
Characterization of Oilseed Crop Noug (Guizotia abyssinica) Using Agro-Morphological Traits

 ,
, 
 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
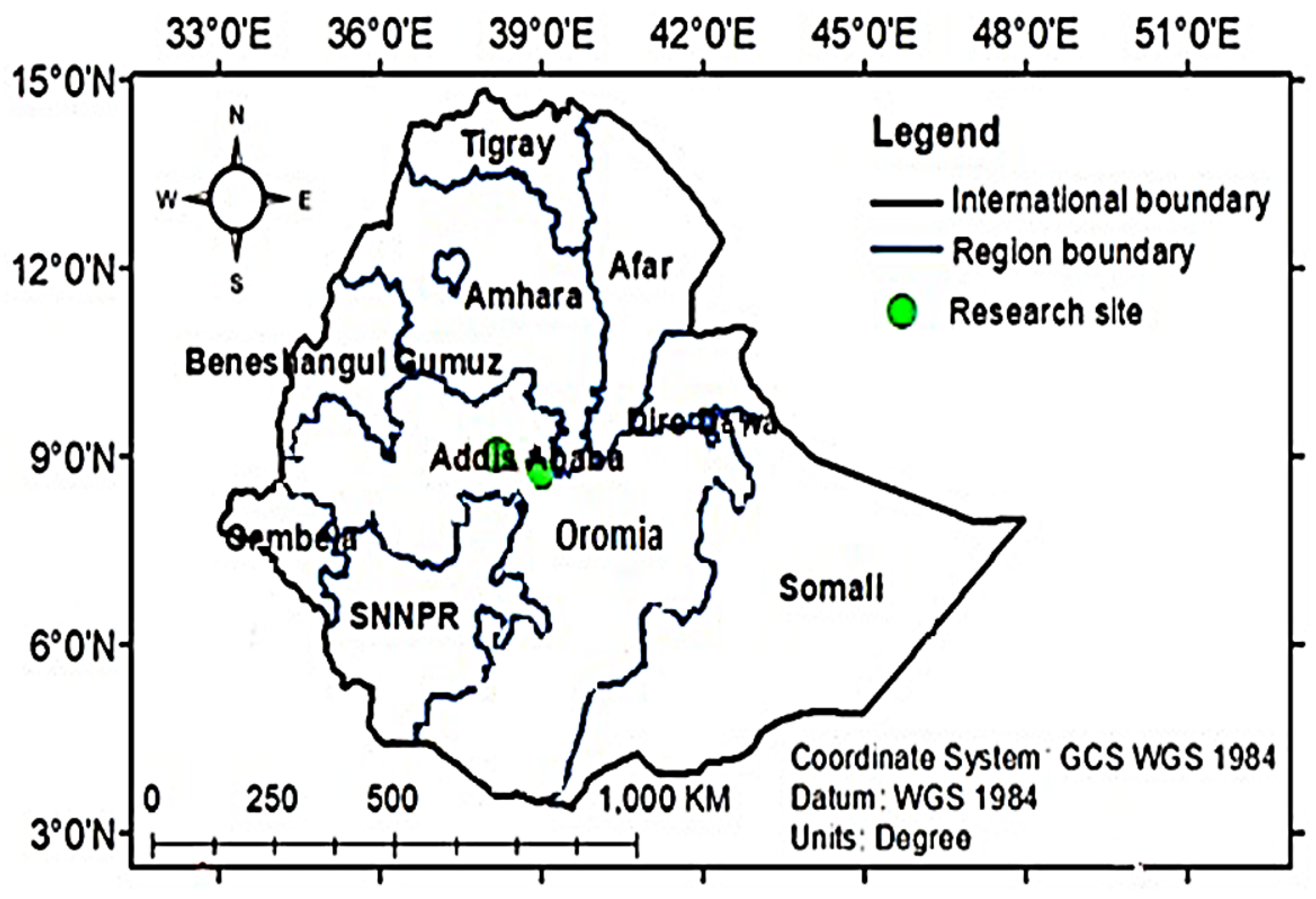
2.2. Experimental Sites and Design
2.3. Recording Phenotypic Data
2.4. Data Analysis
2.5. Estimation of Variance Components
2.6. Broad-Sense Heritability (H2) and Genetic Correlations as Computed by META-R
3. Results
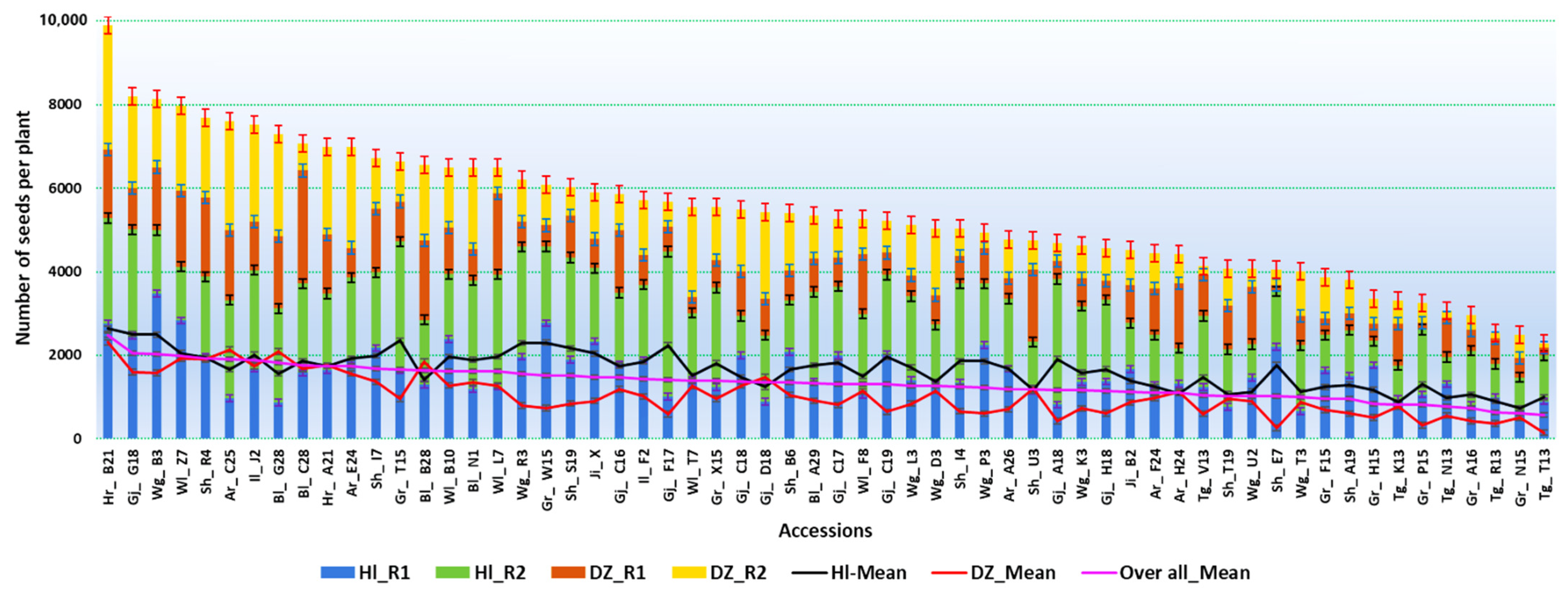
3.1. Analysis of Variance and Mean Performance of the Accessions
3.2. Genotypic and Phenotypic Correlation
3.3. Estimation of Variance Components and Broad-Sense Heritability
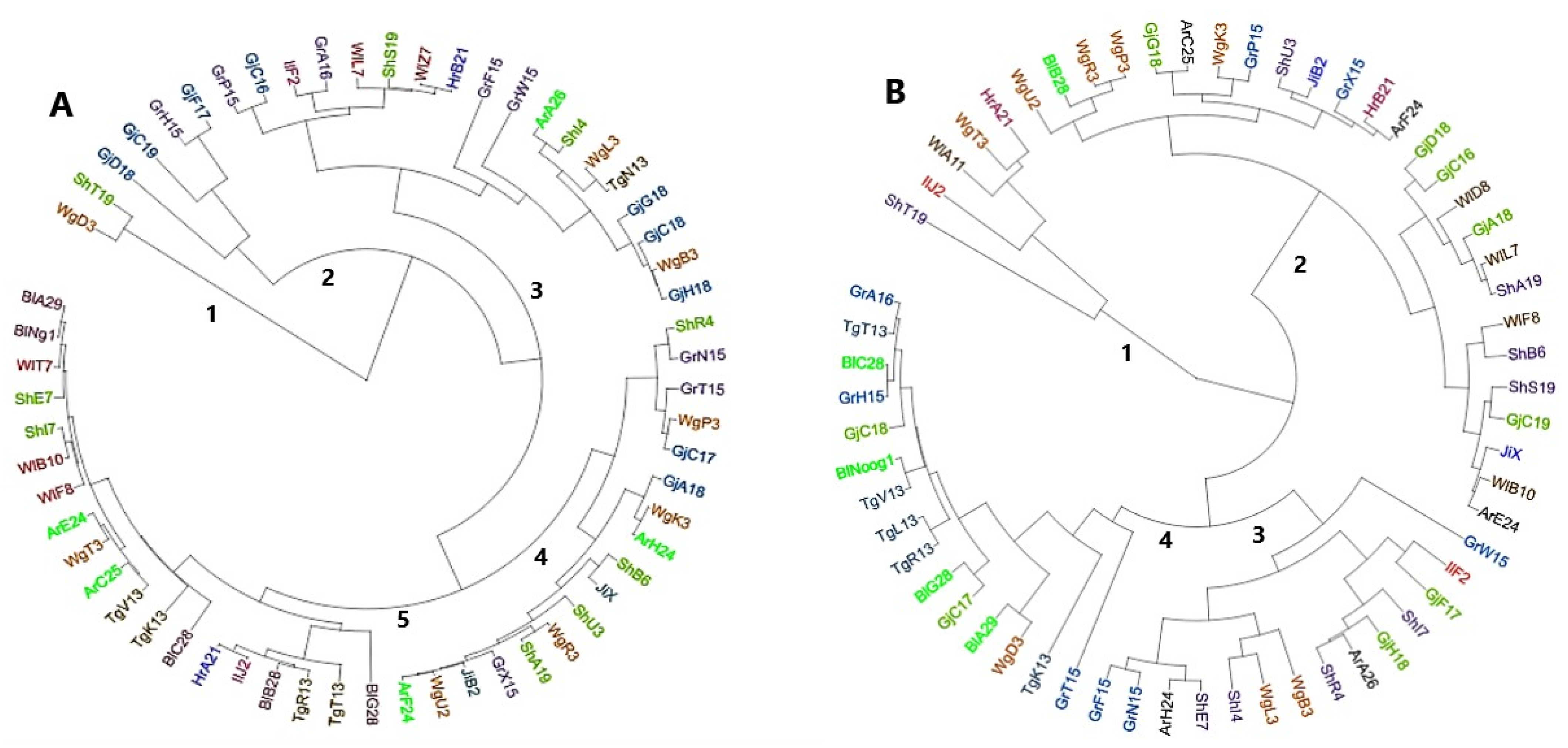
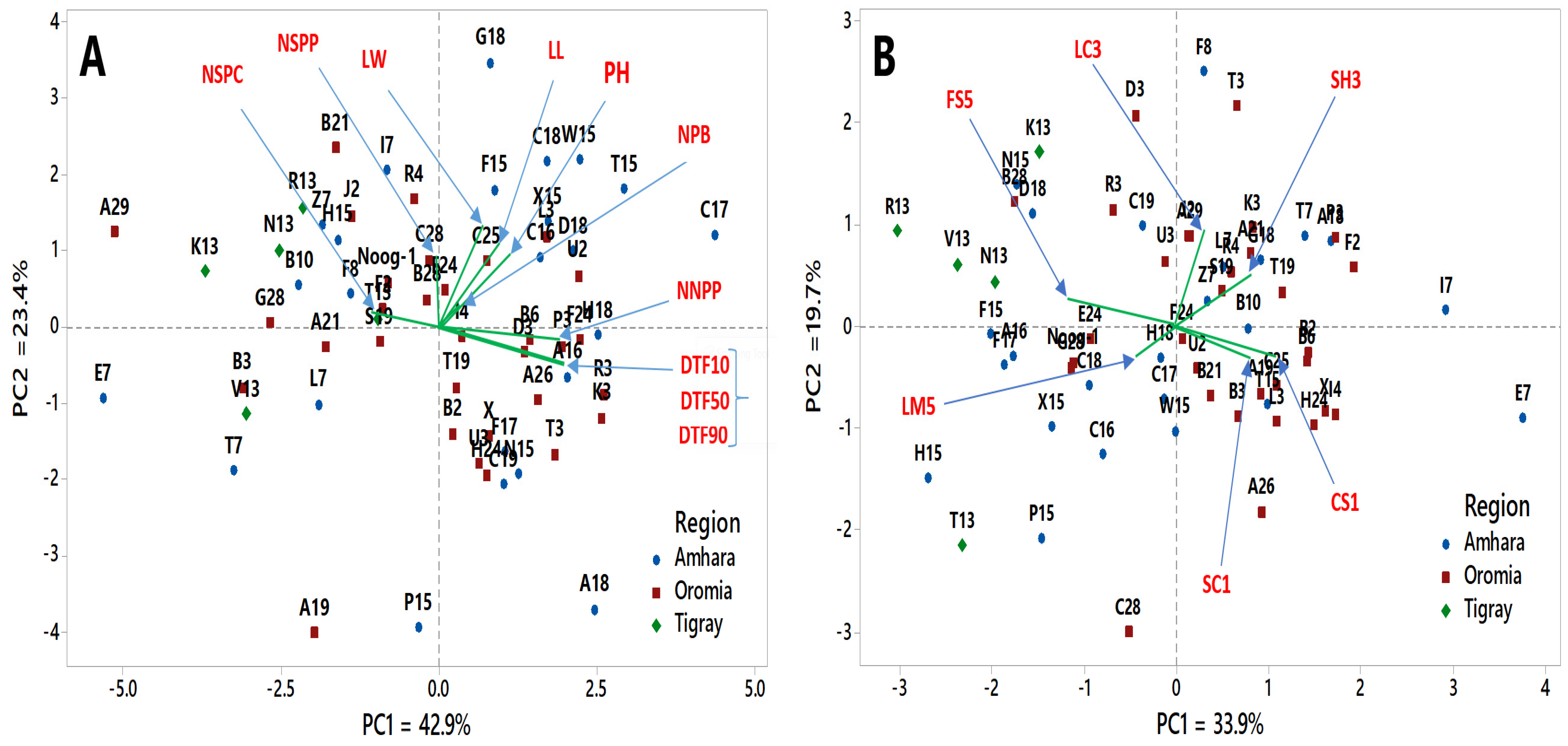
