
Yield Stability and Genotype Environment Interaction of Water Deficit Stress Tolerant Mung Bean (Vigna radiata L. Wilczak) Genotypes of Bangladesh

 ,
,  ,
,  , ,
, ,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
Statistical Analysis
3. Results and Discussion
3.1. Chemical Properties of the Initial Soil of Five Studied Locations
3.2. Variation in Phenology as Influenced by Different Environments
3.3. Environmental Impact on Yield Attributing Traits
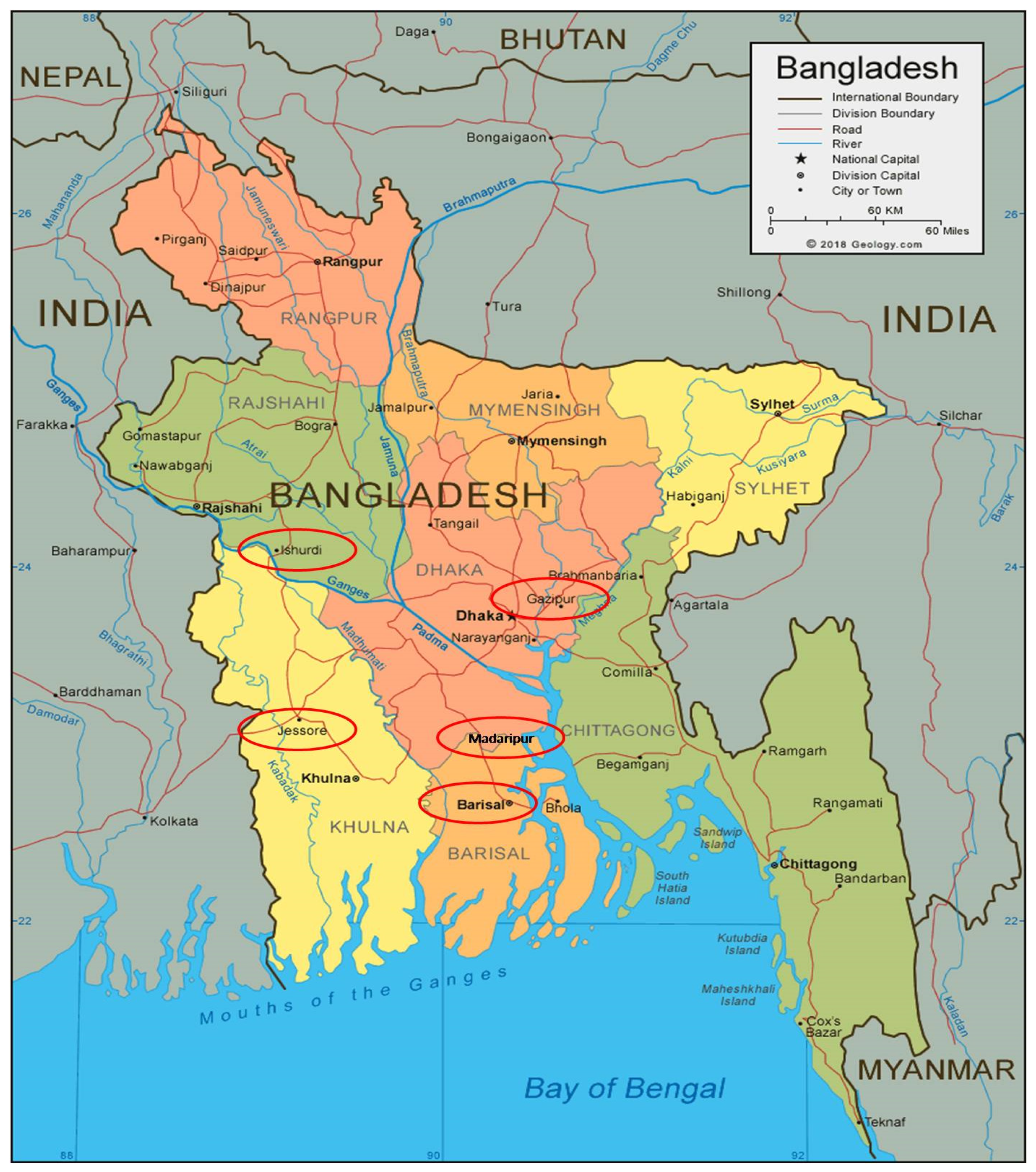
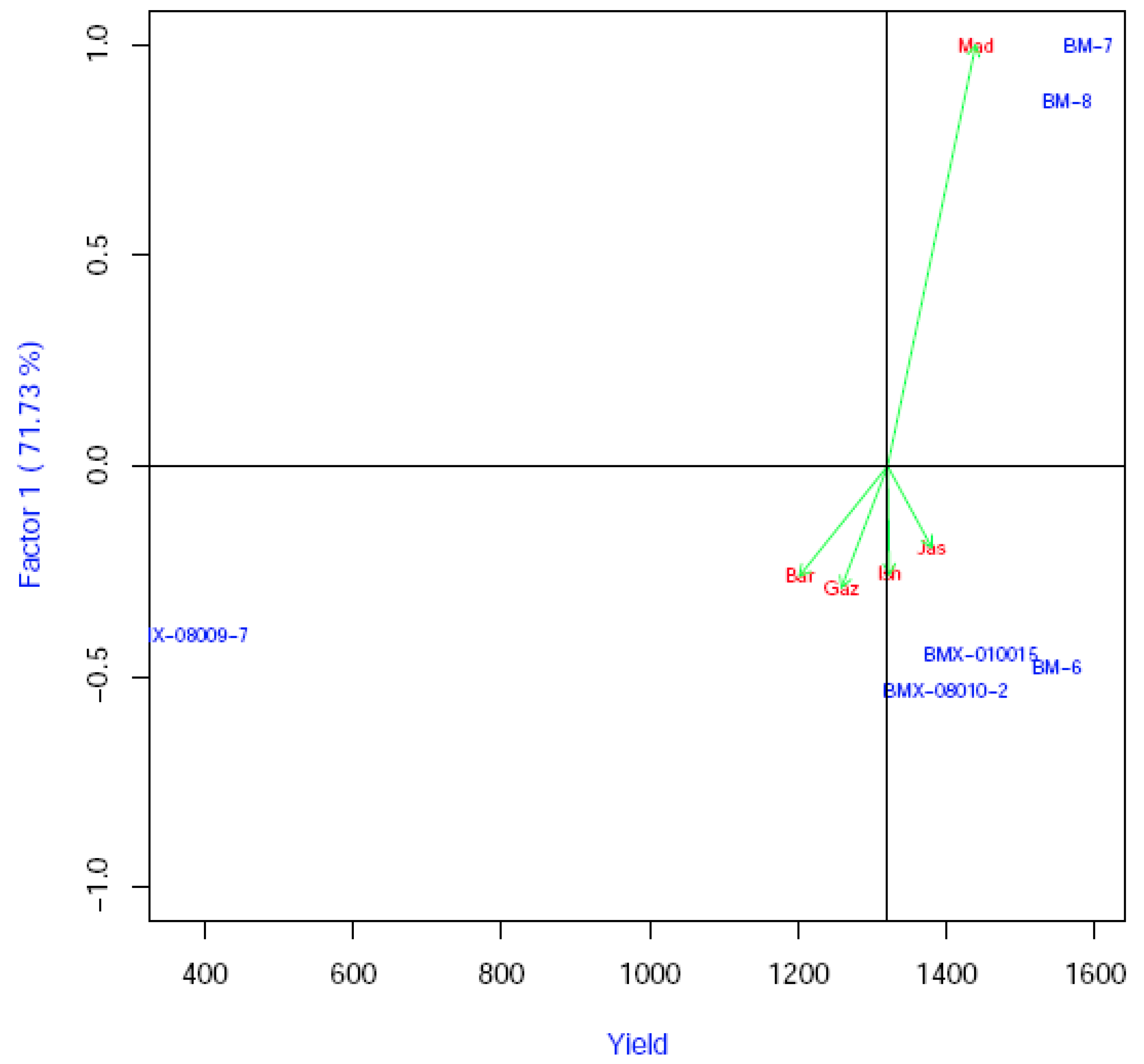
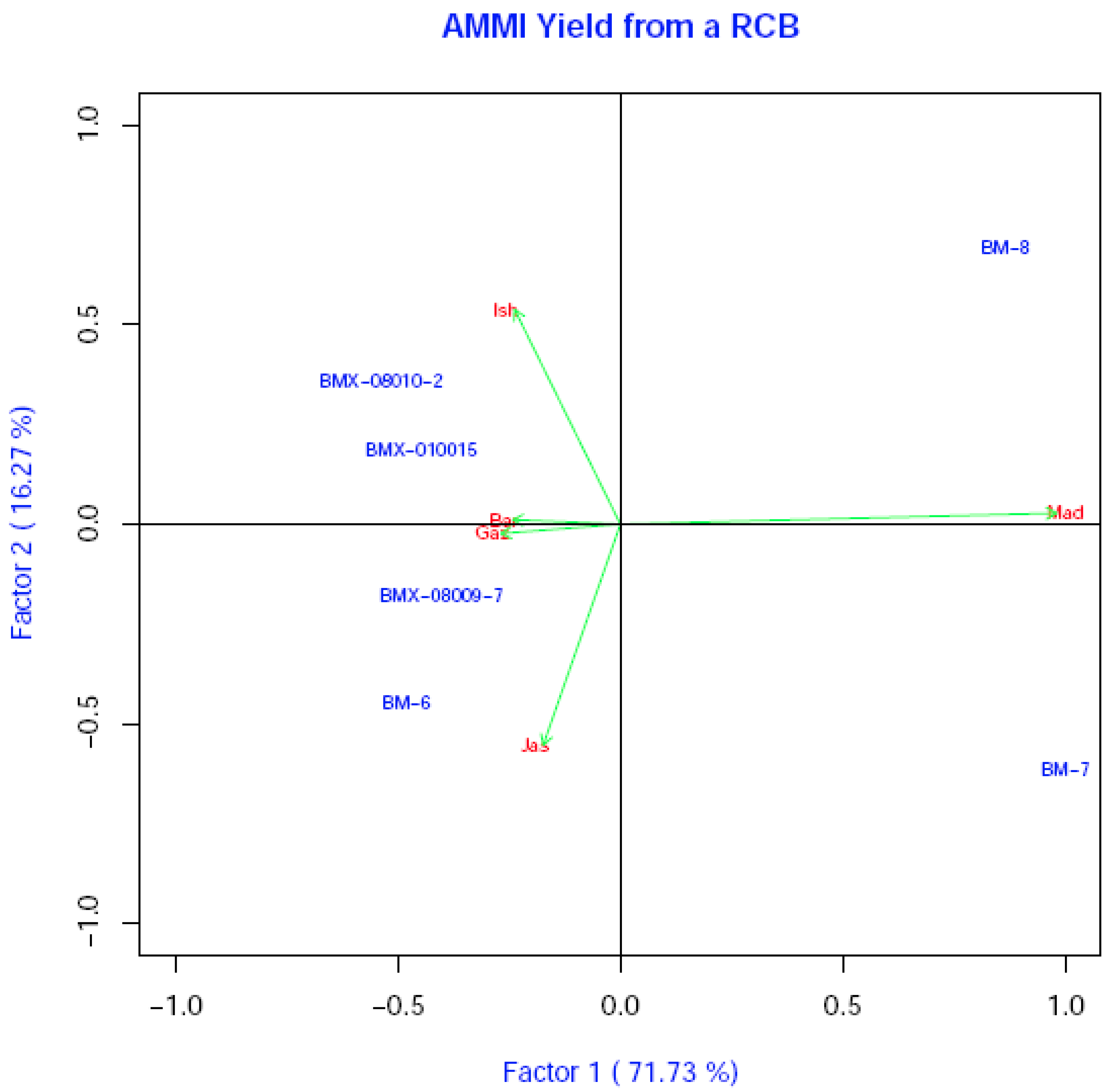
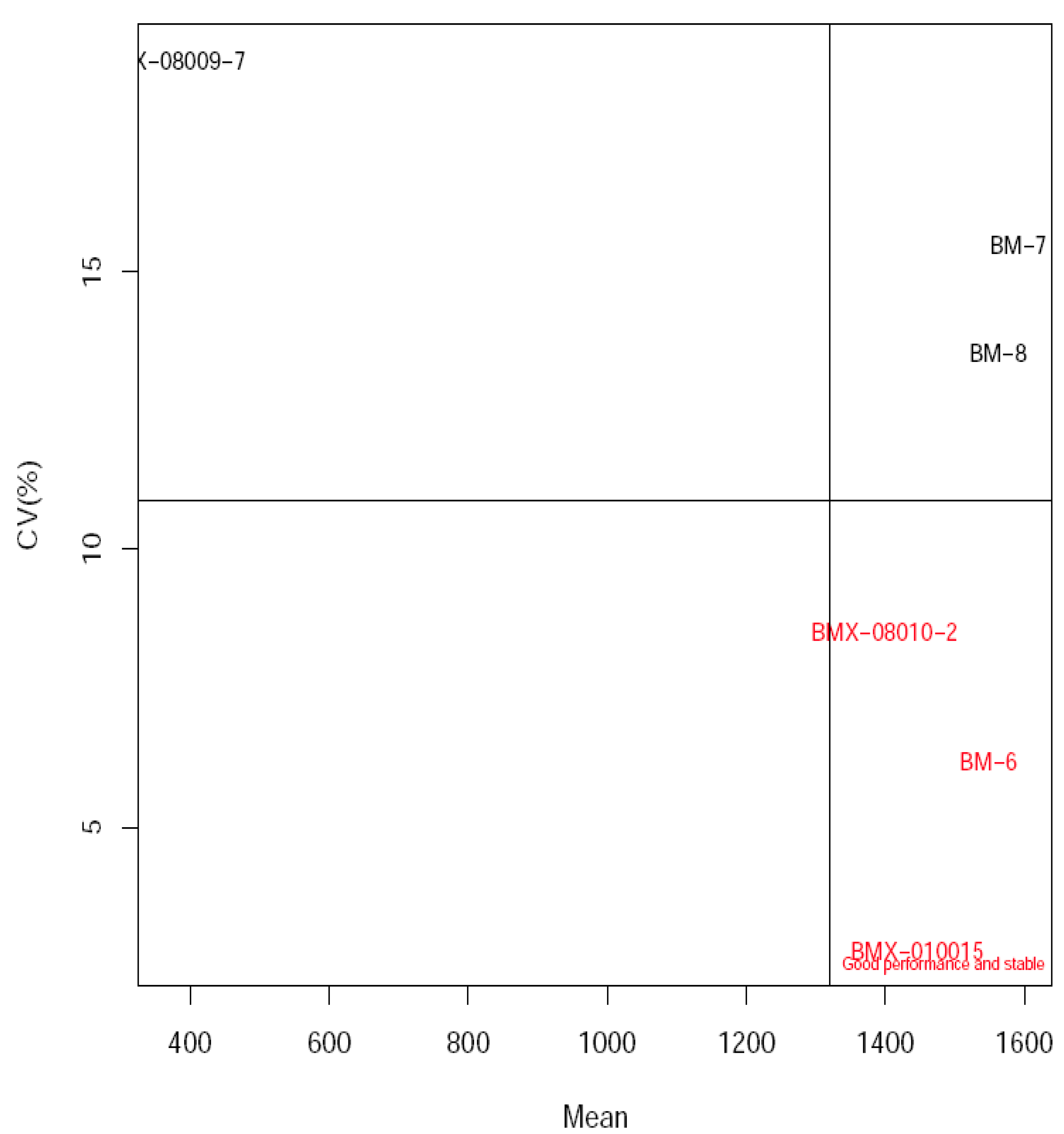
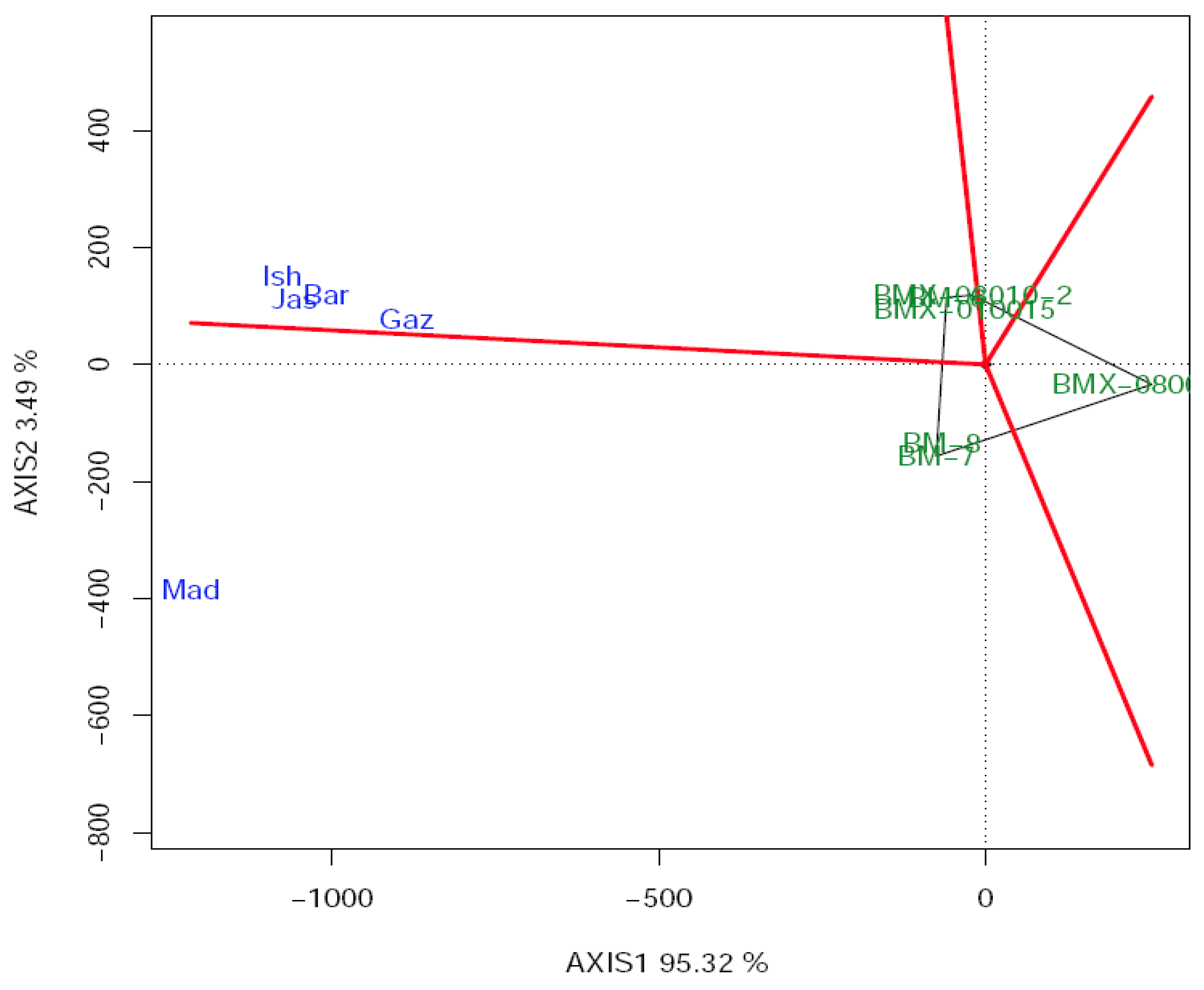
3.4. Estimation of Grain Yield Stability and Genotype Environment Interaction
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tomooka, N.; Lairungreeng, C.; Nakeeraks, P.; Egawa, Y.; Thavarasook, C. Production of mung bean and black gram. In Mung Bean and the Genetic Resources; TARC: Tohoku, Japan, 1991. [Google Scholar]
- Chauhan, Y.; Douglas, S.; Rachaputi, C.; Agius, R.C.N.; Martin, P.; King, W.K. Physiology of mungbean and development of the mungbean crop model. In Proceedings of the 1st Australian Summer Grains Conference Australia, Gold Coast, QLD, Australia, 21–24 June 2010; pp. 21–24. [Google Scholar]
- Sarker, U.; Oba, S. Salinity stress enhances color parameters, bioactive leaf pigments, vitamins, polyphenols, flavonoids and antioxidant activity in selected Amaranthus leafy vegetables. J. Sci. Food Agric. 2019, 99, 2275–2284. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Catalase, superoxide dismutase and ascorbate-glutathione cycle enzymes confer drought tolerance of Amaranthus tricolor. Sci. Rep. 2018, 8, 16496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, U.; Oba, S. The Response of Salinity Stress-Induced A. tricolor to Growth, Anatomy, Physiology, Non-Enzymatic and Enzymatic Antioxidants. Front. Plant Sci. 2020, 11, 559876. