
Incidence of GLMD-Like Symptoms on Grapevines Naturally Infected by Grapevine Pinot gris virus, Boron Content and Gene Expression Analysis of Boron Metabolism Genes

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Sampling and Collection of Plant Samples
2.2. Sampling and Collection of Soil Samples
2.3. RNA Extraction and RT-PCR Detection of GPGV and Sequencing of MP/CP and RdRp PCR Amplicons
2.4. Phylogenetic Analysis
2.5. GPGV Relative Quantification
2.6. Gene Expression Analysis
2.7. Analysis of Boron Content in Soil and Grapevine Shoots and Leaves
2.8. Statistical Analysis
3. Results
3.1. Symptomatology and GPGV Detection
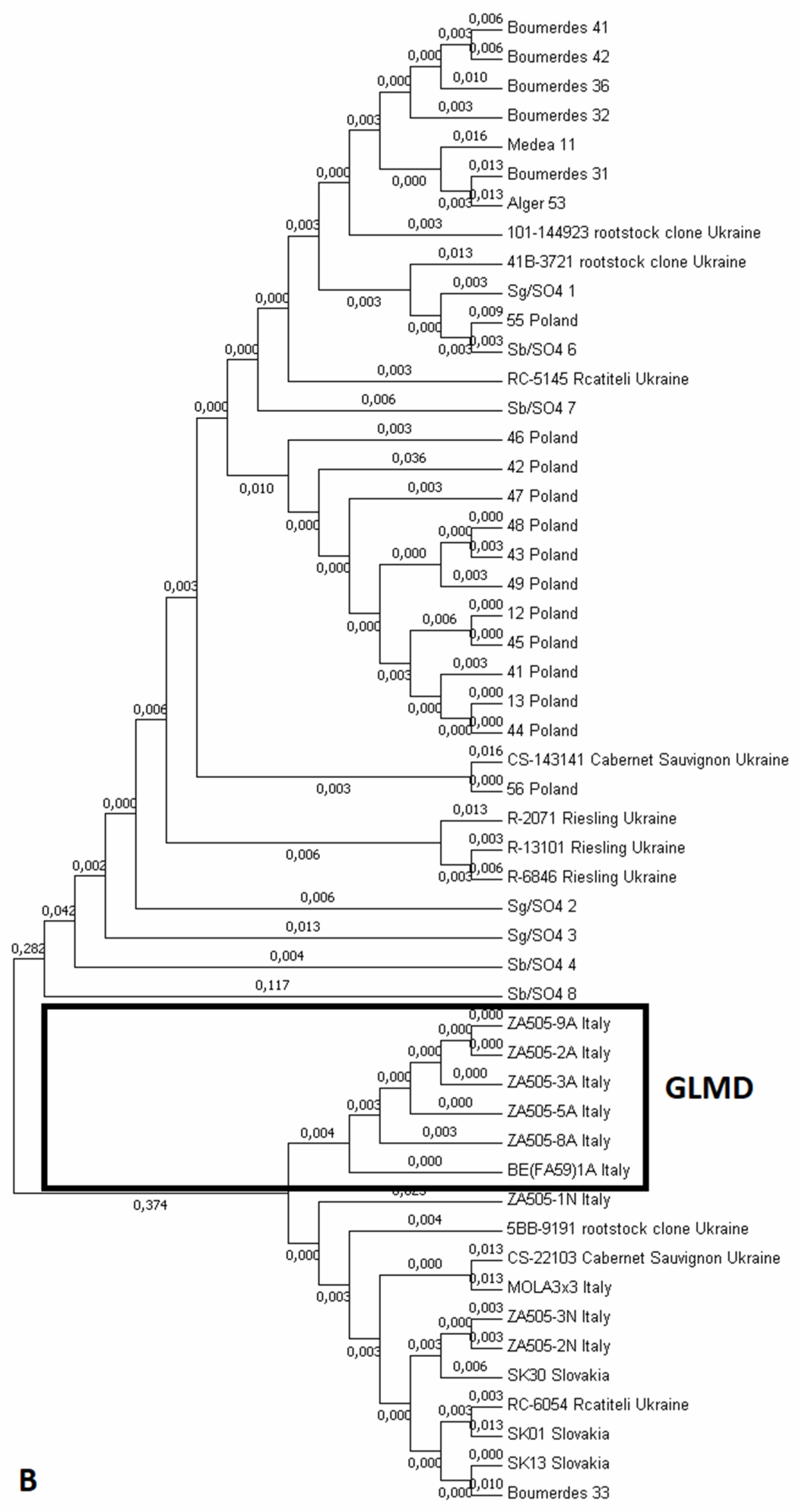
3.2. GPGV Phylogenetic Analysis
3.3. GPGV Relative Quantification
3.4. Boron Content
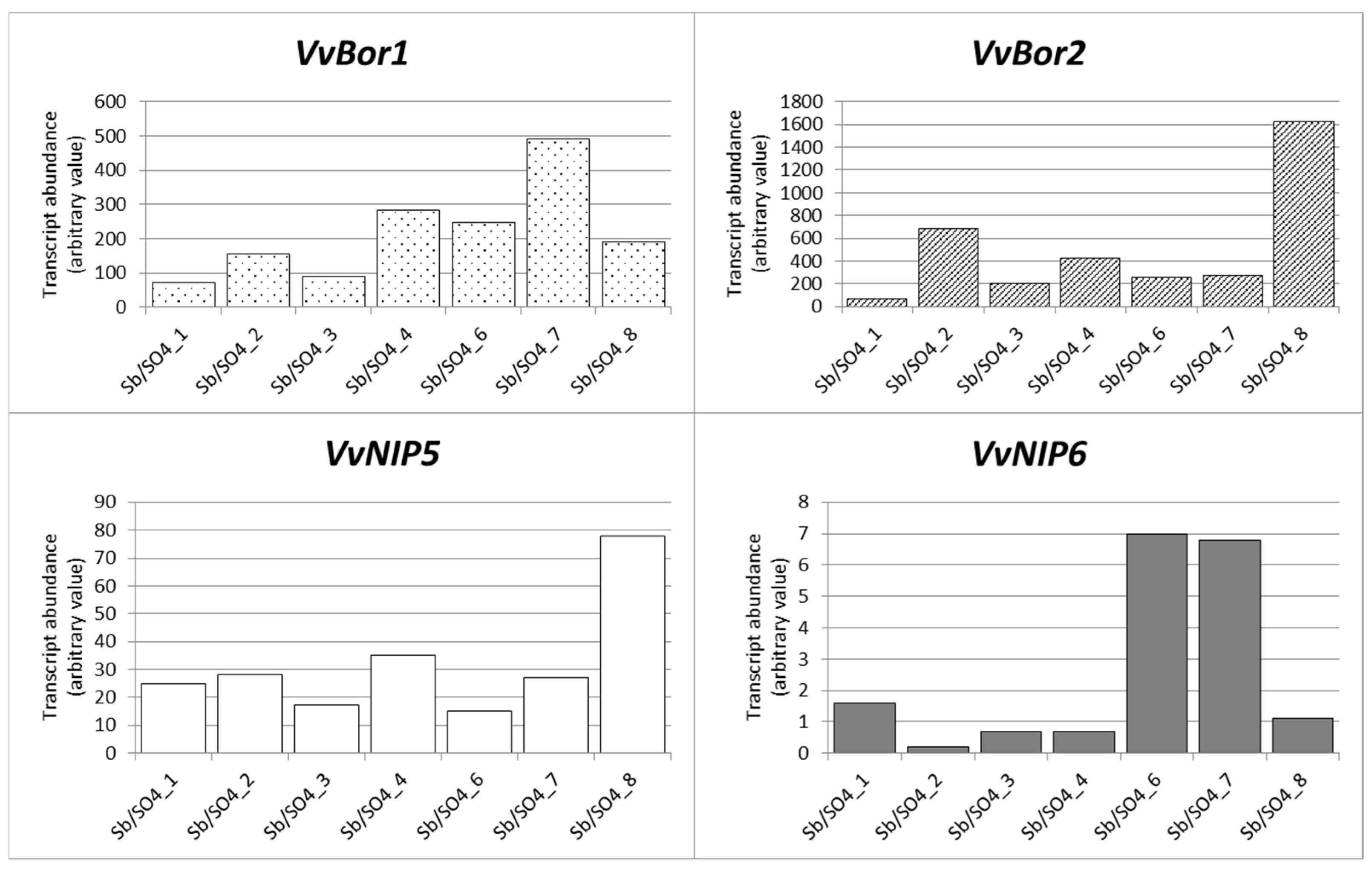
3.5. Gene Expression Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Giampetruzzi, A.; Roumi, V.; Roberto, R.; Malossini, U.; Yoshikawa, N.; La Notte, P.; Terlizzi, F.; Credi, R.; Saldarelli, P. A new grapevine virus discovered by deep sequencing of virus- and viroid-derived small RNAs in Cv Pinot gris. Virus Res. 2012, 163, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Saldarelli, P.; Giampetruzzi, A.; Morelli, M.; Malossini, U.; Pirolo, C.; Bianchedi, P.; Gualandri, V. Genetic Variability of Grapevine Pinot gris virus and Its Association with Grapevine Leaf Mottling and Deformation. Phytopathology 2015, 105, 555–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichmeier, A.; Penazova, E.; Muljukina, N. Survey of Grapevine Pinot gris virus in certified grapevine stocks in Ukraine. Eur. J. Plant Pathol. 2018, 152, 555–560. [Google Scholar] [CrossRef]
- Poojari, S.; Lowery, T.; Rott, M.; Schmidt, A.M.; Urbez-Torres, J.R. First Report of Grapevine Pinot gris virus in British Columbia, Canada. Plant Dis. 2016, 100, 1513. [Google Scholar] [CrossRef]
- Eichmeier, A.; Penazova, E.; Nebish, A. First Report of Grapevine Pinot gris Virus on Grapevines in Armenia. Plant Dis. 2020, 104, 1000. [Google Scholar] [CrossRef]
- Wu, Q.; Habili, N. The recent importation of Grapevine Pinot gris virus into Australia. Virus Genes 2017, 53, 935–938. [Google Scholar] [CrossRef]
- Fajardo, T.V.M.; Eiras, M.; Nickel, O. First report of Grapevine Pinot gris virus infecting grapevine in Brazil. Australas Plant. Dis. 2017, 12, 45. [Google Scholar] [CrossRef]