3.4. Cluster Analysis and Principal Component Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dagne, K. Meiosis in interspecific hybrids and genomic interrelationships in Guizotia Cass.(Compositae). Hereditas 1994, 121, 119–129. [Google Scholar] [CrossRef]
- Geleta, M.; Asfaw, Z.; Bekele, E.; Teshome, A. Edible oil crops and their integration with the major cereals in North Shewa and South Welo, Central Highlands of Ethiopia: An ethnobotanical perspective. Hereditas 2002, 137, 29–40. [Google Scholar] [CrossRef]
- Geleta, M.; Bryngelsson, T. Population genetics of self-incompatibility and developing self-compatible genotypes in niger (Guizotia abyssinica). Euphytica 2010, 176, 417–430. [Google Scholar] [CrossRef]
- Baagoe, J. The genus Guizotia (Compositae). A Taxonomic Revision. Bot. Tidsskr. 1974, 69, 1–39. [Google Scholar]
- Geleta, M.; Ortiz, R. The importance of Guizotia abyssinica (niger) for sustainable food security in Ethiopia. Genet. Resour. Crop Evol. 2013, 60, 1763–1770. [Google Scholar] [CrossRef]
- Seegeler, C.J.P. Oil Plants in Ethiopia: Their Taxonomy and Agricultural Significance; Center of Agricultural Publishing and Documentation: Wageningen, The Netherlands, 1983; pp. 122–146. [Google Scholar]
- Bulcha, W. Guizotia abyssinica (Lf) Cass. Record from PROTA4U; van der Vossen, H.A.M., Mkamilo, G.S., Eds.; PROTA (Plant Resources of Tropical Africa/Ressources végétales de l’Afrique tropicale): Wageningen, The Netherlands, 2007. [Google Scholar]
- Riley, K.; Belayneh, H. Niger Seed: Guizotia abyssinica Cass, in Oil Crops in the World: Their Breeding and Utilization; McGraw Hill Publishing Company: New York, NY, USA, 1989; pp. 394–403. [Google Scholar]
- Getinet, A.; Sharma, S. Niger, Guizotia abyssinica (Lf) Cass; Bioversity International: Rome, Italy, 1996; Volume 5. [Google Scholar]
- Oilseeds Research Strategy 2016–2023; Ethiopian Institute of Agricultural Research: Addis Ababa, Ethiopia, 2017.
- Petros, Y. Genetic Diversity and Oil Quality of Guizotia Cass (Asteraceae). Ph.D. Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2008. [Google Scholar]
- CSA, R. The Federal Democratic Republic of Ethiopia Report on Area and Production of Major Crops, Addis Ababa, Ethiopia, 2016.