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Drought Stress Effects on Growth, ROS Markers, Compatible Solutes, Phenolics, Flavonoids, and Antioxidant Activity in Amaranthus tricolor. Appl. Biochem. Biotechnol. 2018, 186, 999–1016. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S. Response of nutrients, minerals, antioxidant leaf pigments, vitamins, polyphenol, flavonoid and antioxidant activity in selected vegetable amaranth under four soil water content. Food Chem. 2018, 252, 72–83. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Drought stress enhances nutritional and bioactive compounds, phenolic acids and antioxidant capacity of Amaranthus leafy vegetable. BMC PlantBiol. 2018, 18, 258. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Islam, M.T.; Oba, S. Salinity stress accelerates nutrients, dietary fiber, minerals, phytochemicals and antioxidant activity in Amaranthus tricolor leaves. PLoS ONE 2018, 13, e0206388. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Nutritional and bioactive constituents and scavenging capacity of radicals in Amaranthus hypochondriacus. Sci. Rep. 2020, 10, 19962. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Color attributes, betacyanin, and carotenoid profiles, bioactive components, and radical quenching capacity in selected Amaranthus gangeticus leafy vegetables. Sci. Rep. 2021, 11, 11559. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S. Nutrients, minerals, pigments, phytochemicals, and radical scavenging activity in Amaranthus blitum leafy vegetables. Sci. Rep. 2020, 10, 3868. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Leaf pigmentation, its profiles and radical scavenging activity in selected Amaranthus tricolor leafy vegetables. Sci. Rep. 2020, 10, 18617. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Hossain, M.M.; Oba, S. Nutritional and antioxidant components and antioxidant capacity in green morph Amaranthus leafy vegetable. Sci. Rep. 2020, 10, 1336. