryza sativa L.) under drought stress. PLoS ONE 2021, 16, e0254452. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Basra, S.M.A.; Nawaz, M.; Hussain, I.; Foidl, N. Combined application of moringa leaf extract and chemical growth-promoters enhances the plant growth and productivity of wheat crop (Triticum aestivum L.). S. Afri. J. Bot. 2020, 129, 74–81. [Google Scholar] [CrossRef]
- Khan, S.; Basra, S.M.A.; Afzal, I.; Nawaz, M.; Rehman, H.U. Growth promoting potential of fresh and stored Moringa oleifera leaf extracts in improving seedling vigor, growth and productivity of wheat crop. Environ. Sci. Pollut. Res. 2017, 24, 27601–27612. [Google Scholar] [CrossRef] [PubMed]
- Tamme, A.A.; Kerr, K.L.; Donovan, L.A. Vigour/tolerance trade-off in cultivated sunflower (Helianthus annuus) response to salinity stress is linked to leaf elemental composition. J. Agron. Crop Sci. 2019, 5, 508–518. [Google Scholar] [CrossRef]
- Nachabe, M.H. Refining the definition of field capacity in the literature. J. Irrig. Drain. Eng. 1998, 124, 230–232. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzyme polyphenoloxides in isolated chloroplast in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Julkkenon, R. Phenolics constituents in the leaves of northern willows methods for analysis of certain phenolics. Agric. Food Chem. 1985, 33, 213–217. [Google Scholar]
- USDA Laboratory Staff. Diagnosis and improvement of saline and alkali soils. In Superintendent of Documents; Richards, L.A., Ed.; U. S. Government Printing Office Washington: Washington, DC, USA, 1954. [Google Scholar]
- Chance, B.; Maehly, A.C. Assay of catalaseand peroxidase. Methods Enzymol. 1955, 2, 764–775. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for quantitation or microgram quantities of protein nitiling the principle protein dychinding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Long, S.P.; Farage, P.K.; Garcia, R.L. Measurement of leaf and canopy photosynthetic CO2 exchange in the field. J. Exp. Bot. 1996, 47, 1629–1642. [Google Scholar] [CrossRef] [Green Version]
- Steel, R.G.D.; Torrie, J.H.; Dickey, D.A. Principles and Procedures of Statistics: A Biometrical Approach, 3rd ed.; Mc Graw Hill Book Co. Inc.: New York, NY, USA, 1997; Volume 3, pp. 336–352. [Google Scholar]
- Hussain, R.A.; Ashraf, M.Y.; Ahmad, R. Foliar nitrogen and potassium applications improve photosynthetic activities and water relations in sunflower under moisture deficit condition. Pak. J. Bot. 2016, 48, 1805–1811. [Google Scholar]
- Latef, A.A.A.; Chaoxing, H. Does the inoculation with Glomus mosseae improves salt tolerance in pepper plants? J. Plant Growth Regul. 2014, 33, 644–653. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Fujita, M.; Tran, L.S.P. Nitric oxide mediates hydrogen peroxideand salicylic acid induced salt tolerance in rice (Oryza sativa L.) seedlings. Plant Growth Regul. 2015, 77, 265–277. [Google Scholar] [CrossRef]
- Kosová, K.; Vítámvás, P.; Prášil, I.T. Plant proteome changes under abiotic stress—contribution of proteomics studies to understanding plant stress response. J. Proteom. 2011, 74, 1301–1322. [Google Scholar] [CrossRef]
- Farooq, O.; Ali, M.; Sarwar, N.; Rehman, A.; Iqbal, M.M.; Naz, T.; Asghar, M.; Ehsan, F.; Nasir, M.; Hussain, Q.M.; et al. Foliar applied brassica water extract improves the seedling development of wheat and chickpea. Asian J. Agric. Biol. 2021. [Google Scholar] [CrossRef]