- Geleta, M.; Bryngelsson, T.; Bekele, E.; Dagne, K. Assessment of genetic diversity of Guizotia abyssinica (Lf) Cass (Asteraceae) from Ethiopia using amplified fragment length polymorphism. Plant Genet. Resour. 2008, 6, 41–51. [Google Scholar] [CrossRef]
- Dempewolf, H.; Tesfaye, M.; Teshome, A.; Bjorkman, A.; Andrew, R.; Scascitelli, M.; Black, S.; Bekele, E.; Engels, J.M.M.; Cronk, Q.; et al. Patterns of domestication in the Ethiopian oil-seed crop noug (Guizotia abyssinica). Evol. Appl. 2015, 8, 464–475. [Google Scholar] [CrossRef] [PubMed]
- Geleta, M. Genetic Diversity, Phylogenetics and Molecular Systematics of Guizotia Cass (Asteraceae). Ph.D. Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2007. [Google Scholar]
- Geleta, M.; Bryngelsson, T.; Bekele, E.; Dagne, K. AFLP and RAPD analyses of genetic diversity of wild and/or weedy Guizotia (Asteraceae) from Ethiopia. Hereditas 2007, 144, 53–62. [Google Scholar] [CrossRef]
- Teklewold, A.; Wakjira, A. Seed filling and oil accumulation in noug [Guizotia abyssinica (LF) Cass.]. Sinet Ethiop. J. Sci. 2004, 27, 25–32. [Google Scholar]
- Adarsh, M.N.; Kumari, P.; Devi, S. A review of Guizotia abyssinica: A multipurpose plant with an economic prospective. J. Ind. Pollut. Control 2014, 30, 277–280. [Google Scholar]
- Kandel, H.J.; Porter, P.M.; Johnson, B.L.; Henson, R.A.; Hanson, B.K.; Weisberg, S.; LeGare, D.G. Plant population influences niger seed yield in the northern Great Plains. Crop Sci. 2004, 44, 190–197. [Google Scholar] [CrossRef]
- Hill, W.G. Understanding and using quantitative genetic variation. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 73–85. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Boddupalli, P.; Zou, C.; Lu, Y.; Xie, C.; Zhang, X.; Prasanna, B.M.; Olsen, M.S. Enhancing genetic gain in the era of molecular breeding. J. Exp. Bot. 2017, 68, 2641–2666. [Google Scholar] [CrossRef]
- Genet, T.; Belete, K. Phenotypic diversity in the Ethiopian noug germplasm. Afr. Crop Sci. J. 2000, 8, 137–143. [Google Scholar] [CrossRef]
- Sivasubramanian, S.; Menon, M. Heterosis and inbreeding depression in rice. Madras Agric. J. 1973, 60, 1139–1140. [Google Scholar]
- Robinson, H.F.; Comstock, R.E.; Harvey, P.H. Estimates of Heritability and the Degree of Dominance in Corn 1. Agron. J. 1949, 41, 353–359. [Google Scholar] [CrossRef]
- Alvarado, G.; Rodríguez, F.M.; Pacheco, A.; Burgueño, J.; Crossa, J.; Vargas, M.; Pérez-Rodríguez, P.; Lopez-Cruz, M.A. META-R: A software to analyze data from multi-environment plant breeding trials. Crop. J. 2020, 8, 745–756. [Google Scholar] [CrossRef]
- Buzatti, R.S.D.O.; Pfeilsticker, T.R.; Muniz, A.C.; Ellis, V.A.; De Souza, R.P.; Lemos-Filho, J.P.; Lovato, M.B. Disentangling the environmental factors that shape genetic and phenotypic leaf trait variation in the tree Qualea grandiflora across the Brazilian savanna. Front. Plant Sci. 2019, 10, 1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, M.; Deng, J.; Yao, B.; Chen, R.; Fan, Z.; Guan, J.; Li, X.; Wu, F.; Niklas, K.J. Ecogeographical variation of 12 morphological traits within Pinus tabulaeformis: The effects of environmental factors and demographic histories. J. Plant Ecol. 2017, 10, 386–396. [Google Scholar]
