1. Introduction
Although lettuce (
Lactuca sativa L.) was historically grown and used in the “Old Continent” (Mediterranean areas), nowadays, this tender and fresh leafy vegetable belonging to the Asteraceae has also widely spread in more industrialized countries such as China and the USA [
1,
2]. Due to its high water content, the nutritional quality of lettuce is often undervalued [
3]. However, its low caloric, sodium, and fat contents have made it a well-liked and popular vegetable, electing it as a staple ingredient for fresh meals like salads [
4,
5]. The genetic variability of the genus
Lactuca, which is the fruit of intense breeder selection over the years, provides consumers with a wide assortment of leaf typology that differs in color (red to green), morphology (smooth or curly), size, and nutritional characteristics [
3,
6,
7]. Lettuce is usually consumed raw, exalting its crispness and freshness and preserving its nutritional value. The health attributes of this leafy vegetable are ascribed to its dietary fibers and bioactive compounds such as vitamins (A, B9, and C), carotenoids, and phenolic compounds [
8,
9]. Nowadays, the demand for lettuce is steadily growing due to its appreciated beneficial effects on human well-being [
5].
The industrialization process has led to a depopulation of the rural areas by accelerating the urban development process and unavoidably causing the loss of agricultural land [
10]. In this scenario, hydroponic cultivation is a practical approach to address this challenge as it fits well with the urban landscape, supplying fresh food and solving the agricultural soil loss problem [
11]. As if that were not enough, traditional agricultural systems consume large volumes of water, endangering the water supply of future generations [
2]. These systems’ environmental and economic unsustainability highlights the urgent need to move towards innovative agronomic methods, such as closed-loop hydroponics [
12]. Among the different hydroponic techniques, the floating raft system (FRS) has become the most successful for producing leafy vegetables such as lettuce, basil, rocket, and radish [
13,
14]. Compared to conventional soil-based cultivation, FRS guarantees the (i) de-seasonalization of production, (ii) speeding up the growth cycle, (iii) increment of water use efficiency, (iv) reduction of labor costs, (v) increment of unit production through higher planting density, (vi) achievement of more stable and repeatable growth conditions, (vii) reduction of the incidence of telluric pathogens, and (viii) overcoming of the limiting conditions such as high salinity, heavy metal contamination, and suboptimal soil reactions (alkalinity and acidity) [
5,
8,
13,
14,
15]. FRS bestows a benefit for both consumers and fresh-cut vegetable growers who can produce residue-free soil products, which results in fewer washing procedures with less water waste [
12,
13].
However, it is noteworthy that using FRS to grow leafy vegetables does not inevitably result in quality improvement [
16]. The imperative to achieve higher yield with lower costs has prompted the use of nutrient solutions often exceeding in nitrogen, causing a hyper-accumulation of nitrate, especially in green leafy vegetables, thus posing a potential hazard for the environment and food safety [
17,
18]. Several studies have proven that nitrate is safe, but salivary bacteria can reduce it into more harmful metabolites such as nitrites, nitric oxide, and nitrosamines, which are recognized to play a critical role in the processes of carcinogenesis and blood diseases like methemoglobinemia [
19,
20]. Since nitrate exposure depends on leafy vegetable consumption, the European Community has set restrictive thresholds for their commercialization [
21]. At present, hydroponic systems, through the possibility of promptly managing the composition of the nutrient solution, could be used to reduce nitrate levels in leafy tissues. For this purpose, several authors proposed different agronomic strategies such as nutrient solution replacement with water, the use of low-nitrate nutrient solution, or nitrate replacement with chloride a few days before harvest [
15,
20,
22,
23,
24,
25].
Moreover, the potential to apply eustress (positive stress) through the management of the nutrient solution could not only reduce the amount of antinutritional compounds (e.g., nitrate) but also trigger the biosynthesis of specialized metabolites such as carotenoids, flavonoids, phenols, and vitamin C which are increasingly sought after by consumers due to their antioxidant properties [
7,
26]. To date, few works have investigated the effects induced by nutrient solution deprivation close to harvest, on the primary and secondary metabolism of leafy vegetables. For this purpose, our research was aimed to evaluate the effects of nutrient solution replacement with water, three and six days before harvest, on yield and nitrate accumulation as well as on the biosynthesis of antioxidant metabolites of ready-to-eat lettuce (
Lactuca sativa L.) in a closed-loop hydroponic system.