- Raza, M.A.S.; Saleem, M.F.; Shah, G.M. Exogenous application of glycinebetaine and potassium for improving water relations and grain yield of wheat under drought. J. Soil Sci. Plant Nutr. 2014, 14, 348–364. [Google Scholar] [CrossRef] [Green Version]
- Waraich, E.A.; Ahmad, R.; Ashraf, M.Y. Improving agricultural water use efficiency by nutrients management. Acta. Agric. Scand. Soil. Plant. Sci. 2011, 61, 291–304. [Google Scholar]
- Ashfaq, A.; Hussain, N.; Athar, M. Role of potassium fertilizers in plant growth, crop yield and quality fiber production of cotton. Fuuast. J. Biol. 2015, 5, 27–35. [Google Scholar]
- Nazar, R.; Iqbal, N.; Syeed, S. Salicylic acid alleviates decreases in photosynthesis under salt stress by enhancing nitrogen and sulfur assimilation and antioxidant metabolism differentially in two mungbean cultivars. J. Plant Physiol. 2011, 168, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Netondo, G.W.; Onyango, J.C.; Beck, E. Sorghum and salinity: Response of growth, water relations and ion accumulation to NaCl salinity. Crop Sci. 2004, 44, 797–805. [Google Scholar] [CrossRef]
- Tabaxi, I.; Ζisi, C.; Karydogianni, S.; Folina, A.E.; Kakabouki, I.; Kalivas, A.; Bilalis, D. Effect of organic fertilization on quality and yield of oriental tobacco (Nicotiana tabacum L.) under Mediterranean conditions. Asian J. Agric. Biol. 2021. [Google Scholar] [CrossRef]
- Safaa, R.; Magdi, T.; Abde, H. Effect of Potassium Application on Wheat (Triticum aestivum L.) Cultivars grown under salinity stress. World Appl. Sci. J. 2013, 26, 840–850. [Google Scholar]
- Kabir, E.; Karimand, M.A.; Azad, M.A.K. Effect of Potassium on salinity tolerance of mungbean. J. Biol. Sci. 2004, 4, 103–110. [Google Scholar]
- Zahid, N.; Ahmed, M.J.; Tahir, M.M.; Maqbool, M.; Shah, S.Z.A.; Hussain, S.J.; Khaliq, A.; Rehmani, M.I.A. Integrated effect of urea and poultry manure on growth, yield and postharvest quality of cucumber (Cucumis sativus L.). Asian J. Agric. Biol. 2021. [Google Scholar] [CrossRef]
- Safdar, M.E.; Aslam, A.; Qamar, R.; Ali, A.; Javaid, M.M.; Hayyat, M.S.; Raza, A. Allelopathic effect of prickly chaff flower (Achyranthes aspera L.) used as a tool for managing noxious weeds. Asian J. Agric. Biol. 2021, 10. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Antioxidant constituents of three selected red and green color Amaranthus leafy vegetable. Sci. Rep. 2019, 9, 18233. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Augmentation of leaf color parameters, pigments, vitamins, phenolic acids, flavonoids and antioxidant activity in selected Amaranthus tricolor under salinity stress. Sci. Rep. 2018, 8, 12349. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Hossain, M.N.; Iqbal, M.A.; Oba, S. Bioactive Components and Radical Scavenging Activity in Selected Advance Lines of Salt-Tolerant Vegetable Amaranth. Front. Nutr. 2020, 7, 587257. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S. Nutraceuticals, phytochemicals, and radical quenching ability of selected drought tolerant advance lines of vegetable amaranth. BMC Plant Biol. 2020, 20, 564. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S. Polyphenol and flavonoid profiles and radical scavenging activity in leafy vegetable Amaranthus gangeticus. BMC Plant Biol. 2020, 20, 499. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Phenolic profiles and antioxidant activities in selected drought tolerant leafy vegetable amaranth. Sci. Rep. 2020, 10, 18287. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S. Color attributes, betacyanin, and carotenoid profiles, bioactive components, and radical quenching capacity in selected Amaranthus gangeticus leafy vegetables. Sci. Rep. 2021, 11, 11559. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Nutritional and bioactive constituents and scavenging capacity of radicals in Amaranthus hypochondriacus. Sci. Rep. 2020, 10, 19962. [Google Scholar] [CrossRef]
