
Does Leaf Waxiness Confound the Use of NDVI in the Assessment of Chlorophyll When Evaluating Genetic Diversity Panels of Wheat?


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Field Trials
2.3. Field Data Collection
2.4. Phenotypic Data Analysis
3. Results
3.1. The Response of Traits to Different Growing Environments
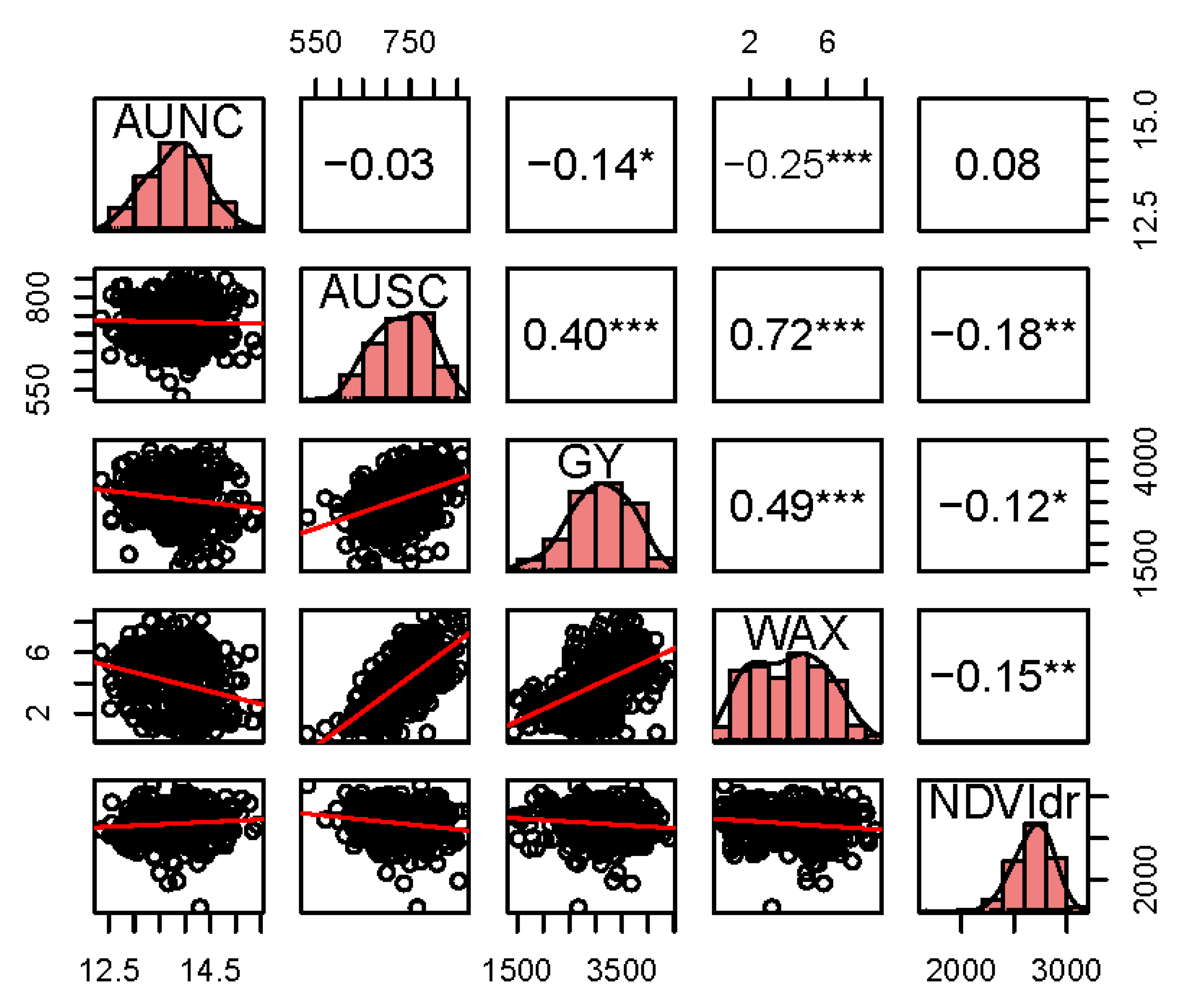
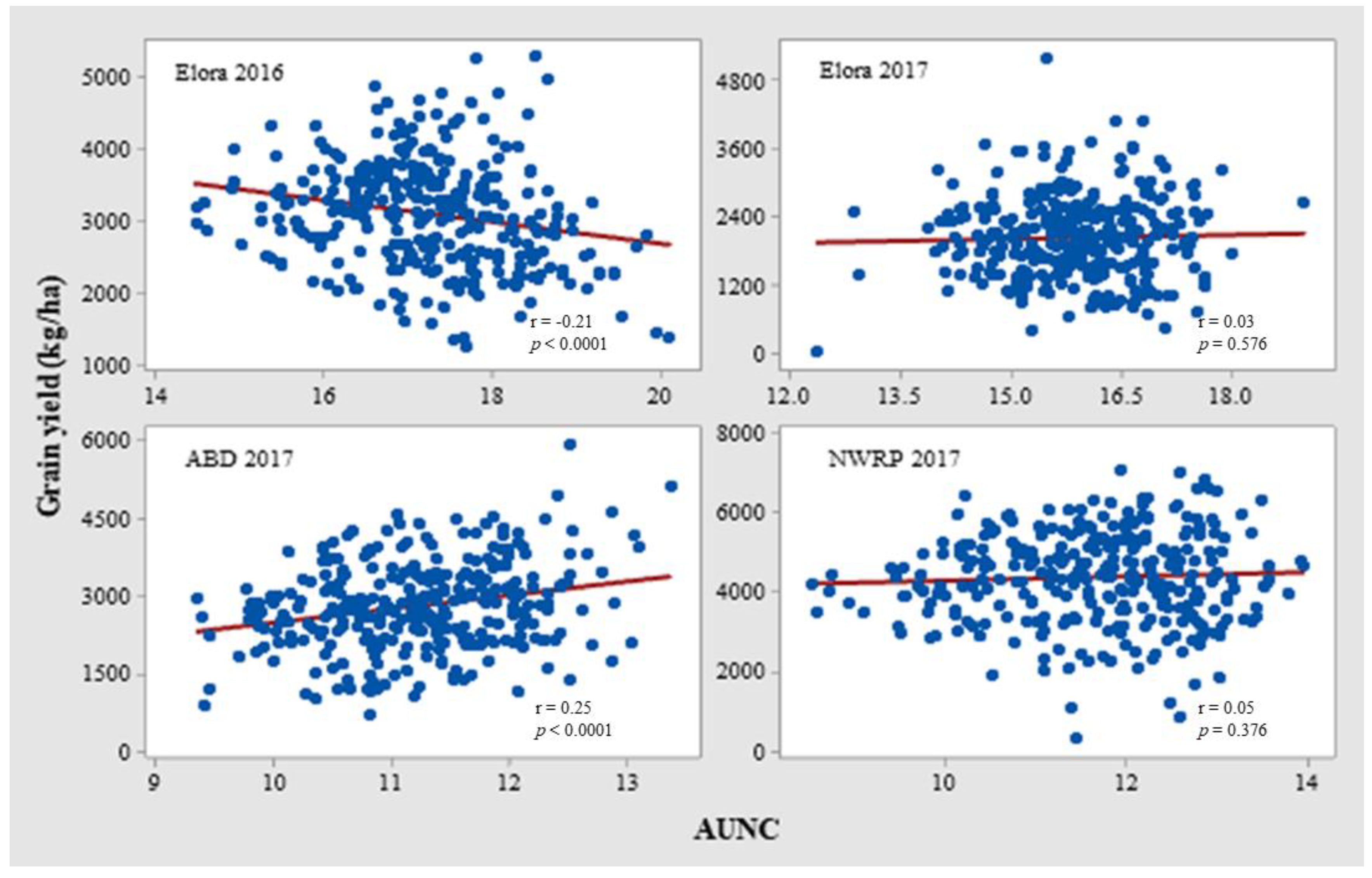
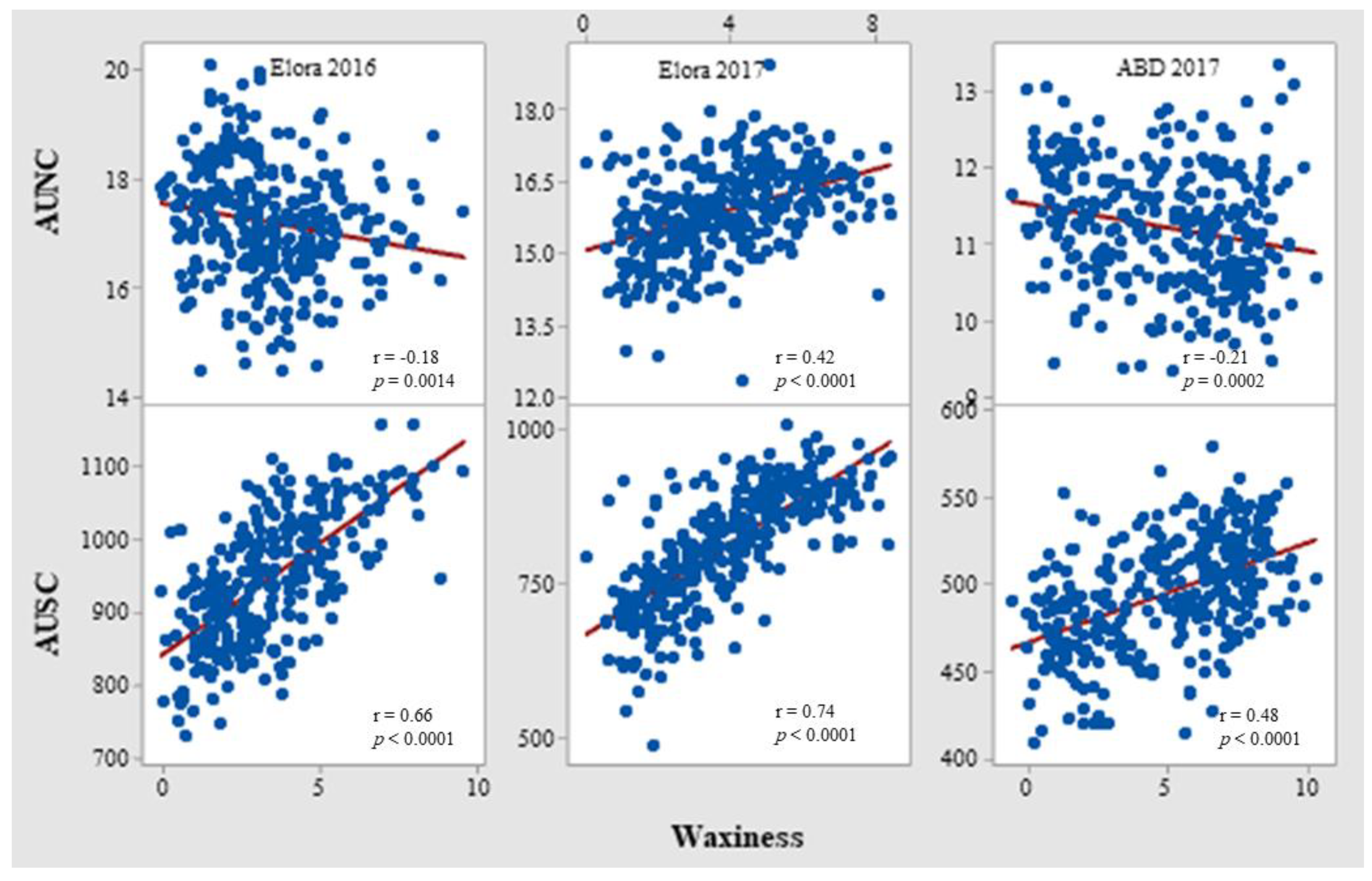
3.2. Correlation of Chlorophyll-Related Traits with Waxiness and Grain Yield
3.3. Principal Component Analysis
3.4. Within Trial Correlation Analysis
4. Discussion
4.1. Does Wax Interfere with NDVI?
4.2. UAV versus NDVI Measurements
4.3. Grain Yield and its Association with Physiological Traits
4.4. Should NDVI Be Used in Future Diversity Studies for Wheat Grain Yield?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shiferaw, B.; Smale, M.; Braun, H.-J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global food security. Food Secur. 2013, 5, 291–317. [Google Scholar] [CrossRef] [Green Version]
- Hickey, L.T.; Hafeez, A.; Robinson, H.; Jackson, S.A.; Leal-Bertioli, S.C.M.; Tester, M.; Gao, C.; Godwin, I.D.; Hayes, B.J.; Wulff, B.B.H. Breeding crops to feed 10 billion. Nat. Biotechnol. 2019, 37, 744–754. [Google Scholar] [CrossRef] [Green Version]
- Abhinandan, K.; Skori, L.; Stanic, M.; Hickerson, N.M.N.; Jamshed, M.; Samuel, M.A. Abiotic Stress Signaling in Wheat—An Inclusive Overview of Hormonal Interactions During Abiotic Stress Responses in Wheat. Front. Plant Sci. 2018, 9, 734. [Google Scholar] [CrossRef] [PubMed]
- Sarto, M.V.M.; Sarto, J.R.W.; Rampim, L.; Rosset, J.S.; Bassegio, D.; Da Costa, P.F.; Inagaki, A.M. State University of Mato Grosso do Sul—UEMS Wheat phenology and yield under drought: A review. Aust. J. Crop. Sci. 2017, 11, 941–946. [Google Scholar] [CrossRef]
- Daryanto, S.; Wang, L.; Jacinthe, P.-A. Global Synthesis of Drought Effects on Maize and Wheat Production. PLoS ONE 2016, 11, e0156362. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Min, D.; Yasir, T.A.; Hu, Y.-G. Evaluation of 14 morphological, yield-related and physiological traits as indicators of drought tolerance in Chinese winter bread wheat revealed by analysis of the membership function value of drought tolerance (MFVD). Field Crop. Res. 2012, 137, 195–201. [Google Scholar] [CrossRef]
- Barakat, M.N.; Saleh, M.; Al-Doss, A.; Moustafa, K.A.; Elshafei, A.A.; Al-Qurainy, F.H. Identification of new SSR markers linked to leaf chlorophyll content, flag leaf senescence and cell membrane stability traits in wheat under water stressed condition. Acta Biol. Hung. 2015, 66, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Khadka, K.; Raizada, M.N.; Navabi, A. Recent Progress in Germplasm Evaluation and Gene Mapping to Enable Breeding of Drought-Tolerant Wheat. Front. Plant Sci. 2020, 11, 1149. [Google Scholar] [CrossRef] [PubMed]
- Khadka, K.; Earl, H.J.; Raizada, M.N.; Navabi, A. A Physio-Morphological Trait-Based Approach for Breeding Drought Tolerant Wheat. Front. Plant Sci. 2020, 11, 715. [Google Scholar] [CrossRef] [PubMed]