2. Materials and Methods
2.1. Plant Material and Experimental Design
The experiment was conducted in a passively ventilated greenhouse (15 m × 5 m), located at the Department of Agriculture (DIA) of the University of Naples Federico II (Portici, Italy; 40°48′ N, 14°20′ E, 29 m.s.l.). During the crop cycle, air temperature, air relative humidity, and photosynthetic photon flux density (PPFD) measurements were recorded every 30 min using WatchDog A150 dataloggers (Spectrum Technologies Inc., Aurora, IL, USA) (
Supplementary Figure S1). On 2 October 2020 (18 days after sowing), seedlings of lettuce (
Lactuca sativa L.) ‘Maravilla De Verano Canasta’ hereafter ‘Canasta’ (Butterhead lettuce; Pagano Domenico & Figli, Scafati, Salerno, Italy) were transplanted at the two true-leaf stage in polystyrene trays with a planting density of 70 plants m
−2. Experimental treatments consisted of a control (0 DD) managed with nutrient solution (NS) throughout the growing cycle and two deprivation treatments applied 3 (3 DD) and 6 (6 DD) days prior to harvest by completely replacing nutrient solution with osmotic water only (0.156 mM potassium, 0.217 mM magnesium, 0.205 mM calcium, 0.065 mM nitrate, 0.010 sulfate, 0.003 phosphate; pH 6, electrical conductivity (EC) 150 µS cm
−1) arranged in a randomized complete block design (RCBD) with three replicates. Each experimental unit consisted of a 35 L plastic tank (0.60 × 0.40 × 0.24 m; L × W × D) on which floated a polystyrene tray (52 × 33 m) anchoring 12 plants (total of 36 plants per treatment). Oxygenation of the aqueous medium (NS or water) was provided by immersion pumps equipped with a Venturi system (Aquaball 60, Eheim, Stuttgart, Germany).
The macronutrient concentration of the NS was: 8.2 mM nitrate, 0.7 mM phosphate, 1.0 mM sulfate, 2.7 mM potassium, 2.9 mM calcium, 0.7 mM magnesium, and 1.0 mM ammonium, while the micronutrient concentration was: 20 µM iron, 20 µM boron, 9 µM manganese, 1.6 µM zinc, 0.3 µM copper, and 0.3 µM molybdenum. The electrical conductivity of NS was 1.2 ± 0.1 mS cm−1, whereas pH was monitored every day and maintained at 5.8 ± 0.3 using a portable pH meter (HI 991301, Hanna Instruments, Milan, Italy) by adding an acid mixture (mainly phosphoric acid) with the same anion ratio as NS.
2.2. Sampling and CIELab Colorimetric Index
The experimental trial was carried out from 2 October to 30 October 2020. At 29 days after transplanting (DAT), ten plants per replicate were harvested, weighed for fresh weight determination, and separated into leaves and stems. Leaves of each plant were counted, and the leaf area was quantified by digital image analysis with ImageJ v1.52a software (U.S. National Institutes of Health, Bethesda, MD, USA) after image acquisition with Nikon D80 camera (Nikon Corporation, Tokyo, Japan). Two plants per replicate were stored at −20 °C for further qualitative analysis. Contextually to harvest, colorimetric measurements were performed on the upper side of ten fully expanded young leaves for each experimental unit. Specifically, a Minolta CR-300 Chroma Meter (Minolta Camera Co. Ltd., Osaka, Japan) was used to obtain the color space parameters (L, a, and b) and then to calculate the chroma and HUE angle values. The leaf tissues were placed in a ventilated oven for ~72 h at 60 °C until constant weight to determine the dry biomass and leaf dry matter percentage (%). Dry matter percentage was also calculated using the following formula: DM = 100 × Dry weight/Fresh weight.