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S. Nutraceuticals, antioxidant pigments, and phytochemicals in the leaves of Amaranthus spinosus and Amaranthus viridis weedy species. Sci. Rep. 2019, 9, 20413. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Protein, dietary fiber, minerals, antioxidant pigments and phytochemicals, and antioxidant activity in selected red morph Amaranthus leafy vegetable. PLoS ONE 2019, 14, e0222517. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Islam, T.; Rabbani, G.; Oba, S. Antioxidant leaf pigments and variability in vegetable amaranth. Genetika 2018, 50, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Variability in total antioxidant capacity, antioxidant leaf pigments and foliage yield of vegetable amaranth. J. Int. Agr. 2018, 17, 1145–1153. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Antioxidant constituents of three selected red and green color Amaranthus leafy vegetable. Sci. Rep. 2019, 9, 18233. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Augmentation of leaf color parameters, pigments, vitamins, phenolic acids, flavonoids and antioxidant activity in selected Amaranthus tricolor under salinity stress. Sci. Rep. 2018, 8, 12349. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Nutraceuticals, phytochemicals, and radical quenching ability of selected drought-tolerant advance lines of vegetable amaranth. BMC Plant Biol. 2020, 20, 564. [Google Scholar] [CrossRef]
- Sarker, U.; Hossain, M.N.; Iqbal, M.A.; Oba, S. Bioactive Components and Radical Scavenging Activity in Selected Advance Lines of Salt-Tolerant Vegetable Amaranth. Front. Nutr. 2020, 7, 587257. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Phenolic profiles and antioxidant activities in selected drought-tolerant leafy vegetable amaranth. Sci. Rep. 2020, 10, 18287. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Polyphenol and flavonoid profiles and radical scavenging activity in leafy vegetable Amaranthus gangeticus. BMC Plant Biol. 2020, 20, 499. