
Morphological, Genetic and Biochemical Evaluation of Dasypyrum villosum (L.) P. Candargy in the Gene Bank Collection



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Cultivation
2.2. Genetic Evaluation
2.3. Disease Resistance Evaluation
2.4. Biochemical Evaluation
3. Results
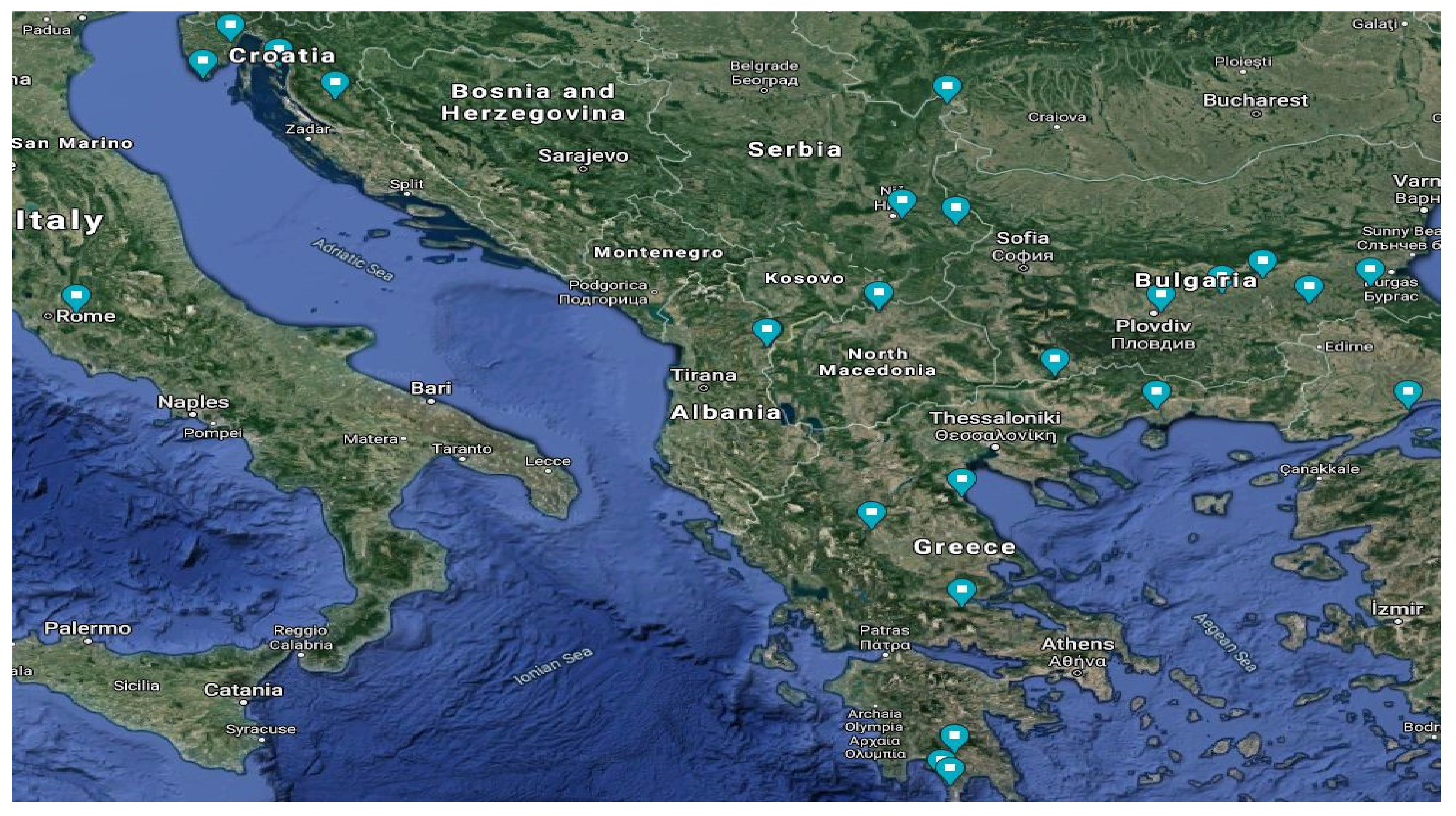
3.1. Characterization of Original Sites
3.2. Morphological Phenotyping of the Dasypyrum Collection
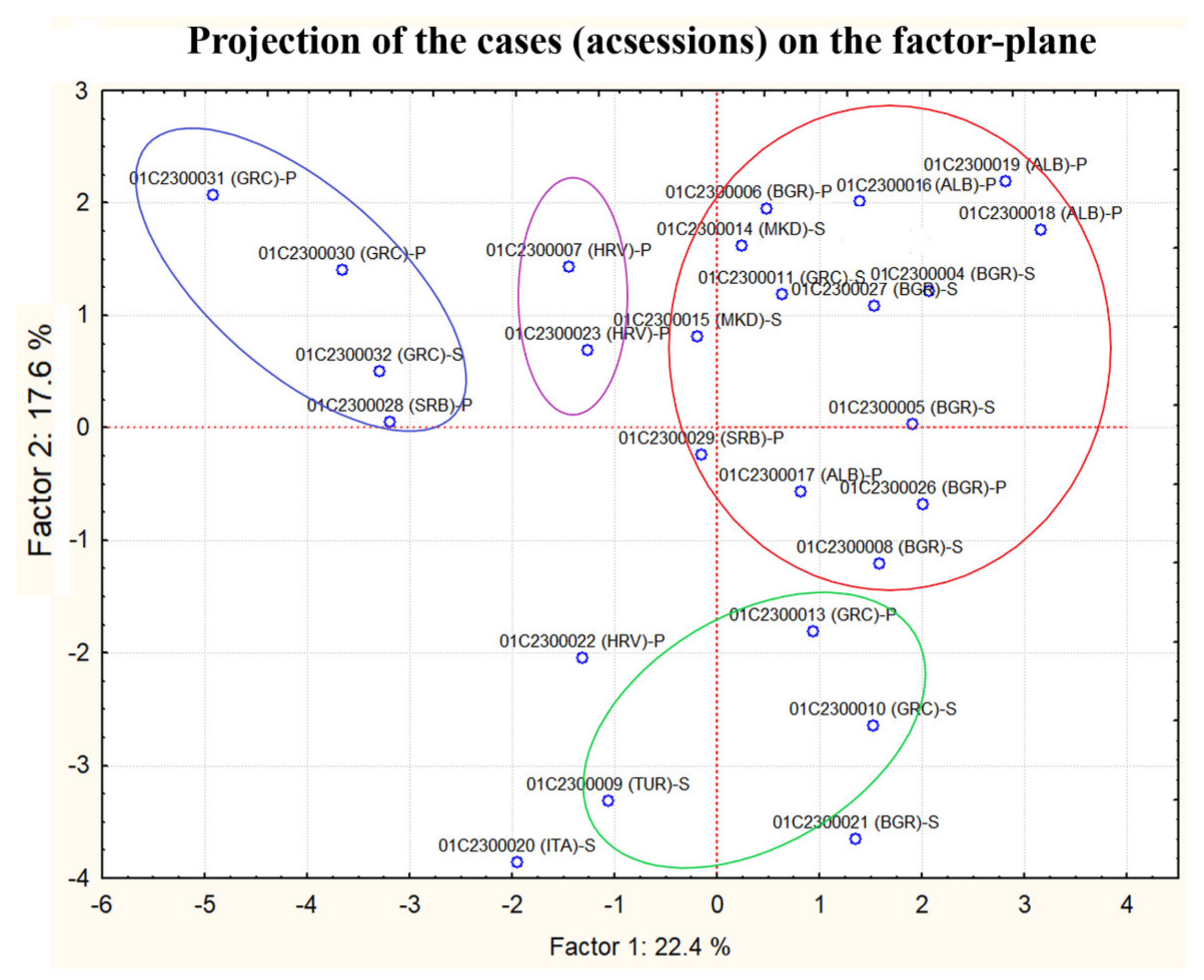
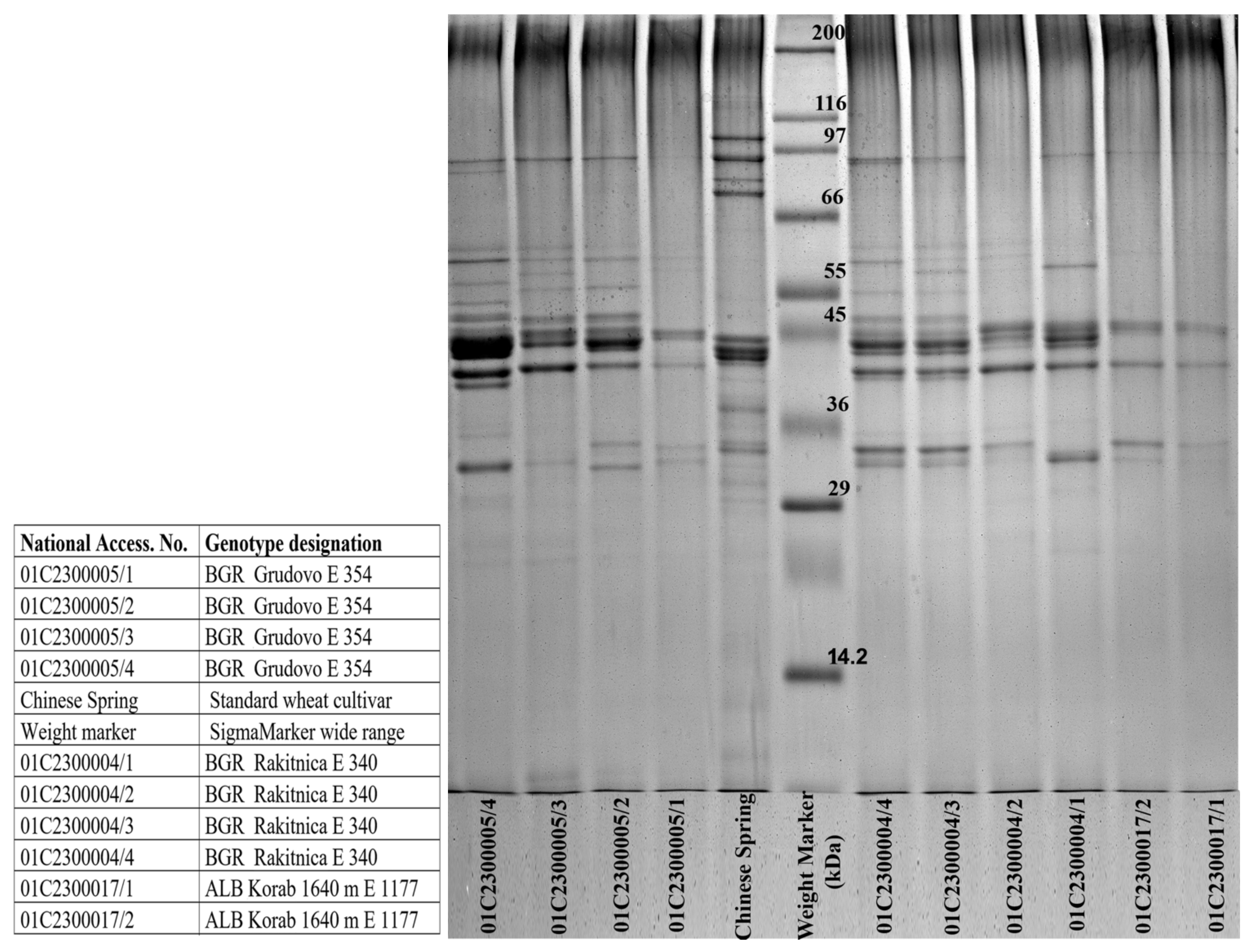
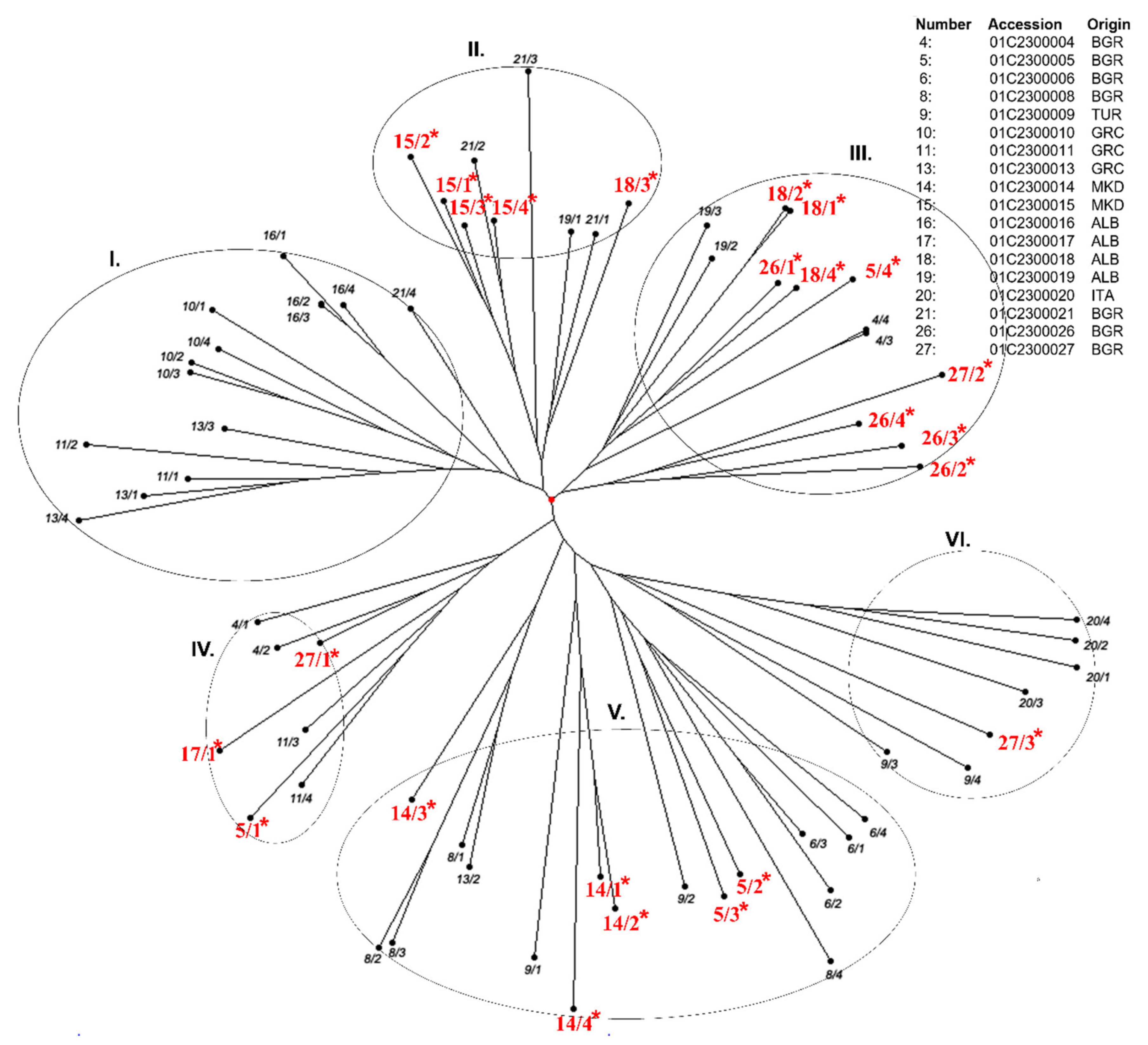
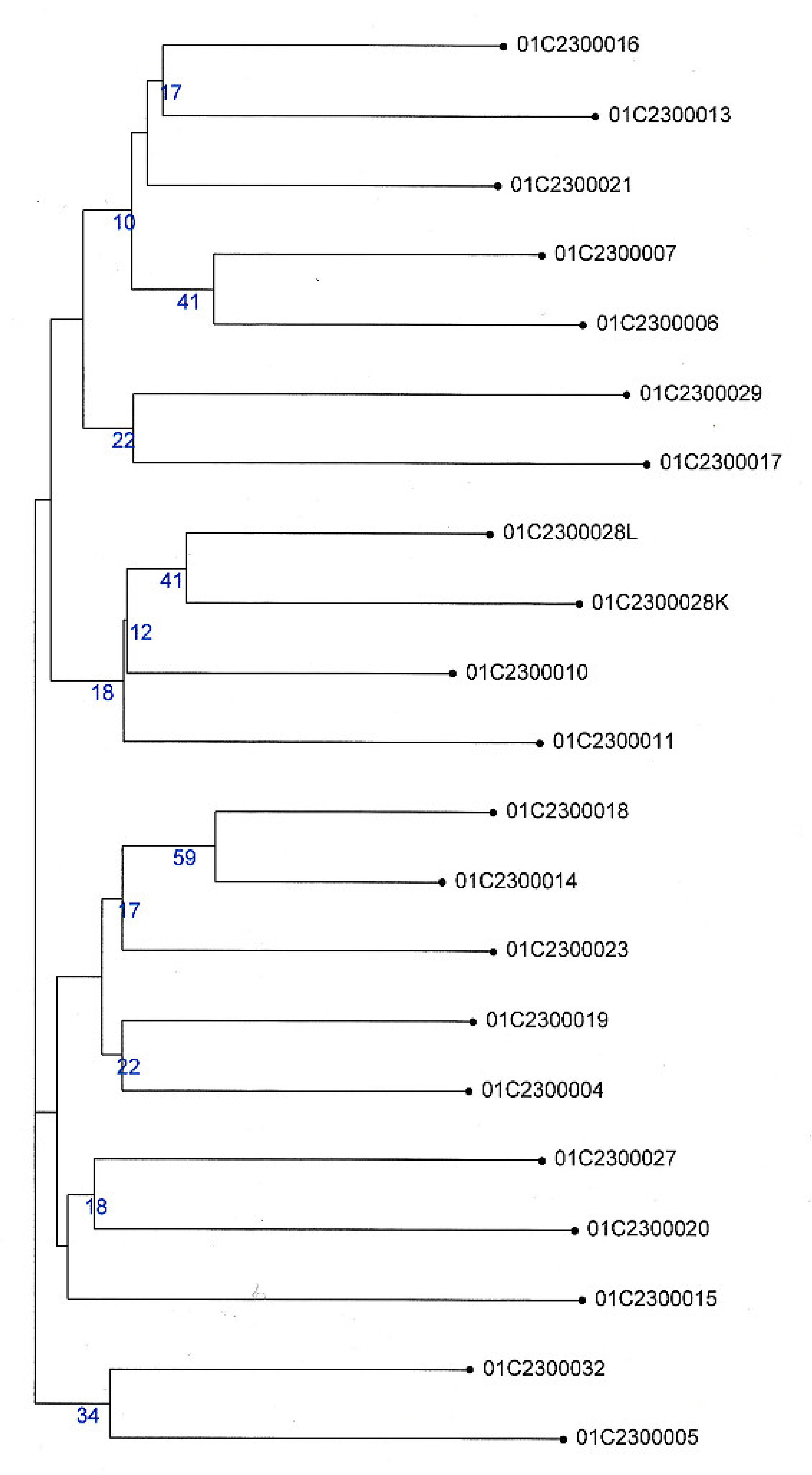
3.3. Genotyping of the Dasypyrum Collection
3.4. Resistance of Dasypyrum villosum to Leaf Diseases
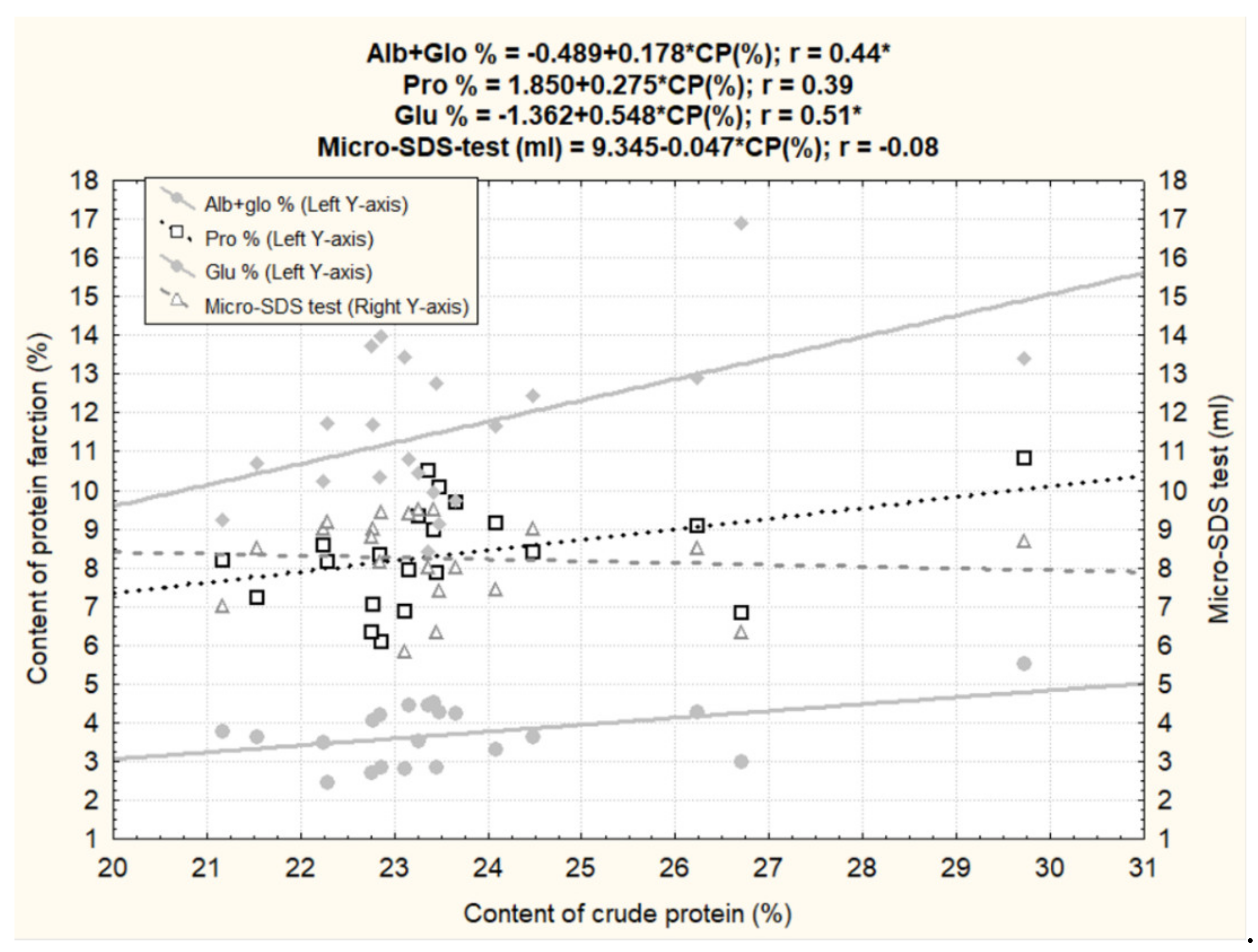
3.5. Biochemical Evaluation of the Dasypyrum Collection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hodgkin, T.; Hajjar, R. Using crop wild relatives for crop improvement: Trends and perspectives. In Crop Wild Relative Con-servation and Use; Maxted, N., Ford-Lloyd, B.V., Kell, S.P., Iriondo, J., Dulloo, E., Turok, J., Eds.; CAB International: Wallingford, UK, 2008; pp. 535–548. [Google Scholar]
