1. Introduction
Tef (also written as “teff”) (
Eragrostis tef (
Zucc.) Trotter) is an annual small grain, panicle bearing, C
4 cereal crop native to Ethiopia, where it is a major staple food, accounting for two thirds of the daily protein intake of the population, and 11% of the caloric intake [
1]. It is resistant to many biotic and abiotic stresses, making it an attractive crop for smallholder farmers [
1]. It is known for its many health benefits, including its low glycemic index and high concentrations of essential amino acids, and it is also gluten free [
2].
The majority of tef cultivation is located in Ethiopia [
1], and it is mostly cultivated under low input, rainfed conditions However, in recent years interest has increased in cultivating tef outside of its native environment [
1], including drylands where irrigation is required. Interest has also increased in cultivating tef in a more intensive manner [
1,
3]. One way of intensifying tef cultivation would be to apply water and fertilizer through irrigation (fertigation), which could allow for a high yield even in conditions of low rainfall or low soil fertility. Applying nutrients through fertigation requires a detailed understanding of how the crop responds to different concentration of nutrients, of which nitrogen (N), phosphorus (P) and potassium (K) are the most prominent. The response of tef to N fertilizer has been described in a previous paper [
4], and in this paper we will discuss the response of tef to P and K.
Phosphorus fertilization of tef is common in Ethiopia [
5]. A blanket recommendation by the Ethiopian Ministry of Agriculture was to apply between 30 and 40 kg P ha
−1, depending on the soil type [
5], which was later changed to 100 kg DAP ha
−1 regardless of the soil type. In recent years, soil tests have been used to improve accuracy of P fertilization [
5]. A number of field trials have been published in recent years showing the effect of different levels of P on tef growth parameters. Dubale [
6] conducted a field experiment in the North Shewa Highlands of Ethiopia with three different rates of P fertilizer (0, 50, and 100 kg P
2O
5 ha
−1) and found that broadcasting different amounts of P fertilizer at sowing did not significantly affect P concentration in plant tissue or grain yield, perhaps because there was already sufficient P in the soil in which the experiment was conducted or the P that was applied became unavailable to plants. Dereje et al. [
7] conducted a full factorial field experiment in the Assosa Zone in Ethiopia in which five levels of N and four levels of P were tested. They found significant main effects and interactive effects of N and P on many different growth parameters including yield. After economic analysis, they concluded that in soils similar to their test plots, the best practice would be to apply 10 Kg P ha
−1, along with 46 Kg N ha
−1.
Although K is universally recognized as important for crop growth and yield, for many years tef grown in Ethiopia was usually fertilized only with N and P and not with K [
8]. This was due to the common belief, stemming from studies conducted in the 1960s and 1970s, that claimed that K was not limiting in Ethiopian agricultural soils [
8,
9]. Recently, this idea was called into question with largescale mapping of Ethiopian soils in a project initiated by the Ethiopian government, called the Ethiopian Soil Information System (Ethiosis) [
10]. A number of field trials have been recently published indicating that K application can increase tef yield [
9,
11]. Similarly, a greenhouse trial in which different levels of K were applied to tef growing in pots with different Ethiopian soils was conducted in order to determine an optimum K fertilization rate [
12].
The purpose of this study is to elucidate the response of tef to different levels of P and K applied in the irrigation water. It was carried out in two phases. In the first phase, tef was grown under controlled conditions in a greenhouse, in perlite, an inert medium that allowed full control of the nutrient concentration in the root zone. This is in contrast to soil, which can absorb, fix, or release ions of K
+ and PO
4−, making it difficult to determine the exact nutrient concentrations to which the roots are exposed. The second phase involved growing tef out in the field in a semi-arid Mediterranean climate in natural soil. While the first phase was designed to clarify the basic response of tef to different P and K concentrations, the second phase was intended to show how the lessons learned from the first phase play out in a realistic agricultural environment. The same experiments were used to study N fertigation in tef, recently published by Gashu et al. [
4].