- Simkin, A.J.; López-Calcagno, P.; A Raines, C. Feeding the world: Improving photosynthetic efficiency for sustainable crop production. J. Exp. Bot. 2019, 70, 1119–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, J.R. Improving Photosynthesis. Plant Physiol. 2013, 162, 1780–1793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinheiro, C.; Chaves, M.M. Photosynthesis and drought: Can we make metabolic connections from available data? J. Exp. Bot. 2010, 62, 869–882. [Google Scholar] [CrossRef] [Green Version]
- Zait, Y.; Schwartz, A. Climate-Related Limitations on Photosynthesis and Drought-Resistance Strategies of Ziziphus spina-christi. Front. For. Glob. Chang. 2018, 1, 3. [Google Scholar] [CrossRef]
- Rungrat, T.; Awlia, M.; Brown, T.; Cheng, R.; Sirault, X.; Fajkus, J.; Trtilek, M.; Furbank, B.; Badger, M.; Tester, M.A.; et al. Using Phenomic Analysis of Photosynthetic Function for Abiotic Stress Response Gene Discovery. Arab. Book 2016, 14, e0185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, E.M.; Tietz, S.; Froehlich, J.E. Photosystem I-LHCII megacomplexes respond to high light and aging in plants. Photosynth. Res. 2017, 136, 107–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roach, T.; Krieger-Liszkay, A.K. Regulation of Photosynthetic Electron Transport and Photoinhibition. Curr. Protein Pept. Sci. 2014, 15, 351–362. [Google Scholar] [CrossRef] [Green Version]
- Shah, S.H.; Houborg, R.; McCabe, M.F. Response of chlorophyll, carotenoid and SPAD-502 measurement to salinity and nu-trient stress in wheat (Triticum aestivum L.). Agronomy 2017, 7, 61. [Google Scholar] [CrossRef] [Green Version]
- Rosyara, U.R.; Khadka, K.; Subedi, S.; Sharma, R.C.; Duveiller, E. Heritability of stay green traits and association with spot blotch resistance in three spring wheat populations. J. Genet. Breed. 2007, 61, 75–82. [Google Scholar]
- Rosyara, U.R.; Khadka, K.; Subedi, S.; Sharma, R.C.; Duveiller, E. Field resistance to spot blotch is not associated with unde-sirable physio-morphological traits in three spring wheat populations. J. Plant Pathol. 2009, 91, 113–122. [Google Scholar]
- Kira, O.; Linker, R.; Gitelson, A. Non-destructive estimation of foliar chlorophyll and carotenoid contents: Focus on informative spectral bands. Int. J. Appl. Earth Obs. Geoinf. 2015, 38, 251–260. [Google Scholar] [CrossRef]
- Guendouz, A.; Maamari, K. Grain-filling, chlorophyll content in relation with grain yield component of durum wheat in a Mediterranean environment. Afr. Crop Sci. J. 2012, 20, 31–37. [Google Scholar]
- Earl, H.J.; Davis, R.F. Effect of drought stress on leaf and whole canopy radiation use efficiency and yield of maize. Agron. J. 2003, 95, 688–696. [Google Scholar] [CrossRef]
- Rosyara, U.R.; Subedi, S.; Duveiller, E.; Sharma, R.C. Photochemical Efficiency and SPAD Value as Indirect Selection Criteria for Combined Selection of Spot Blotch and Terminal Heat Stress in Wheat. J. Phytopathol. 2010, 158, 813–821. [Google Scholar] [CrossRef]
- Guan, S.; Fukami, K.; Matsunaka, H.; Okami, M.; Tanaka, R.; Nakano, H.; Sakai, T.; Nakano, K.; Ohdan, H.; Takahashi, K. Assessing Correlation of High-Resolution NDVI with Fertilizer Application Level and Yield of Rice and Wheat Crops using Small UAVs. Remote Sens. 2019, 11, 112. [Google Scholar] [CrossRef] [Green Version]
- Condorelli, G.E.; Maccaferri, M.; Newcomb, M.; Andrade-sanchez, P.; White, J.W.; French, A.N.; Sciara, G.; Ward, R.; Tuberosa, R. Comparative aerial and ground based high throughput phenotyping for the genetic dissection of NDVI as a proxy for drought adaptive traits in durum wheat. Front. Plant Sci. 2018, 9, 893. [Google Scholar] [CrossRef] [PubMed]