- Zamorano, A.; Medina, G.; Fernandez, C.; Cui, W.; Quiroga, N.; Fiore, N. First Report of Grapevine Pinot gris virus in Grapevine in Chile. Plant Dis. 2019, 103, 1438–1439. [Google Scholar] [CrossRef]
- Rasool, S.; Naz, S.; Rowhani, A.; Golino, D.A.; Westrick, N.M.; Farrar, K.D.; Al Rwahnih, M. First Report of Grapevine Pinot gris virus Infecting Grapevine in Pakistan. Plant Dis. 2017, 101, 1958. [Google Scholar] [CrossRef]
- Eichmeier, A.; Penazova, E.; Cechova, J.; Berraf-Tebbal, A. Survey and Diversity of Grapevine Pinot gris virusin Algeria and Comprehensive High-Throughput Small RNA Sequencing Analysis of Two Isolates fromVitis viniferacv. Sabel Revealing High Viral Diversity. Genes 2020, 11, 1110. [Google Scholar] [CrossRef] [PubMed]
- Eichmeier, A.; Pieczonka, K.; Penazova, E.; Pecenka, J.; Gajewski, Z. Occurrence of Grapevine Pinot gris virus in Poland and description of asymptomatic exhibitions in grapevines. J. Plant Dis. Prot. 2017, 124, 407–411. [Google Scholar] [CrossRef]
- Bertazzon, N.; Filippin, L.; Forte, V.; Angelini, E. Grapevine Pinot gris virus seems to have recently been introduced to vineyards in Veneto, Italy. Arch. Virol. 2016, 161, 711–714. [Google Scholar] [CrossRef]
- Bertazzon, N.; Forte, V.; Filippin, L.; Causin, R.; Maixner, M.; Angelini, E. Association between genetic variability and titre of Grapevine Pinot gris virus with disease symptoms. Plant Pathol. 2017, 66, 949–959. [Google Scholar] [CrossRef]
- Malagnini, V.; de Lillo, E.; Saldarelli, P.; Beber, R.; Duso, C.; Raiola, A.; Zanotelli, L.; Valenzano, D.; Giampetruzzi, A.; Morelli, M.; et al. Transmission of grapevine Pinot gris virus by Colomerus vitis (Acari: Eriophyidae) to grapevine. Arch. Virol. 2016, 161, 2595–2599. [Google Scholar] [CrossRef]
- Eichmeier, A.; Penazova, E.; Pavelkova, R.; Mynarzova, Z.; Saldarelli, P. Detection of Grapevine Pinot Gris Virus in Certified Grapevine Stocks in Moravia, Czech Republic. J. Plant Pathol. 2016, 98, 155–157. [Google Scholar]
- Glasa, M.; Predajna, L.; Kominek, P.; Nagyova, A.; Candresse, T.; Olmos, A. Molecular characterization of divergent grapevine Pinot gris virus isolates and their detection in Slovak and Czech grapevines. Arch. Virol. 2014, 159, 2103–2107. [Google Scholar] [CrossRef]
