
Regulation of Phosphorus Supply on Nodulation and Nitrogen Fixation in Soybean Plants with Dual-Root Systems

Abstract
:1. Introduction
2. Materials and Methods
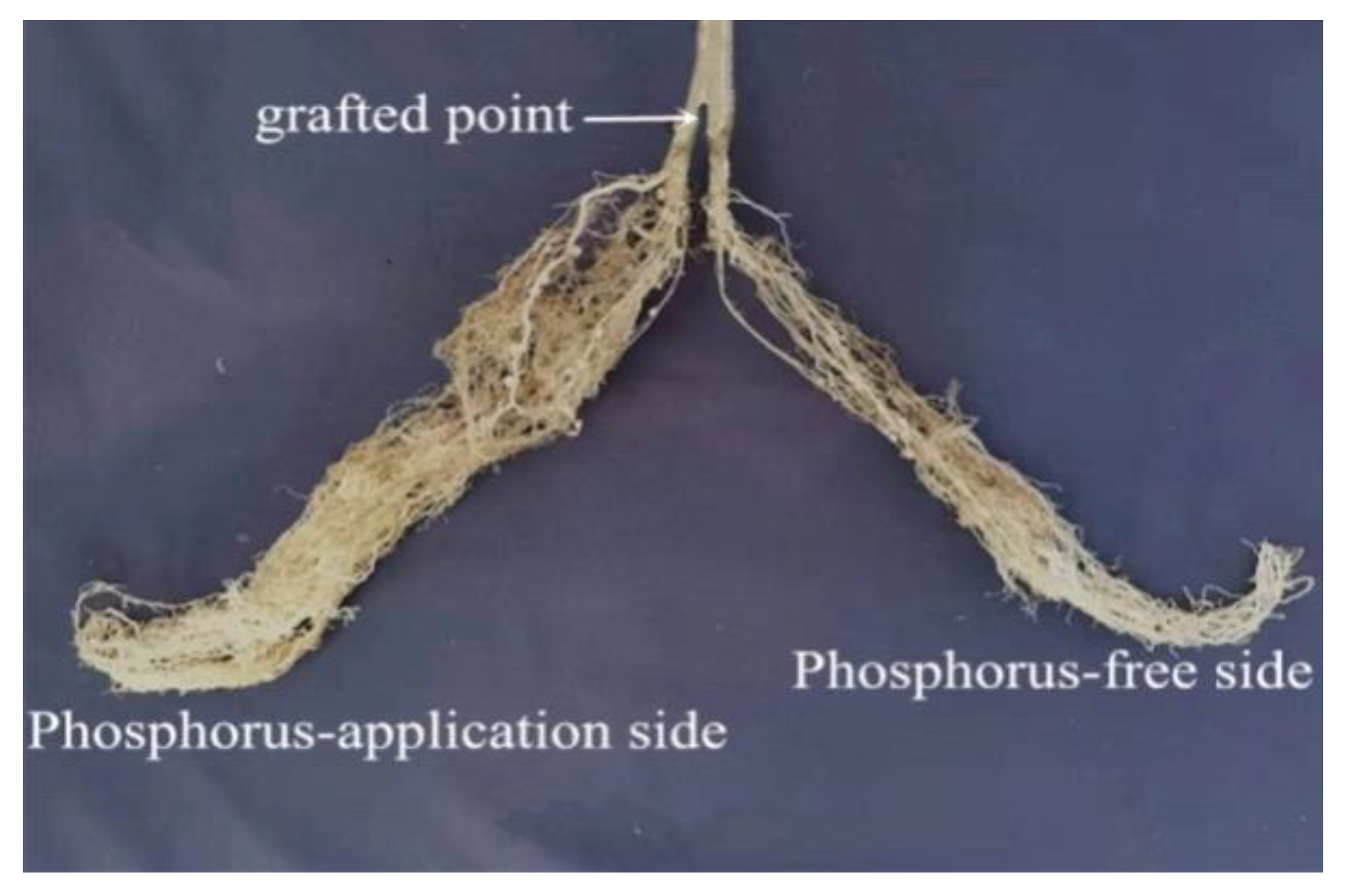
2.1. Experimental Treatments
2.2. Sampling and Measurement
2.3. Statistical Analyses
3. Results
3.1. Effects of P Supply on P Content and Dry Matter Accumulation in Soybean Plants with Dual-Root Systems
3.2. Effect of P Supply on the Activity of Nitrogenase and the Nodule Ureide Content in Soybean Plants with Dual-Root Systems
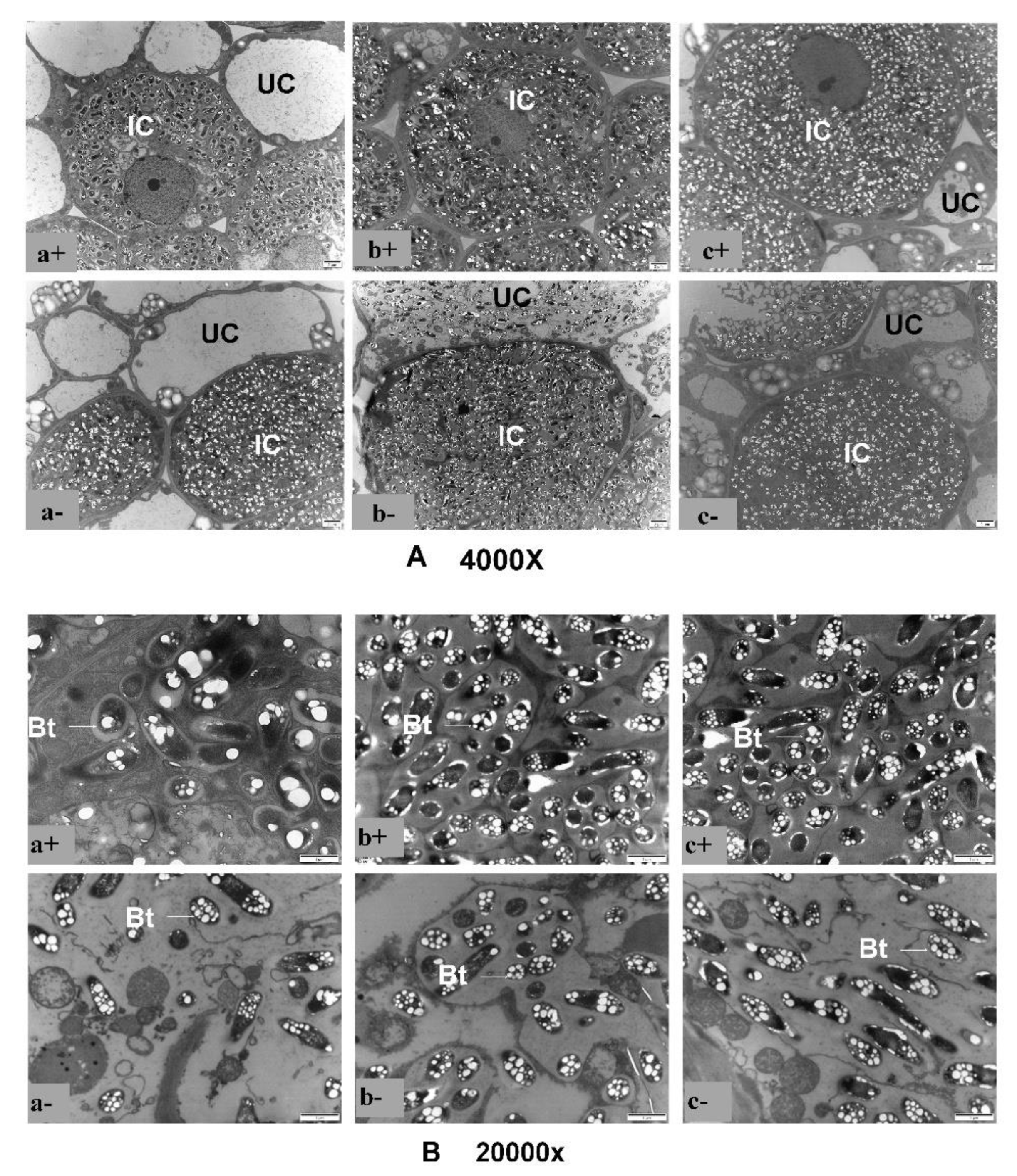
3.3. Effect of P Supply on the Ultrastructure of Soybean Plants with Dual-Root Systems Root Nodules
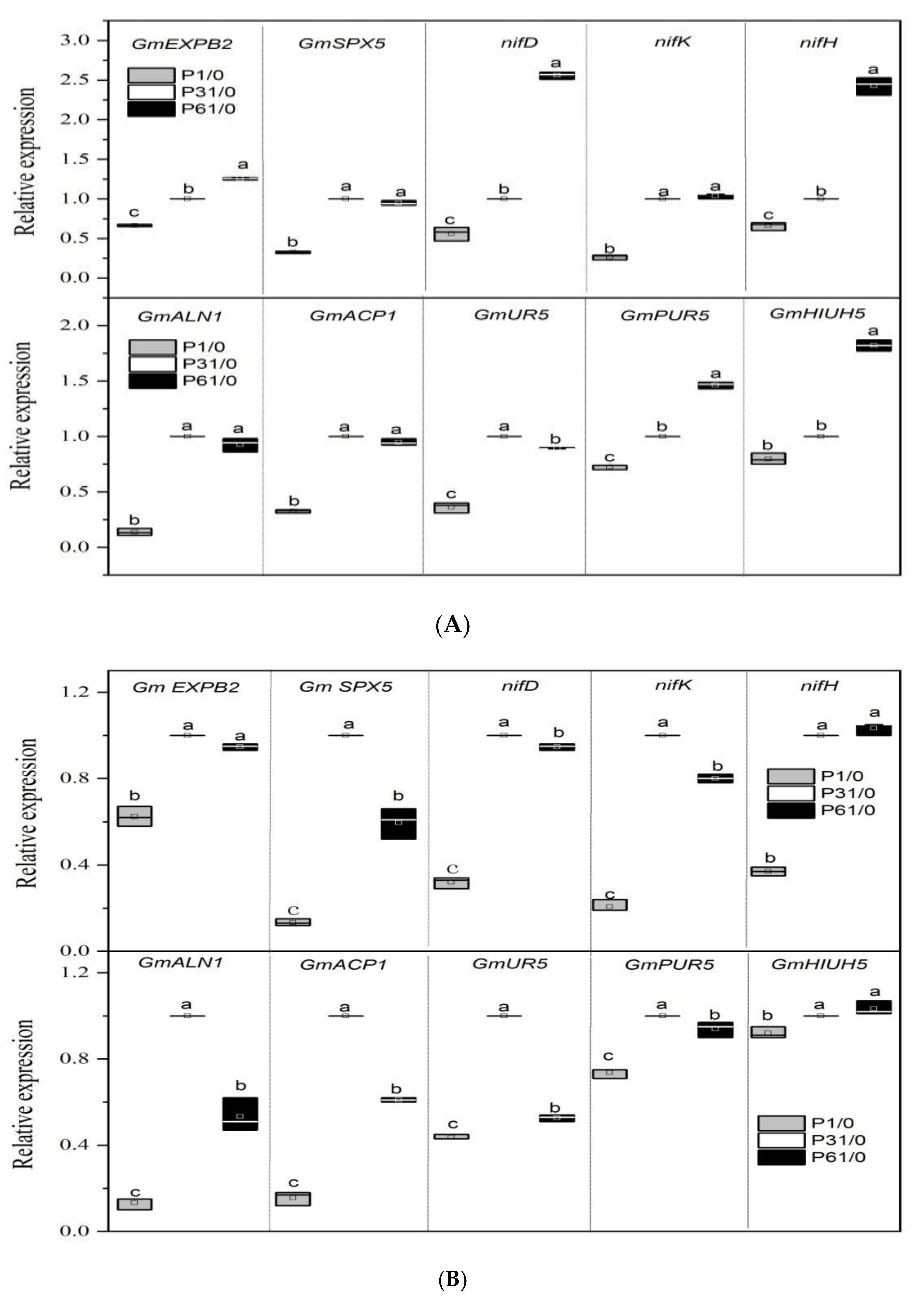
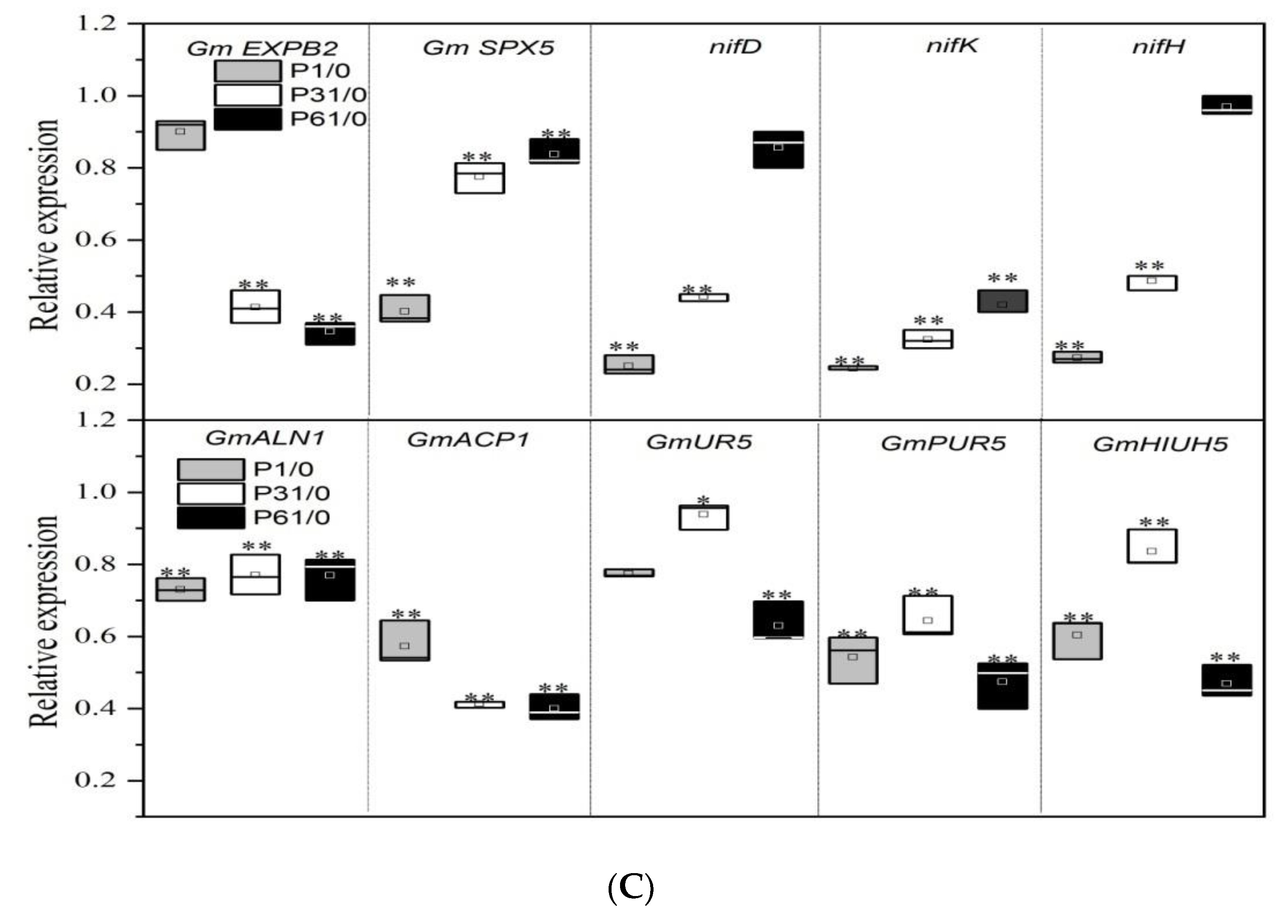
3.4. Effect of P Supply on the Expression of Key Genes Related to Nodulation and Nitrogen Fixation in Soybean Plants with Dual-Root Systems
4. Discussion
4.1. Effects of P Supply on Nodulation and Nitrogen Fixation in Soybean Plants
4.2. Effects of P Supply Level on P Absorption and Transport in Soybean Plants
5. Conclusions
- In the dual-root soybean plants supplied with P on only one side, the nodule weight, ARA, SNA, ureide content, the number of Bts, the number of ICs, and relative expression levels of key genes related to nodulation and nitrogen fixation (GmEXPB2, GmSPX5, nifH, nifD, nifK, GmALN1, GmACP1, GmUR5, GmPUR5, and GmHIUH5) on the P+ side increased with the increase in P supply during the VC-R1 period. Those on the P− side showed the same pattern as those on the P+ side but less prominently. Therefore, P supply regulates soybean nodulation and nitrogen fixation.
- When the level of phosphorus supply was increased from 1 mg/L to 31 mg/L, the increase on the P– side root was significant, and nodule phosphorus content increased by 57.14–85.71% and 68.75–75.00%; ARA and SNA were 218.64–383.33% and 11.41–16.11%, respectively, and ureide content was 118.18–156.44%. When the level of phosphorus supply was increased from 31 mg/L to 61 mg/L, the increases in the regulation ability of the root and nodule phosphorus content were only 0.00–9.09% and 25.00–29.63%, respectively; ARA and SNA were 36.21–50.00% and 10.46–15.78%, respectively, and ureide content was 6.02–9.27%.