4. Discussion
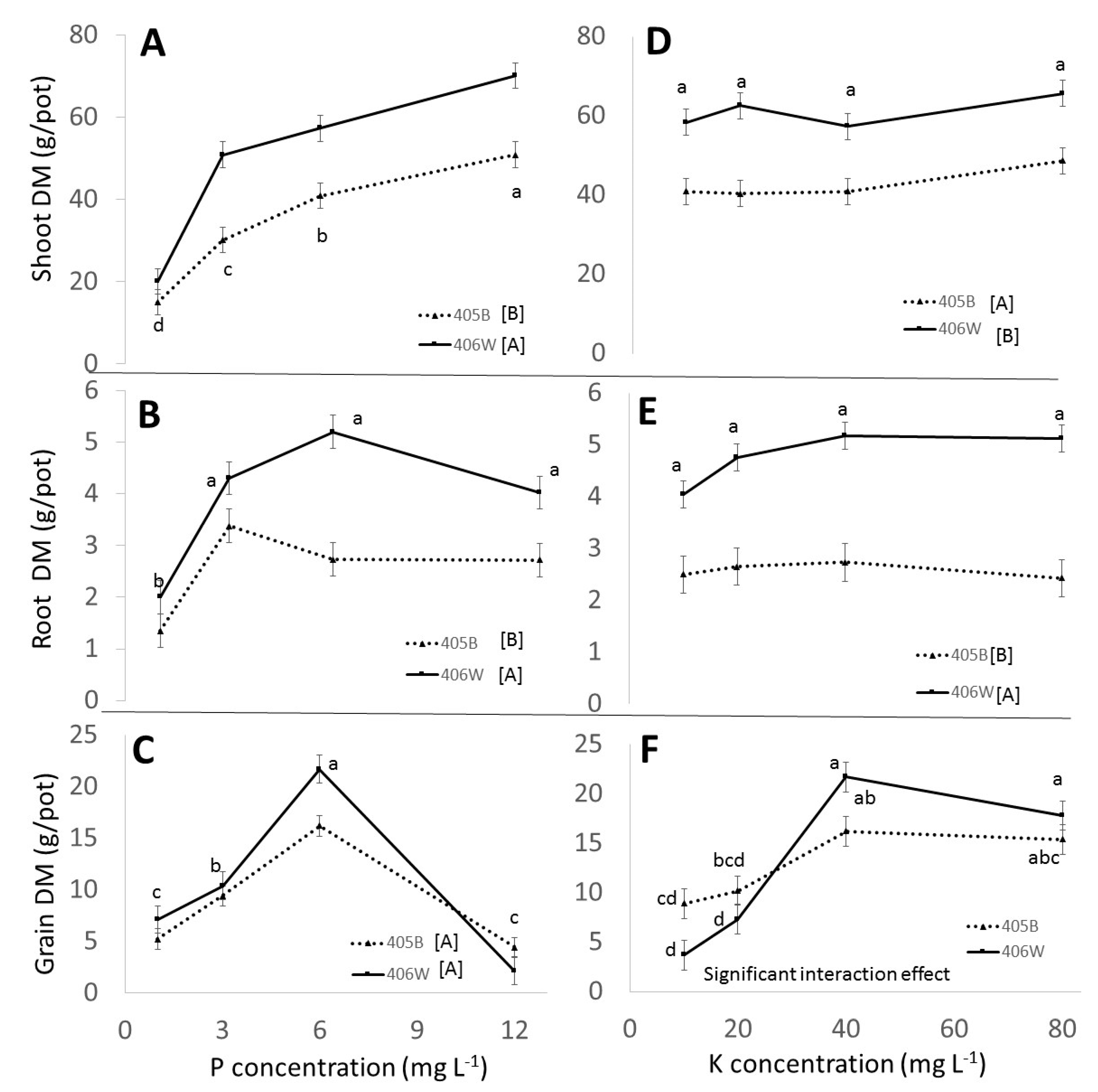
Similar to the response of tef to N fertilization detailed by Gashu et al. [
4], the response to P in the pot experiment can be divided into three ranges: underfertilization, sufficient fertilization, and overfertilization. As fertilization level rose from 1 mg L
−1 P to 6 mg L
−1 P, the grain yield increased by over threefold, indicating that section of the graph was underfertilized (
Figure 1C). At 12 mg L
−1, the yield decreased dramatically, indicating overfertilization. The optimal P concentration is, therefore, between 6 and 12 mg L
−1, and future experiments should include treatments within this range for a clearer picture. Interestingly, while overfertilization severely impacted the grain yield, it did not have any negative effect on the shoot or root DM (
Figure 1A,B).