- Sid’Ko, A.; Botvich, I.; Pisman, T.; Shevyrnogov, A. Estimation of chlorophyll content and yield of wheat crops from reflectance spectra obtained by ground-based remote measurements. Field Crop. Res. 2017, 207, 24–29. [Google Scholar] [CrossRef]
- Lopresti, M.F.; Di Bella, C.M.; Degioanni, A.J. Relationship between MODIS-NDVI data and wheat yield: A case study in Northern Buenos Aires province, Argentina. Inf. Process. Agric. 2015, 2, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Gracia-Romero, A.; Kefauver, S.C.; Vergara-Díaz, O.; Zaman-Allah, M.A.; Prasanna, B.M.; Cairns, J.E.; Araus, J.L. Compar-ative performance of ground vs. Aerially assessed rgb and multispectral indices for early-growth evaluation of maize performance under phosphorus fertilization. Front. Plant Sci. 2017, 8, 2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, G.F.C.; Gonçalves, A.C.A.; da Silva Junior, C.A.; Nanni, M.R.; Facco, C.U.; Cezar, E.; da Silva, A.A. NDVI response to water stress in different phenological stages in culture bean. J. Agron. 2016, 15, 1–10. [Google Scholar]
- Fernández, E.; Gorchs, G.; Serrano, L. Use of consumer-grade cameras to assess wheat N status and grain yield. PLoS ONE 2019, 14, e0211889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, H.; Kovalchuk, N.; Langridge, P.; Tricker, P.J.; Lopato, S.; Borisjuk, N. The impact of drought on wheat leaf cuticle properties. BMC Plant Biol. 2017, 17, 85. [Google Scholar] [CrossRef] [Green Version]
- Cossani, C.M.; Reynolds, M.P. Physiological Traits for Improving Heat Tolerance in Wheat. Plant Physiol. 2012, 160, 1710–1718. [Google Scholar] [CrossRef] [Green Version]
- Buschhaus, C.; Jetter, R. Composition and Physiological Function of the Wax Layers Coating Arabidopsis Leaves: β-Amyrin Negatively Affects the Intracuticular Water Barrier. Plant Physiol. 2012, 160, 1120–1129. [Google Scholar] [CrossRef] [Green Version]
- Huggins, T.D.; Mohammed, S.; Sengodon, P.; Ibrahim, A.M.H.; Tilley, M.; Hays, D.B. Changes in leaf epicuticular wax load and its effect on leaf temperature and physiological traits in wheat cultivars (Triticum aestivum L.) exposed to high temperatures during anthesis. J. Agron. Crop. Sci. 2017, 204, 49–61. [Google Scholar] [CrossRef]
- Jäger, K.; Fábián, A.; Eitel, G.; Szabó, L.; Deák, C.; Barnabäs, B.; Papp, I. A morpho-physiological approach differentiates bread wheat cultivars of contrasting tolerance under cyclic water stress. J. Plant Physiol. 2014, 171, 1256–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammed, S.; Huggins, T.D.; Beecher, F.; Chick, C.; Sengodon, P.; Mondal, S.; Paudel, A.; Ibrahim, A.M.; Tilley, M.; Hays, D.B. The Role of Leaf Epicuticular Wax in the Adaptation of Wheat (Triticum aestivum L.) to High Temperatures and Moisture Deficit Conditions. Crop Sci. 2018, 58, 679–689. [Google Scholar] [CrossRef]
- Pereira, J.F.; Da Cunha, G.R.; Moresco, E.R. Improved drought tolerance in wheat is required to unlock the production potential of the Brazilian Cerrado. Crop Breed. Appl. Biotechnol. 2019, 19, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, T.; Griffiths, D.W. The effects of stress on plant cuticular waxes. New Phytol. 2006, 171, 469–499. [Google Scholar] [CrossRef] [PubMed]