- When the dual-root soybean plants were supplied with different concentrations of P on one side, the P content in the roots and nodules on the P− side increased with the increase in P supply. These findings show that P can be transported to the roots and nodules on the non-supplied side through the shoots, and the nodules are prioritized to receive P.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vance, C.P.; Uhde-Stone, C.; Allan, D.L. Phosphorus acquisition and use: Critical adaptations by plantsfor securing a nonrenewable resource. New Phytol. 2010, 157, 423–447. [Google Scholar] [CrossRef] [Green Version]
- Jakobsen, I. The role of phosphorus m nitrogen fixation by young pea plants (Pisum sativum). Physiol. Plant. 1985, 64, 190–196. [Google Scholar] [CrossRef]
- Chen, Z.; Cui, Q.; Liang, C.; Sun, L.; Tian, J.; Liao, H. Identification of differentially expressed proteins in soybean nodules under phosphorus deficiency through proteomic analysis. Proteomics 2011, 11, 4648–4659. [Google Scholar] [CrossRef]
- Sulieman, S.; Ha, C.V.; Schulze, J.; Tran, L.S. Growth and nodulation of symbiotic Medicago truncatula at different levels of phosphorus availability. J. Exp. Bot. 2013, 64, 2701–2712. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.; Zhuang, Q.; Zhu, S.; Xiao, B.; Liang, C.; Liao, H.; Tian, J. Genome Wide Transcriptome Analysis Reveals Complex Regulatory Mechanisms Underlying Phosphate Homeostasis in Soybean Nodules. Int. J. Mol. Sci. 2018, 19, 2924. [Google Scholar] [CrossRef] [Green Version]
- Taliman, N.; Dong, Q.; Echigo, K.; Raboy, V.; Saneoka, H. Effect of Phosphorus Fertilization on the Growth, Photosynthesis, Nitrogen Fixation, Mineral Accumulation, Seed Yield, and Seed Quality of a Soybean Low-Phytate Line. Plants 2019, 8, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulgarelli, R.G.; De Oliveira, V.H.; de Andrade, S.A.L. Arbuscular mycorrhizal symbiosis alters the expression of PHT1 phosphate transporters in roots and nodules of P-starved soybean plants. Theor. Exp. Plant Physiol. 2020, 32, 243–253. [Google Scholar] [CrossRef]
- Qin, L.; Zhao, J.; Tian, J.; Chen, L.; Sun, Z.; Guo, Y.; Lu, X. The High-Affinity Phosphate Transporter GmPT5 Regulates Phosphate Transport to Nodules and Nodulation in Soybean. Plant Physiol. 2012, 159, 1634–1643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, M.; Cheng, Z.; Zhang, X.-M.; Huang, P.; Fan, C.; Yu, G.; Chen, F.; Xu, K.; Chen, Q.; Miao, Y.; et al. Spatial Divergence of PHR-PHT1 Modules Maintains Phosphorus Homeostasis in Soybean Nodules. Plant Physiol. 2020, 184, 01209–02019. [Google Scholar] [CrossRef]
- Israel, D.W. Investigation of the role of phosphorus in symbiotic dinitrogen fixation. Plant Physiol. 1987, 84, 835–840. [Google Scholar] [CrossRef] [Green Version]
