
Short-Term Responses to Salinity of Soybean and Chenopodium album Grown in Single and Mixed-Species Hydroponic Systems

Abstract
:1. Introduction
2. Materials and Methods
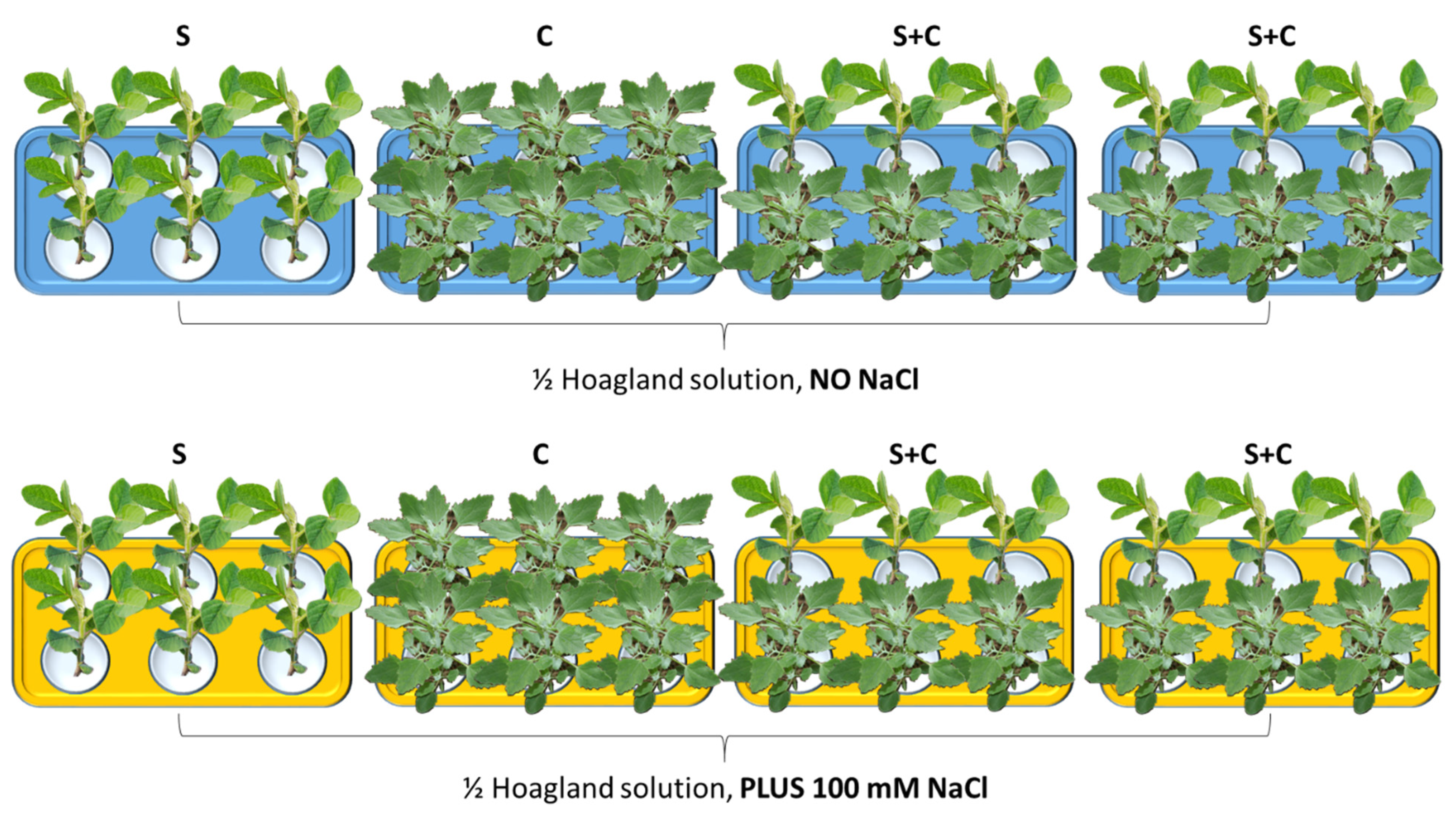
2.1. Plant Growth Conditions and Experimental Design
2.2. Soluble Protein Quantification
2.3. Elemental Content Quantification
2.4. Determination of Total Antioxidant Activity and Phenol Content
2.5. Antioxidant Enzyme Activity
2.6. Lipid Peroxidation
2.7. Proline Quantification
2.8. Statistical Analysis
3. Results
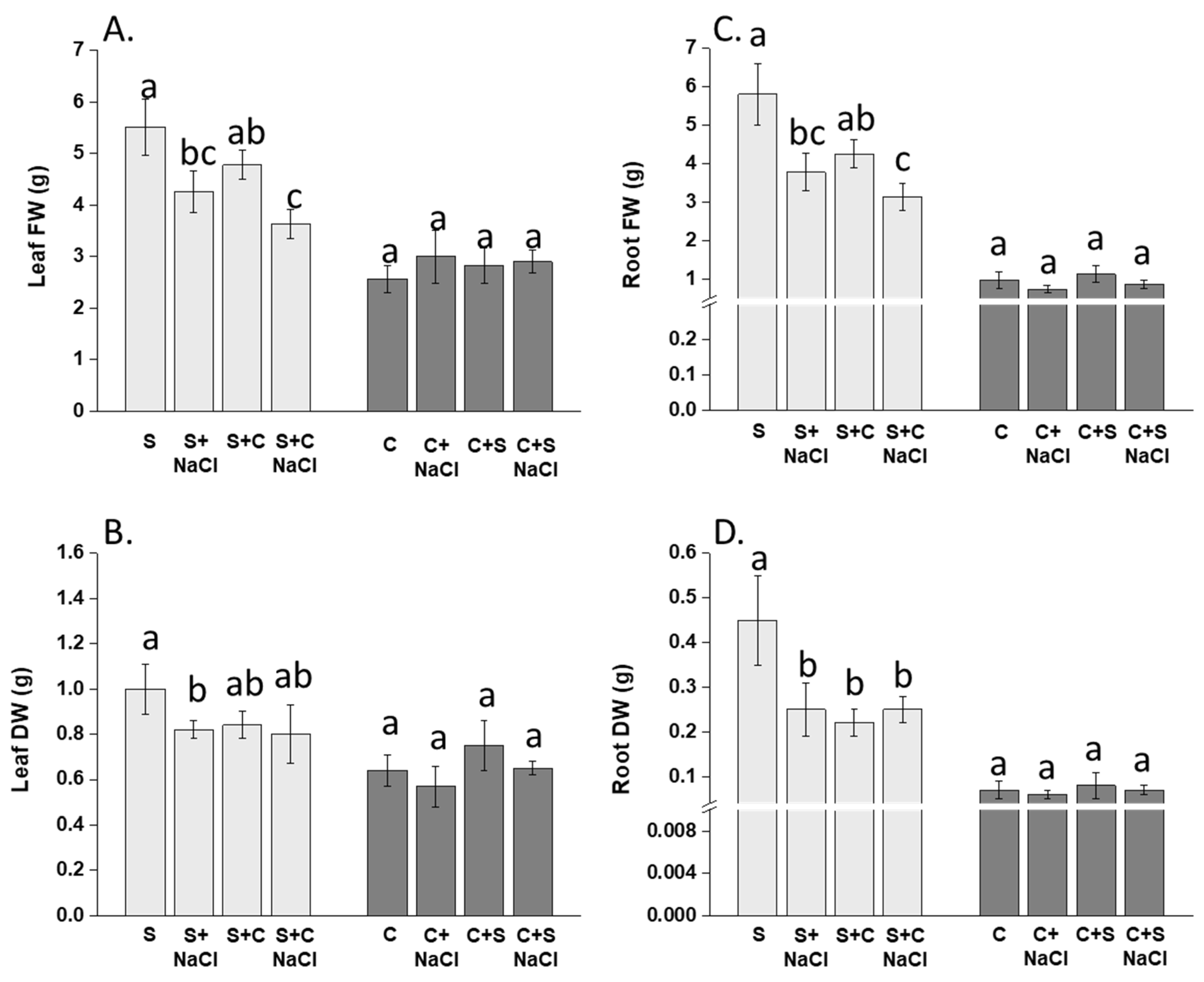
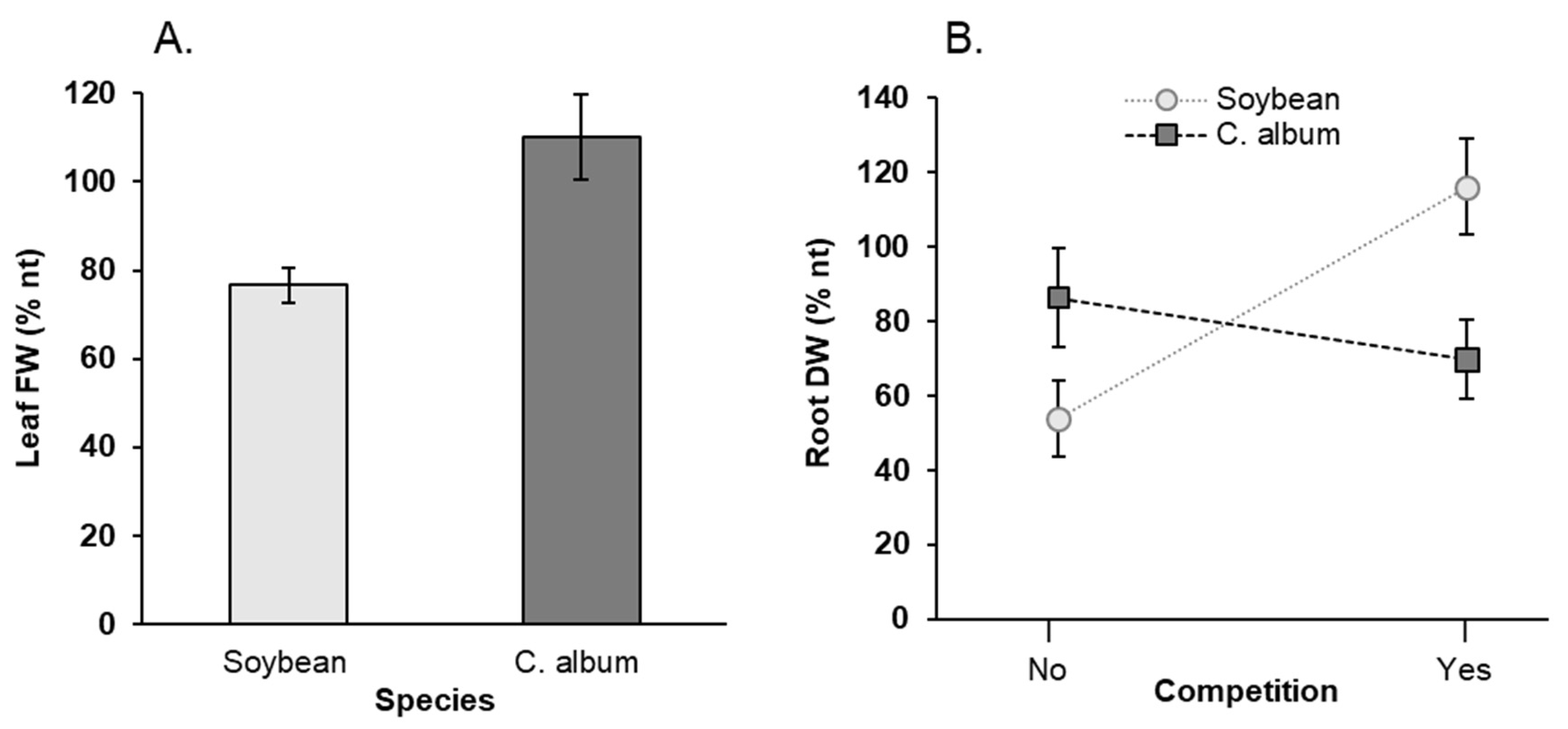
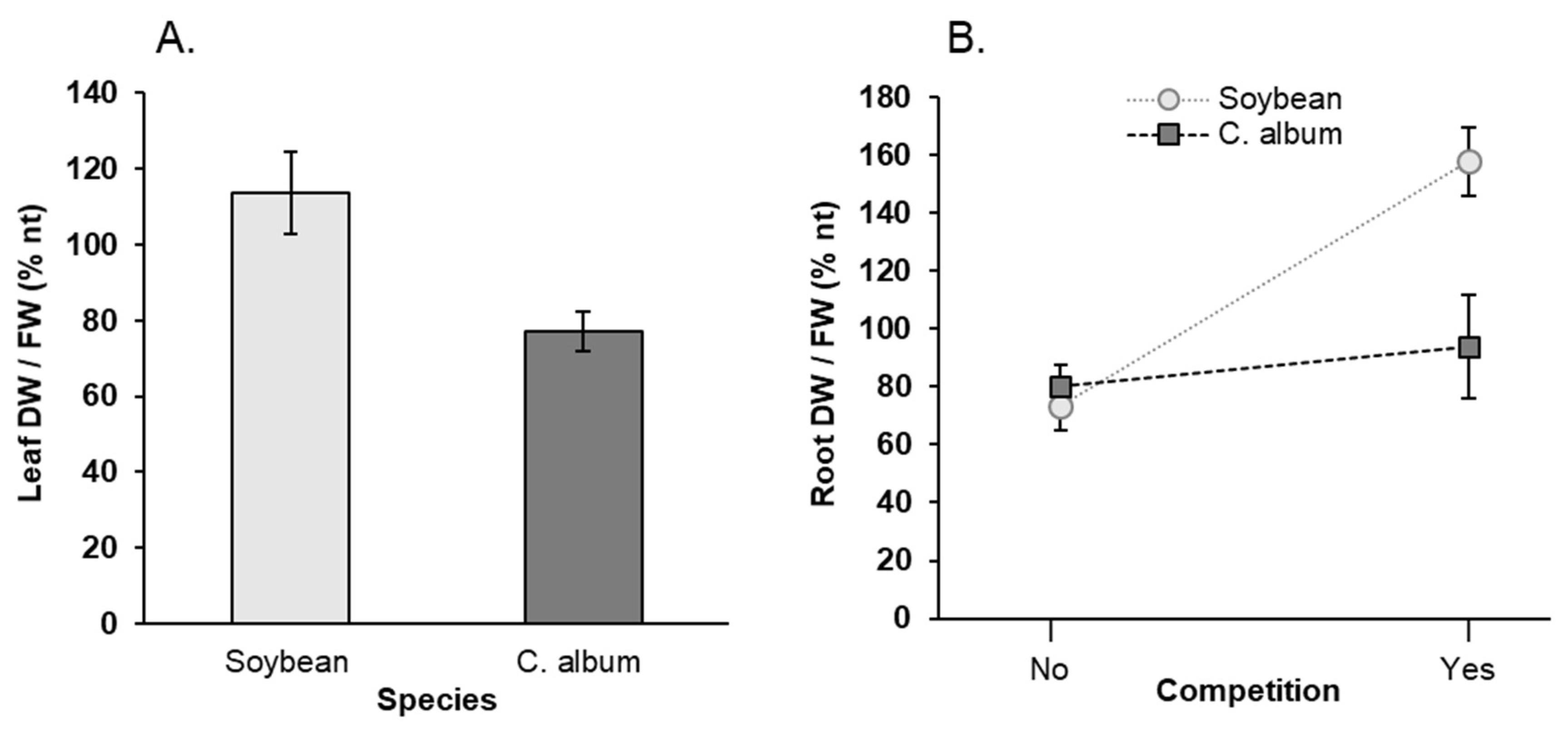
3.1. Effect of NaCl, Plant-Competition and Combination of Both on Plant Biomass
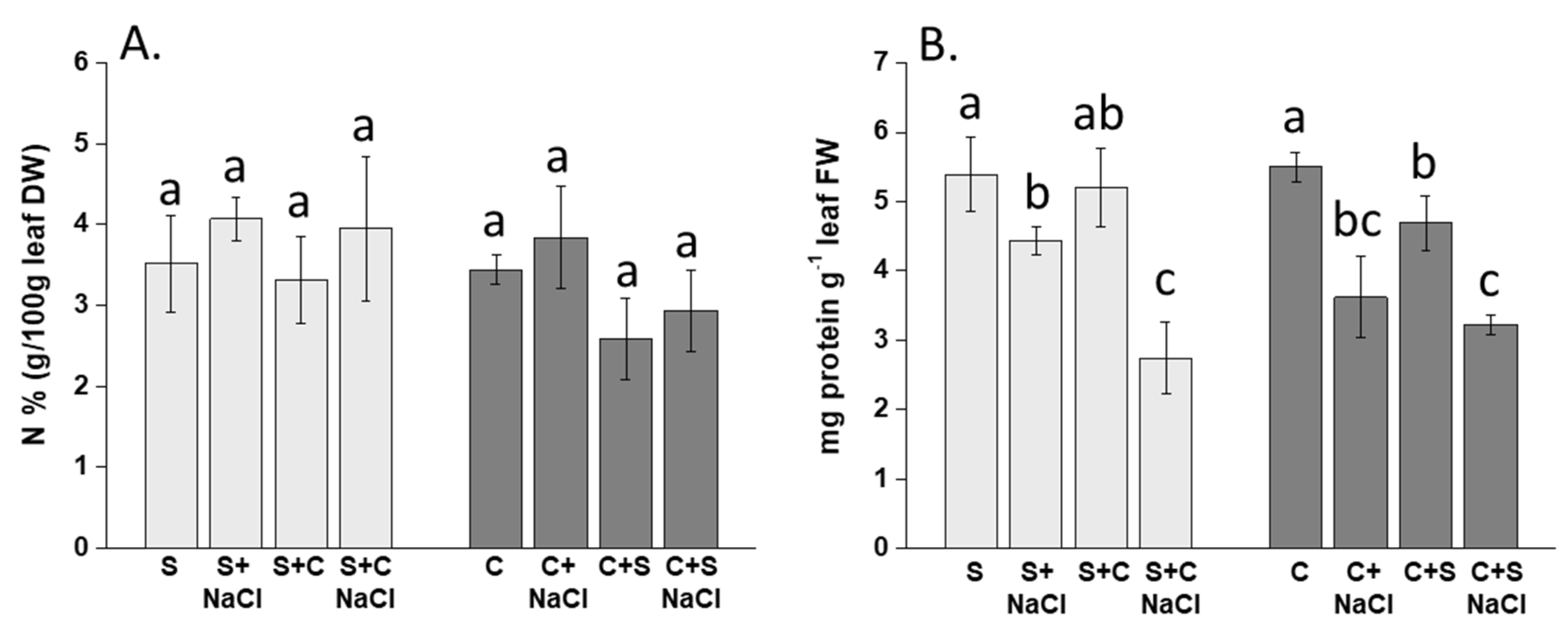
3.2. Effect of NaCl, Plant-Competition and Combination of Both on the Content of N and Soluble Proteins
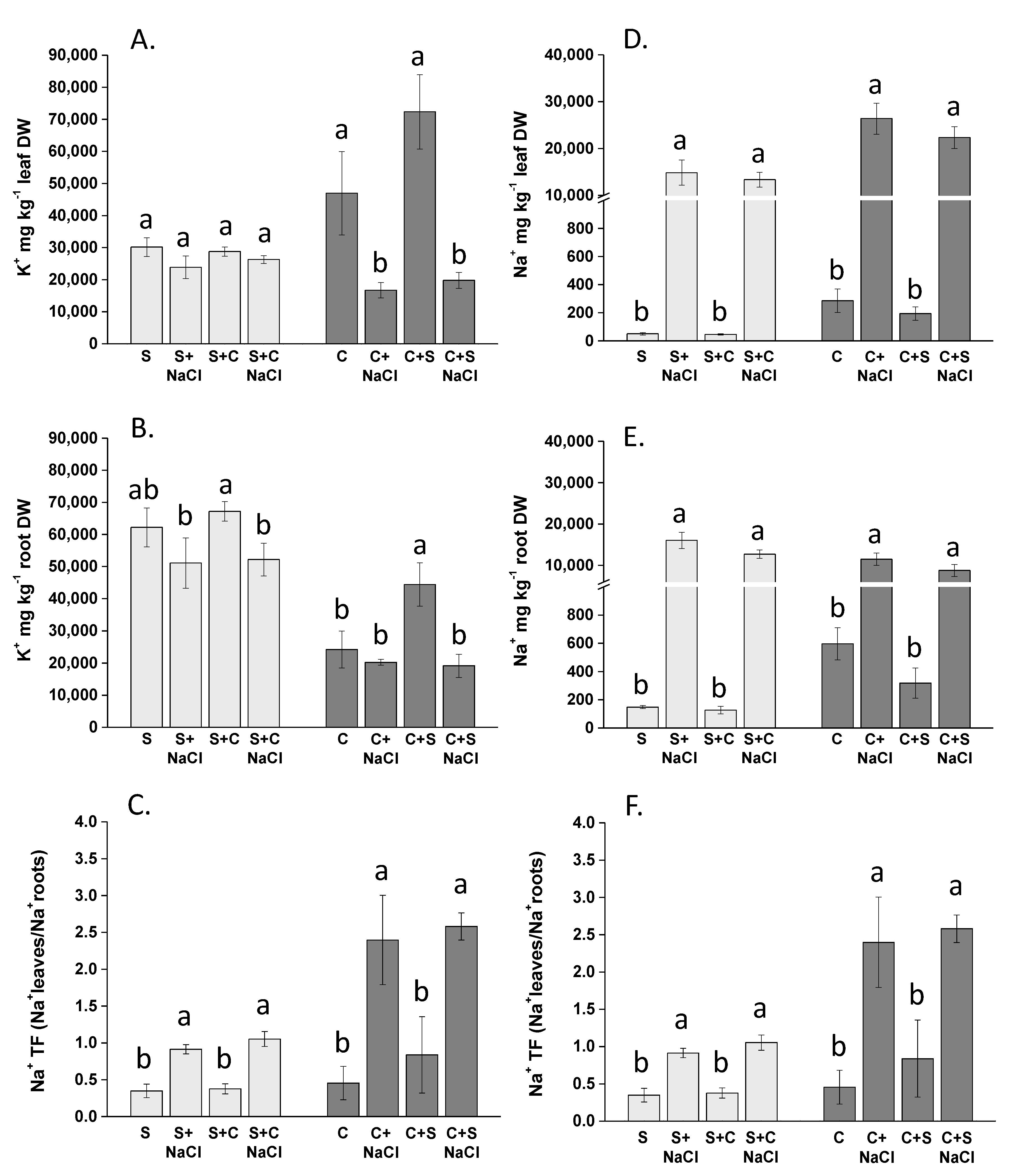
3.3. Effect of NaCl, Plant-Competition and Combination of Both on Na+ and K+ Accumulation
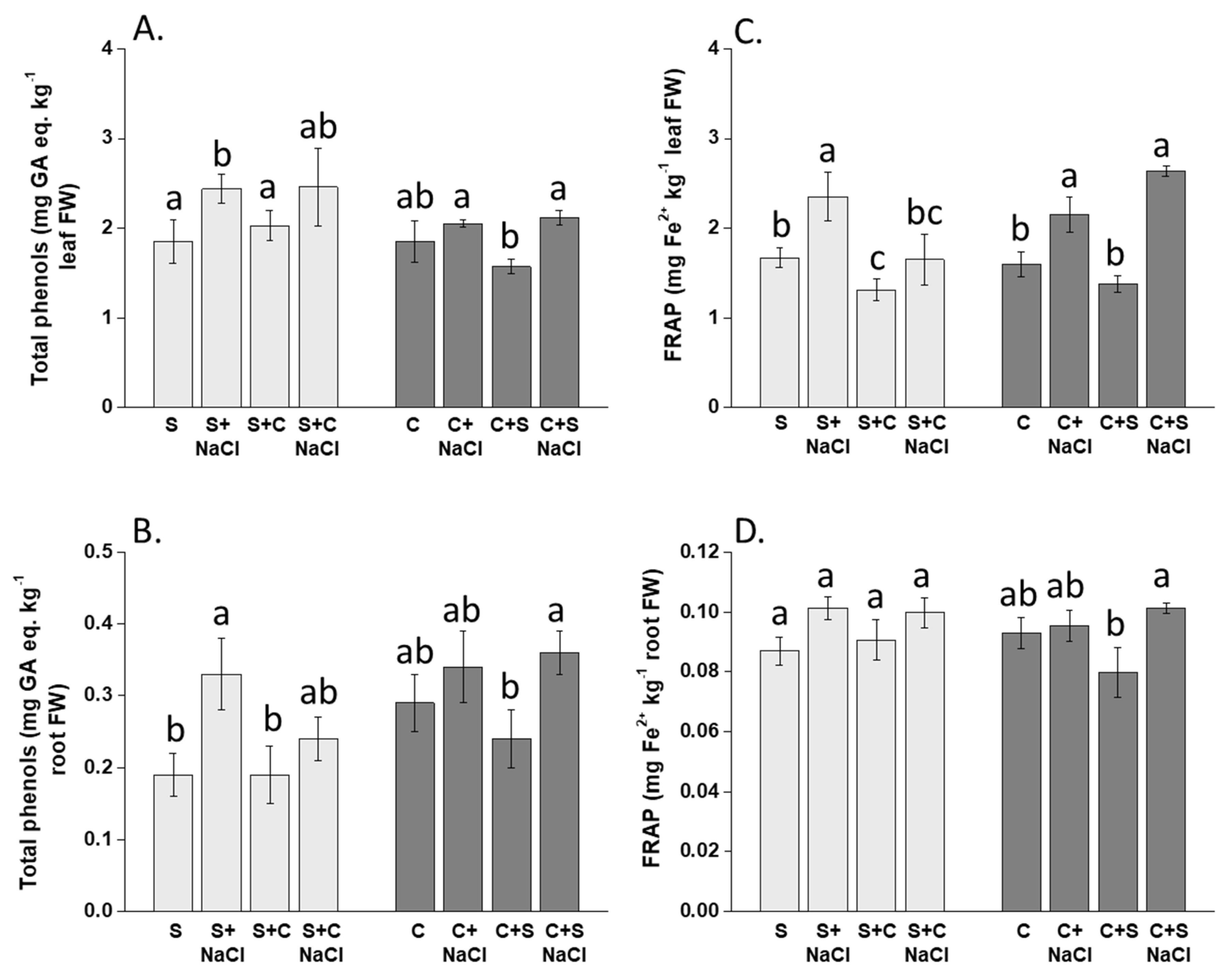
3.4. Effect of NaCl, Plant-Competition and Combination of Both on the Content of Phenolic Compounds and Plant Antioxidant Capacity (FRAP)
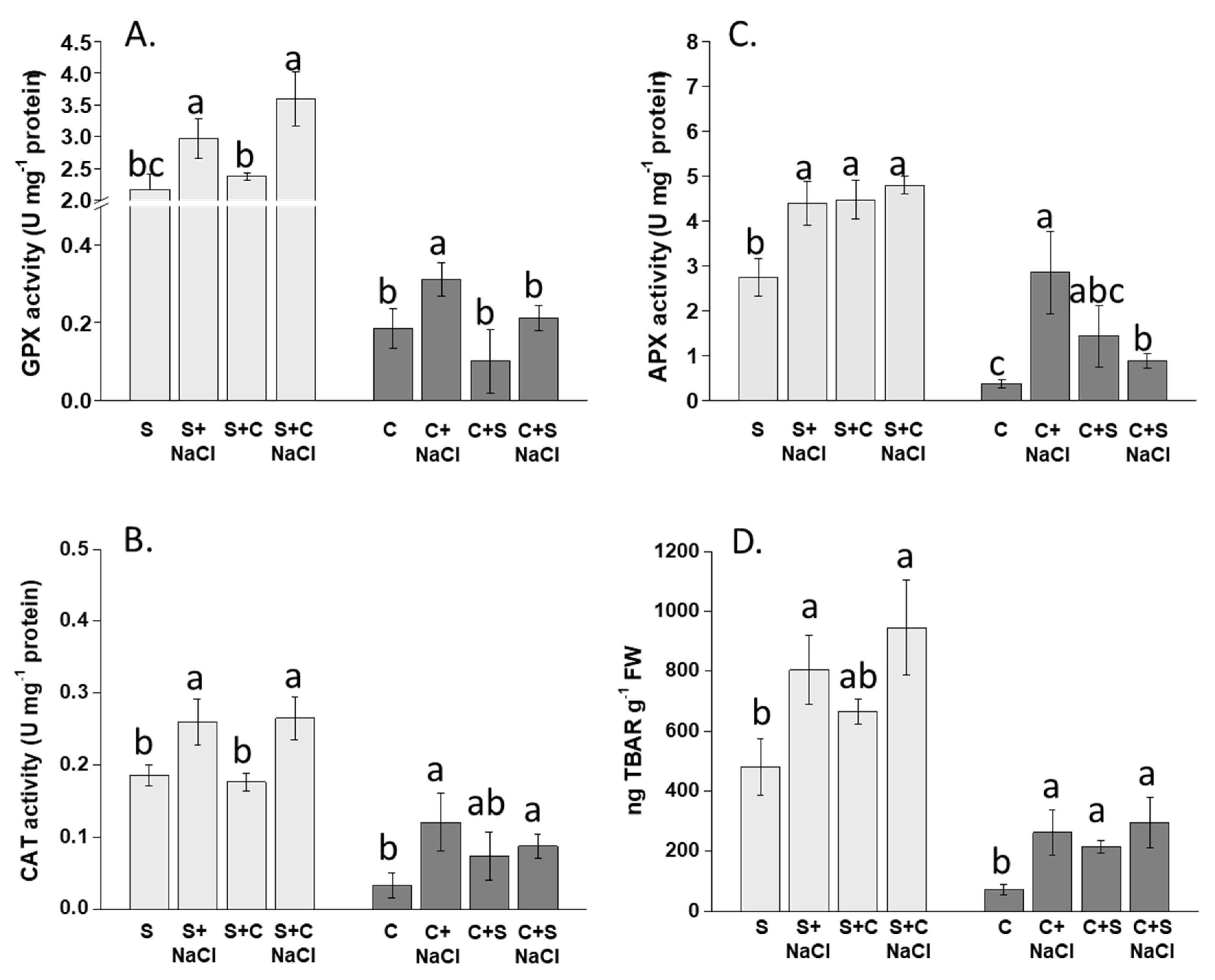
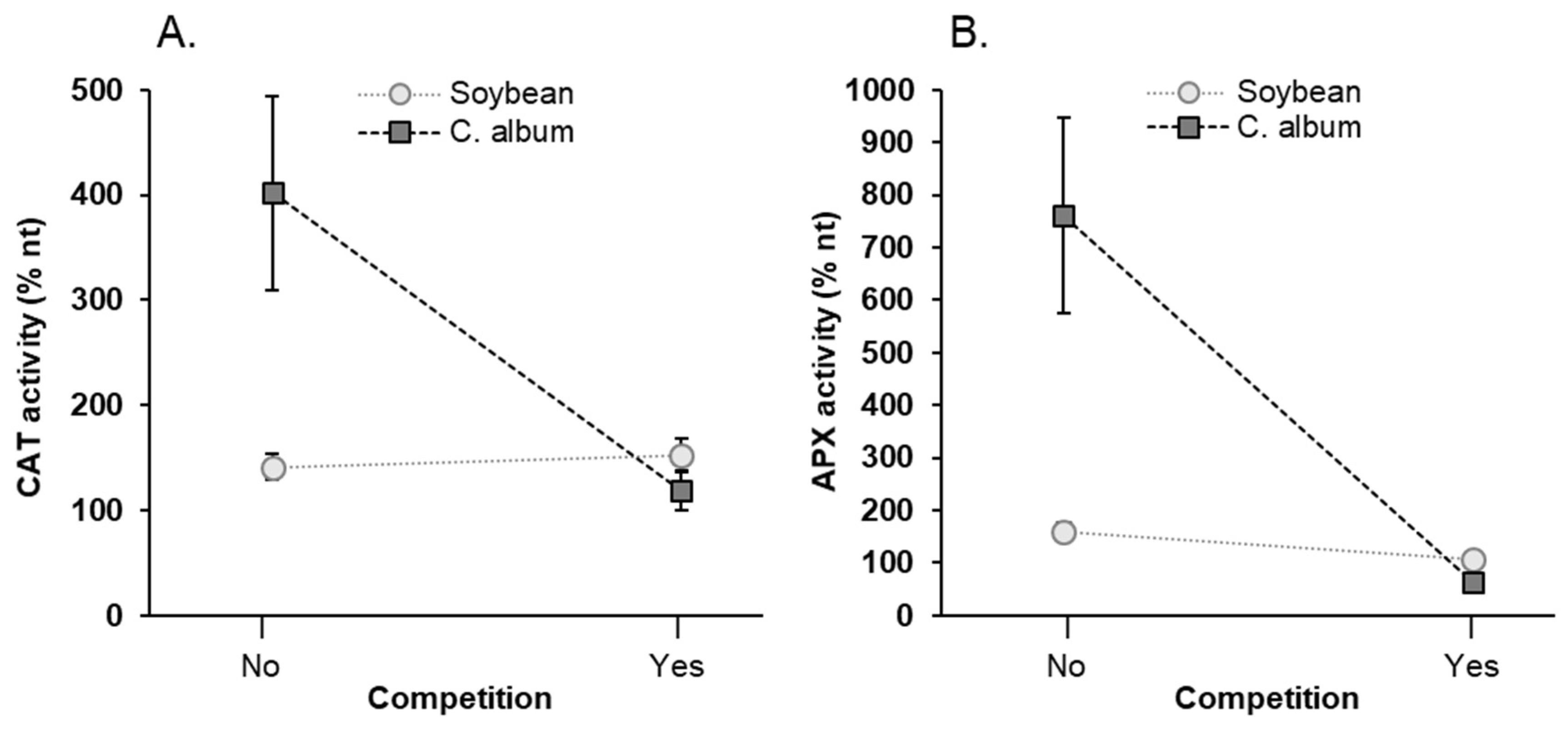
3.5. Effect of NaCl, Plant-Competition and Combination of Both on Antioxidant Enzyme Activity (GPX, APX, CAT) and Lipid Peroxidation
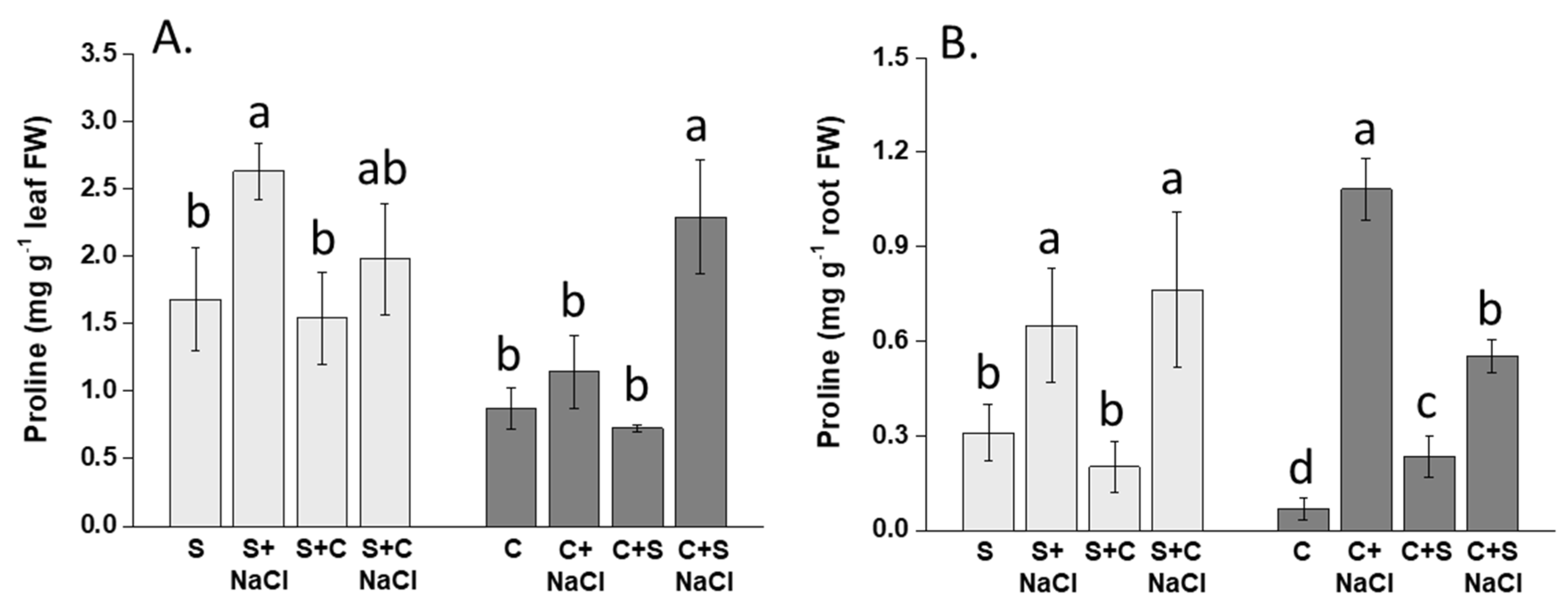
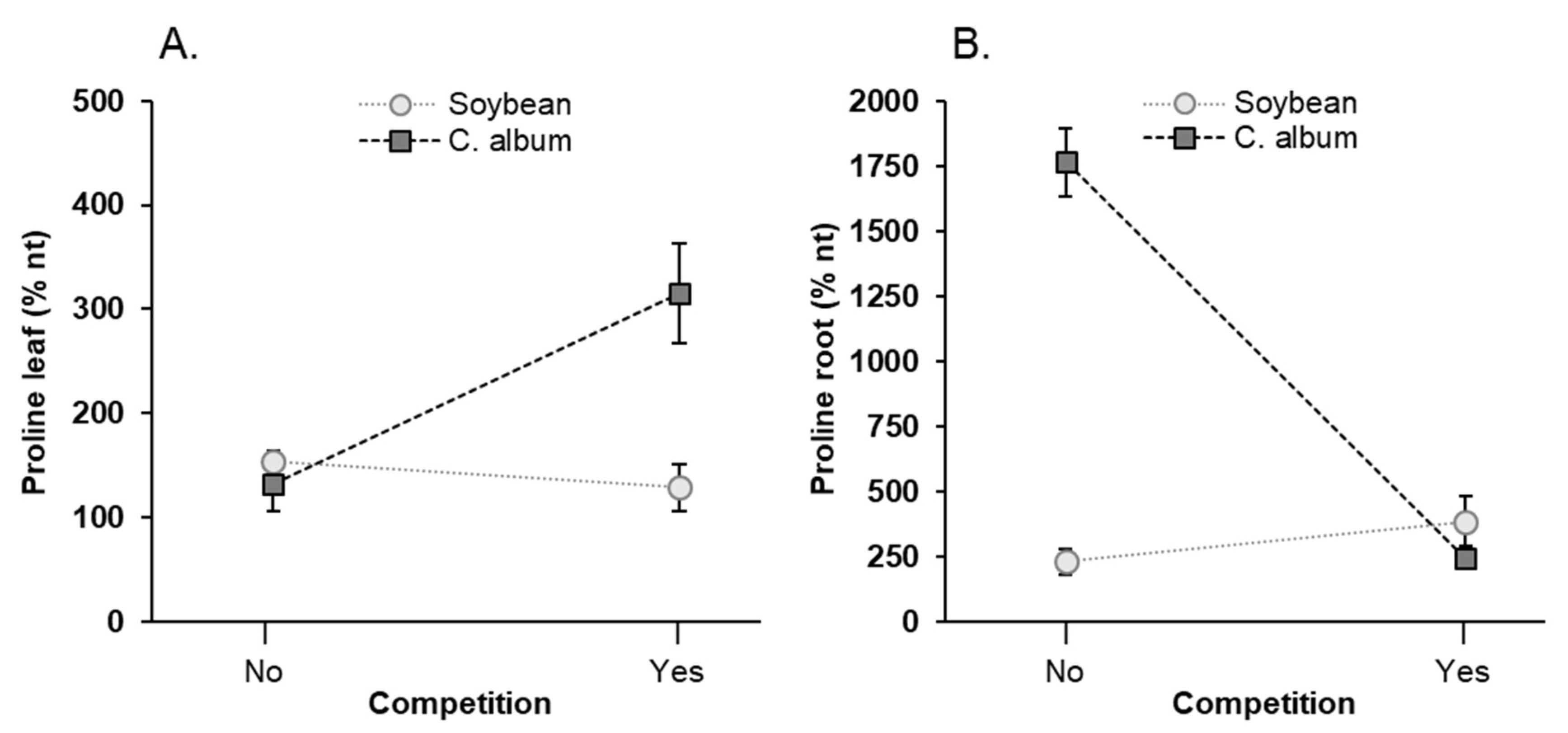
3.6. Effect of NaCl, Plant-Competition and Combination of Both on Proline Accumulation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Almeida, D.M.; Margarida Oliveira, M.; Saibo, N.J.M. Regulation of Na+ and K+ homeostasis in plants: Towards improved salt stress tolerance in crop plants. Genet. Mol. Biol. 2017, 40, 326–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corwin, D.L. Climate change impacts on soil salinity in agricultural areas. Eur. J. Soil Sci. 2021, 72, 842–862. [Google Scholar] [CrossRef]
