1. Introduction
Barley (
Hordeum vulgare L.) is an economically important cereal crop that is known to be dry, cold, salt-tolerant, and well adapted to low-input environmental conditions and changing climates [
1,
2]. It is cultivated at high altitudes, commonly under rain-fed conditions. Barley is often grown in marginal agricultural areas with low annual precipitation, often less than 220 mm [
3]. It is ranked fourth in terms of the most cultivated crop (by area) in the world, after wheat, maize, and rice. Almost half of the world’s barley growing area is in Europe, including Poland, where it is ranked second in terms of the most cultivated crop after wheat. It is used for livestock feed, malt, and foods. Although domestic Polish barley supply has fluctuated substantially in recent years, it tended to decline from 1969–2018 [
4].
Fungal pathogens are an economically significant factor limiting the size and quality of barley grain. Barley is often infected by powdery mildew fungus (
Blumeria graminis DC.
hordei), brown rust caused by
Puccinia hordei (Ph), stripe rust caused by
P. striiformis f. sp.
hordei (Psh), and stem rust caused by
Puccinia graminis f. sp.
tritici (Pgt). The loss of yield caused by powdery mildew can reach up to 30%, at an average of 5–10% [
5,
6]. Yield losses caused by barley brown rust can be up to 60% in susceptible varieties [
6,
7,
8]. However, the average yield losses of barley due to barley brown or stem rust often reach 10–25% [
9,
10,
11]. The obvious alternative to fungicide treatment against plant diseases is the use of genetically resistant cultivars [
12,
13,
14,
15].
The potential yield loss caused by disease depends not only on host susceptibility and weather conditions, but also on the timing and severity of disease outbreaks relative to crop growth stage. The greatest yield losses occur when one or more of these diseases occur before the heading stage of development, and can be potentially important for the detection of early-activated pathogen-associated molecular patterns, which can trigger nonspecific defense cascades. On the other hand, resistance during late stages of growth, such as the milky-waxy stage, can be important for the identification of key resistance proteins. The early detection and proper identification of pathogens are critical to in-season disease management, future variety selection, and the use of these genes in breeding programs [
14,
16,
17].
Two types of barley rusts and powdery mildew resistance have been described: (1) hypersensitive resistance (HR), and (2) adult plant resistance (APR). Resistance genes are pathogen race-specific in their action; they are responsible for HR and mostly encode immune receptors for nucleotide binding leucine rich repeat (NB-LRR). They are effective throughout all growth stages. By contrast, APR genes are only functional in adult plants and usually confer only partial resistance, albeit in a non-race-specific manner [
17,
18].
At least 38 different race-specific resistance genes/alleles to powdery mildew are known and used in varieties grown throughout Europe [
19,
20]. Barley cultivars with effective genes for resistance to powdery mildew have been an efficient means for controlling this disease [
11,
15,
20,
21,
22,
23,
24,
25]. Barley breeders use the following major seedling resistance genes:
Mla6,
Mla7,
Mla9,
Mla12, and
Mla13; and
Mlk,
Mlg,
MlLa,
Mlh, and
Mlra, which originate in landraces and in the subspecies,
H. vulgare ssp.
spontaneum.
More than 25
Rph (resistance to
P. hordei) genes have been identified and mapped in barley [
26], including 21 as seedling resistance [
27].
Rph5 and
Rph6 on chromosome 3H,
Rph9 and
Rph12 on chromosome 5H, and
Rph15 and Rph16 on chromosome 2H have been described as alleles of the same gene. Only
Rph7,
Rph15, and
Rph16 are still effective in Europe [
28] and the number of effective
Rph genes available to breeders is decreasing rapidly [
26]. Among all the known
Rph genes, only
Rph1 has been isolated recently, using a newly developed cloning approach called Mutant Chromosome Sequencing (MutChromSeq) in combination with genetic mapping [
29]. Six genes are known to confer barley resistance upon
Pgt in the US, including well-characterized
Rpg1,
Rpg4, and
Rpg5, as well as the less studied
Rpg2,
Rpg3, and
Rpg6 [
30].
APR is considered potentially more durable for controlling barley rusts or powdery mildew than seedling resistance genes. The use of race-specific resistance genes in barley quickly results in the selection of virulent races of
B. graminis f. sp.
hordei (PM) or
Puccinia spp. When a cultivar containing one dominant resistance gene is grown on a large acreage, new virulent races can emerge within 4–5 years. Virulence has been detected for most seedling resistance genes but is unknown for APR genes [
17].