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S.; Daramy, M.A. Nutrients, minerals, antioxidant pigments and phytochemicals, and antioxidant capacity of the leaves of stem amaranth. Sci. Rep. 2020, 10, 3892. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Islam, T.; Rabbani, G.; Oba, S. Phenotypic divergence in vegetable amaranth for total antioxidant capacity, antioxidant profile, dietary fiber, nutritional and agronomic traits. Acta Agric. Scand. Soil Plant Sci. 2018, 68, 67–76. [Google Scholar] [CrossRef]
- Levitt, J. Response of Plants to Environmental Stress: Chilling, Freezing, and High Temperature Stress; Academic Press: New York, NY, USA, 1980; pp. 26–54. [Google Scholar]
- Mirzaei, A.; Rahim, N.; Parvaneh, V.; Meysam, M. Effects of Drought Stress on Qualitative and Quantitative Traits of Mung bean. Int. J. Biol. Food Vet. Agril. Eng. 2014, 8, 144–148. [Google Scholar]
- Boyer, J.S. Plant productivity and environment. Science 1982, 218, 443–448. [Google Scholar] [CrossRef]
- Sadasivan, R.; Natrajaratnam, N.; Dabu, R.; Muralidharan, V.; Rangasmay, S.R.S. Response of Mung bean cultivars to soil moisture stress at different growth phases. In Proceedings of the Second International Symposium, Bangkok, Thailand, 16–20 November 1987; AVRDC: Shanhua, Taiwan, 1988; pp. 260–262. [Google Scholar]
- Islam, M.R. Improving Mung bean Productivity under Drought Stress through Screening of Genotypes and Potassium Application. Ph.D. Thesis, Department of Agronomy, Hajee Mohammad Danesh Science and Technology University (HSTU), Dinajpur, Bangladesh, 2020. [Google Scholar]
- Akhtar, L.H.; Kashif, M.; Ali, M.; Aziz, T. Stability analysis for grain yield in Mung bean (Vigna radiata L. Wilczek) grown in different agro-climatic regions. Emir. J. Food Agri. 2010, 22, 490–497. [Google Scholar] [CrossRef]