- Bowne, J.B.; Erwin, T.A.; Juttner, J.; Schnurbusch, T.; Langridge, P.; Bacic, A.; Roessner, U. Drought Responses of Leaf Tissues from Wheat Cultivars of Differing Drought Tolerance at the Metabolite Level. Mol. Plant 2012, 5, 418–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willick, I.R.; Lahlali, R.; Vijayan, P.; Muir, D.; Karunakaran, C.; Tanino, K.K. Wheat flag leaf epicuticular wax morphology and composition in response to moderate drought stress are revealed by SEM, FTIR-ATR and synchrotron X-ray spectroscopy. Physiol. Plant. 2018, 162, 316–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, D.; Zhang, X.; Lu, X.; Chen, G.; Chen, Z.-H. Molecular and Evolutionary Mechanisms of Cuticular Wax for Plant Drought Tolerance. Front. Plant Sci. 2017, 8, 621. [Google Scholar] [CrossRef] [PubMed]
- Khadka, K.; Torkamaneh, D.; Kaviani, M.; Belzile, F.; Raizada, M.N.; Navabi, A. Population structure of Nepali spring wheat (Triticum aestivum L.) germplasm. BMC Plant Biol. 2020, 20, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mo, A.D. Statistical Information on Nepalese Agriculture; Government of Nepal, Ministry of Agricultural Development, Agribusiness Promotion and Statistics Division, Agri Statistics Section: Kathmandu, Nepal, 2017.
- Gurung, D.B. Improving wheat productivity in Asia. In Proceedings of the Regional Consultation on Improving Wheat Productivity in Asia, Bangkok, Thailand, 26–27 April 2012; Paroda, R., Dasgupta, S., Mal, B., Singh, S.S., Jat, M.L., Singh, G., Eds.; FAO; APAARI; CIMMYT; ICARDA; JIRCAS: Bangkok, Thailand, 2013; pp. 112–123. [Google Scholar]
- DHM-MoSTE Nepal. Agroclimatic Atlas of Nepal; Department of Hydrology and Meteorology, Ministory of Science and Teconology, and Environment, Government of Nepal: Kathmandu, Nepal, 2013.
- Shrestha, N.; Raes, D.; Vanuytrecht, E.; Sah, S.K. Cereal yield stabilization in Terai (Nepal) by water and soil fertility man-agement modeling. Agric. Water Manag. 2013, 122, 53–62. [Google Scholar] [CrossRef]
- Depauw, R.M.; Knox, R.E.; McCaig, T.N.; Clarke, F.R.; Clarke, J.M. Carberry hard red spring wheat. Can. J. Plant Sci. 2011, 91, 529–534. [Google Scholar] [CrossRef]
- Patterson, H.D.; Williams, E.R. A new class of resolvable incomplete block designs. Biometrika 1976, 63, 83–92. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef] [Green Version]
- Haghighattalab, A.; Pérez, L.G.; Mondal, S.; Singh, D.; Schinstock, D.; Rutkoski, J.; Ortiz-Monasterio, I.; Singh, R.P.; Goodin, D.; Poland, J. Application of unmanned aerial systems for high throughput phenotyping of large wheat breeding nurseries. Plant Methods 2016, 12, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, A.; Pietragalla, J. Chapter 19: Crop morphological traits. In Physiological Breeding II: A Field Guide to Wheat Breeding; Pask, A., Pietragalla, J., Mullan, D., Reynolds, M., Eds.; CIMMYT: Mexico City, Mexico, 2012; pp. 106–112. [Google Scholar]