- Jamil, A.; Riaz, S.; Ashraf, M.; Foolad, M.R. Gene expression profiling of plants under salt stress. CRC Crit. Rev. Plant Sci. 2011, 30, 435–458. [Google Scholar] [CrossRef]
- Taylor, R.G.; Scanlon, B.; Döll, P.; Rodell, M.; van Beek, R.; Wada, Y.; Longuevergne, L.; Leblanc, M.; Famiglietti, J.S.; Edmunds, M.; et al. Ground water and climate change. Nat. Clim. Chang. 2013, 3, 322–329. [Google Scholar] [CrossRef] [Green Version]
- Phang, T.-H.; Shao, G.; Lam, H.-M. Salt Tolerance in Soybean. J. Integr. Plant Biol. 2008, 50, 1196–1212. [Google Scholar] [CrossRef]
- Zeng, L.; Shannon, M.C. Salinity effects on seedling growth and yield components of rice. Crop Sci. 2000, 40, 996–1003. [Google Scholar] [CrossRef] [Green Version]
- Daei, G.; Ardekani, M.R.; Rejali, F.; Teimuri, S.; Miransari, M. Alleviation of salinity stress on wheat yield, yield components, and nutrient uptake using arbuscular mycorrhizal fungi under field conditions. J. Plant Physiol. 2009, 166, 617–625. [Google Scholar] [CrossRef]
- Katerji, N.; Van Hoorn, J.W.; Hamdy, A.; Karam, F.; Mastrorilli, M. Effect of salinity on water stress, growth, and yield of maize and sunflower. Agric. Water Manag. 1996, 30, 237–249. [Google Scholar] [CrossRef]
- Khan, M.H.; Panda, S.K. Alterations in root lipid peroxidation and antioxidative responses in two rice cultivars under NaCl-salinity stress. Acta Physiol. Plant. 2008, 30, 81–89. [Google Scholar] [CrossRef]
- Zhu, J.K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef]