- Israel, D.W. Symbiotic dinitrogen fixation and host-plant growth during development of and recovery from phosphorus deficiency. Physiol. Plant. 1993, 88, 294–300. [Google Scholar] [CrossRef]
- Tsvetkova, G.; Georgiev, G. Effect of phosphorus nutrition on the nodulation, nitrogen fixation and nutrient-use efficiency of Bradyrhizobiumjaponicun soybean (Glycine max L. Merr.) symbiosis. Bulg. J. Plant Physiol. 2003, 3, 315–335. [Google Scholar]
- Miao, S.; Qiao, Y.; Han, X.-Z.; An, M. Nodule Formation and Development in Soybeans (Glycine max L.) in Response to Phosphorus Supply in Solution Culture1. Pedosphere 2007, 17, 36–43. [Google Scholar] [CrossRef]
- Dong, Q.; Echigo, K.; Raboy, V.; Saneoka, H. Seedling growth, physiological characteristics, nitrogen fixation, and root and nodule phytase and phosphatase activity of a low-phytate soybean line. Plant Physiol. Biochem. 2020, 149, 225–232. [Google Scholar] [CrossRef]
- Cassman, K.; Whitney, A.; Stockinger, K. Root Growth and Dry Matter Distribution of Soybean as Affected by Phosphorus Stress, Nodulation, and Nitrogen Source1. Crop. Sci. 1980, 20, 239–244. [Google Scholar] [CrossRef]
- Sa, T.M.; Israel, D.W. Energy status and functioning of phosphorus-deficient soybean nodules. Plant Physiol. 1991, 97, 928–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, C.; Hinsinger, P.; Drevon, J.J.; Jaillard, B. Phosphorus Deficiency Impairs Early Nodule Functioning and Enhances Proton Release in Roots of Medicago truncatula L. Ann. Bot. 2001, 88, 131–138. [Google Scholar] [CrossRef] [Green Version]
- Le Roux, M.; Khan, S.; Valentine, A. Nitrogen and carbon costs of soybean and lupin root systems during phosphate starvation. Symbiosis 2009, 48, 102–109. [Google Scholar] [CrossRef]
- Mandri, B.; Drevon, J.J.; Adnane, B.; Oufdou, K.; Faghire, M.; Plassard, C.; Payre, H.; Ghoulam, C. Interactions between common bean genotypes and rhizobia strains isolated from moroccan soils for growth, phosphatase and phytase activities under phosphorus deficiency conditions. J. Plant Nutr. 2012, 35, 1477–1490. [Google Scholar] [CrossRef]
- Araújo, A.P.; Plassard, C.; Drevon, J.J. Phosphatase and phytase activities in nodules of common bean genotypes at different levels of phosphorus supply. Plant Soil 2008, 312, 129. [Google Scholar] [CrossRef]
- Shu, L.; Shen, J.; Rengel, Z.; Tang, C.; Zhang, F.; Cawthray, G. Formation of cluster roots and citrate exudation by Lupinusalbus in response to localized application of different phosphorus sources. Plant Sci. 2007, 172, 1017–1024. [Google Scholar] [CrossRef]