- Panda, S.; Sial, P. Assessment of existing genetic variability and yield component analysis in noug (Guizotia abyssinica (L.f.) Cass). Indian J. Innov. Dev. 2012, 1, 511–514. [Google Scholar]
- Amsalu, F. Genetic Variability Correlation and Path Coefficient Analysis of Niger Seed (Guizotia abyssinica [Lf] Cass.) Genotypes. Int. J. Res. Stud. Agric. Sci. 2020, 6, 8–16. [Google Scholar]
- Bhoite, K.D.; Kusalkar, D.V.; Patil, H.M. Genetic studies of yield and its component of Niger [(Guizotia abyssinica (Lf) Cass.] in Western Ghat Zone of Maharashtra. J. Pharmacogn. Phytochem. 2020, 9, 1182–1184. [Google Scholar]
- Ahmad, E.; Singh, M.N.; Paul, A.; Ansari, A.M.; Sigh, D.N. Genetic studies of yield and yield component of niger (Guizotia abyssinica Cass.) in rainfed condition of western plateau of Jharkhand. Int. J. Trop. Agric. 2016, 34, 1127. [Google Scholar]
- Tiwari, V.N.; Ahirwar, A.D.; Rai, G.K. Estimation of genetic parameters of variability for yield and its attributing traits in niger [Guizotia abyssinica (LF) Cass.]. Plant Arch. 2016, 16, 157–158. [Google Scholar]
- Wright, S. Correlation and causation. J. Agric. Res. 1921, 20, 557–580. [Google Scholar]
- Pulate, S.C.; Patil, H.S.; Patil, M.R. Multivariate analysis of genetic divergence among niger genotypes in relation to seed oil quality traits. Bioscan 2013, 8, 829–833. [Google Scholar]
- Geleta, M.; Bryngelsson, T.; Bekele, E.; Dagne, K. Comparative analysis of genetic relationship and diagnostic markers of several taxa of Guizotia Cass (Asteraceae) as revealed by AFLPs and RAPDs. Plant Syst. Evol. 2007, 265, 221–239. [Google Scholar] [CrossRef]
- Tsehay, S.; Ortiz, R.; Johansson, E.; Bekele, E.; Tesfaye, K.; Hammenhag, C.; Geleta, M. New Transcriptome-Based SNP Markers for Noug (Guizotia abyssinica) and Their Conversion to KASP Markers for Population Genetics Analyses. Genes 2020, 11, 1373. [Google Scholar] [CrossRef]
- Geleta, M.; Stymne, S.; Bryngelsson, T. Variation and inheritance of oil content and fatty acid composition in niger (Guizotia abyssinica). J. Food Compos. Anal. 2011, 24, 995–1003. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Trait Code | Description |
|---|---|---|
| Quantitative | ||
| Days to 10% flowering | DTF10 | Number of days from planting to the flowering of 10% of plants on a plot |
| Days to 50% flowering | DTF50 | Number of days from planting to the flowering of 50% of plants on a plot |
| Days to 90% flowering | DTF90 | Number of days from planting to the flowering of 90% of plants on a plot |
| Leaf length | LL | The length of leaves in the middle part of a stem measured from base to tip in cm (average of five leaves per plant) |
| Leaf width | LW | The width (at the center) of leaves in the middle part of a stem measured in cm (average of five leaves per plant) |
| Number of capitula per plant | NCPP | Total number of capitula on each plant (count) |
| Number of nodes per plant | NNPP | Total number of nodes on each plant (count) |
| Number of primary branches | NPB | Total number of primary branches on each plant (count) |
| Number of seeds per capitulum | NSPC | An average number of seeds per capitulum, calculated based on five capitula from each plant |
| Plant height | PH | Height of a plant measured in cm at full flowering stage |