- Eichmeier, A.; Kominkova, M.; Kominek, P.; Baranek, M. Comprehensive Virus Detection Using Next Generation Sequencing in Grapevine Vascular Tissues of Plants Obtained from the Wine Regions of Bohemia and Moravia (Czech Republic). PLoS ONE 2016, 11, e0167966. [Google Scholar] [CrossRef]
- Cooper, J.I.; Jones, R.A.C.; Harrison, B.D. Field and Glasshouse Experiments on Control of Potato Mop-Top Virus. Ann. Appl. Biol. 1976, 83, 215–230. [Google Scholar] [CrossRef]
- Shimomura, T. Effects of Boron on the Formation of Local Lesions and Accumulation of Callose in French Bean and Samsun Nn Tobacco-Leaves Inoculated with Tobacco Mosaic-Virus. Physiol. Plant Pathol. 1982, 20, 257–261. [Google Scholar] [CrossRef]
- Buoso, S.; Pagliari, L.; Musetti, R.; Fornasier, F.; Martini, M.; Loschi, A.; Fontanella, M.C.; Ermacora, P. With or Without You: Altered Plant Response to Boron-Deficiency in Hydroponically Grown Grapevines Infected by Grapevine Pinot Gris Virus Suggests a Relation Between Grapevine Leaf Mottling and Deformation Symptom Occurrence and Boron Plant Availability. Front. Plant Sci. 2020, 11, 226. [Google Scholar] [CrossRef] [Green Version]
- Takano, J.; Miwa, K.; Yuan, L.X.; von Wiren, N.; Fujiwara, T. Endocytosis and degradation of BOR1, a boron transporter of Arabidopsis thaliana, regulated by boron availability. Proc. Natl. Acad. Sci. USA 2005, 102, 12276–12281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.N.; Yang, C.Q.; Pan, Z.Y.; Liu, Y.Z.; Peng, S. Boron deficiency in woody plants: Various responses and tolerance mechanisms. Front. Plant Sci. 2015, 6, 916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miwa, K.; Wakuta, S.; Takada, S.; Ide, K.; Takano, J.; Naito, S.; Omori, H.; Matsunaga, T.; Fujiwara, T. Roles of BOR2, a Boron Exporter, in Cross Linking of Rhamnogalacturonan II and Root Elongation under Boron Limitation in Arabidopsis. Plant Physiol. 2013, 163, 1699–1709. [Google Scholar] [CrossRef] [Green Version]
- Takano, J.; Wada, M.; Ludewig, U.; Schaaf, G.; von Wiren, N.; Fujiwara, T. The Arabidopsis major intrinsic protein NIP5;1 is essential for efficient boron uptake and plant development under boron limitation. Plant Cell 2006, 18, 1498–1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, J.C.; Liu, Y.Z.; Yang, C.Q.; Zhou, G.F.; Wei, Q.J.; Peng, S.A. Isolation and expression analysis of CiNIP5, a citrus boron transport gene involved in tolerance to boron deficiency. Sci. Hortic. 2012, 142, 149–154. [Google Scholar] [CrossRef]