- Scott, B.; Robson, A. The Distribution of Mg, P and K in the Split Roots of Subterranean Clover. Ann. Bot. 1991, 67, 251–256. [Google Scholar] [CrossRef]
- Snapp, S.; Lynch, J. Phosphorus Distribution and Remobilization in Bean Plants as Influenced by Phosphorus Nutrition. Crop. Sci. 1996, 36, 929–935. [Google Scholar] [CrossRef]
- Xia, X.; Ma, C.; Dong, S.; Xu, Y.; Gong, Z. Effects of nitrogen concentrations on nodulation and nitrogenase activity in dual root systems of soybean plants. Soil Sci. Plant Nutr. 2017, 63, 470–482. [Google Scholar] [CrossRef]
- Shen, J.; Li, H.; Neumann, G.; Zhang, F. Nutrient uptake, cluster root formation and exudation of protons and citrate in Lupinusalbus as affected by localized supply of phosphorus in a split-root system. Plant Sci. 2005, 168, 837–845. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water culture method for growing plants without soil. Calif. Agric. Exp. Stn. Circ. 1950, 347, 33. [Google Scholar]
- Yao, Y.; Wu, D.; Gong, Z.; Zhao, J.; Ma, C. Variation of nitrogen accumulation and yield in response to phosphorus nutrition of soybean (Glycine max L. Merr.). J. Plant Nutr. 2018, 41, 1138–1147. [Google Scholar] [CrossRef]
- Fehr, W.R.; Caviness, C.E.; Burmood, D.T.; Pennington, J.S. Stage of Development Descriptions for Soybeans, Glycine Max (L.) Merrill1. Crop. Sci. 1971, 11, 929–931. [Google Scholar] [CrossRef]
- Gremaud, M.F.; Harper, J.E. Selection and initial characterization of partially nitrate tolerant nodulation mutants of soybean. Plant Physiol. 1989, 89, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Goodchild, D.J.; Bergersen, F.J. Electron microscopy of the infection and subsequent development of soybean nodule cells. J. Bacteriol 1966, 92, 204–213. [Google Scholar] [CrossRef] [Green Version]
- Trijbels, F.; Vogels, G.D. Degradation of allantoin by Pseudomonas acidovorans. Biochim. Acta 1966, 113, 292–301. [Google Scholar] [CrossRef]
- Sulieman, S.; Kusano, M.; Ha, C.; Watanabe, Y.; Abdalla, M.; Abdelrahman, M. Divergent metabolic adjustments in nodules are indispensable for efficient N2 fixation of soybean under phosphate stress. Plant Sci. 2019, 289, 110249. [Google Scholar] [CrossRef]
- Carter, A.M.; Tegeder, M. Increasing Nitrogen Fixation and Seed Development in Soybean Requires Complex Adjustments of Nodule Nitrogen Metabolism and Partitioning Processes. Curr. Biol. 2016, 26, 2044–2051. [Google Scholar] [CrossRef] [Green Version]