| Qualitative/Categorical | ||
| Flower size | FS | Three categories were given based on the diameter of fully developed flowers: 1 = small (<3 cm); 3 = medium (3 to 4 cm); 5 = large (>4 cm) |
| Capitulum size | CS | Three categories were given based on the cross-sectional circumference of mature capitulum: 1 = small (<3 cm); 3 = medium (3 to 4 cm); 5 = large (>4 cm) |
| Leaf color | LC | Three categories were given based on the intensity of green color of mature leaves: 1= light green; 3 = green; 5 = deep green |
| Leaf margin | LM | Three categories were given based on the indent size of the margins of mature leaves: 1 = fine; 3 = medium; 5 = course |
| Stem color | SC | Two categories were given based on the color of the middle portion of a stem: 1 = green; 2 = purple |
| Stem hairiness | SH | Three categories were given based on the level of hairiness of the middle part of stems: 1 = no or little observable hair; 3 = medium; 5 = clearly visible dense hair |
| Traits | Sources of Variation | Coefficient of Variation (%) | |||
|---|---|---|---|---|---|
| MSG (59) | MSE (1) | MSG*E (59) | MSR (120) | ||
| DTF10 | 179.2 *** | 18,357.5 *** | 22.1 * | 14.4 | 4.9 |
| DTF50 | 188.0 *** | 20,813.4 *** | 26.9 *** | 12.6 | 4.4 |
| DTF90 | 185.8 *** | 21,606.8 *** | 33.4 *** | 12.7 | 4.2 |
| NNPP | 4.7 *** | 97.3 *** | 0.7 ** | 0.4 | 6.1 |
| PH | 610.0 *** | 91.2 | 370.1 | 290.4 | 18.1 |
| NPB | 9.0 *** | 18.5 * | 4.8 | 3.9 | 18.1 |
| NCPP | 243.5 | 7695.4 *** | 235 | 188 | 35.1 |
| LL | 0.6 * | 16.3 *** | 0.4 | 0.4 | 24.3 |
| LW | 1.2 | 3.2 | 1.2 | 1.0 | 33.4 |
| NSPC | 103.7 * | 20,419.2 *** | 104.0 * | 63.0 | 12.3 |
| Character | Pearson χ2 | LR χ2 | p-Value | Acc | DZ_R1 | DZ_R2 | HL_R1 | HL_R2 | All | |
|---|---|---|---|---|---|---|---|---|---|---|
| CS-1 | 1629.7 | 1802.6 | <0.001 | Gj_D18 | Obs. freq. | 0 | 2 | 6 | 0 | 8 |
| Exp. freq. | 1.0 | 1.8 | 2.7 | 2.5 | ||||||
| χ2 | 1.0 | 0.01 | 4.1 | 2.5 | ||||||
| Sh_E7 | Obs. freq. | 78 | 71 | 78 | 90 | 317 | ||||
| Exp. freq. | 41.0 | 70.0 | 106.5 | 99.5 | ||||||
| χ2 | 33.4 | 0.01 | 7.6 | 0.9 | ||||||
| FS-5 | 1162.3 | 1251.9 | <0.001 | Sh_I7 | Obs. freq. | 16 | 0 | 5 | 8 | 29 |
| Exp. freq. | 4.7 | 6.5 | 8.9 | 8.9 | ||||||
| χ2 | 27.1 | 6.5 | 1.7 | 0.1 | ||||||
| Tg_R13 | Obs. freq. | 55 | 48 | 94 | 95 | 292 | ||||
| Exp. freq. | 47.4 | 65.7 | 89.3 | 89.6 | ||||||
| χ2 | 1.22 | 4.79 | 0.25 | 0.33 | ||||||
| LC-3 | 227.9 | 231.1 | <0.01 | Tg_T13 | Obs. freq. | 44 | 62 | 44 | 62 | 212 |
| Exp, freq. | 51.3 | 50.8 | 55.6 | 54.3 | ||||||
| χ2 | 1.0 | 2.4 | 2.4 | 1.1 | ||||||
| Wg_T3 | Obs. freq. | 100 | 100 | 100 | 100 | 400 | ||||
| Exp. freq. | 96.8 | 95.9 | 104.9 | 102.4 | ||||||
| χ2 | 0.1 | 0.2 | 0.2 | 0.1 | ||||||
| LM-5 | 804.9 | 881.8 | <0.001 | Sh_E7 | Obs. freq. | 0 | 6 | 3 | 0 | 9 |
| Exp. freq. | 2.3 | 2.0 | 2.3 | 2.3 | ||||||
| χ2 | 2.3 | 7.9 | 0.2 | 2.3 | ||||||
| Gj_C17 | Obs. freq. | 84 | 62 | 88 | 62 | 296 | ||||
| Exp. freq. | 76.79 | 66.08 | 77.43 | 75.69 | ||||||
| χ2 | 0.68 | 0.25 | 1.44 | 2.48 | ||||||
| SC-1 | 817.1 | 926.6 | <0.001 | Tg_K13 | Obs. freq. | 8 | 33 | 10 | 0 | 51 |
| Exp. freq. | 12.3 | 13.0 | 12.6 | 13.1 | ||||||
| χ2 | 1.5 | 30.7 | 0.5 | 13.1 | ||||||
| Ar_H24 | Obs. freq. | 70 | 64 | 72 | 53 | 259 | ||||