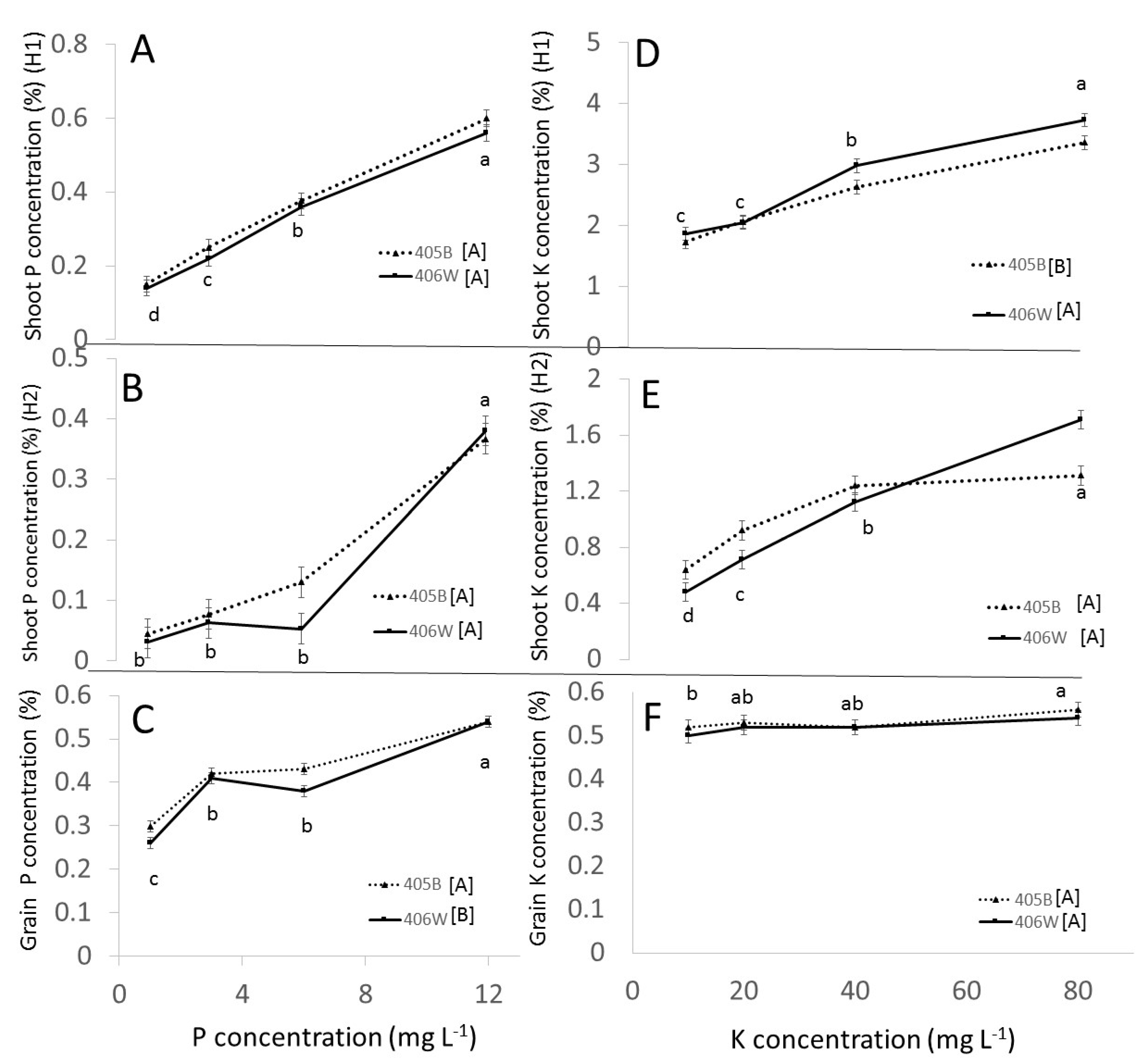
At flowering, P concentration in the shoot increased linearly with added P regardless of the concentration (
Figure 2A). However, at maturity, between 1 mg L
−1 P and 6 mg L
−1 P, there was no significant increase in shoot P, while between 6 and 12 mg L
−1 P the shoot P concentration increased by 400% (
Figure 2B). Hawkesford et al. [
17] stated that plants rarely show signs of P toxicity since they are able to downregulate P transporters involved in net root P uptake. This seems to be the case between 1 and 6 mg L
−1 P, since at maturity (
Figure 2B) the concentration in the shoot was not significantly increased in that range. The sharp increase in shoot P concentration between 6 and 12 mg L
−1 seems to indicate that the tef plant was unable to regulate the P concentration effectively. That being said, there was little evidence of actual toxicity, since other than a reduction in grain yield, plant growth was unaffected by P overfertilization (