- Dempewolf, H.; Baute, G.; Anderson, J.; Kilian, B. Past and future use of wild relatives in crop breeding. Crop Science 2017, 57, 1070–1082. [Google Scholar] [CrossRef]
- Bilz, M.; Kell, S.P.; Maxted, N.; Lansdown, R.V. European Red List of Vascular Plants; Publication Office of the European Union: Luxembourg, 2011. [Google Scholar]
- Kell, S.P.; Maxted, N.; Bilz, M. European crop wild relative threat assessment: Knowledge gained and lessons learnt. In Agro-biodiversity Conservation: Securing the Diversity of Crop Wild Relatives and Landraces; Maxted, Ehsan Dulloo, M., Ford-Lloyd, B., Frese, L., Iriondo, J., de Carvalho, M.P., Eds.; CAB International: Wallingford, UK, 2012; pp. 218–242. [Google Scholar]
- Maxted, N.; Scholten M, M.; Codd, R.; Ford-Lloyd, B. Creation and use of a national inventory of crop wild relatives. Biol. Conserv. 2007, 140, 142–159. [Google Scholar] [CrossRef]
- Sears E., R. Addition of the genome of H. villosa to T. aestivum. Am. J. Bot. 1953, 40, 168–174. [Google Scholar] [CrossRef]