- Kulsum, U.; Sarker, U.; Karim, A.; Mian, K. Additive main effects and multiplicative interaction (AMMI) analysis for yield of hybrid rice in Bangladesh. Trop. Agr. Develop. 2012, 56, 53–61. [Google Scholar]
- Pham, H.N.; Kang, M.S. Interrelationships among repeatability of several stability statistics estimated from international maize trials. Crop Sci. 1988, 28, 925–928. [Google Scholar] [CrossRef]
- Fikere, M.; Tadesse, T.; Letta, T. Genotype-environment interactions and stability parameters for grain yield of faba beans (Vacia faba L.) genotypes grown in south eastern Ethiopia. Int. J. Sustain. Crop Prod. 2008, 3, 80–87. [Google Scholar]
- Yan, W.; Cornelius, P.L.; Crossa, J.; Hunt, L.A. Two Types of GGE Biplots for Analyzing Multi-environment Trial Data. Crop Sci. 2001, 41, 656–663. [Google Scholar] [CrossRef] [Green Version]
- Piepho, H.P. Methods for comparing the yield stability of cropping systems. J. Agron. Crop Sci. 1998, 180, 193–213. [Google Scholar] [CrossRef]
- Becker, H.C.; Leon, J. Stability analysis in plant breeding. Plant Breed. 1988, 101, 1–23. [Google Scholar] [CrossRef]
- Lin, C.S.; Binns, M.R.; Lefkovitch, L.P. Stability analysis; where do we stand? Crop Sci. 1986, 26, 894–900. [Google Scholar] [CrossRef] [Green Version]
- Westcott, B. Some methods of analyzing genotype-environment interaction. Heredity 1986, 56, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Eberhart, S.A.; Russell, W.A. Stability parameters for comparing varieties. Crop Sci. 1966, 6, 36–40. [Google Scholar] [CrossRef] [Green Version]
- Gauch, H.G. Statistical Analysis of Regional Yield Trials: AMMI Analysis of Factorial Designs, 1st ed.; Elsevier Science: Amsterdam, The Netherlands, 1992. [Google Scholar]
- BARI (Bangladesh Agricultural Research Institute). Krishi Projukyi Hatboi (Handbook on Agro-Technology), 7th ed.; Farm Technology Group: Gazipur, Bangladesh, 2017. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria, 2019. Available online: http://www.R-project.org/ (accessed on 23 August 2021).