- Silva, R.; Gomes de Faria, A.; Neto, A.; Pereira, T.; Dias, J.; Martins dos Santos, A.; Junior, A.; Freitas, G.; Nascimento, V. High P Availability in Brazilian Cerrado Soils Reduces Nodulation and Grain Yield of Soybean. Commun. Soil Sci. Plant Anal. 2019, 50, 1267–1277. [Google Scholar] [CrossRef]
- Rotaru, V.; Sinclair, T. Interactive influence of phosphorus and iron on nitrogen fixation by soybean. Environ. Exp. Bot. 2009, 66, 94–99. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, Z.; Youbin, K.; Li, X.; Li, W.; Du, H.; Zhang, C. GmPAP12 Is Required for Nodule Development and Nitrogen Fixation Under Phosphorus Starvation in Soybean. Front. Plant Sci. 2020, 11, 450. [Google Scholar] [CrossRef]
- Magadlela, A.; Steenkamp, E.; Valentine, A. Variable P supply affects N metabolism in a legume tree, Virgiliadivaricata, from nutrient-poor Mediterranean-Type ecosystems. Funct. Plant Biol. 2015, 43, 287–297. [Google Scholar] [CrossRef] [Green Version]
- Isidra-Arellano, M.; Reyero-SaavedraI, M.; Sánchez-Correa, M.; Pingault, L.; Sen, S.; Joshi, T. Phosphate Deficiency Negatively Affects Early Steps of the Symbiosis between Common Bean and Rhizobia. Genes 2018, 9, 498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gentili, F.; Wall, L.G.; Huss-Danell, K. Effects of phosphorus and nitrogen on nodulation are seen already at the stage of early cortical cell divisions in Alnusincana. Ann. Bot. 2006, 98, 309–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhao, J.; Tan, Z.; Zeng, R.; Liao, H. GmEXPB2, a Cell Wall b-Expansin, Affects Soybean Nodulation through Modifying Root Architecture and Promoting Nodule Formation and Development. Plant Physiol. 2015, 169, 2640–2653. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Xie, J.; Liao, H.; Wang, X. Overexpression of β-expansin gene GmEXPB2 improves phosphorus efficiency in soybean. Physiol. Plant. 2013, 150, 194–204. [Google Scholar] [CrossRef]
- Yao, Z.; Tian, J.; Liao, H. Comparative characterization of GmSPX members reveals that GmSPX3 is involved in phosphate homeostasis in soybean. Ann. Bot. 2014, 114, 477–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curatti, L.; Brown, C.S.; Ludden, P.W.; Rubio, L.M. Genes required for rapid expression of nitrogenase activity in Azotobactervinelandii. Proc. Natl. Acad. Sci. USA 2005, 102, 6291–6296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esfahani, M.N.; Sulieman, S.; Schulze, J.