| Exp. freq. | 62.5 | 66.1 | 64.1 | 66.7 | ||||||
| χ2 | 0.9 | 0.1 | 1.0 | 2.7 | ||||||
| SH-3 | 706.6 | 772.3 | <0.001 | Wg_B3 | Obs. freq. | 8 | 17 | 4 | 23 | 52 |
| Exp. freq. | 13.1 | 13.1 | 13.1 | 12.7 | ||||||
| χ2 | 2.0 | 1.1 | 6.3 | 8.4 | ||||||
| Wl_F8 | Obs. freq. | 90 | 86 | 52 | 56 | 284 | ||||
| Exp. freq. | 71.8 | 71.6 | 71.5 | 69.1 | ||||||
| χ2 | 4.6 | 2.9 | 5.3 | 2.5 |
| Traits | DTF10 | DTF50 | DTF90 | NNPP | PH | NPB | NCPP | LL | LW | NSPC |
|---|---|---|---|---|---|---|---|---|---|---|
| DTF10 | 0.91 *** | 0.94 *** | 0.59 *** | 0.22 | 0.38 | −0.04 | 0.23 | −0.06 | −0.23 *** | |
| DTF50 | 0.99 *** | 0.95 *** | 0.57 *** | 0.23 * | 0.36 | −0.03 | 0.24 | −0.08 | −0.30 ** | |
| DTF90 | 0.99 *** | 0.99 *** | 0.57 *** | 0.25 * | 0.35 | −0.10 | 0.21 | −0.06 | −0.30 *** | |
| NNPP | 0.68 *** | 0.72 *** | 0.74 *** | 0.41 *** | 0.73 *** | 0.06 | 0.32 * | −0.01 | −0.09 * | |
| PH | 0.25 *** | 0.38 | 0.39 | 0.44 *** | 0.50 | 0.27 * | 0.65 *** | 0.63 *** | −0.11 | |
| NPB | 0.51 | 0.55 *** | 0.52 *** | 0.89 *** | 0.29 *** | 0.43 | 0.53 ** | 0.37 | 0.12 | |
| NCPP | 0.11 *** | 0.60 *** | 0.24 *** | 0.36 *** | 0.65 *** | 0.99 *** | 0.35 *** | 0.25 *** | 0.24 * | |
| LL | 0.38 | 0.57 *** | 0.47 | 0.48 *** | 0.72 *** | 0.48 *** | 0.33 *** | 0.66 *** | −0.11 | |
| LW | −0.14 *** | 0.01 | −0.03 *** | −0.26 *** | 0.50 | −0.21 *** | 0.40 *** | 0.42 *** | −0.03 | |
| NSPC | −0.36 *** | −0.61 *** | −0.54 *** | −0.05 *** | −0.08 *** | 0.41 *** | −0.77 *** | −0.04 *** | 0.49 *** |
| Traits | Minimum | Maximum | Mean ± SD | Genotypic Variance | Phenotypic Variance | H2 (%) | GCV (%) | PCV (%) |
|---|---|---|---|---|---|---|---|---|
| DTF10 | 60.00 | 102.0 | 77.45 ± 3.80 | 0.2914 | 0.3413 | 85.38 | 0.69 | 0.75 |
| DTF50 | 63.00 | 107.0 | 81.00 ± 3.60 | 0.2031 | 0.2823 | 71.94 | 0.55 | 0.65 |
| DTF90 | 65.00 | 109.0 | 84.07 ± 3.60 | 0.2195 | 0.2844 | 77.14 | 0.56 | 0.63 |
| NNPP | 6.60 | 13.82 | 10.05 ± 0.61 | 0.4468 | 0.5676 | 78.71 | 6.65 | 7.49 |
| PH | 29.60 | 144.7 | 87.15 ± 13.71 | 0.1586 | 0.5575 | 28.45 | 1.02 | 1.91 |
| NPB | 5.10 | 16.82 | 10.85 ± 1.97 | 0.1611 | 0.4234 | 38.05 | 3.69 | 5.99 |
| NCPP | 10.78 | 87.08 | 94.22 ± 17.04 | 0.1808 | 0.4691 | 38.54 | 0.45 | 0.72 |
| LL | 1.04 | 4.66 | 2.54 ± 0.62 | 0.0506 | 0.3050 | 16.59 | 8.85 | 21.74 |
| LW | 1.00 | 6.00 | 3.01 ± 1.01 | 0.1383 | 1.2228 | 11.31 | 12.35 | 36.73 |
| NSPC | 39.40 | 96.92 | 64.52 ± 7.94 | 0.1154 | 0.3649 | 31.62 | 0.53 | 0.93 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gebeyehu, A.; Hammenhag, C.; Ortiz, R.; Tesfaye, K.; Geleta, M. Characterization of Oilseed Crop Noug (Guizotia abyssinica) Using Agro-Morphological Traits. Agronomy 2021, 11, 1479. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081479
Gebeyehu A, Hammenhag C, Ortiz R, Tesfaye K, Geleta M. Characterization of Oilseed Crop Noug (Guizotia abyssinica) Using Agro-Morphological Traits. Agronomy. 2021; 11(8):1479. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081479
Chicago/Turabian StyleGebeyehu, Adane, Cecilia Hammenhag, Rodomiro Ortiz, Kassahun Tesfaye, and Mulatu Geleta. 2021. "Characterization of Oilseed Crop Noug (Guizotia abyssinica) Using Agro-Morphological Traits" Agronomy 11, no. 8: 1479. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081479