Two forms of durable resistance to powdery mildew are described. Mlo resistance (gene
mlo) was identified as durable resistance to powdery mildew in barley landraces [
31]. Since 1984, it has been well described and deployed in many barley cultivars throughout Europe [
20,
25,
32,
33]. The second form of durable resistance involves genes other than major
R-genes. These may be expressed at one or more growth stages and include partial or quantitative resistance and adult plant resistance (APR). Three novel major-effect powdery mildew APR genes from landraces (
Rbgh1,
Rbgh2, and
Rbgh3) were identified in the terminal regions of the barley chromosomes, 5HL, 7HS, and 1HS, respectively [
34]. Among the genes for barley brown rust resistance, three genes,
Rph20 and
Rph24 on chromosome 5HS and 7HS, respectively, and
Rph23 on chromosome 6HS, confer high, moderate, and low levels of APR, respectively [
8,
22,
29,
35,
36,
37,
38,
39].
World-wide, there are about 1800 gene banks, including more than 600 in Europe [
40], with about 7.4 million accessions stored globally [
41]. However, only 25–30% of these accessions are genetically unique [
42]. There is a need for characterization of this germplasm in terms of its agronomic potential, including resistance to biotic and abiotic stresses and for establishing associations between markers and phenotypes. This knowledge is necessary to enable the use of specific accessions in breeding programs [
42,
43,
44,
45,
46]. However, the genetic studies were conducted on a still limited number of accessions [
47]. To determine the markers that determine a qualitative trait controlled by many genes, including APR, the assessed collection should be highly variable for these traits. Old European barley cultivars or landraces are an important source of genetic variation and resistance to biotic stresses, including powdery mildew and rusts [
2,
23,
48,
49,
50,
51,
52,
53,
54].
The aim of this study was to associate genetic loci with adult plant resistance (APR) to powdery mildew (PM), barley brown rust (BBR) and barley stem rust (SR) at heading and the milky-waxy plant seed development stages. To achieve this, we used GWAS analysis of DArTseq-derived markers and phenotypic data relating to 431 barley accessions, which are stored in the Polish Gene Bank, segregating for these disease resistance traits.
2. Materials and Methods
2.1. Plant Material
A collection of 431 barley accessions, including landraces and old cultivars, stored at the Polish Gene Bank (National Centre for Plant Genetic Resources: NCPGR) were phenotyped and evaluated using DArTseq: 137 POL, 67 DEU, 38 SWE, 35 CSK, 34 FRA, 27 GBR, 25 DNK, 21 NLD, 12 AUT, 8 SUN, 6 NOR, 4 FIN, 3 IRL, 3 CAN, 2 USA, 2 HUN, 1 each from UKR, TUR, PRK, NZL, JPN, BEL, and 1 of unknown origin were evaluated. For evaluation using DArTseq 23, additional control genotypes were included.
Barley was one of the principal crops that accompanied the spread of agriculture into Europe during the 6th and 5th millennia BC and was dispersed along two main routes: a southern route along the Mediterranean, reaching the Iberian peninsula, and a northwards route passing through central Europe, eventually reaching northern Scotland [
55,
56]. Because of this, the accessions were grouped into sub-collections, either by country of origin or by European region. These sub-collections were: Polish; West-Central European (CSK, DEU, DNK, AUT); French; Great British; North European (FIN, NOR); and Swedish. The accessions were further classified into three groups: group A (206 accessions), representing old cultivars cultivated prior to 1985; group B (178 accessions), representing moderate and modern cultivars cultivated after 1985; and group C (37 accessions), representing Polish landraces (
Table 1). These sub-collections correspond to barley domestication and adaptation in Europe and temporal trends in genetic diversity in European cultivars [
57].
Landrace accessions are genotypes that were not improved by breeders. Old cultivars were improved and obtained based on selection during breeding programs.