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L. Approaches for enhancement of N2 fixation efficiency of chickpea (Cicer arietinum L.) under limiting nitrogen conditions. Plant Biotechnol. J. 2014, 12, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Nasr Esfahani, M.; Kusano, M.; Nguyen, K.H.; Watanabe, Y.; Ha, C.V.; Saito, K.; Sulieman, S.; Herrera-Estrella, L.; Tran, L.S. Adaptation of the symbiotic Mesorhizobium-chickpea relationship to phosphate deficiency relies on reprogramming of whole-plant metabolism. Proc. Natl. Acad. Sci. USA 2016, 113, 4610–4619. [Google Scholar] [CrossRef] [Green Version]
- Alamillo, J.M.; Díaz-Leal, J.L.; Sánchez-Moran, M.V.; Pineda, M. Molec-ular analysis of ureide accumulation under drought stress in Phaseolus v-ulgaris L. Plant Cell Env. 2010, 33, 1828–1837. [Google Scholar] [CrossRef]
- Díaz-Leal, J.L.; Gálvez-Valdivieso, G.; Fernández, J.; Pineda, M.; Alamillo, J.M. Developmental effects on ureide levels are mediated by tissue-specific regulation of allantoinase in Phaseolus vulgaris L. J. Exp. Bot. 2012, 63, 4095–4106. [Google Scholar] [CrossRef]
- Vance, C.; Graham, P.; Allan, D. Biological Nitrogen Fixation: Phosphorus—A Critical Future Need? Nitrogen Fixat. Mol. Crop. Product. 2002, 38, 509–514. [Google Scholar] [CrossRef]
- Zhang, Y.; Hongling, Q.I.; Fengxia, L.U.; Han, M.; Wang, P. Differences in Phosphorus Absorption and Utilization Efficiency of Soybean in Mature Period under Phosphorus Stress. Agric. Biotechnol. 2014, 3, 24–27. [Google Scholar]
- Xue, A.; Guo, X.; Zhu, Q.; Zhang, H.; Wang, H. Effect of Phosphorus Fertilization to P Uptake and Dry Matter Accumulation in Soybean with Different P Efficiencies. J. Integr. Agric. 2014, 13, 326–334. [Google Scholar] [CrossRef]
- Georgiev, G.I.; Tsvetkova, G.E. Changes in phosphate fractions growth r-ate, nodulation and nitrogn2 fixation of phosphorus starved soybean pla-nts. J. Plant Nutr. 2011, 34, 2055–2068. [Google Scholar] [CrossRef]
- Thuynsma, R.; Valentine, A.; Kleinert, A. Phosphorus deficiency affects the allocation of below-ground resources to combined cluster roots and nodules in Lupinusalbus. J. Plant Physiol. 2014, 171, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Lazali, M.; Drevon, J.J. The nodule conductance to O₂ diffusion increases with phytase activity in N₂-fixing Phaseolus vulgaris L. Plant Physiol. Biochem. 2014, 80, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Al-Niemi, T.S.; Kahn, M.L.; Mc De Rmott, T.R. Phosphorus uptake by bean nodules. Plant Soil 1998, 198, 71–78. [Google Scholar] [CrossRef]