- Sarker, U.; Hossain, M.M.; Oba, S. Nutritional and antioxidant components and antioxidant capacity in green morph Amaranthus leafy vegetable. Sci. Rep. 2020, 10, 1336. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Nutrients, minerals, pigments, phytochemicals, and radical scavenging activity in Amaranthus blitum leafy vegetables. Sci. Rep. 2020, 10, 3868. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Protein, dietary fiber, minerals, antioxidant pigments and phytochemicals, and antioxidant activity in selected red morph Amaranthus leafy vegetable. PLoS ONE 2019, 14, e0222517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, U.; Oba, S. Leaf pigmentation, its profiles and radical scavenging activity in selected Amaranthus tricolor leafy vegetables. Sci. Rep. 2020, 10, 18617. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Islam, M.T.; Rabbani, M.G.; Oba, S. Antioxidant Leaf Pigments and Variability in Vegetable Amaranth. Genetika 2018, 50, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Nutraceuticals, antioxidant pigments, and phytochemicals in the leaves of Amaranthus spinosus and Amaranthus viridis weedy species. Sci. Rep. 2019, 9, 20413. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Islam, M.T.; Rabbani, M.G.; Oba, S. Variability in total antioxidant capacity, antioxidant leaf pigments and foliage yield of vegetable amaranth. J. Integ. Agric. 2018, 17, 1145–1153. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S.; Daramy, M.A. Nutrients, minerals, antioxidant pigments and phytochemicals, and antioxidant capacity of the leaves of stem amaranth. Sci. Rep. 2020, 10, 3892. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Islam, M.T.; Rabbani, M.G.; Oba, S. Phenotypic divergence in vegetable amaranth for total antioxidant capacity, antioxidant profile, dietary fiber, nutritional and agronomic traits. Acta Agric. Scand. Sect. B Soil Plant Sci. 2018, 68, 67–76. [Google Scholar] [CrossRef]
- Akram, A.N.; Ashraf, M.; Al-Qurainyc, F. Aminolevulinic acid-induced changes in somekey physiological attributes and activities of antioxidant enzymes in sunflower (Helianthus annuus L.) plants under saline regimes. Sci. Hortic. 2012, 142, 143–148. [Google Scholar] [CrossRef]
- Luo, X.; Dai, Y.; Zheng, C.; Yang, Y.; Chen, W.; Wang, Q.; Chandrasekaran, U.; Du, J.; Liu, W.; Shu, K. The ABI4-RbohD/VTC2 regulatory module promotes reactiveoxygen species (ROS) accumulation to decrease seed germination under salinity stress. New Phytol. 2021, 229, 950–962. [Google Scholar] [CrossRef]
- Santos, C.V. Regulation of chlorophyll biosynthesis and degradation by salt stress in sunflower leaves. Sci. Hortic. 2014, 103, 93–99. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| SOV | DF | No of Leaves | Leaf Area | Shoot Length | Root Length | Plant Fresh Weight | Plant Dry Weight | Achene Weight | Stomatal Conductance | Transpiration Rate | WUE | CO2 AR | TSP |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Salinity stress (S) | 1 | 165 ** | 1137 ** | 294 ** | 8.88 ** | 19.561 ** | 2.877 ** | 29.15 ** | 0.018 ** | 0.126 ** | 5.771 ** | 0.083 ** | 1.736 ** |