The Polish accessions were selected to reflect the diversity of the Polish accessions held at the Gene Bank, with priority given to those with key phenotypic traits in Polish breeding programs. This was then supplemented with non-Polish accessions from countries where a particular trait is most frequent. The passport data, listed in Additional file 1 include: accession number (ACCENUMB), accession name (ACCENAME), country of origin (ORIGCTY), institute code (INSTCODE)/institute name, acquisition date (ACQDATE), donor institution code (DONORCODE)/donor institute name (DONORNAME), and type of germplasm storage (STORAGE).
2.2. Field Experiments and Phenotypic Evaluation
Field experiments were conducted in 2018 and in 2019 on the experimental fields of the Plant Breeding and Acclimatization Institute—National Research Institute (PBAI-NRI), Radzikow, near Warsaw, Poland. No specific permissions were required. No endangered or protected species were involved.
The experimental trials were conducted in a randomized complete block design. In 2018, seeds were sown in three replications (blocks), and in 2019 in two replications, in rows (row length of 2.0 m), with a plant spacing of 4.0 cm and a row spacing of 20.0 cm.
In 2019, a trait measurement was conducted at the heading stage (HA), when half of the heads had emerged for 50% of the plants in a plot (Z55, according to the Zadoks growth scale) and, 2 weeks later, at the early milky-waxy seed maturity stage (MW; Z75, according to the Zadoks growth scale) [
58]. In 2018, the measurement was conducted once, in the milky-waxy stage.
2.3. Weather Conditions
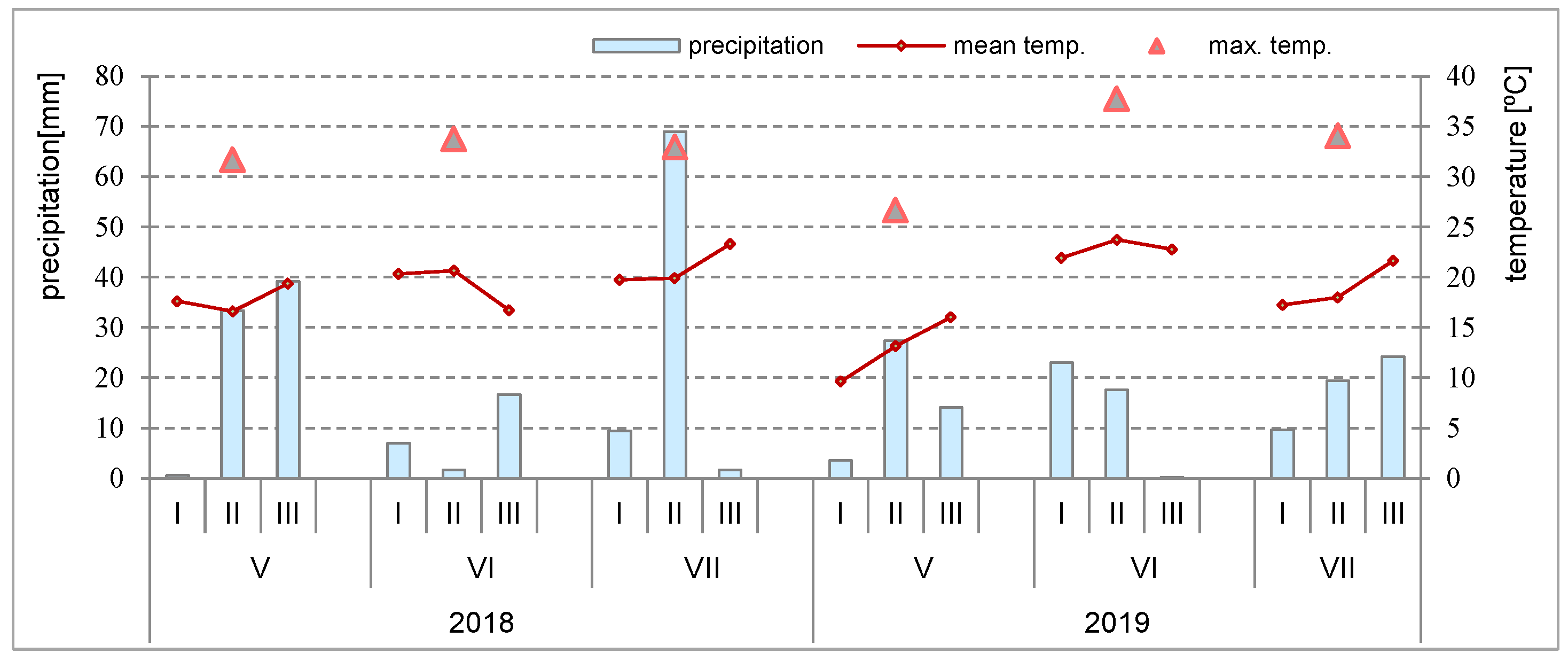
The weather conditions, including the mean and maximum temperatures (°C) and precipitation (mm), were monitored during the second quarter of 2018 and 2019 (May–July) (
Figure 1). This was a period when the plants were in the heading to physiological maturity and harvest stages.