- Guo, J.; Xu, W.; Yu, X.; Shen, H.; Li, H.; Cheng, D.; Liu, A.; Liu, J.; Liu, C.; Zhao, S.; et al. Cuticular wax accumulation is as-sociated with drought tolerance in wheat near-isogenic lines. Front. Plant Sci. 2016, 7, 1809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medeiros, C.; Falcão, H.; Almeida-Cortez, J.; Santos, D.; Oliveira, A.; Santos, M. Leaf epicuticular wax content changes under different rainfall regimes, and its removal affects the leaf chlorophyll content and gas exchanges of Aspidosperma pyrifolium in a seasonally dry tropical forest. S. Afr. J. Bot. 2017, 111, 267–274. [Google Scholar] [CrossRef]
- Gates, D.M.; Keegan, H.J.; Schleter, J.C.; Weidner, V.R. Spectral Properties of Plants. Appl. Opt. 1965, 4, 11–20. [Google Scholar] [CrossRef]
- Ballester, C.; Brinkhoff, J.; Quayle, W.C.; Hornbuckle, J. Monitoring the Effects of Water Stress in Cotton using the Green Red Vegetation Index and Red Edge Ratio. Remote Sens. 2019, 11, 873. [Google Scholar] [CrossRef] [Green Version]
- Walter-Shea, E.A.; Blad, B.L.; Hays, C.J.; Mesarch, M.A.; Deering, D.W.; Middleton, E.M. Biophysical properties affecting vegetative canopy reflectance and absorbed photsynthetically active radiation at the FIFE site. J. Geophys. Res. 1992, 97, 925–934. [Google Scholar]
- Holmes, M.G.; Keiller, D.R. Effects of pubescence and waxes on the reflectance of leaves in the ultraviolet and photosynthetic wavebands: A comparison of a range of species. Plant Cell Environ. 2002, 25, 85–93. [Google Scholar] [CrossRef]
- Mohammed, G.H.; Noland, T.L.; Irving, P.H.; Sampson, P.H.; Zarco-Tejada, P.J.; Miller, J.R. Natural and Stress-Induced Effects on Leaf Spectral Reflectance in Ontario Species; Ontario Forest Research Institute, Ontario Ministry of Natural Resources: Sault Ste. Marie, ON, Canada, 2000; Volume 156, ISBN 0777893878. [Google Scholar]
- Xue, J.; Su, B. Significant Remote Sensing Vegetation Indices: A Review of Developments and Applications. J. Sens. 2017, 2017, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Morgounov, A.; Gummadov, N.; Belen, S.; Kaya, Y.; Keser, M.; Mursalova, J. Association of digital photo parameters and NDVI with winter wheat grain yield in variable environments. Turk. J. Agric. For. 2014, 38, 624–632. [Google Scholar] [CrossRef]
- Araus, J.L. Integrative physiological criteria associated with yield potential. In Increasing Yield Potential in Wheat: Breaking the Barriers; Reynolds, M.P., Rajaram, S., McNab, A., Eds.; International Maize and Wheat Improvement Center (CIMMYT): Mexico City, Mexico, 1996; pp. 150–166. [Google Scholar]
- Vergara-Díaz, O.; Chairi, F.; Vicente, R.; A Fernandez-Gallego, J.; Nieto-Taladriz, M.T.; Aparicio, N.; Kefauver, S.C.; Araus, J.L. Leaf dorsoventrality as a paramount factor determining spectral performance in field-grown wheat under contrasting water regimes. J. Exp. Bot. 2018, 69, 3081–3094. [Google Scholar] [CrossRef] [Green Version]
- Samborski, S.M.; Gozdowski, D.; Walsh, O.S.; Lamb, D.W.; Stępień, M.; Gacek, E.S.; Drzazga, T. Winter wheat genotype effect on canopy reflectance: Implications for using NDVI for in-season nitrogen topdressing recommendations. Agron. J. 2015, 107, 2097–2106. [Google Scholar] [CrossRef]
- Sreeman, S.M.; Vijayaraghavareddy, P.; Sreevathsa, R.; Rajendrareddy, S.; Arakesh, S.; Bharti, P.; Dharmappa, P.; Soola-nayakanahally, R. Introgression of physiological traits for a comprehensive improvement of drought adaptation in crop plants. Front. Chem. 2018, 6, 92. [Google Scholar] [CrossRef] [Green Version]