- Gradzielewska, A. The genus Dasypyrum-part 2. Dasypyrum villosum: A wild species used in wheat improvement. Euphytica 2006, 152, 441–454. [Google Scholar] [CrossRef]
- De Bolos, O.; Vigo, J. Flora del Països Catalans; Editorial Barcino: Barcelona, Spain, 1984. [Google Scholar]
- Zhang, R.; Cao, Y.; Wang, X.; Feng, Y.; Chen, P. Development and characterization of a Triticum aestivum-H. villosa T5VS. 5DL translocation line with soft grain texture. J. Cereal Sci. 2010, 51, 220–225. [Google Scholar] [CrossRef]
- Chen, P.D.; Qi, L.L.; Zhou, B.; Zhang, S.Z.; Liu, D.J. Development and molecular cytogenetic analysis of wheat Haynaldia villosa 6VS/6AL translocation lines specifying resistance to powdery mildew. Theor. Appl. Genet. 1995, 91, 1125–1128. [Google Scholar] [CrossRef]
- Zhang, R.; Fan, Y.; Kong, L.; Wang, Z.; Wu, J.; Xing, L.; Cao, A.; Feng, Y. Pm62, an adult plant powdery mildew resistance gene introgressed from Dasypyrum villosum chromosome arm 2VL into wheat. Theor. Appl. Genet. 2018, 131, 2613–2620. [Google Scholar] [CrossRef]
- Yildirim, A.; Jones, S.S. Evaluation of Dasypyrum villosum populations for resistance to cereal eyespot and stripe rust pathogens. Plant Dis. 2000, 84, 40–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, T.D.; De La Pena, R.C.; Yildirim, A.; Jones, S.S. A new source of resistance to Pseudocercosporella herpotrichoides, cause of eyespot disease of wheat, located on chromosome 4V of Dasypyrum villosum. Plant Breed. 1994, 113, 281–286. [Google Scholar] [CrossRef]
- Liu, C.; Han, R.; Wang, X.L.; Gong, W.P.; Cheng, D.G.; Ca, X.Y.; Liu, A.F.; Li, H.S.; Liu, J.J. Research Progress of Wheat Wild Hy-bridization, Disease Resistance Genes Transfer and Utilization. Sci. Agric. Sin. 2020, 53, 1287–1308. [Google Scholar]
- McIntosh, R.A.; Dubcovsky, J.; Rogers, J.; Morris, C.; Appels, R.; Xia, X.C. Catalogue of Gene Symbols for Wheat: Komugi Integrated Wheat Science Database. 2013. Available online: http://www.shigen.nig.ac.jp/wheat/komugi/genes/download.jsp (accessed on 30 May 2021).