- Purchase, J.L.; Hatting, H.; van Deventer, C.S. Genotype × environment interaction of winter wheat (Triticum aestivum L.) in South Africa: II. Stability analysis of yield performance. S. Afr. J. Plant Soil. 2000, 17, 101–107. [Google Scholar] [CrossRef]
- Gollob, H.F. A statistical model which combines features of factor analytic and analysis of variance techniques. Psychometrika 1968, 33, 73–115. [Google Scholar] [CrossRef] [PubMed]
- Finlay, K.W.; Wilkinson, G.N. The analysis of adaptation in a plant breeding program. Aust. J. Agric. Res. 1963, 14, 742–754. [Google Scholar] [CrossRef] [Green Version]
- Atta, B.M.; Tariq, M.S.; Ghulam, A.; Muhammad, A.H. Genotype x environment interaction for grain yield in kabuli chickpea (Cicer arietinum L.) Genotypes Developed through mutation breeding. Pak. J. Bot. 2009, 41, 1883–1890. [Google Scholar]
- Asfaw, A.; Gurum, F.; Alemayehu, F.; Rezene, Y. Analysis of multi-environment grain yield trials in Mung bean Vigna radiata (L.) Wilczek based on GGE Bipot in Southern Ethiopia. J. Agr. Sci. Technol. 2012, 14, 389–398. [Google Scholar]
- Hussain, A.; Fatima, N. A bibliometric analysis of the ‘Chinese librarianship: An International Electronic Journal, (2006–2010)’. Chin. Librariansh. Int. Electron. J. 2011, 31, 1–14. [Google Scholar]
- Waniale, A.; Wanyera, N.; Talwana, H. Morphological and agronomic traits variations for Mung bean variety selection and improvement in Uganda. Afr. Crop Sci. J. 2014, 22, 123–136. [Google Scholar]
- Tarakanovas, P.; Ruzgus, V. Additive main effect and multiplicative interaction analysis of grain yield of wheat varieties in Lithuania. Agro. Res. 2006, 4, 91–98. [Google Scholar]
- Misra, R.C.; Das, S.; Patnaik, M.C. AMMI model analysis of stability and adaptability of late duration finger millet (Eleusinecoracana) genotypes. World Appl. Sci. J. 2009, 6, 1650–1654. [Google Scholar]
- Das, S.; Misra, R.C.; Patnaik, M.C.; Das, S.R. Genotype x environment interaction, adaptability and yield stability of mid—Early rice genotypes. Indian J. Agric. Res. 2010, 44, 104–111. [Google Scholar]
- Khan, A.A.; Alam, M.A.; Kabir, M.R. AMMI analysis for stability and environmental effects on grain yield of eight spring wheat varieties (Triticum aestivum L.) in Bangladesh. Bull. Inst. Trop Agri. Kyushu Univ. 2014, 37, 93–103. [Google Scholar]
- Win, K.S.; Kyi, W.; Than, D.M.; Nyo, M.H.; Tun, S. Genotype by environment interaction and stability analysis of seed yield, agronomic characters in Mung bean (Vigna radiata L. Wilczek) genotypes. Int. J. Adv. Res. 2018, 6, 926–934. [Google Scholar] [CrossRef] [Green Version]
- Yan, W.; Kang, M.S. GGE Biplot Analysis: A Graphical Tool for Breeders, Geneticists and Agronomists; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Yan, W.; Tinker, N.A. Biplot analysis of multi-environment trial data: Principles and applications. Can. J. Plant Sci. 2006, 86, 623–645. [Google Scholar] [CrossRef] [Green Version]
- Varma, P.N. Studies on Yield and Stress Tolerance in Green Gram. Master’s Thesis, Department of Plant Breeding and Genetics, Odisha University of Agriculture and Technology, Bhubaneswar, India, 2016. [Google Scholar]
- Yan, W.; Kang, M.S.; Ma, B.; Wood, S.; Cornelius, P.L. GGE Biplot vs. AMMI Analysis of Genotype-by environment Data. Crop Sci. 2007, 47, 643–655. [Google Scholar] [CrossRef]
- Mahmood, T.; Wang, X.; Ahmar, S.; Abdullah, M.; Iqbal, M.S.; Rana, R.M.; Yasir, M.; Khalid, S.; Javed, T.; Mora-Poblete, F.; et al. Genetic potential and inheritance pattern of phenological growth and drought tolerance in cotton (Gossypium hirsutum L.). Front. Plant Sci. 2021, 12, 915. [Google Scholar]
- Chowdhury, M.K.; Hasan, M.A.; Bahadur, M.M.; Islam, M.; Hakim, M.; Iqbal, M.A.; Javed, T.; Raza, A.; Shabbir, R.; Sorour, S.; et al. Evaluation of drought tolerance of some wheat (Triticum aestivum L.) genotypes through phenology, growth, and physiological indices. Agronomy 2021, 11, 1792. [Google Scholar] [CrossRef]
- Javed, T.; Shabbir, R.; Ali, A.; Afzal, I.; Zaheer, U.; Gao, S.J. Transcription factors in plant stress responses: Challenges and potential for sugarcane improvement. Plants 2020, 9, 491. [Google Scholar] [CrossRef] [PubMed]
- Shabbir, R.; Javed, T.; Afzal, I.; Sabagh, A.E.; Ali, A.; Vicente, O.; Chen, P. Modern biotechnologies: Innovative and sustainable approaches for the improvement of sugarcane tolerance to environmental stresses. Agronomy 2021, 11, 1042. [Google Scholar] [CrossRef]