Figure 1A,B). Given the large number of tillers in the tef that received 12 mg L
−1 P compared to the other treatments (
Table 1), it seems that excess P simply caused an increase in tiller production at the expense of grain production. The detrimental effects of P overfertilization were reported by Girma et al. [
18] in a field trial in Oklahoma, who found that even low levels of P fertilization decreased yield.
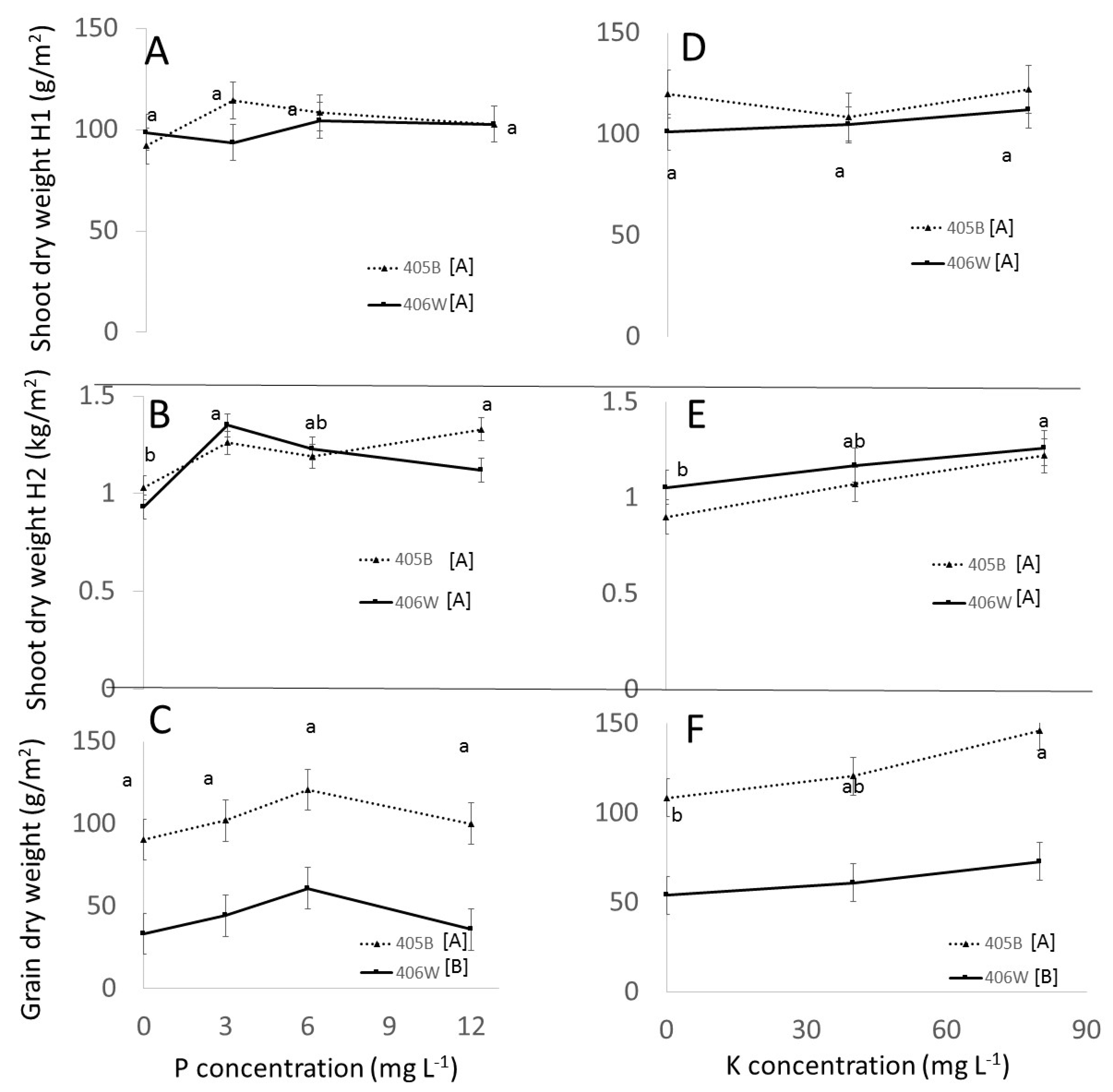
In the field experiment, there was a trend towards higher yields at 6 mg L
−1 P, which is similar to the pot experiment, but the difference in grain yield between the minimum and maximum was 33% but statistically insignificant (
Figure 3B). It is noteworthy that the grain yield and HI in the field were exceedingly low. Possible reasons for this have been discussed in detail by Gashu et al. [
4], including the heat and long daylight hours in mid-summer months.