- Wiersma, A.T.; Pulman, J.A.; Brown, L.K.; Cowger, C.; Olson, E.L. Identification of Pm58 from Aegilops tauschii. Theor. Appl. Genet. 2017, 130, 1123–1133. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Bowden, R.L.; Krishna Jagadish, S.V.; Gill, B. Genetic variation for tolerance to terminal heat stress in Dasypyrum villosum. Crop. Sci. 2017, 57, 1–7. [Google Scholar] [CrossRef]
- De Pace, C.; Snidaro, D.; Ciaffi, M.; Vittori, D.; Ciofo, A.; Cenci, A.; Tanzarella, O.A.; Qualset, C.O.; Scarascia-Mugnozza, G.T. Introgression of Dasypyrum villosum chromatin into common wheat improves grain protein quality. Euphytica 2001, 117, 67–75. [Google Scholar] [CrossRef]
- Montebove, L.; De Pace, C.; Jan, C.C.; Scarascia-Mugnozza, G.T.; Qualset, C.O. Chromosomal location of isozyme and seed storage protein genes in Dasypyrum villosum (L.) Candargy. Theor. Appl. Genet. 1987, 73, 836–845. [Google Scholar] [CrossRef]
- Shewry, P.R.; Parmar, S.; Pappin, D.J.C. Characterization and genetic control of the prolamins of Haynaldia villosa: Relationship to cultivated species of the Triticeae (rye, wheat and barley). Biochem. Genet. 1987, 25, 309–325. [Google Scholar] [CrossRef] [PubMed]
- Kozub, N.O.; Sozinova, O.I.; Blume, Y.B. Variation of Storage Proteins in Crimean Populations of Dasypyrum villosum. Cytol. Genet. 2020, 54, 91–95. [Google Scholar] [CrossRef]
- Li, G.R.; Lang, T.; Yang, E.N.; Liu, C.; Yang, Z.J. Characterization and phylogenetic analysis of α-gliadin gene sequences reveals significant genomic divergence in Triticeae species. J. Genet. 2014, 93, 725–731. [Google Scholar] [CrossRef]
- Blanco, A.; Simeone, R.; Tanzarella, O.A.; Greco, B. Morphology and chromosome pairing of a hybrid between Triticum durum Desf. and Haynaldia villosa (L.) Schur. Theor. Appl. Genet. 1983, 64, 333–337. [Google Scholar] [CrossRef]
- Baum, B.R.; Edwards, T.; Johnson, D.A. What does the nr5S DNA multigene family tell us about the genomic relationship be-tween Dasypyrum breviaristatum and D. villosum (Triticeae: Poaceae)? Mol. Genet. Genom. 2014, 289, 553–565. [Google Scholar] [CrossRef]
- Liu, C.; Li, G.; Yan, H.; Zhou, J.; Hu, L.; Lei, M.; Ran, L.; Yang, Z. Molecular and cytogenetic identification of new wheat—D. breviaristatum additions conferring resistance to stem rust and powdery mildew. Breed. Sci. 2011, 61, 366–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Gao, D.; Zhang, H.; Li, J.; Wang, H.; La, S.; Ma, J.; Yang, Z. Molecular cytogenetic characterization of Dasypyrum breviaristatum chromosomes in wheat background revealing the genomic divergence between Dasypyrum species. Mol. Cytogenet. 2016, 9, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holubec, V.; Dumalasová, V.; Hanzalová, A. Triticeae genebank collection as a source of resistance to fungal diseases. Czech J. Genet. Plant Breed. 2005, 41, 189–192. [Google Scholar] [CrossRef] [Green Version]
- Holubec, V.; Matějovič, M. Descriptor list Aegilops. 2020. Available online: https:/www.gzr.cz/klasifikarory (accessed on 30 May 2021).
- Bareš, I.; Sehnalová, J.; Vlasák, M.; Vlach, M.; Kryštof, Z.; Amler, P.; Malý, J.; Beránek, V. Descriptor list genus Triticum. In Genové Zdroje 21; VURV: Praha, Czechia, 1985; p. 78. [Google Scholar]
- Saghai-Maroof, M.A.; Soliman, K.M.; Jorgensen, R.A.; Allard, R.W. Ribosomal DNA spacer-length polymorphisms in barley: Mendelian inheritance, chromosomal location, and population dynamics. Proc. Natl. Acad. Sci. USA 1984, 81, 8014–8018. [Google Scholar]
- Rőder, M.S.; Korzun, V.; Wendehake, K.; Plaschke, J.; Tixier, M.H.; Leroy, P.; Ganal, M.W. A microsatellite map of wheat. Genetics 1998, 149, 2007–2023. [Google Scholar]
- Roussel, V.; Leisova, L.; Exbrayat, F.; Stehno, Z.; Balfourier, F. SSR allelic diversity changes in 480 European bread wheat varieties released from 1840 to 2000. Theor. Appl. Genet. 2005, 111, 162–170. [Google Scholar]
- Perrier, X.; Jacquemoud-Collet, J.P. DARwin software. 2006. Available online: https://darwin.cirad.fr/darwin (accessed on 30 May 2020).