- Niu, X.; Bressan, R.A.; Hasegawa, P.M.; Pardo, J.M. lon Homeostasis in NaCl stress environments. Plant Physiol 1995, 109, 735–742. [Google Scholar] [CrossRef] [Green Version]
- Xing, J.; Cai, M.; Chen, S.; Chen, L.; Lan, H. Seed germination, plant growth and physiological responses of Salsola ikonnikovii to short-term NaCl stress. Plant Biosyst. 2013, 147, 285–297. [Google Scholar] [CrossRef]
- Garg, N.; Manchanda, G. ROS generation in plants: Boon or bane? Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2009, 143, 81–96. [Google Scholar] [CrossRef]
- Jbir-Koubaa, R.; Charfeddine, S.; Ellouz, W.; Saidi, M.N.; Drira, N.; Gargouri-Bouzid, R.; Nouri-Ellouz, O. Investigation of the response to salinity and to oxidative stress of interspecific potato somatic hybrids grown in a greenhouse. Plant Cell. Tissue Organ Cult. 2015, 120, 933–947. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive Oxigen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [PubMed] [Green Version]
- McKersie, B.D.; Leshem, Y.Y. Stress and Stress Coping in Cultivated Plants; Springer: Dordrecht, The Netherlands, 1994. [Google Scholar]
- Chang, R.Z.; Chen, Y.W.; Shao, G.H.; Wan, C.W. Effect of salt stress on agronomic characters and chemical quality of seeds in soybean. Soybean Sci. 1994, 13, 101–105. [Google Scholar]
- Cirillo, V.; Masin, R.; Maggio, A.; Zanin, G. Crop-weed interactions in saline environments. Eur. J. Agron. 2018, 99, 51–61. [Google Scholar]
- Lu, P.; Li, J.; Jin, C.; Jiang, B.; Bai, Y. Different growth responses of an invasive weed and a native crop to nitrogen pulse and competition. PLoS ONE 2016, 11, e0156285. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.H.; Gols, R.; Benrey, B. Crop domestication and its impact on naturally selected trophic interactions. Annu. Rev. Entomol. 2015, 60, 35–58. [Google Scholar] [CrossRef] [Green Version]