2.3. Spad Index and Chlorophyll Fluorescence
On the harvest day (October 30, 29 DAT), on the adaxial side of twenty fully expanded young leaves of twelve plants per experimental unit, SPAD index measurements were made using a portable SPAD-502 m (Minolta Camera Co. Ltd., Osaka, Japan). On the same date and the same leaves used for SPAD index determination, between 11:00 a.m. and 1:00 p.m., measurements of the maximum quantum efficiency of PSII expressed as Fv/Fm were determined using a portable fluorometer (Plant Stress Kit, Opti-Sciences, Hudson, NH, USA).
2.4. Pigments Determination
As proposed by Wellburn [
27], for pigment determination (i.e., total chlorophyll and carotenoids), five hundred mg of two fresh frozen leaves per replicate were extracted in 3 mL of 90% (
v/v) ammonia acetone (Carlo Erba Reagents Srl, Milan, Italy) and ground using a ceramic pestle and mortar. Chlorophyll a, b, and carotenoids contents were determined by UV-Vis spectrophotometer (DR 4000, Hach Co, Loveland, CO, USA) with absorbance of 647, 664, and 470 nm, respectively. Total chlorophyll, calculated as the sum of chlorophylls a and b, and carotenoids were expressed as mg g
−1 of fresh weight (fw).
2.5. Total Nitrogen and Mineral Profile Determination
The total nitrogen (N) concentration of dried and finely ground lettuce samples by means of MF10.1 cutting-grinding head mill (IKA
®, Staufen im Breisgau, Baden-Württemberg, Germany) and MF0.5 sieve (0.5 mm hole size; IKA
®, Staufen im Breisgau, Baden-Württemberg, Germany) was determined by the Kjeldahl method after mineralization with sulfuric acid (Carlo Erba Reagents Srl, Milan, Italy) and a catalyst (3.5 g K
2SO
4 + 0.1 g CuSO
4 × 5H
2O; Velp
® Scientifica, Usmate Velate, Monza Brianza, Italy) as described in detail by Formisano et al. [
28]. Minerals (P, K, S), nitrate, and organic acids (malate, tartrate, oxalate, citrate, and isocitrate) were determined by ion chromatography (ICS3000, Thermo Scientific™ Dionex™, Sunnyvale, CA, USA) coupled to an electrical conductivity detector using the Chromeleon™ 6.8 software (Thermo Scientific™ Dionex™, Sunnyvale, CA, USA) according to the method described by Pannico et al. [
29]. Minerals and organic acids concentrations were expressed as mg g
−1 of dry weight (dw), while nitrate was expressed as mg kg
−1 fw.
2.6. Total Phenol, Total Ascorbic Acid, Anthocyanins, Hydrogen Peroxide, and Antioxidant Activities Determination
For compositional and phytochemical analyses, samples of fresh matter from randomly selected lettuce leaves were instantly frozen in liquid nitrogen and stored at −80 °C before lyophilized in a Christ, Alpha 1–4 (Martin Christ Gefriertrocknungsanlagen GmbH, Osterode am Harz, Germany) freeze drier [
30].
Total phenol and total ascorbic acid analysis were performed by spectrophotometry according to the method proposed by Singleton et al. [
31] and Kampfenkel et al. [
32], respectively. Gallic acid and ascorbate standards (Sigma Aldrich, Milan, Italy) standards were used to create calibration curves to quantify total phenols and total ascorbic acid content. A UV-VIS spectrophotometry (Hach DR 4000, Hach CO., Loveland, CO, USA) with an absorbance of 765 and 525 nm, respectively, was used to determine total phenols and total ascorbic acid, and the results were expressed as mg gallic acid equivalent 100 g
−1 dw and mg 100 g
−1 fw, respectively. Total anthocyanin analysis was performed according to Rouphael et al. [
33] by suspending 20 mg lyophilized leaf samples in 180 µL of 40% (
v/v) ethyl alcohol and incubating it on ice for 20 min. After 14,000 rpm centrifugation for 10 min, the pellet was re-extracted in 180 µL of 40% (