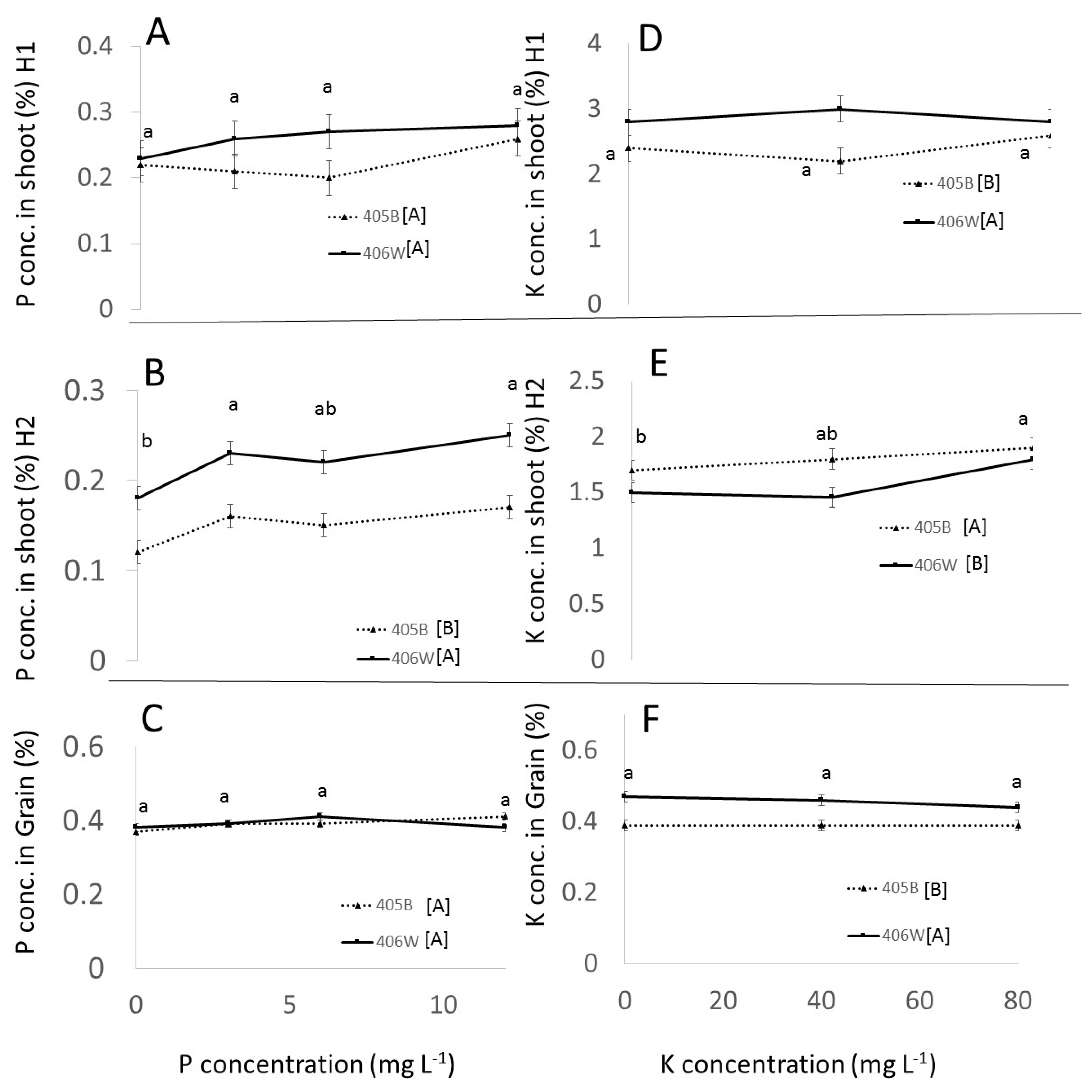
It is instructive to compare the P concentration in the shoot in the field to that in the pot experiment at flowering in order to determine, at least on a basic level, what degree of P availability occurred in the field under different treatments. In the field, shoot P concentration at flowering under 0 P application, averaged across cultivars, was 0.22% (
Figure 4A), while under the 12 mg L
−1 P treatment it was 0.27%. Both these concentrations are close to the P concentration found in the pot experiment in the shoot at flowering time (0.24%) for the 3 mg L
−1 P treatment (
Figure 2A). This suggests that regardless of the treatment, the P availability in the soil was approximately 3 mg L
−1 P in the root zone. Evidently, there was already some available P in the soil, somewhat below the 6 mg L
−1, which gave an optimum grain yield in the pot experiment. However, adding P did not increase the availability of P in the field at flowering. Perhaps the added P was immobilized through biotic or abiotic processes.
The P concentration at maturity tells a slightly different story. At maturity, plots receiving 0 mg L
−1 in the irrigation water had a significantly smaller P concentration in the shoots than those that received even low amounts of P concentrations (
Figure 4B). Furthermore, the total P uptake was significantly higher in those plots that received P in the irrigation water, with no significant difference between those treatments (data not shown). Evidently, as the plant grew larger and P became more limiting, there was a significant advantage to fertigating with P.
This may explain why the grain yield at 6 mg L−1 in the field was 33% higher than the grain yield when 0 P was applied, though not a statistically significant difference. This follows the same trend as the pot experiment, in which grain yield reached a maximum at 6 mg L−1 P.