- Clements, D.R.; Ditommaso, A.; Jordan, N.; Booth, B.D.; Cardina, J.; Doohan, D.; Onguevergne, L.; Leblanc, M.; Famiglietti, J.S.; Edmunds, M.; et al. Adaptability of plants invading North American cropland. Ecosyst. Environ. 2004, 104, 379–398. [Google Scholar] [CrossRef]
- Tanveer, M.; Shah, A.N. An insight into salt stress tolerance mechanisms of Chenopodium album. Environ. Sci. Pollut. Res. 2017, 24, 16531–16535. [Google Scholar] [CrossRef]
- Kraehmer, H.; Baur, P. Weed Anatomy, 1st ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2013. [Google Scholar]
- Sartorato, I.; Berti, A.; Zanin, G. Estimation of economic thresholds for weed control in soybean (Glycine max (L.) Merr.). Crop Prot. 1996, 15, 63–68. [Google Scholar] [CrossRef]
- Shurtleff, J.L.; Coble, H.D. Interference of certain broadleaf weed species in soybeans (Glycine max). Weed Sci. 1985, 33, 654–657. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the quantitation of microgram quantities of protein utilizing the principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Nicoletto, C.; Santagata, S.; Bona, S.; Sambo, P. Influence of cut number on qualitative traits in different cultivars of sweet basil. Ind. Crops Prod. 2013, 44, 465–472. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Rezaie, F.; Yarnia, M. Allelopathic effects of Chenopodium album, Amaranthus retroflexus and Cynodon dactylon on germination and growth of safflower. J. Food Agric. Environ. 2005, 7, 516–521. [Google Scholar]
- Reinhardt, C.F.; Meissner, R.; Labuschagne, N. Allelopathic interaction between Chenopodium album L. and certain crop species. S. Afr. J. Plant Soil 1994, 11, 45–49. [Google Scholar] [CrossRef]