- Yeh, F.C.; Boyle, T.; Rongcai, Y.; Ye, Z.; Xiyan, J.M. Popgene Version 1.31. Microsoft Window-Based Freeware for Population Genetic Analysis; University of Alberta: Edmonton, AB, Canada, 1999; Available online: http://www.ualberta.ca/~fyeh/ (accessed on 30 May 2021).
- Stakman, E.C.; Stewart, P.M.; Loegering, W.O. Identification of Physiologic Races of Puccinia Graminis var. Tritici; Agricultural Research Service E617; United States Department of Agriculture: Washington, DC, USA, 1962; pp. 1–53.
- Singh, N.K.; Shepherd, K.W.; Cornish, G.B. Simplified SDS-PAGE Procedure for Separating LMW Subunits of Glutenin. J. Cereal Sci. 1991, 14, 203–208. [Google Scholar] [CrossRef]
- Zhong, G.Y.; Qualset, C.O. Allelic diversity of highmolecular- weight glutenin protein subunits in natural populations of Da-sypyrum villosum (L.) Candargy. Theor. Appl. Genet. 1993, 86, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Axford, D.W.E.; McDermott, E.E.; Redman, D.G. Small-scale tests of bread-making quality. Milling Feed Fertil. 1978, 66, 18–20. [Google Scholar]
- Dvořáček, V.; Curn, V. Evaluation of protein fractions as biochemical markers for identification of spelt wheat cultivars (Triticum spelta L.). Plant Soil Environ. 2011, 49, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Amylose/Amylopectin, Assay Procedure, K-Amyl 09/14, for the Measurement of the Amylose and Amylopectin Contents of Starch; Megazyme International Ireland Ltd.: Bray, Ireland, 2014; p. 11.
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europaea; Cambridge University Press: Cambridge, UK, 1980; Volume 5, p. 203. [Google Scholar]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Fanelli, G. The ecology of Dasypyrum villosum in Italy: Highlights from phytosociology. In Transferring Genes from the Wild Species D. Villosum to Wheat for Increasing Adaptation to Sustainable Agricultural Systems; Conference paper; Acad. Nazionale delle Scienze, Scritti e Documenti XLIV; University of Rome Tor Vergata: Rome, Italy, 2011; pp. 105–119. [Google Scholar]
- Ljubičić, I.; Paulik, H.; Bogdanović, S. Habitat mapping of Protected Landscape of Donji Kamenjak, Istria (Croatia). J. Cent. Eur. Agric. 2020, 21, 676–685. [Google Scholar] [CrossRef]
- Qualset, C.O.; De Pace, C.; Jan, C.C.; Scarascia Mugnozza, G.T.; Tanzarella, O.A.; Greco, B. Haynaldia villosa (L.) Schur: A species with potential use in wheat breeding. Am. Soc. Agron. Abstr. 1981, 70. [Google Scholar]
- Zhong, G.Y.; Dvorak, J. Evidence for common genetic mechanisms controlling the tolerance of sudden salt stress in the tribe Triticeae. Plant Breed. 1995, 114, 297–302. [Google Scholar] [CrossRef]
- Cabi, E.; Pehlivan, S. Morphological and palynological features of the genus Dasypyrum (Poaceae) in Turkey. Phytol. Balc. 2009, 15, 393–400. [Google Scholar]
- He, H.; Ji, Y.; Zhu, S.; Li, B.; Zhao, R.; Jiang, Z.; Bie, T. Genetic, Physical and Comparative Mapping of the Powdery Mildew Resistance Gene Pm21 Originating from Dasypyrum villosum. Front. Plant Sci. 2017, 8, 1914. [Google Scholar] [CrossRef] [Green Version]
- Li, G.R.; Zhao, J.M.; Li, D.H.; Yang, E.N.; Huang, Y.F.; Liu, C. A novel wheat—Dasypyrum breviaristatum substitution line with stripe rust resistance. Cytogenet. Genome Res. 2014, 143, 280–287. [Google Scholar] [CrossRef]
- Payne, P.I.; Lawrence, G.J. Catalogue of alleles for the complex gene loci, Glu-A1, Glu-B1, and Glu-D1 which code for high-molecular-weight subunits of glutenin in hexaploid wheat. Cereal Res. Commun. 1983, 11, 29–35. [Google Scholar]
- Vaccino, P.; Banfi, R.; Corbellini, M.; De Pace, C. Improving the Wheat Genetic Diversity for End-Use Grain Quality by Chromatin Introgression from the Wheat Wild Relative Dasypyrum villosum. Crop. Sci. 2010, 50, 528–540. [Google Scholar] [CrossRef] [Green Version]