In the pot experiment, the response to K fertilization can be divided into two ranges: underfertilization and sufficient fertilization. The grain DM responded positively to K fertilization up to 40 mg L
−1 K, and then plateaued (
Figure 1F). Unlike the P (
Figure 1A–C) and N (Gashu et al., 2020) segments of this experiment, there was no significant negative effect of K at the highest level of K fertilization, although there was a small, non-significant decline in grain DM between the 40 mg L
−1 and 80 mg L
−1 treatment (
Figure 1F). This is similar to the response of tef grain yield to K fertilization observed by Misskire et al. [
19] in a field experiment in Ethiopia. They reported yield rising and then plateauing with increased K, with a slight, non-significant drop at the highest K level. It is interesting that no decline in grain yield occurred in the higher K treatments, since Mulugeta et al. [
9] found a decline in grain yield in plants with much lower shoot K concentrations (0.6–0.9%) than the concentrations we measured in the highest K fertilization treatments (
Figure 2C). Furthermore, maximum yield was observed at a much higher vegetative K concentration in our experiment (1%) compared to the 0.63% K reported by Mulugeta et al. [
9] as the internal K requirement of tef. It is possible that the decreased grain yield in high K treatments reported by Mulugeta et al. [
9] was not caused by the increase in K itself, but rather a secondary effect caused by an imbalance of other nutrients. In our experiment, all other nutrients were provided at high rates and in available forms, so imbalances of other nutrients did not occur. This underscores the importance of conducting experiments in perlite, where minerals can be provided in a highly controlled manner.
Interestingly, in the pot experiment, there were no significant changes in shoot or root biomass across the entire K treatment range (
Figure 1D,E). In the field experiment there was an increase in shoot DM as the K concentration in the fertigation water increased from 0 to 80 mg L
−1, but this difference was not apparent at flowering and became apparent only at maturity (
Figure 2E). The lack of response of the vegetative portions of the plant to K fertilization in the pot experiment and the late-stage response in the field experiment shows that the completion of the lifecycle of the plant and measurement of grain yield are essential for showing the effect of K fertilization on tef. The increase in straw yield due to K fertilization which we observed in the field and not in the pot experiment has been reported previously. Gebrehawariat et al. [
11] reported an increase in straw with increased K fertilization in tef. Interestingly, while Gebrehawatiat et al. [
11] reported that increased K caused an increase in the number of fertile tillers, we observed a decrease in tillers with increased K in the pot experiment (
Table 2), and no clear effect of K on tiller number in the field experiment (
Table 4).
In the pot experiment, K concentration in the shoot increased with increasing K fertilization (
Figure 2D,E), but the K concentration in the grain was not responsive to K fertilization, remaining at around 0.5% regardless of cultivar or K treatment. In the field experiment, there were significant differences between the cultivars (0.46% in 406W and 0.39% in 405B) but no significant effect of K treatment on grain K concentration. Evidently, the K concentration in the shoot is flexible, but the K concentration in the grain needed to be at a certain range in order to allow grain development. This is in contrast to the findings of Gebrehawariyat et al. [
11], who found that on average grain K concentration increased by 33% from the lowest K treatment (0 kg K
2O ha
−1) to the highest K treatment (120 kg K
2O ha
−1).
In the field, shoot K concentration at flowering under 0 K application, averaged across cultivars, was 2.6%, while under 80 mg L
−1 K treatment it was 2.7%. Both these concentrations are close to the K concentration found in the pot experiment in shoot at flowering (2.8%) under the 40 mg L
−1 K treatment (
Figure 2D). This suggests that at the beginning of the field experiment, regardless of the treatment, the K availability in the soil was approximately 40 mg L
−1 K in the root zone. At maturity, however, there was a 16% increase in shoot K concentration between the 0 and 80 mg L
−1 K treatments, as well as a 56% increase in total K uptake between those two treatments (data not shown), indicating that fertigation with K increased K availability later on in the lifecycle.





{kind=link}
{kind=link}
{kind=link}
{kind=link}