- Alam, S.M.; Azmi, A.R.; Naqvi, S.S.M.; Khan, M.A.; Khanzada, B. Effect of aqueous leaf extract of common lambsquarters (Chenopodium album L.) and NaCl on germination and seedling growth of rice. Acta Physiol. Plant. 1997, 19, 91–94. [Google Scholar] [CrossRef]
- Mallik, M.A.B.; Puchala, R.; Grosz, F.A. A growth inhibitory factor from lambsquarters (Chenopodium album). J. Chem. Ecol. 1994, 20, 957–967. [Google Scholar] [CrossRef]
- Namvar, A.; Seyed Sharifi, R.; Molaei Igdir Üniversitesi, P. Influence of extracts of Chenopodium album and NaCl salinity on germination and seedling growth of soybean. Allelopath. J. 2009, 23, 193–202. [Google Scholar]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.; Chen, S.; Xu, D.; Lan, H. Plant growth and responses of antioxidants of Chenopodium album to long-term NaCl and KCl stress. Plant Growth Regul. 2010, 60, 115–125. [Google Scholar] [CrossRef]
- Wu, H.; Shabala, L.; Zhou, M.; Su, N.; Wu, Q.; Ul-Haq, T.; Zhu, J.; Mancuso, S.; Azzarello, E.; Shabala, S. Root vacuolar Na+ sequestration but not exclusion from uptake correlates with barley salt tolerance. Plant J. 2019, 100, 55–67. [Google Scholar] [CrossRef]
- Osmond, C.B.; Björkman, O.; Anderson, D.J. Physiological Processes in Plant Ecology. Toward a Synthesis with Atriplex; Springer: Berlin/Heidelberg, Germany, 1980. [Google Scholar]
- Wu, H.; Zhang, P.; Chen, G.; Pan, L.; Li, J.; Dong, L. Environmental factors on seed germination and seedling emergence of Phleum paniculatum Huds. Chil. J. Agric. Res. 2018, 78, 370–377. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Pottosin, I.I.; Cuin, T.A.; Fuglsang, A.T.; Tester, M.; Jha, D.; Zepeda-Jazo, I.; Zhou, M.; Palmgren, M.G.; Newman, I.A.; et al. Root plasma membrane transporters controlling K+/Na+ homeostasis in salt-stressed barley. Plant Physiol. 2007, 145, 1714–1725. [Google Scholar] [CrossRef] [Green Version]