- Perdeaux, S. Climate Change Adaptation ‘Stories’ of Ontario: A Collection of Five Adaptation Initiatives from across Ontario; Ontario Centre for Climate Impacts and Adaptation Resources (OCCIAR); Laurentian University: Greater Sudbury, ON, Canada, 2017. [Google Scholar]
- Foley, S.; Rivard, B.; Sanchezazofeifa, G.; Calvo, J. Foliar spectral properties following leaf clipping and implications for handling techniques. Remote Sens. Environ. 2006, 103, 265–275. [Google Scholar] [CrossRef]
- Khadka, K. Genotypic and Phenotypic Analysis of a Nepali Spring Wheat (Triticum aestivum L.) Population. Ph.D. Thesis, University of Guelph, Guelph, ON, Canada, 2020. [Google Scholar]
- Kyratzis, A.C.; Skarlatos, D.P.; Menexes, G.C.; Vamvakousis, V.F.; Katsiotis, A. Assessment of Vegetation Indices Derived by UAV Imagery for Durum Wheat Phenotyping under a Water Limited and Heat Stressed Mediterranean Environment. Front. Plant Sci. 2017, 8, 1114. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Thomasson, J.A.; Murray, S.C.; Pugh, N.A.; Rooney, W.L.; Shafian, S.; Rajan, N.; Rouze, G.; Morgan, C.L.S.; Neely, H.L.; et al. Unmanned Aerial Vehicles for High-Throughput Phenotyping and Agronomic Research. PLoS ONE 2016, 11, e0159781. [Google Scholar] [CrossRef] [Green Version]
- Tattaris, M.; Reynolds, M.P.; Chapman, S.C. A direct comparison of remote sensing approaches for high-throughput pheno-typing in plant breeding. Front. Plant Sci. 2016, 7, 1131. [Google Scholar] [CrossRef]
- Hollington, P. Technological breakthroughs in creening/breeding wheat varieties for salt toleracne. In Proceedings of the Na-tional Conference on “Salinity Management in Agriculture”, Karnal, India, 2 December 1998; p. 36. [Google Scholar]
- Zhang, Y.; Wang, Z.; Fan, Z.; Li, J.; Gao, X.; Zhang, H.; Zhao, Q.; Wang, Z.; Liu, Z. Phenotyping and evaluation of CIMMYT WPHYSGP nursery lines and local wheat varieties under two irrigation regimes. Breed. Sci. 2019, 69, 55–67. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.R.; Haque, K.M.S.; Akter, N.; Karim, M.A. Leaf chlorophyll dynamics in wheat based on SPAD meter reading and its relationship with grain yield. Sci. Agric. 2014, 4, 4–9. [Google Scholar]
- Sultana, S.R.; Ali, A.; Ahmad, A.; Mubeen, M.; Zia-Ul-Haq, M.; Ahmad, S.; Ercisli, S.; Jaafar, H.Z.E. Normalized difference vegetation index as a tool for wheat yield estimation: A case study from Faisalabad, Pakistan. Sci. World J. 2014, 2014, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebetzke, G.J.; Jimenez-Berni, J.A.; Bovill, W.D.; Deery, D.M.; James, R.A. High-throughput phenotyping technologies allow accurate selection of stay-green. J. Exp. Bot. 2016, 67, 4919–4924. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.; Wang, D.; Zhou, J.; Du, Y.; Luo, M.; Zhang, Y.; Guo, W. Remotely Assessing Fraction of Photosynthetically Active Radiation (FPAR) for Wheat Canopies Based on Hyperspectral Vegetation Indexes. Front. Plant Sci. 2018, 9, 776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Wang, J.; Qin, H.; Wei, Z.; Hang, L.; Zhang, P.; Reynolds, M.; Wang, D. Assessment of the individual and combined effects of Rht8 and Ppd-D1a on plant height, time to heading and yield traits in common wheat. Crop. J. 2019, 7, 845–856. [Google Scholar] [CrossRef]