- Zhou, G.F.; Liu, Y.Z.; Sheng, O.; Wei, Q.J.; Yang, C.Q.; Peng, S.A. Transcription profiles of boron-deficiency-responsive genes in citrus rootstock root by suppression subtractive hybridization and cDNA microarray. Front. Plant Sci. 2015, 5, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, M.; Wallace, I.S.; Takano, J.; Roberts, D.M.; Fujiwara, T. NIP6;1 Is a Boric Acid Channel for Preferential Transport of Boron to Growing Shoot Tissues in Arabidopsis. Plant Cell 2008, 20, 2860–2875. [Google Scholar] [CrossRef] [Green Version]
- Eichmeier, A.; Baranek, M.; Pidra, M. Analysis of genetic diversity and phylogeny of partial coat protein domain in Czech and Italian GFLV isolates. Plant Prot. Sci. 2010, 46, 145–148. [Google Scholar] [CrossRef] [Green Version]
- Morelli, M.; Catarino, A.D.M.; Susca, L.; Saldarelli, P.; Gualandri, V.; Martelli, G.P. First Report of Grapevine Pinot Gris Virus from Table Grapes in Southern Italy. J. Plant Pathol. 2014, 96, 439. [Google Scholar]
- Pfaffl, M.W. MIQE challenges and solutions: The use of -omics technologies for transcriptional biomarker discovery. Int. J. Mol. Med. 2015, 36, S58. [Google Scholar]
- Berger, K.C. Boron in Soils and Crops. Adv. Agron. 1949, 1, 321–351. [Google Scholar]
- Krug, B.A.; Whipker, B.E.; Fonteno, W.C.; McCall, I.; Frantz, J. Incidence of Boron Deficiency in Bedding Plants Caused by Drought Stress or Abscicic Acid Application. Acta Hortic. 2011, 891, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial-DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Abreu, C.A.; van Raij, B.; de Abreu, M.F.; González, A.P. Routine soil testing to monitor heavy metals and boron. Sci. Agric. 2005, 62, 564–571. [Google Scholar] [CrossRef]
- Reuter, D.J.; Robinson, J.B. Plant Analysis: An Interpretation Manual, 2nd ed.; CSIRO Publishing: Melbourne, VIC, Australia, 1997. [Google Scholar]