- Raza, A.; Charagh, S.; Sadaqat, N.; Jin, W. Arabidopsis thaliana: Model plant for the study of abiotic stress responses. In The Plant Family Brassicaceae; Springer: Singapore, 2020; pp. 129–180. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Genotypes | Distinction | Pedigree | Remark |
|---|---|---|---|---|
| 1 | BARI Mung-8 | ST | Selection from the local collection (LM-101) | RV |
| 2 | BMX-08010-2 | ST | BARI Mung-6 × BMX-9902-2 | AL |
| 3 | BMX-010015 | ST | NM-94 × BARI Mung -3 | AL |
| 4 | BMX-08009-7 | ST | BARI Mung-6 × BU Mung-2 | AL |
| 5 | BARI Mung-6 | Check | NM-36 × VC-2768A (AVRDC) | RV |
| 6 | BARI Mung-7 | Check | VC-3960A-88 × VC-6173C | RV |
| Location | Variable | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Monthly Total Rainfall(mm) | Monthly Average Temperature (°C) | |||||||||||
| Altitude (m) | Geographical Position | Soil Texture | March | April | May | June | March | April | May | June | ||
| Ishurdi | 16.00 | 24° 03′ N 89° 05′ E | CL | 27.2 | 61.9 | 127.1 | 167.9 | Tmax Tmin | 32.2 17.8 | 34.1 22.9 | 35.7 25.7 | 34.9 26.2 |
| Jashore | 6.10 | 23° 17′ N 89° 21′ E | CL | 85.0 | 155.0 | 152.0 | 155.0 | Tmax Tmin | 32.2 19.3 | 34.9 23.4 | 36.7 26.2 | 36.0 26.7 |
| Barishal | 2.10 | 22° 48′ N 90° 37′ E | SC | 38.0 | 78.30 | 125.40 | 173.8 | Tmax Tmin | 31.6 20.5 | 33.1 23.0 | 34.8 26.0 | 33.2 26.6 |
| Madaripur | 7.00 | 23°10 N 90°12′ E | SL | 52.3 | 117.2 | 229.1 | 370.9 | Tmax Tmin | 31.6 20.5 | 33.1 23.7 | 32.9 25.1 | 31.7 25.9 |
| Gazipur | 14.00 | 22° 46′ N 90° 39′ E | SCL | 126.0 | 112.0 | 233.6 | 185.0 | Tmax Tmin | 32.0 20.0 | 33.5 22.3 | 35.3 25.3 | 34.1 26.5 |
| Location | pH | OM (%) | Total N (%) | Exchangeable K meq 100 g Soil−1 | P | S | Zn | B |
|---|---|---|---|---|---|---|---|---|
| µg g−1 | ||||||||
| Ishurdi | 7.36 | 1.10 | 0.060 | 0.31 | 31.12 | 10.75 | 1.43 | 0.35 |
| Jashore | 7.6 | 1.17 | 0.065 | 0.18 | 13.00 | 14.00 | 0.56 | 0.16 |
| Barishal | 6.8 | 0.92 | 0.080 | 0.07 | 12.00 | 10.20 | 0.60 | 0.54 |
| Madaripur | 7.4 | 1.45 | 0.065 | 0.16 | 16.00 | 18.30 | 1.10 | 0.16 |
| Gazipur | 6.25 | 1.09 | 0.087 | 0.08 | 7.41 | 10.07 | 0.26 | 0.17 |
| Critical level (CL) | - | - | 0.12 | 0.12 | 10 | 10 | 0.60 | 0.20 |
| Entries | Days to Flowering | Pi | bi | S2di | |||||
|---|---|---|---|---|---|---|---|---|---|
| Ish | Gaz | Jas | Bar | Mad | Mean | ||||
| BMX-010015 | 43 | 41 | 38 | 40 | 38 | 40 | 0.60 | 0.75 * | 10.64 |
| BMX-08009-7 | 38 | 48 | 40 | 45 | 45 | 43 | 3.60 | 0.56 *** | 5.78 ** |
| BMX-08010-2 | 37 | 38 | 40 | 35 | 40 | 38 | −1.40 | 1.48 ** | 4.54 *** |
| BARI Mung-6 | 34 | 43 | 36 | 41 | 38 | 39 | −0.40 | 0.66 ** | 2.87 *** |
| BARI Mung-7 | 39 | 40 | 42 | 34 | 36 | 39 | 0.00 | 0.40 *** | 14.57 ** |
| BARI Mung-8 | 36 | 37 | 39 | 34 | 36 | 37 | −2.40 | 2.15 | 12.63 |
| Mean | 37.83 | 41.17 | 39.17 | 38.17 | 38.83 | 39.40 | |||
| Environmental index (Ij) | −1.57 | 1.77 | −0.23 | −1.23 | −0.57 | ||||
| CV (%) | 1.65 | 2.60 | 1.90 | 2.79 | 4.60 | - | |||
| LSD (0.05) | 2.12 | 2.42 | 3.53 | 2.43 | 2.81 | - | |||
| Entries | Days to Maturity | Pi | bi | S2di | |||||
|---|---|---|---|---|---|---|---|---|---|
| Ish | Gaz | Jas | Bar | Mad | Mean | ||||
| BMX-010015 | 70 | 67 | 69 | 70 | 66 | 68 | 3.63 | 0.75 *** | 2.46 |
| BMX-08009-7 | 64 | 70 | 70 | 74 | 66 | 69 | 4.63 | 1.59 *** | 3.16 *** |
| BMX-08010-2 | 63 | 63 | 64 | 64 | 68 | 64 | 0.03 | 1.62 *** | 16.47 *** |
| BARI Mung-6 | 61 | 64 | 60 | 66 | 56 | 61 | −3.37 | 0.17 ** | 3.03 *** |
| BARI Mung-7 | 61 | 63 | 63 | 60 | 60 | 62 | −2.17 | 0.46 *** | 12.84 ** |
| BARI Mung-8 | 61 | 61 | 62 | 61 | 60 | 62 | −2.77 | 1.39 *** | 1.13 *** |
| Mean | 63.33 | 64.67 | 64.67 | 65.83 | 62.67 | 64.37 | |||
| Environmental index (Ij) | −1.03 | 0.30 | 0.30 | 1.47 | −1.70 | 0.00 | |||
| CV (%) | 1.94 | 2.00 | 0.80 | 2.77 | 7.07 | - | |||
| LSD (0.05) | 2.73 | 2.66 | 2.86 | 3.89 | 4.08 | - | |||
| Entries | Pods Plant−1 | Pi | bi | S2di | |||||
|---|---|---|---|---|---|---|---|---|---|
| Ish | Gaz | Jas | Bar | Mad | Mean | ||||
| BMX-010015 | 26.21 | 17.42 | 21.31 | 19.42 | 20.2 | 20.91 | 0.04 | 2.89 *** | 26.52 *** |
| BMX-08009-7 | 22.12 | 26.32 | 22.10 | 11.43 | 25.22 | 21.40 | 0.53 | 0.75 | 20.25 |
| BMX-08010-2 | 25.64 | 23.25 | 24.00 | 18.6 | 24.27 | 23.15 | 2.28 | 0.11 *** | 10.61 ** |