v/v) ethyl alcohol, and the two supernatants joined. Duplicate aliquots of 150 µL of the supernatant (or 40%
w/v ethanol solution as blank) were placed in a polypropylene microplate with 75 µL of 25 mM KCl (pH 1.0) or 75 µL of 400 mM sodium acetate (pH 4.5). The absorbance was recorded at 520 and 700 nm by a Synergy HT spectrophotometer (BioTEK Instruments, Bad Friedrichshall, Germany). The anthocyanin content was expressed as mg cyanidin−3-glucoside equivalents (C3Geq) g
−1 dw. Hydrogen peroxide (H
2O
2) content was determined according to Dell’Aversana et al. [
34] with few modifications. Aliquots of 20 mg of lyophilized leaf tissues were suspended in 230 µL of 0.1% (
w/v) of trichloroacetic acid, vortexed and, after at 4 °C for 10 min, 50 µL of clear supernatants were placed in wells of a polypropylene microplate with 100 µL of 1 M KI and 50 µL of 10 mM phosphate buffer (pH 7.0). H
2O
2 concentration was assayed at 390 nm for comparison with standard curves of known concentrations of H
2O
2 and expressed as µmol·g
−1 dw. To quantify the antioxidant activity, 0.2 g of freeze-dried lettuce leaves were subjected to different extraction procedures. Hydrophilic (HAA) and 2,2’-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) antioxidant activities were measured according to the protocol proposed by Fogliano et al. [
35] and Pellegrini et al. [
36]. For ABTS antioxidant activity, a radical cation decolorization assay 3-ethylbenzothiazoline-6-sulfonic acid (2, 2′-azinobis (Sigma Aldrich, Milan, Italy) was used. The HAA and ABTS content of the obtained solutions were quantified by UV-VIS spectrophotometry (Hach DR 4000, Hach CO., Loveland, CO, USA) with absorbance at 505 and 734 nm, respectively. Each treatment was analyzed in triplicate and the results were expressed as mmol of ascorbic acid equivalent 100 g
−1 dw and mmol Trolox equivalent 100 g
−1 dw for HAA and ABTS, respectively.
2.7. Carbohydrates, Soluble Proteins, and Amino Acids Analysis
The analysis of soluble carbohydrates content (expressed as µmol g
−1 dw) was performed in ethanolic extracts of lyophilized lettuce leaf tissues, while the starch (expressed as µmol glucose equivalents g
−1 dw) was estimated in the pellet of the same ethanolic extract after hydrolysis to glucose, by supplying oxidized pyridine nucleotide and measuring its rate of reduction in the presence of sugars according to Carillo et al. [
24]. The absorbance was recorded at 340 nm by a Synergy HT spectrophotometer (BioTEK Instruments, Bad Friedrichshall, Germany). Soluble proteins were analyzed by suspending 20 mg of lyophilized leaf tissues in 1 mL of Tris-HCl 200 mM (pH 7.5) containing 500 mM MgCl
2. Samples were then centrifuged and the proteins measured in the supernatant by using the Protein Assay Dye Reagent Concentrate (Bio-Rad, Milan, Italy), according to Rouphael et al. [
32]. Proteins were expressed as mg g
−1 dw and primary amino acids were determined in ethanolic extracts of lyophilized leaf samples and assayed by FLD-HPLC, according to Carillo et al. [
24]. Amino acids were expressed as µmol g
−1 dw.
2.8. Statistics
The statistical analysis was performed using IBM SPSS Statistics (IBM Corp., Armonk, NY, USA), version 26.0 package for Microsoft Windows 10 (Microsoft Corporation, Redmond, WA, USA). The mean effect of all parameters investigated was compared according to one-way ANalysis of VAriance (ANOVA). Statistical significance was determined at the p = 0.05 level using Tukey’s HSD test. All data are presented as mean, n = 3.