- Rutkoski, J.; Poland, J.A.; Mondal, S.; Autrique, E.; Pérez, L.G.; Crossa, J.; Reynolds, M.; Singh, R. Canopy Temperature and Vegetation Indices from High-Throughput Phenotyping Improve Accuracy of Pedigree and Genomic Selection for Grain Yield in Wheat. G3 Genes Genomes Genet. 2016, 6, 2799–2808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, M.S.; Reynolds, M.P. Stay-green in spring wheat can be determined by spectral reflectance measurements (normalized difference vegetation index) independently from phenology. J. Exp. Bot. 2012, 63, 3789–3798. [Google Scholar] [CrossRef] [Green Version]
- Prasad, B.; Babar, M.A.; Carver, B.F.; Raun, W.R.; Klatt, A.R. Association of biomass production and canopy spectral reflec-tance indices in winter wheat. Can. J. Plant Sci. 2009, 89, 485–496. [Google Scholar] [CrossRef]
- Thapa, S.; Rudd, J.C.; Xue, Q.; Bhandari, M.; Reddy, S.K.; Jessup, K.E.; Liu, S.; Devkota, R.N.; Baker, J.; Baker, S. Use of NDVI for characterizing winter wheat response to water stress in a semi-arid environment. J. Crop. Improv. 2019, 33, 633–648. [Google Scholar] [CrossRef]
- Christopher, J.T.; Christopher, M.J.; Borrell, A.K.; Fletcher, S.; Chenu, K. Stay-green traits to improve wheat adaptation in well-watered and water-limited environments. J. Exp. Bot. 2016, 67, 5159–5172. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.; Jiang, J.; Gao, Y.; Krienke, B.; Wang, M.; Zhong, K.; Cao, Q.; Tian, Y.; Zhu, Y.; Cao, W.; et al. Wheat Growth Monitoring and Yield Estimation based on Multi-Rotor Unmanned Aerial Vehicle. Remote Sens. 2020, 12, 508. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | |||||
|---|---|---|---|---|---|
| Location | AUNC | AUSC | GY | WAX | |
| Elora 2016 | AUSC | 0.06 | |||
| GY | −0.21 *** | 0.33 *** | |||
| WAX | −0.18 ** | 0.66 *** | 0.34 *** | ||
| Elora 2017 | AUSC | 0.57 *** | |||
| GY | 0.03 | 0.06 | |||
| WAX | 0.42 *** | 0.74 *** | 0.12 * | ||
| NDVIdr | −0.18 ** | −0.31 *** | −0.03 | −0.21 *** | |
| ABD 2017 | AUSC | −0.14 * | |||
| GY | 0.25 *** | 0.20 ** | |||
| WAX | −0.21 ** | 0.48 *** | 0.44 *** | ||
| NWRP 2017 | GY | 0.05 | |||
| Traits | Groups Based on Waxiness Scores | Groups Based on Breeding History | ||||
|---|---|---|---|---|---|---|
| Group 1—Low wax (N = 88) GY (kg/ha) (mean ± SEM) = 2720.1 ± 69.97 WAX (mean ± SE) = 1.2 ± 0.05 | Landraces (N = 166) GY (kg/ha) (mean ± SEM) = 2705.7 ± 48.43 WAX (mean ± SE) = 2.3 ± 0.10 | |||||
| AUNC | AUSC | GY | AUNC | AUSC | GY | |
| AUSC | 0.23 * | 0.15 | ||||
| GY | −0.16 | 0.19 | −0.24 * | 0.11 | ||
| WAX | 0.19 | 0.22 * | −0.23 * | −0.23 * | 0.47 *** | 0.11 |
| Group 2—Medium wax (N = 100) GY (kg/ha) (mean ± SEM) = 3037.3 ± 78.38 WAX (mean ± SE) = 2.9 ± 0.04 | Commercial varieties (N = 34) GY (kg/ha) (mean ± SEM) = 3608.3 ± 108.12 WAX (mean ± SE) = 2.9 ± 0.19 | |||||
| AUSC | 0.10 | 0.61 *** | ||||
| GY | −0.34 ** | 0.11 | 0.48 * | 0.14 | ||
| WAX | −0.22 * | 0.35 ** | 0.22 * | 0.18 | 0.38 * (check) | 0.12 |
| Group 3—Highest wax (N = 130) GY (kg/ha) (mean ± SEM) = 3413.3 ± 54.53 WAX (mean ± SE) = 5.2 ± 0.11 | CIMMYT lines (N = 115) GY (kg/ha) (mean ± SEM) = 3474.7 ± 53.89 WAX (mean ± SE) = 4.9 ± 0.15 | |||||
| AUSC | 0.45 *** | 0.30 ** | ||||
| GY | 0.14 | 0.15 | 0.09 | 0.06 | ||
| WAX | 0.17 | 0.46 *** | 0.03 | 0.18 | 0.45 *** | 0.00 |
| Traits | Groups Based on Waxiness Scores | Groups Based on Breeding History | ||||||
|---|---|---|---|---|---|---|---|---|