- Ai-Rawahy, S.A.; Stroehlein, J.L.; Pessarakli, M. Dry-matter yield and nitrogen-15, Na+, and K+ content of tomatoes under sodium chloride stress. J. Plant Nutr. 1992, 15, 341–358. [Google Scholar] [CrossRef]
- Hamada, A.M.; El-Enany, A.E. Effect of NaCl salinity on growth, pigment and mineral element contents, and gas exchange of broad bean and pea plants. Biol. Plant. 1994, 36, 75–81. [Google Scholar] [CrossRef]
- Bulut, F.; Akıncı, Ş. The effect of salinity on growth and nutrient composition in broad bean (Vicia faba L.) seedlings Al and humic acid interactions on broad bean seedlings. Fresenius Environ. Bull. 2010, 19, 2901–2910. [Google Scholar]
- Tester, M.; Davenport, R. Na+ Tolerance and Na+ Transport in Higher Plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef]
- Debouba, M.; Gouia, H.; Suzuki, A.; Ghorbel, M.H. NaCl stress effects on enzymes involved in nitrogen assimilation pathway in tomato “Lycopersicon esculentum” seedlings. J. Plant Physiol. 2006, 163, 1247–1258. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Song, J.; Dong, L.; Wang, D.; Zhang, S.; Liu, J. Physiological responses of three soybean species (Glycine soja, G. gracilis, and G. max cv. Melrose) to salinity stress. J. Plant Res. 2017, 130, 723–733. [Google Scholar] [CrossRef]
- Osman, M.S.; Badawy, A.A.; Osman, A.I.; Abdel Latef, A.A.H. Ameliorative impact of an extract of the halophyte Arthrocnemum macrostachyum on growth and biochemical parameters of soybean under salinity stress. J. Plant Growth Regul. 2020, 9, 1–12. [Google Scholar]
- Heuer, B. Role of proline in plant response to drought and salinity. In Handbook of Plant and Crop Stress; Pessarakli, M., Ed.; CRC Press: Boca Raton, FL, USA, 2010; pp. 213–238. ISBN 9781439813997. [Google Scholar]