| Treatment (T) | 1 | 145 ** | 724 ** | 77.7 ** | 9.63 ** | 4.182 ** | 0.246 ** | 18.71 ** | 0.026 ** | 0.024 NS | 0.451 NS | 0.0001 NS | 0.045 NS |
| Hybrid (H) | 1 | 22.0 * | 4.86 NS | 0.202 NS | 0.24 NS | 0.0072 * | 0.015 NS | 0.782 NS | 0.001 NS | 0.151 ** | 0.622 NS | 14.56 ** | 0.0003 NS |
| S × T | 1 | 7.04 NS | 0.47 NS | 3.23 NS | 0.202 NS | 0.841 * | 0.049 NS | 0.435 NS | 0.002 NS | 0.028 NS | 1.456 ** | 10.73 ** | 0.688 ** |
| S × H | 1 | 22.0 * | 1.05 NS | 0.04 NS | 0.042 NS | 0.061 NS | 0.004 * | 0.234 NS | 0.002 NS | 0.003 NS | 0.002 NS | 0.199 NS | 0.002 NS |
| T × H | 1 | 3.37 NS | 82.2 ** | 8.82 NS | 0.007 NS | 0.0017 NS | 0.0006 NS | 3.161 ** | 0.003 * | 0.004 NS | 0.202 NS | 2.131 NS | 0.034 NS |
| S × T × H | 1 | 0.37 NS | 0.015 NS | 8.40 NS | 0.015 NS | 0.0198 NS | 0.00022 NS | 1.607 * | 0.001 NS | 0.001 NS | 0.261 NS | 0.609 NS | 0.006 NS |
| SOV | DF | Chl a | Chl b | Phenolics | Carotenoids | Catalase | Peroxidase | Root Ca2+ | Shoot Ca2+ | Root K+ | Shoot K+ | Root Na+ | Shoot Na+ |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Salinity stress (S) | 1 | 0.004 ** | 0.004 ** | 0.064 ** | 1.391 ** | 4.313 ** | 0.276 ** | 3.375 NS | 266.7 ** | 280.2 ** | 384 ** | 368 ** | 513 ** |
| Treatment (T) | 1 | 0.009 ** | 0.043 ** | 0.012 ** | 1.271 ** | 0.792 ** | 0.015 NS | 22.04 * | 280.7 ** | 266.7 ** | 294 ** | 8.17 NS | 222 ** |
| Hybrid (H) | 1 | 0.001 NS | 0.001 NS | 0.004 NS | 0.014 NS | 0.243 ** | 0.029 NS | 5.042 NS | 10.67 NS | 8.167 NS | 10.67 NS | 37.5 * | 30.38 * |
| S × T | 1 | 0.004 * | 0.001 NS | 0.008 ** | 0.026 NS | 0.143 ** | 0.001 NS | 3.375 NS | 4.167 NS | 2.667 NS | 0.667 NS | 28.17 NS | 70.04 ** |
| S × H | 1 | 0.006 NS | 0.007 NS | 0.001 NS | 0.192 ** | 0.282 ** | 0.035 NS | 0.375 NS | 10.67 NS | 13.5 NS | 6.01 NS | 8.167 NS | 30.38 * |
| T × H | 1 | 0.007 NS | 0.002 * | 0.0062 * | 0.029 NS | 0.202 ** | 0.001 NS | 0.042 NS | 73.5 ** | 80.67 ** | 10.67 NS | 1.5 NS | 0.375 NS |
| S × T × H | 1 | 0.008 NS | 0.001 NS | 0.0012 NS | 0.047 NS | 0.145 ** | 0.002 NS | 0.042 NS | 13.5 NS | 16.67 NS | 16.67 * | 0.167 NS | 2.042 NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aslam, A.; Khan, S.; Ibrar, D.; Irshad, S.; Bakhsh, A.; Gardezi, S.T.R.; Ali, M.; Hasnain, Z.; Al-Hashimi, A.; Noor, M.A.; et al. Defensive Impact of Foliar Applied Potassium Nitrate on Growth Linked with Improved Physiological and Antioxidative Activities in Sunflower (Helianthus annuus L.) Hybrids Grown under Salinity Stress. Agronomy 2021, 11, 2076. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11102076
Aslam A, Khan S, Ibrar D, Irshad S, Bakhsh A, Gardezi STR, Ali M, Hasnain Z, Al-Hashimi A, Noor MA, et al. Defensive Impact of Foliar Applied Potassium Nitrate on Growth Linked with Improved Physiological and Antioxidative Activities in Sunflower (Helianthus annuus L.) Hybrids Grown under Salinity Stress. Agronomy. 2021; 11(10):2076. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11102076
Chicago/Turabian StyleAslam, Anisa, Shahbaz Khan, Danish Ibrar, Sohail Irshad, Ali Bakhsh, Syed Tahir Raza Gardezi, Madad Ali, Zuhair Hasnain, Abdulrahman Al-Hashimi, Mehmood Ali Noor, and et al. 2021. "Defensive Impact of Foliar Applied Potassium Nitrate on Growth Linked with Improved Physiological and Antioxidative Activities in Sunflower (Helianthus annuus L.) Hybrids Grown under Salinity Stress" Agronomy 11, no. 10: 2076. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11102076