| BARI Mung-6 | 13.14 | 25.22 | 25.17 | 9.64 | 38.03 | 22.24 | 1.37 | −0.43 *** | 7.93 |
| BARI Mung-7 | 19.44 | 17.05 | 17.80 | 24.20 | 18.07 | 19.31 | −1.56 | 1.24 | 19.76 *** |
| BARI Mung-8 | 19.97 | 17.58 | 18.33 | 16.60 | 18.60 | 18.22 | −2.65 | 1.45 | 31.42 *** |
| Mean | 21.09 | 21.14 | 21.45 | 16.65 | 24.07 | 20.87 | |||
| Environmental index (Ij) | 0.22 | 0.27 | 0.58 | −4.22 | 3.19 | ||||
| CV (%) | 12.55 | 10.16 | 10.91 | 6.24 | 11.18 | - | |||
| LSD (0.05) | 4.98 | 2.59 | 3.34 | 8.94 | 3.61 | - | |||
| Entries | 100 Grain Weight | Pi | bi | S2di | |||||
|---|---|---|---|---|---|---|---|---|---|
| Ish | Gaz | Jas | Bar | Mad | Mean | ||||
| BMX-010015 | 4.47 | 4.34 | 4.29 | 4.42 | 4.52 | 4.41 | −0.02 | 2.44 *** | 0.08 |
| BMX-08009-7 | 3.8 | 4.63 | 5.00 | 3.13 | 5.30 | 4.37 | −0.06 | 1.32 | 0.16 *** |
| BMX-08010-2 | 3.91 | 3.43 | 3.55 | 4.52 | 3.93 | 3.87 | −0.56 | 0.64 | 0.45 *** |
| BARI Mung-6 | 4.47 | 3.92 | 5.03 | 3.40 | 5.44 | 4.45 | 0.02 | −0.08 *** | −0.08 |
| BARI Mung-7 | 5.12 | 4.64 | 4.97 | 5.94 | 5.60 | 5.25 | 0.82 | 1.68 ** | 0.59 *** |
| BARI Mung-8 | 4.15 | 3.67 | 3.78 | 4.75 | 4.80 | 4.23 | −0.20 | 0.002 *** | 0.06 |
| Mean | 4.32 | 4.11 | 4.44 | 4.36 | 4.93 | 4.43 | |||
| Environmental index (Ij) | −0.11 | −0.33 | 0.01 | −0.07 | 0.50 | ||||
| CV (%) | 2.01 | 1.53 | 1.48 | 2.45 | 7.28 | - | |||
| LSD (0.05) | 0.89 | 0.41 | 0.08 | 1.05 | 0.62 | - | |||
| SV | DF | SS | MSS | % Treatment SS | % Interaction SS | Cumulative % |
|---|---|---|---|---|---|---|
| Gen | 5 | 16496312.7 | 3299263 ** | 91.03 | ||
| Env | 4 | 640406.65 | 160101.7 ** | 3.53 | ||
| Gen x Env | 20 | 985227.21 | 49261.36 ** | 5.44 | ||
| IPCA I | 8 | 706752.70 | 88344.09 ** | 71.74 | 71.74 | |
| IPCA II | 6 | 160280.95 | 26713.49 ** | 16.27 | 88.00 | |
| IPCA III | 4 | 109876.28 | 27469.07 ** | 11.15 | 99.16 | |
| IPCA IV | 2 | 8317.28 | 4158.639 ns | 0.84 | 100 | |
| Residuals | 60 | 110366.84 | 1839.45 |
| Entries | Ish | Gaz | Jas | Bar | Mad | Mean | Pi | bi | S2di | ASV | YSI | RBY | RBASV | CV(%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BMX-010015 | 1483.27 | 1434.80 | 1478.92 | 1383.68 | 1448.30 | 1446.07 | 125.66 | 0.29 | 7804.83 | 1.98 | 6 | 4 | 2 | 2.78 |
| BMX-08009-7 | 327.14 | 455.74 | 424.73 | 283.96 | 394.41 | 375.20 | −945.21 | 0.30 * | 11,548.75 ** | 1.78 | 7 | 6 | 1 | 18.81 |
| BMX-08010-2 | 1549.44 | 1299.88 | 1493.91 | 1278.86 | 1372.24 | 1398.99 | 78.58 | 0.65 ** | 21,838.87 | 2.39 | 9 | 5 | 4 | 8.51 |
| BARI Mung-6 | 1531.48 | 1482.66 | 1712.74 | 1478.51 | 1533.33 | 1548.20 | 227.79 | 0.57 *** | 597.2057 *** | 2.16 | 6 | 3 | 3 | 6.18 |
| BARI Mung-7 | 1455.67 | 1433.48 | 1708.75 | 1387.67 | 1969.62 | 1591.71 | 271.30 | 2.41 *** | 4988.177 * | 4.45 | 7 | 1 | 6 | 15.48 |
| BARI Mung-8 | 1589.96 | 1443.45 | 1459.94 | 1397.65 | 1917.04 | 1562.28 | 241.87 | 1.78 ** | 13,323.05 *** | 3.88 | 7 | 2 | 5 | 13.55 |
| Mean | 1322.83 | 1258.34 | 1379.83 | 1201.72 | 1439.16 | 1320.41 | ||||||||
| Ei (Ij) | 2.42 | −62.07 | 59.42 | −118.69 | 118.75 | |||||||||
| CV (%) | 3.20 | 3.00 | 1.68 | 2.76 | 4.10 | - | ||||||||
| LSD (0.05) | 54.40 | 48.54 | 29.74 | 42.61 | 75.84 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.R.; Sarker, B.C.; Alam, M.A.; Javed, T.; Alam, M.J.; Zaman, M.S.U.; Azam, M.G.; Shabbir, R.; Raza, A.; Habib-ur-Rahman, M.; et al. Yield Stability and Genotype Environment Interaction of Water Deficit Stress Tolerant Mung Bean (Vigna radiata L. Wilczak) Genotypes of Bangladesh. Agronomy 2021, 11, 2136. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11112136
Islam MR, Sarker BC, Alam MA, Javed T, Alam MJ, Zaman MSU, Azam MG, Shabbir R, Raza A, Habib-ur-Rahman M, et al. Yield Stability and Genotype Environment Interaction of Water Deficit Stress Tolerant Mung Bean (Vigna radiata L. Wilczak) Genotypes of Bangladesh. Agronomy. 2021; 11(11):2136. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11112136
Chicago/Turabian StyleIslam, Mohammad Rafiqul, Bikash Chandra Sarker, Mohammad Ashraful Alam, Talha Javed, Mohammad Jahangir Alam, Mohammad Shahin Uz Zaman, Mohammad Golam Azam, Rubab Shabbir, Ali Raza, Muhammad Habib-ur-Rahman, and et al. 2021. "Yield Stability and Genotype Environment Interaction of Water Deficit Stress Tolerant Mung Bean (Vigna radiata L. Wilczak) Genotypes of Bangladesh" Agronomy 11, no. 11: 2136. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11112136