- Haq, N.U.; Ammar, M.; Bano, A.; Luthe, D.S.; Heckathorn, S.A.; Shakeel, S.N. Molecular Characterization of Chenopodium album chloroplast small heat shock protein and its expression in response to different abiotic stresses. Plant Mol. Biol. Report. 2013, 31, 1230–1241. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Progressive Number | Treatment | Acronym |
|---|---|---|
| 1 1 | Soybean No NaCl | S |
| 2 | Soybean plus 100 mM NaCl | S+NaCl |
| 3 2 | Soybean and C. album No NaCl | S+C |
| 4 | Soybean and C. album plus 100 mM NaCl | S+C+NaCl |
| 5 | C. album No NaCl | C |
| 6 | C. album plus 100 mM NaCl | C+NaCl |
| 7 2 | C. album and Soybean No NaCl | C+S |
| 8 | C. album and Soybean plus 100 mM NaCl | C+S+NaCl |
| A | Leaves | Roots | Leaves | Roots | Leaves | Roots |
| FW | FW | DW | DW | DW/FW | DW/FW | |
| Species | 0.005 | ns | ns | ns | 0.007 | 0.028 |
| Competition | ns | ns | ns | ns | ns | 0.001 |
| Species × Competition | ns | ns | ns | 0.003 | ns | 0.008 |
| B | leaves | leaves | ||||
| N tot | Proteins | |||||
| Species | ns | ns | ||||
| Competition | ns | ns | ||||
| Species × Competition | ns | 0.038 | ||||
| C | leaves | leaves | roots | roots | Na leaves/ | K leaves/ |
| Na | K | Na | K | Na roots | K roots | |
| Species | 0 | 0 | 0 | 0.045 | 0.047 | 0 |
| Competition | ns | ns | ns | 0.011 | ns | 0.043 |
| Species × Competition | ns | ns | ns | 0.02 | ns | ns |
| D | leaves | roots | leaves | roots | ||
| Phenols | Phenols | FRAP | FRAP | |||
| Species | ns | ns | 0.01 | ns | ||
| Competition | ns | ns | ns | ns | ||
| Species × Competition | ns | 0.012 | 0.012 | 0.003 | ||
| E | leaves | leaves | leaves | leaves | ||
| MDA | GPX | APX | CAT | |||
| Species | ns | 0.038 | 0.017 | 0.047 | ||