- Mefleh, M.; Conte, P.; Fadda, C.; Giunta, F.; Motzo, R. From seed to bread: Variation in quality in a set of old durum wheat cultivars. J. Sci. Food Agric. 2020, 100, 4066–4074. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Number | Pl height (cm) | Spike Length (cm) | Awn Length, Apical Spikelets (cm) | Spike Color in Flower | Awn Color in Flower | Spike Color maturity | Awn Color Maturity | Spikelet Length (mm) | Glume1_Length (mm) | Awn glume2 Length (mm) | Glume2 Length (mm) | Awn Glume2 Length (mm) | Lemma Length (mm) | Awn Lemma Length (mm) | Caryopsis Length (mm) | Caryopsis Width (mm) | Caryopsis Color | Anther Length (mm) | Anthocynin in Anthers | Glume Indumentun |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 01C2300004 | 107 | 8.0 | 2.0 | 9 | 9 | 8 | 8 | 16 | 6 | 18 | 6 | 16 | 12 | 12 | 5.5 | 1.4 | 8 | 6 | 0 | 1 |
| 01C2300005 | 88 | 6.4 | 2.4 | 8 | 8 | 3 | 3 | 14 | 5 | 17 | 6 | 26 | 11 | 18 | 4.9 | 1.3 | 8 | 7 | 1 | 2 |
| 01C2300006 | 88 | 6.4 | 2.3 | 9 | 9 | 8 | 8 | 17 | 6 | 18 | 7 | 30 | 11 | 25 | 5.8 | 1.6 | 4 | 7 | 0 | 1 |
| 01C2300007 | 90 | 8.0 | 3.0 | 9 | 9 | 8 | 8 | 18 | 7 | 30 | 7 | 35 | 13 | 30 | 5.8 | 1.6 | 8 | 6 | 0 | 2 |
| 01C2300008 | 100 | 7.0 | 2.0 | 3 | 9 | 9 | 9 | 14 | 6 | 22 | 6 | 17 | 11 | 19 | 6.0 | 1.4 | 4 | 6 | 0 | 1 |
| 01C2300009 | 130 | 7.0 | 3.0 | 3 | 9 | 9 | 9 | 18 | 7 | 18 | 7 | 18 | 14 | 32 | 5.5 | 2.0 | 9 | 6 | 0 | 1 |
| 01C2300010 | 100 | 8.0 | 2.5 | 3 | 3 | 1 | 1 | 13 | 5 | 18 | 6 | 25 | 12 | 24 | 5.3 | 1.7 | 9 | 6 | 0 | 1 |
| 01C2300011 | 83 | 8.0 | 2.0 | 9 | 9 | 8 | 8 | 16 | 7 | 18 | 7 | 30 | 15 | 35 | 5.1 | 1.2 | 9 | 7 | 0 | 2 |
| 01C2300013 | 85 | 8.0 | 3.0 | 3 | 8 | 1 | 3 | 15 | 5 | 23 | 5 | 24 | 12 | 9 | 6.2 | 1.5 | 9 | 6 | 1 | 2 |
| 01C2300014 | 93 | 7.4 | 1.9 | 8 | 8 | 3 | 3 | 19 | 7 | 18 | 8 | 30 | 12 | 13 | 4.4 | 1.3 | 4 | 5 | 0 | 2 |
| 01C2300015 | 99 | 6.2 | 2.8 | 9 | 9 | 8 | 8 | 17 | 6 | 28 | 7 | 33 | 11 | 30 | 5.5 | 1.6 | 8 | 6 | 1 | 1 |
| 01C2300016 | 73 | 6.0 | 2.5 | 9 | 9 | 8 | 8 | 15 | 5 | 22 | 6 | 22 | 10 | 13 | 6.2 | 1.6 | 4 | 7 | 0 | 1 |
| 01C2300017 | 77 | 4.7 | 1.2 | 3 | 9 | 9 | 9 | 18 | 6 | 15 | 7 | 17 | 11 | 10 | 5.5 | 1.8 | 4 | 7 | 0 | 1 |
| 01C2300018 | 86 | 6.4 | 2.2 | 9 | 9 | 8 | 8 | 15 | 4 | 12 | 5 | 20 | 10 | 11 | 5.1 | 1.4 | 8 | 6 | 0 | 1 |
| 01C2300019 | 85 | 7.0 | 2.1 | 9 | 9 | 8 | 8 | 16 | 5 | 15 | 5 | 16 | 10 | 22 | 5.0 | 1.4 | 4 | 6 | 0 | 1 |
| 01C2300020 | 90 | 4.5 | 3.6 | 3 | 3 | 1 | 1 | 17 | 6 | 17 | 7 | 32 | 13 | 45 | 6.8 | 1.9 | 9 | 6 | 1 | 2 |
| 01C2300021 | 90 | 3.4 | 3.2 | 3 | 3 | 1 | 1 | 13 | 6 | 10 | 6 | 12 | 13 | 16 | 5.5 | 1.6 | 9 | 6 | 1 | 1 |
| 01C2300022 | 105 | 4.6 | 2.0 | 3 | 3 | 1 | 1 | 14 | 8 | 31 | 8 | 29 | 11 | 8 | 5.5 | 1.8 | 9 | 6 | 0 | 2 |
| 01C2300023 | 105 | 4.1 | 3.1 | 8 | 8 | 3 | 3 | 14 | 8 | 31 | 8 | 29 | 11 | 8 | 5.2 | 1.5 | 4 | 6 | 0 | 2 |
| 01C2300026 | 80 | 5.8 | 1.6 | 3 | 3 | 1 | 1 | 16 | 6 | 13 | 7 | 15 | 12 | 8 | 4.3 | 1.5 | 4 | 6 | 0 | 1 |
| 01C2300027 | 80 | 7.3 | 2.2 | 8 | 8 | 3 | 3 | 15 | 7 | 12 | 7 | 15 | 11 | 8 | 5.1 | 1.4 | 4 | 5 | 0 | 2 |
| 01C2300028 | 104 | 8.3 | 3.5 | 9 | 9 | 4 | 9 | 20 | 9 | 31 | 8 | 41 | 12 | 10 | 5.5 | 1.9 | 4 | 7 | 0 | 1 |
| 01C2300029 | 80 | 6.8 | 2.3 | 9 | 3 | 4 | 1 | 16 | 7 | 26 | 9 | 22 | 11 | 11 | 6.0 | 1.5 | 4 | 7 | 0 | 1 |
| 01C2300030 | 70 | 3.9 | 3.1 | 3 | 9 | 1 | 8 | 19 | 8 | 26 | 11 | 43 | 11 | 19 | 5.3 | 1.8 | 4 | 6 | 0 | 2 |
| 01C2300031 | 84 | 3.6 | 4.2 | 9 | 9 | 8 | 8 | 23 | 9 | 18 | 13 | 34 | 12 | 16 | 5.6 | 1.9 | 8 | 6 | 0 | 2 |
| 01C2300032 | 84 | 5.0 | 3.1 | 3 | 8 | 1 | 3 | 18 | 7 | 36 | 9 | 49 | 11 | 12 | 5.5 | 1.8 | 4 | 6 | 0 | 2 |
| Legend to Table 1. Description of characters (Descriptor list genus Triticum and Aegilops) | ||||||||||||||||||||
| 18. spike color after heading | ||||||||||||||||||||
| 3 | green (spike after heading) | |||||||||||||||||||
| 8 | violet-brown (spike after heading) | |||||||||||||||||||
| 9 | mixed | |||||||||||||||||||
| 35. glume and awn color in ripeness | ||||||||||||||||||||
| 1 | white (glumes and awns in ripeness) | |||||||||||||||||||
| 3 | black (glumes and awns in ripeness) | |||||||||||||||||||
| 4 | brown | |||||||||||||||||||
| 8 | mixed white-black | |||||||||||||||||||