- Tarquini, G.; Ermacora, P.; Bianchi, G.L.; De Amicis, F.; Pagliari, L.; Martini, M.; Loschi, A.; Saldarelli, P.; Loi, N.; Musetti, R. Localization and subcellular association of Grapevine Pinot Gris Virus in grapevine leaf tissues. Protoplasma 2018, 255, 923–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, G.L.; De Amicis, F.; De Sabbata, L.; Di Bernardo, N.; Governatori, G.; Notino, F. Occurrence of Grapevine Pinot gris virus in Friuli Venezia Giulia (Italy): Field monitoring and virus quantification by real-time RT -PCR. EPPO Bull. 2015, 45, 22–32. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | GLMD Symptom Severity | GPGV | ||
|---|---|---|---|---|
| MP/CP Acc. No. GenBank | RdRp Acc. No. GenBank | ΔΔCt (Arbitrary Value) | ||
| Sb/SO4_1 | 0 | MW147696 | MW147704 | 5.98 |
| Sb/SO4_2 | 4 | MW147697 | MW147705 | 28.37 |
| Sb/SO4_3 | 4 | MW147698 | MW147706 | 38.42 |
| Sb/SO4_4 | 2 | MW147699 | MW147707 | 2.21 |
| Sb/SO4_6 | 3 | MW147701 | MW147709 | 7.56 |
| Sb/SO4_7 | 1 | MW147702 | MW147710 | 1.26 |
| Sb/SO4_8 | 3 | MW147703 | MW147711 | 14.22 |
| Sample | B (mg.kg−1) in the Soil | B (mg.kg−1) in the Shoots and Leaves |
|---|---|---|
| Sb/SO4_1 | 0.97 | 46.1 |
| Sb/SO4_2 | 0.46 | 39.1 |
| Sb/SO4_3 | 0.40 | 33.8 |
| Sb/SO4_4 | 0.26 | 32.4 |
| Sb/SO4_6 | 0.49 | 52.4 |
| Sb/SO4_7 | 0.37 | 34.4 |
| Sb/SO4_8 | 0.19 | 46.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiss, T.; Kocanová, M.; Vavřiník, A.; Tekielska, D.; Pečenka, J.; Hakalová, E.; Špetík, M.; Štůsková, K.; Mahamedi, A.E.; Baránek, M.; et al. Incidence of GLMD-Like Symptoms on Grapevines Naturally Infected by Grapevine Pinot gris virus, Boron Content and Gene Expression Analysis of Boron Metabolism Genes. Agronomy 2021, 11, 1020. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11061020
Kiss T, Kocanová M, Vavřiník A, Tekielska D, Pečenka J, Hakalová E, Špetík M, Štůsková K, Mahamedi AE, Baránek M, et al. Incidence of GLMD-Like Symptoms on Grapevines Naturally Infected by Grapevine Pinot gris virus, Boron Content and Gene Expression Analysis of Boron Metabolism Genes. Agronomy. 2021; 11(6):1020. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11061020
Chicago/Turabian StyleKiss, Tomáš, Mária Kocanová, Aleš Vavřiník, Dorota Tekielska, Jakub Pečenka, Eliška Hakalová, Milan Špetík, Kateřina Štůsková, Alla Eddine Mahamedi, Miroslav Baránek, and et al. 2021. "Incidence of GLMD-Like Symptoms on Grapevines Naturally Infected by Grapevine Pinot gris virus, Boron Content and Gene Expression Analysis of Boron Metabolism Genes" Agronomy 11, no. 6: 1020. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11061020