| Competition | 0.031 | ns | 0.004 | 0.023 | ||
| Species × Competition | ns | ns | 0.008 | 0.016 | ||
| F | leaves | roots | ||||
| Proline | Proline | |||||
| Species | 0.024 | 0 | ||||
| Competition | 0.028 | 0 | ||||
| Species × Competition | 0.008 | 0 |
| Treatment | Na+/K+ | |
|---|---|---|
| Leaves | Roots | |
| Soybean No NaCl | 0.002 ± 0.000 d | 0.002 ± 0.000 d |
| Soybean plus 100 mM NaCl | 0.618 ± 0.028 b | 0.324 ± 0.029 b |
| Soybean and C. album No NaCl | 0.002 ± 0.000 d | 0.002 ± 0.000 d |
| Soybean and C. album plus 100 mM NaCl | 0.506 ± 0.038 c | 0.249 ± 0.043 b |
| C. album No NaCl | 0.008 ± 0.004 d | 0.028 ± 0.012 c |
| C. album plus 100 mM NaCl | 1.601 ± 0.177 a | 0.569 ± 0.076 a |
| C. album and Soybean No NaCl | 0.003 ± 0.000 d | 0.007 ± 0.002 d |
| C. album and Soybean plus 100 mM NaCl | 1.156 ± 0.205 a | 0.483 ± 0.117 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghirardelli, A.; Schiavon, M.; Zanin, G.; Ostapczuk, P.; Masin, R. Short-Term Responses to Salinity of Soybean and Chenopodium album Grown in Single and Mixed-Species Hydroponic Systems. Agronomy 2021, 11, 1481. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081481
Ghirardelli A, Schiavon M, Zanin G, Ostapczuk P, Masin R. Short-Term Responses to Salinity of Soybean and Chenopodium album Grown in Single and Mixed-Species Hydroponic Systems. Agronomy. 2021; 11(8):1481. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081481
Chicago/Turabian StyleGhirardelli, Aurora, Michela Schiavon, Giuseppe Zanin, Piotr Ostapczuk, and Roberta Masin. 2021. "Short-Term Responses to Salinity of Soybean and Chenopodium album Grown in Single and Mixed-Species Hydroponic Systems" Agronomy 11, no. 8: 1481. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081481