| 9 | mixed white-brown | |||||||||||||||||||
| 40. caryopsis color in ripeness | ||||||||||||||||||||
| 4 | light brown (caryopsis in ripeness) | |||||||||||||||||||
| 5 | dark brown (caryopsis in ripeness) | |||||||||||||||||||
| 9 | mixed light brown-dark brown | |||||||||||||||||||
| glume indumentum: | ||||||||||||||||||||
| 1 | coarse, bristle-like hairs, rather remote | |||||||||||||||||||
| 2 | soft, softer hairs, usually connected | |||||||||||||||||||
| anthers: anthocyanin presence | ||||||||||||||||||||
| 1 | anthocyanin present | |||||||||||||||||||
| 0 | anthocyanin absent | |||||||||||||||||||
| National Acc. No. | No of Eval. YY | Powdery Mildew * | Yellow Rust * | Leaf Rust * | Stem Rust * |
|---|---|---|---|---|---|
| 01C2300004 | 7 | 9 | 9 | 9 | 9 |
| 01C2300005 | 9 | 9 | 9 | 9 | 9 |
| 01C2300006 | 9 | 9 | 3 (2/50) ** | 9 | 9 |
| 01C2300007 | 2 | 9 | 9 | 9 | 9 |
| 01C2300008 | 1 | - | - | 9 | 9 |
| 01C2300009 | 1 | - | - | 9 | 9 |
| 01C2300010 | 5 | 9 | 2 (2/60) ** | 9 | 9 |
| 01C2300011 | 6 | 9 | 9 | 9 | 9 |
| 01C2300013 | 1 | 9 | 9 | 9 | 9 |
| 01C2300014 | 7 | 9 | 9 | 9 | 9 |
| 01C2300015 | 7 | 9 | 9 | 9 | 9 |
| 01C2300016 | 3 | 9 | 9 | 9 | 9 |
| 01C2300017 | 2 | 9 | 9 | 9 | 9 |
| 01C2300018 | 7 | 9 | 9 | 9 | 9 |
| 01C2300019 | 3 | 9 | 9 | 9 | 9 |
| 01C2300020 | 7 | 9 | 6 (2/10) ** | 9 | 9 |
| 01C2300021 | 5 | 9 | 9 | 9 | 9 |
| 01C2300026 | 4 | 9 | 9 | 9 | 9 |
| 01C2300027 | 4 | 9 | 9 | 9 | 8 (2/1) ** |
| 01C2300028 | 4 | 9 | 9 | 9 | 9 |
| 01C2300029 | 5 | 9 | 9 | 9 | 7 (2/1) ** |
| 01C2300030 | 1 | 9 | 9 | 9 | 9 |
| 01C2300031 | 1 | 9 | 9 | 9 | 9 |
| Parameter | Mean | Min. | Max. | St. Dev. | RSD (%) | Granny (T. aestivum) Mean ± St. Dev. |
|---|---|---|---|---|---|---|
| Crude Protein (%) | 23.7 | 21.2 | 29.7 | 1.9 | 8.0 | 13.6 ± 0.4 |
| Starch (%) | 51.3 | 41.2 | 55.9 | 3.6 | 7.0 | 63.8 ± 1.4 |
| Alb + Glo (%) | 3.7 | 2.4 | 5.5 | 0.8 | 20.8 | 3.6 ± 0.1 |
| Pro (%) | 8.4 | 6.1 | 10.8 | 1.3 | 15.9 | 4.6 ± 0.2 |
| ΣGlu (%) | 11.6 | 8.4 | 16.9 | 2.0 | 17.6 | 5.4 ± 0.3 |
| Amylosa (%) | 26.3 | 20.1 | 34.9 | 3.4 | 12.9 | 27.7 ± 2.3 |
| Micro-SDS test(ml) | 8.2 | 5.8 | 9.5 | 1.1 | 13.7 | 6.7 ± 0.3 |
| Genotype | Origin | CP (%) | Starch (%) | Alb + Glo (%) | Pro (%) | ΣGlu (%) | Amylose (%) | M-SDS Test (ml) |
|---|---|---|---|---|---|---|---|---|
| 01C2300005 | BGR | 22.2 a | 53.7 a | 3.2 a | 8.5 ab | 10.5 a | 29.5 a | 8.6 a |
| 01C2300014 | MAK | 23.7 a | 50.1 ab | 4.0 b | 9.8 b | 9.9 a | 25.3 a | 7.8 a |
| 01C2300015 | MAK | 23.1 a | 53. 7 a | 3.5 ab | 8.8 ab | 10.7 ab | 24.9 a | 7.6 a |
| 01C2300017 | ALB | 23.5 a | 53.3 a | 3.6 ab | 7.5 a | 12.4 c | 26.0 a | 9.3 b |
| 01C2300018 | ALB | 22.8 a | 50.9 ab | 3.6 ab | 7.3 a | 11.9 bc | 25.7 a | 8.6 a |
| 01C2300026 | BGR | 22.7 a | 52.3 a | 3.7 ab | 7.7 a | 11.4 abc | 27.2 a | 7.9 a |
| 01C2300027 | BGR | 26.5 b | 47.0 b | 3.6 ab | 8.0 ab | 14.9 d | 25.3 a | 7.8 a |
| Year—2008 | 23.5 a | 51.7 a | 3.8 b | 8.5 b | 11.2 b | 23.9 b | 8.8 a | |
| Year—2009 | 22.9 a | 51.1 a | 4.3 c | 9.0 b | 9.6 a | 26.6 a | 8.3 a | |
| Year—2011 | 23.6 a | 52.5 a | 2.9 a | 7.3 a | 13.4 c | 28.5 a | 7.6 b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holubec, V.; Dvořáček, V.; Svobodová Leišová, L.; Ercisli, S. Morphological, Genetic and Biochemical Evaluation of Dasypyrum villosum (L.) P. Candargy in the Gene Bank Collection. Agronomy 2021, 11, 1316. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11071316
Holubec V, Dvořáček V, Svobodová Leišová L, Ercisli S. Morphological, Genetic and Biochemical Evaluation of Dasypyrum villosum (L.) P. Candargy in the Gene Bank Collection. Agronomy. 2021; 11(7):1316. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11071316
Chicago/Turabian StyleHolubec, Vojtěch, Václav Dvořáček, Leona Svobodová Leišová, and Sezai Ercisli. 2021. "Morphological, Genetic and Biochemical Evaluation of Dasypyrum villosum (L.) P. Candargy in the Gene Bank Collection" Agronomy 11, no. 7: 1316. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11071316