- Burleigh, S.H.; Harrison, M.J. The down-regulation of Mt4-like genes by phosphate fertilization occurs systemically and involves phosphate translocation to the shoots. Plant Physiol. 1999, 119, 241–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wouterlood, M.; Lambers, H.; Veneklaas, E. Plant phosphorus status has a limited influence on the concentration of phosphorus-mobilising carboxylates in the rhizosphere of chickpea. Funct. Plant Biol. 2005, 32, 153–159. [Google Scholar] [CrossRef]
- Bonser, A.; Lynch, J.; Snapp, S. Effect of phosphorus deficiency on growth angle of basal roots in Phaseolus vulgaris. New Phytol. 1996, 132, 281–288. [Google Scholar] [CrossRef]
- Isidra-Arellano, M.C.; Pozas-Rodríguez, E.A.; RocíoReyero-Saavedra, M.; Arroyo-Canales, J.; Ferrer-Orgaz, S.; Socorro Sánchez-Correa, M.; Cardenas, L.; Covarrubias, A.A.; Valdés-López, O. Inhibition of legume nodulation by Pi deficiency is dependent on the autoregulation of nodulation (AON) pathway. Plant J. 2020, 103, 1125–1139. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Roots | Nodules | Shoot(%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| P+(%) | P−(%) | IP–(%) | P−/P+ | P+(%) | P−(%) | IP–(%) | P−/P+ | |||
| V4 | P1/0 | 0.07 ± 0.00 b | 0.07 ± 0.00 b | 85.71 | 1.00 | 0.20 ± 0.01 c** | 0.16 ± 0.01 c | 68.75 | 0.80 | 0.07 ± 0.01 b |
| P31/0 | 0.21 ± 0.01 a** | 0.13 ± 0.01 a | 0.62 | 0.32 ± 0.01 b** | 0.27 ± 0.01 b | 0.84 | 0.16 ± 0.01 a | |||
| 0.00 | 29.63 | |||||||||
| P61/0 | 0.24 ± 0.02 a** | 0.13 ± 0.01 a | 0.54 | 0.44 ± 0.01 a** | 0.35 ± 0.01 a | 0.80 | 0.18 ± 0.01 a | |||
| R1 | P1/0 | 0.07 ± 0.01 b | 0.07 ± 0.00 b | 57.14 | 1.00 | 0.16 ± 0.01 b | 0.16 ± 0.01 c | 75.00 | 1.00 | 0.09 ± 0.00 b |
| P31/0 | 0.20 ± 0.01 a** | 0.11 ± 0.01 a | 0.55 | 0.34 ± 0.01 a* | 0.28 ± 0.01 b | 0.82 | 0.21 ± 0.01 a | |||
| 9.09 | 25.00 | |||||||||
| P61/0 | 0.19 ± 0.01 a** | 0.12 ± 0.01 a | 0.63 | 0.36 ± 0.01 a | 0.35 ± 0.02 a | 0.97 | 0.22 ± 0.01 a | |||
| Treatments | Roots | Nodules | Shoots (g/Plant) | |||||
|---|---|---|---|---|---|---|---|---|
| P+(g/Plant) | P−(g/Plant) | P−/P+ | P+(g/Plant) | P−(g/Plant) | P−/P+ | |||
| V4 | P1/0 | 0.78 ± 0.03 c | 0.71 ± 0.01 b | 0.91 | 0.01 ± 0.000 c | 0.01 ± 0.001 b | 1.00 | 1.78 ± 0.11 c |
| P31/0 | 1.72 ± 0.05 b** | 0.84 ± 0.05 a | 0.49 | 0.18 ± 0.006 b** | 0.04 ± 0.001 a | 0.22 | 3.99 ± 0.52 b | |
| P61/0 | 1.87 ± 0.01 a** | 0.93 ± 0.03 a | 0.50 | 0.23 ± 0.011 a** | 0.05 ± 0.001 a | 0.22 | 5.39 ± 0.40 a | |
| R1 | P1/0 | 0.92 ± 0.04 c | 0.80 ± 0.03 b | 0.87 | 0.02 ± 0.003 c | 0.02 ± 0.003 b | 1.00 | 2.09 ± 0.04 b |