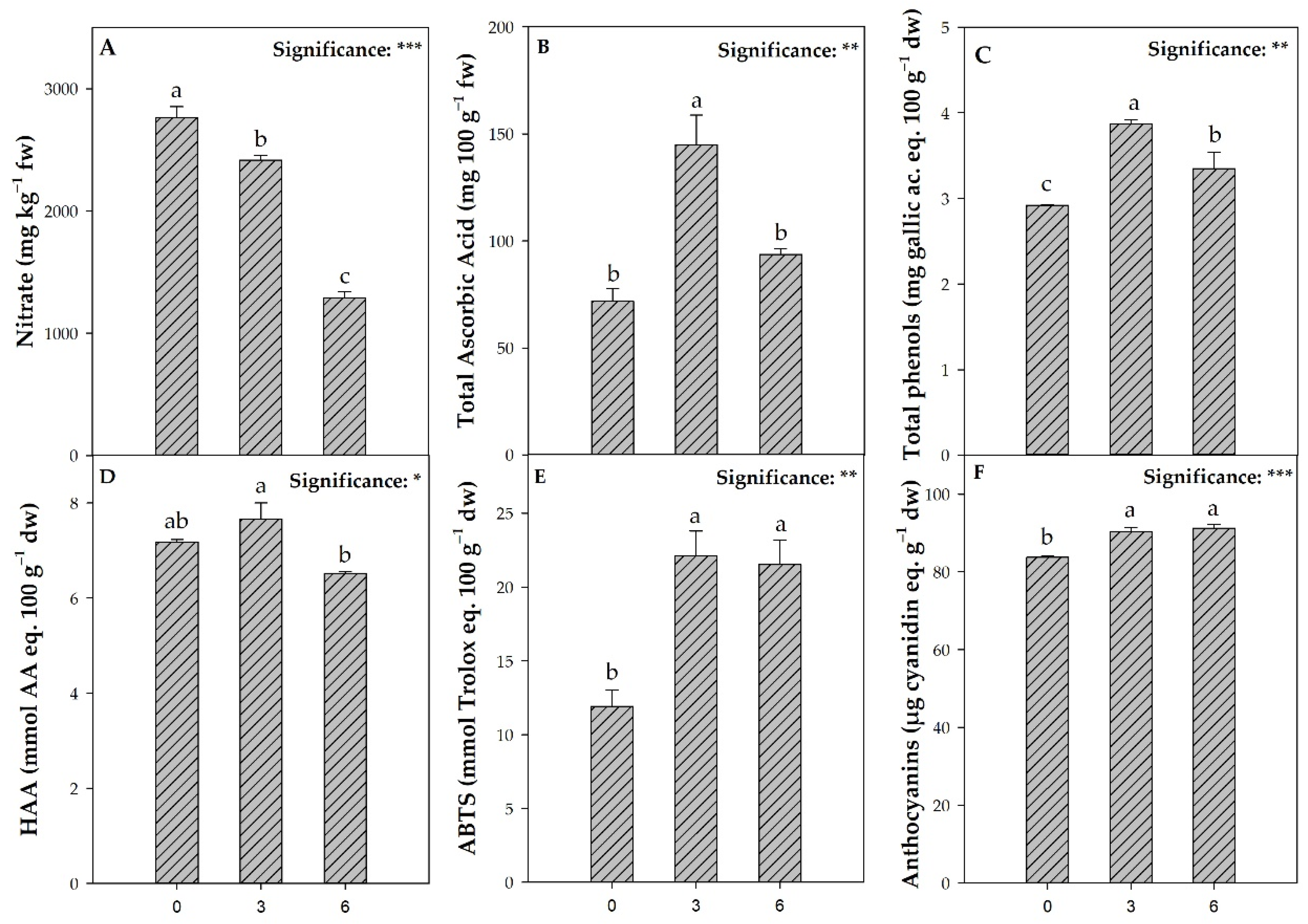
4. Discussion
Nutrient solutions (NS) used in hydroponics have high levels of nutrients, more than twice of those used in traditional soil-based cultivation [
13]. Minerals’ over-rich NS, though capable of increasing productivity within a certain threshold, can lead to quality losses in leafy vegetables (e.g., sensory flattening, delay in harvesting and nitrate accumulation in leaf tissues), and pose an environmental hazard [
15]. Nowadays, researchers are continuously searching for innovative NS management strategies to reduce vegetables’ nitrate content without affecting crop yield. Among these, partial nitrate replacement with chloride in the NS is the most investigated and effective technique [
15,
25]. However, residual chlorine-rich NS disposal can decimate soil microbiological and edaphic populations, thus jeopardizing “soil health” [
37,
38]. Based on the above, we evaluated if replacing NS with only water close to harvest could be a proper eco-friendly strategy to improve the qualitative and quantitative performance of lettuce grown in floating raft systems (FRS).