| Group 1—Low wax (N = 69), GY (kg/ha) (mean ± SEM) = 1917.9 ± 87.31 WAX (mean ± SE) = 1.3 ± 0.05 | Landraces (N = 166) GY (kg/ha) (mean ± SEM) = 1987.3 ± 53.07 WAX (mean ± SE) = 2.9 ± 0.13 | |||||||
| AUNC | AUSC | GY | WAX | AUNC | AUSC | GY | WAX | |
| AUSC | 0.50 *** | 0.51 *** | ||||||
| GY | −0.13 | −0.24 | −0.03 | −0.01 | ||||
| WAX | −0.14 | 0.04 | −0.28 * | −0.30 *** | 0.67 *** | 0.03 | ||
| NDVIdr | −0.30* | −0.37 * | 0.16 | 0.21 | 0.19 * | −0.42 *** | −0.01 | −0.25 ** |
| Group 2—Medium wax (N = 105) GY (kg/ha) (mean ± SEM) = 1977.6 ± 61.45 WAX (mean ± SE) = 2.9± 0.05 | Commercial varieties (N = 34) GY (kg/ha) (mean ± SEM) = 2240.3 ± 120.29 WAX (mean ± SE) = 4.1 ± 0.25 | |||||||
| AUSC | 0.41 *** | 0.55 ** | ||||||
| GY | −0.12 | −0.10 | − 0.10 | −0.09 | ||||
| WAX | 0.17 | 0.43 *** | 0.01 | 0.22 | 0.74 *** | 0.24 | ||
| NDVIdr | 0.07 | −0.24* | −0.09 | −0.18 | −0.13 | −0.10 | 0.10 | −0.11 |
| Group 3—Highest wax (N = 144) GY (kg/ha) (mean ± SEM) = 2126.8 ± 63.58 WAX (mean ± SE) = 5.6 ± 0.09 | CIMMYT lines (N = 115) GY (kg/ha) (mean ± SEM) = 3474.7 ± 53.89 WAX (mean ± SE) = 5.1 ± 0.15 | |||||||
| AUSC | 0.35 *** | 0.33 ** | ||||||
| GY | 0.09 | 0.12 | 0.08 | 0.15 | ||||
| WAX | 0.10 | 0.39 *** | 0.08 | 0.28 * | 0.64 *** | 0.20 * | ||
| NDVIdr | −0.16 | −0.17* | −0.01 | −0.02 | −0.16 | −0.20 * | −0.06 | −0.17 |
| Elora 2016 | ||||||
| Traits | Groups based on waxiness scores | Groups based on breeding history | ||||
| Group 1—Low wax (N = 88) | Landraces (N = 166) | |||||
| AUNC | AUSC | * Discrepancy | AUNC | AUSC | * Discrepancy | |
| WAX | 0.19 | 0.22 * | 0.03 | −0.23 * | 0.47 *** | 0.70 |
| Group 2—Medium wax (N = 100) | Commercial varieties (N = 34) | |||||
| WAX | −0.22 * | 0.35 ** | 0.57 | 0.18 | 0.38 * | 0.20 |
| Group 3—Highest wax (N = 130) | CIMMYT lines (N = 115) | |||||
| WAX | 0.17 | 0.46 *** | 0.29 | 0.18 | 0.45 *** | 0.27 |
| Elora 2017 | ||||||
| Traits | Groups based on waxiness scores | Groups based on breeding history | ||||
| Group 1 (N = 69) | Landraces (N = 166) | |||||
| AUNC | AUSC | Discrepancy | AUNC | AUSC | Discrepancy | |
| WAX | −0.14 | 0.04 | 0.18 | −0.30 *** | 0.67 *** | 0.97 |
| Group 2 (N = 105) | Commercial varieties (N = 34) | |||||
| WAX | 0.17 | 0.43 *** | 0.26 | 0.22 | 0.74 *** | 0.52 |
| Group 3 (N = 144) | CIMMYT lines (N = 115) | |||||
| WAX | 0.10 | 0.39 *** | 0.29 | 0.28 * | 0.64 *** | 0.36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khadka, K.; Burt, A.J.; Earl, H.J.; Raizada, M.N.; Navabi, A. Does Leaf Waxiness Confound the Use of NDVI in the Assessment of Chlorophyll When Evaluating Genetic Diversity Panels of Wheat? Agronomy 2021, 11, 486. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030486
Khadka K, Burt AJ, Earl HJ, Raizada MN, Navabi A. Does Leaf Waxiness Confound the Use of NDVI in the Assessment of Chlorophyll When Evaluating Genetic Diversity Panels of Wheat? Agronomy. 2021; 11(3):486. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030486
Chicago/Turabian StyleKhadka, Kamal, Andrew J. Burt, Hugh J. Earl, Manish N. Raizada, and Alireza Navabi. 2021. "Does Leaf Waxiness Confound the Use of NDVI in the Assessment of Chlorophyll When Evaluating Genetic Diversity Panels of Wheat?" Agronomy 11, no. 3: 486. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030486