| P31/0 | 2.61 ± 0.01 b** | 1.07 ± 0.03 a | 0.41 | 0.22 ± 0.003 b** | 0.06 ± 0.003 a | 0.27 | 6.49 ± 0.71 a | |
| P61/0 | 2.73 ± 0.03 a** | 1.11 ± 0.04 a | 0.41 | 0.31 ± 0.012 a** | 0.07 ± 0.003 a | 0.23 | 7.20 ± 0.26 a | |
| Treatments | ARA | SNA | |||||||
|---|---|---|---|---|---|---|---|---|---|
| P+(C2H4 μmol−1h −1Plant−1) | P-(C2H4 μmol−1h −1Plant−1) | IP– | P−/P+ | P+(C2H4 μmol−1 g−1h−1) | P-(C2H4 μmol−1 g−1h−1) | IP– | P−/P+ | ||
| V4 | P1/0 | 0.25 ± 0.01 c | 0.24 ± 0.01 c | 383.33 | 0.96 | 25.76 ± 0.87 c | 24.96 ± 0.47 c | 16.11 | 0.97 |
| P31/0 | 5.97 ± 0.04 b** | 1.16 ± 0.06 b | 0.19 | 33.23 ± 0.31 b** | 28.98 ± 0.54 b | 0.87 | |||
| 36.21 | 10.46 | ||||||||
| P61/0 | 9.10 ± 0.35 a** | 1.58 ± 0.01 a | 0.17 | 39.69 ± 1.27 a** | 32.01 ± 0.17 a | 0.81 | |||
| R1 | P1/0 | 0.60 ± 0.01 c | 0.59 ± 0.02 c | 218.64 | 0.98 | 29.81 ± 0.49 c | 29.81 ± 1.18 c | 11.41 | 1.00 |
| P31/0 | 10.01 ± 0.17 b** | 1.88 ± 0.07 b | 0.19 | 46.54 ± 0.34 b** | 33.21 ± 0.82 b | 0.75 | |||
| 50.00 | 15.78 | ||||||||
| P61/0 | 16.08 ± 0.76 a** | 2.82 ± 0.16 a | 0.18 | 51.30 ± 0.90 a** | 38.45 ± 0.65 a | 0.75 | |||
| Treatments | Ureide Content (mg/g DW) | IP–(%) | P−/P+ | ||
|---|---|---|---|---|---|
| P+ | P− | ||||
| V4 | P1/0 | 1.02 ± 0.02 c | 0.99 ± 0.04 b | 118.18 | 0.97 |
| P31/0 | 3.41 ± 0.08 b** | 2.16 ± 0.08 a | 0.63 | ||
| 6.02 | |||||
| P61/0 | 3.68 ± 0.11 a** | 2.29 ± 0.05 a | 0.62 | ||
| R1 | P1/0 | 1.16 ± 0.01 b | 1.01 ± 0.01 b | 156.44 | 0.87 |
| P31/0 | 4.42 ± 0.09 a** | 2.59 ± 0.13 a | 0.59 | ||
| 9.27 | |||||
| P61/0 | 4.38 ± 0.03 a** | 2.83 ± 0.11 a | 0.65 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Wang, X.; Liang, Q.; Lyu, X.; Li, S.; Gong, Z.; Dong, S.; Yan, C.; Ma, C. Regulation of Phosphorus Supply on Nodulation and Nitrogen Fixation in Soybean Plants with Dual-Root Systems. Agronomy 2021, 11, 2354. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11112354
Li H, Wang X, Liang Q, Lyu X, Li S, Gong Z, Dong S, Yan C, Ma C. Regulation of Phosphorus Supply on Nodulation and Nitrogen Fixation in Soybean Plants with Dual-Root Systems. Agronomy. 2021; 11(11):2354. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11112354
Chicago/Turabian StyleLi, Hongyu, Xiangxiang Wang, Quanxi Liang, Xiaochen Lyu, Sha Li, Zhenping Gong, Shoukun Dong, Chao Yan, and Chunmei Ma. 2021. "Regulation of Phosphorus Supply on Nodulation and Nitrogen Fixation in Soybean Plants with Dual-Root Systems" Agronomy 11, no. 11: 2354. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11112354