The fresh yield and dry biomass reduction resulting from NS deprivation different days before harvest highlight that leafy vegetable yield in hydroponics is affected by nutrient supply [
39,
40,
41,
42]. Probably, the yield and dry biomass reduction of lettuce (
Lactuca sativa L.) deprived of NS three and six days before harvest was caused by an osmotic effect and, in particular, by macronutrient deficiency [
8]. On the other hand, Santamaria et al. [
22] did not report a reduction of these parameters in arugula and chicory grown under nitrate deficiency. The reduction of biometric parameters observed in the present experiment would probably not be due solely to nitrate deficiency but to the deprivation of other macronutrients crucial for the maintenance of metabolic processes [
43,
44].
Compared to the 6 DD treatment, plants subjected to 3 DD were able to sustain growth due to the still-sufficient nutrients pool, thus guaranteeing higher fresh yield. This emphasizes that nutritional stresses depend on their magnitude and especially on the time of exposure to the stress [
45,
46]. Regardless of the NS deprivation treatment, the number of leaves was unaffected. However, at 6 DD, a significant reduction in leaf area was observed. Long-term nutrient deprivation (particularly nitrogen) probably resulted in the redirection of accumulated nutrient reserves to the root system and young leaves at the expense of mature ones, ensuring their growth and survival [
47].
Similarly, as observed for biometric parameters, NS deprivation reduced F
v/F
m values. However, the values recorded at 6 DD (0.829) and 3 DD (0.834) were not indicative of stress oxidative damage [
48,
49]. To support the above, it is noteworthy that total chlorophyll did not change, in contrast to the findings of several authors on nitrogen-depleted leafy vegetables [
50,
51,
52], falling within the optimal range for lettuce (0.482–0.821 mg g
−1 fw) [
53]. Moreover, chlorophyll affects the greenness of leafy vegetables, driving consumer choice [
54,
55]. The greenness (a) variation in plants deprived of NS 6 days before harvest was not attributable to reduced chlorophyll but to an increased anthocyanins content due to macronutrient deficiency, as supported by several authors [
9,
56,
57].
Similar to previous work carried out on several leafy vegetables grown under nutrient-limited conditions [
8,
41,
58,
59], in our work, the replacement of the NS with water six days before harvest reduced the nitrogen and potassium uptake compared to the control, thus justifying the lower yield. Relative to potassium, although a positive correlation with carotenoids content in leaves is well established [
39], in our experiment, potassium reduction at 6 DD was not associated with a decrease in carotenoids. Probably, potassium levels in plants at 6 DD, being above the optimum value (53.7 mg g
−1 dw) for the Butterhead lettuce cultivar ‘Maravilla De Verano Canasta’ type [
60], did not affect carotenoids biosynthesis, in comparison with control (0 DD) and 3 DD treatment. The reduction in potassium content obtained in the present experiment could be helpful to dialysis patients for whom potassium intake must be restricted [
61,
62,
63,
64,
65,
66].
As reported by several authors [
41,
67,
68,
69], nitrate accumulation in leafy vegetables, other than depending on genotype, is also affected by crop management techniques and light conditions, that differ when plants are grown in different seasons of the year [
20]. In effect, nitrate assimilation depends on irradiance and thus on photosynthesis as it provides the carbon skeletons necessary to trigger nitrogen assimilation into amino acids [
70]. As expected, in our experiment, NS deprivation treatments reduced leaf nitrate content compared to the control (0 DD). Specifically, the values obtained at 3 DD and 6 DD were about two and four-fold lower, respectively, than the threshold limit of 5000 mg kg
−1 fw set by the EU Regulation 1258/2011 [
21]. Interestingly, the nitrate reduction after NS deprivation was higher than that of total nitrogen, which agrees with Gent’s [
71] findings in a previous study. The lower nitrate concentration at 6 DD could be attributable to the reduction of NO
3− reserve (vacuolar nitrate) content that plants use to support metabolic processes [
22]. During the nitrate assimilation process, vacuolar NO
3- ions are replaced by organic acids to prevent alkalization [
72]. The role of organic acids as crucial counter-anions for nitrogen metabolism justifies the lower malate, tartrate, and isocitrate values at 6 DD because a high deficiency of nitrate could impair the enzymatic activity involved in organic acid biosynthesis [
72]. In contrast, deprivation treatments (3 DD and 6 DD) increased starch content in leaves, compared to the control (0 DD), as nitrate depletion may have prompted plants to accumulate starch as an osmotically inactive form of carbon [
72]. Moreover, since nitrate is sensed by plants that adjust their metabolism according to its concentration, the decrease of this important nutrient results in coordinate alterations in the activities of enzymes in several metabolic pathways that are directly or indirectly involved in its reduction and assimilation, including ammonium assimilation, organic acid synthesis, and starch metabolism [
72]. Thus, at 3 DD and 6 DD, starch, sucrose, and glucose increased in lettuce plants because they were not used as the source of carbon skeletons for nitrogen assimilation, while organic acids as well as glutamine and glutamate decreased.
Under sub-optimal environmental conditions, plants activate defense mechanisms for their survival [
73], producing a wide range of secondary metabolites with antioxidant activity with well-known benefits for human health [
4]. In this perspective, nutritional eustress is an excellent strategy to trigger gene activation involved in the biosynthesis of ascorbic acid and phenols, which improves the nutraceutical quality of vegetables [
74]. In agreement with the above, deprivation-induced nutritional stress three days before harvest, in comparison with control (0 DD), increased total phenols, total ascorbic acid, and thus hydrophilic (HAA) and ABTS antioxidant activities. It is worth noting that deprivation of the nutrient solution at 6 DD also resulted in a significant increase in ABTS content, in contrast to HAA. As suggested by El-Nakhel [
8] and Zhou [
4], the total phenols and total ascorbic acid concentration increase after macronutrient deprivation can be attributed to an upregulation of the enzymatic activity of phenylalanine ammonia-lyase (PAL) and L-galactose dehydrogenase (L-GalDH), respectively.
Because antioxidant compounds scavenge hydrogen peroxide (H
2O
2) [
75,
76], the non-increase in H
2O
2 levels in nutrient-deprived plants at 3 DD, compared with the control (0 DD), could be attributable to the increase in antioxidant activity, thus contributing to the scavenging of harmful reactive oxygen species (ROS). In contrast, at 6 DD, the opposite trend was observed, with a significant increase in H
2O
2 reflecting the HAA decrease. Interestingly, long-term nutrient deprivation (6 DD) did not increase total phenols, total ascorbic acid, and HAA than their values at 3 DD. Our results are corroborated by a previous study by Zhou [
4], in which a decrease in transcriptional levels of genes involved in regulating the biosynthesis of antioxidant compounds was observed after a long-term deprivation of nitrate [
4].
However, it is interesting that, in lettuce at 6 DD, the perturbation in H
2O
2 homeostasis can probably affect cytosolic pH inducing the decarboxylation of glutamate to produce GABA [
77,
78,
79]. Since, for the synthesis of this, non-protein amino acid protons are consumed, this process may enable the stationary control of pH. Moreover, GABA has a proved scavenging activity not only against H
2O
2, but also against superoxide anion radicals and singlet oxygen able to stabilize and protect thylakoids and ensure the viability and photosynthetic efficiency of the plants [
80,
81]. As reported by several authors, GABA levels increase in response to biotic and abiotic stresses, especially during nutrient starvation [
81,
82]. Specifically, GABA, after being rapidly synthetized for cell protection in response to stress—mainly functioning as a pH stabilizer and antioxidant—is broken down upon relief of stress to provide nitrogen, NADH, and or/anaplerotic succinate to the Krebs cycle for restoring the energy supply, which is crucial to guarantee that plant recovery and repair stress induced damage [
83]. Nonetheless, GABA is an important health-promoting functional compound that can prevent degenerative diseases; for this reason, diverse GABA-enriched foods are currently on the market [
84], and a lettuce naturally enriched with GABA could be an interesting premium quality produce. In addition, BCAAs also increased in lettuce at 6 DD, which have been proved to actively function as antioxidants not only in plants, but also in rats and mice by an unknown mechanism [
85].


 ,
, 




{kind=link}