The average air temperature and precipitation are presented for 10 days of each month. The maximum temperature is the maximum temperature that occurred in the month.
2.4. Statistical Analysis
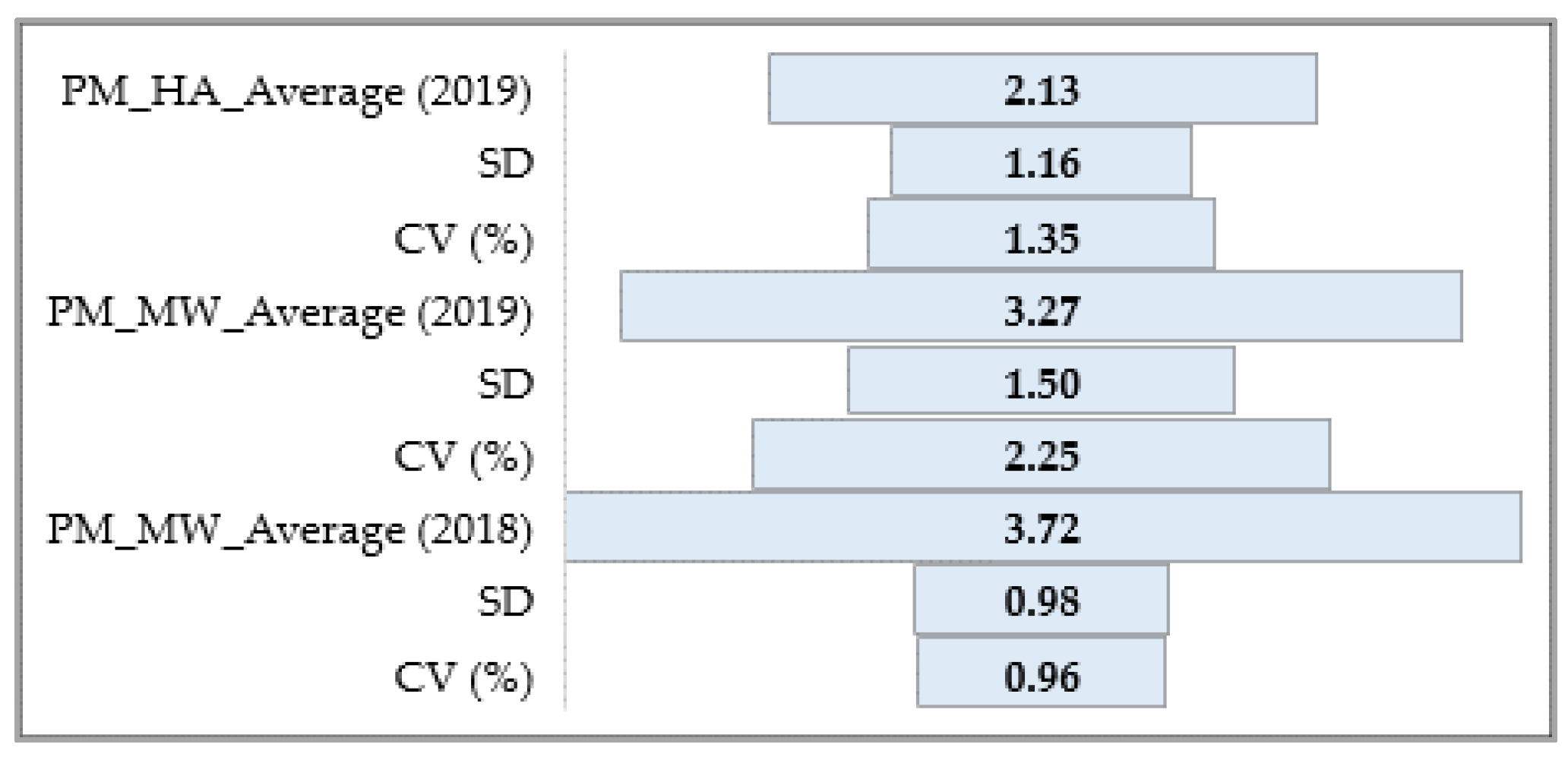
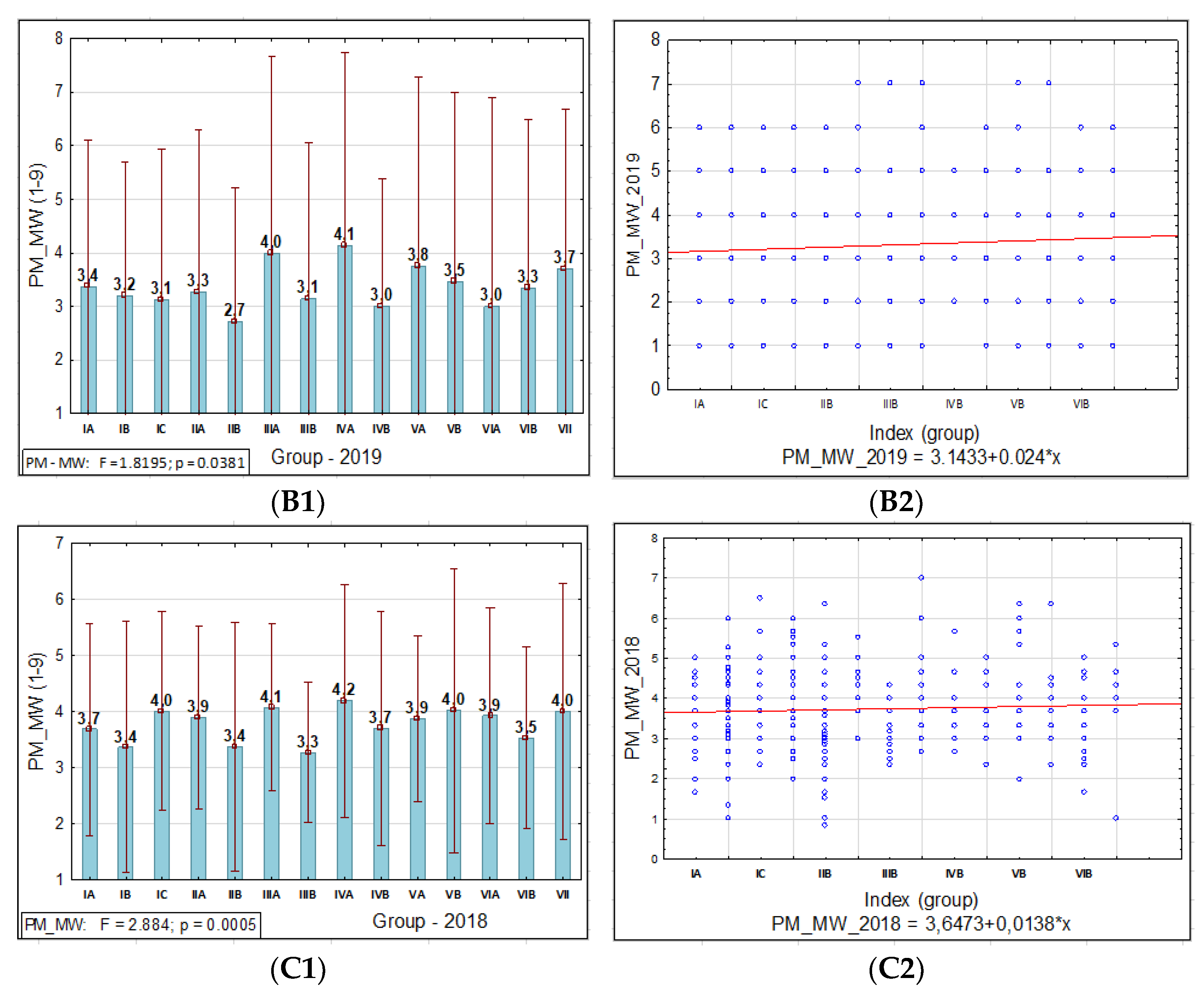
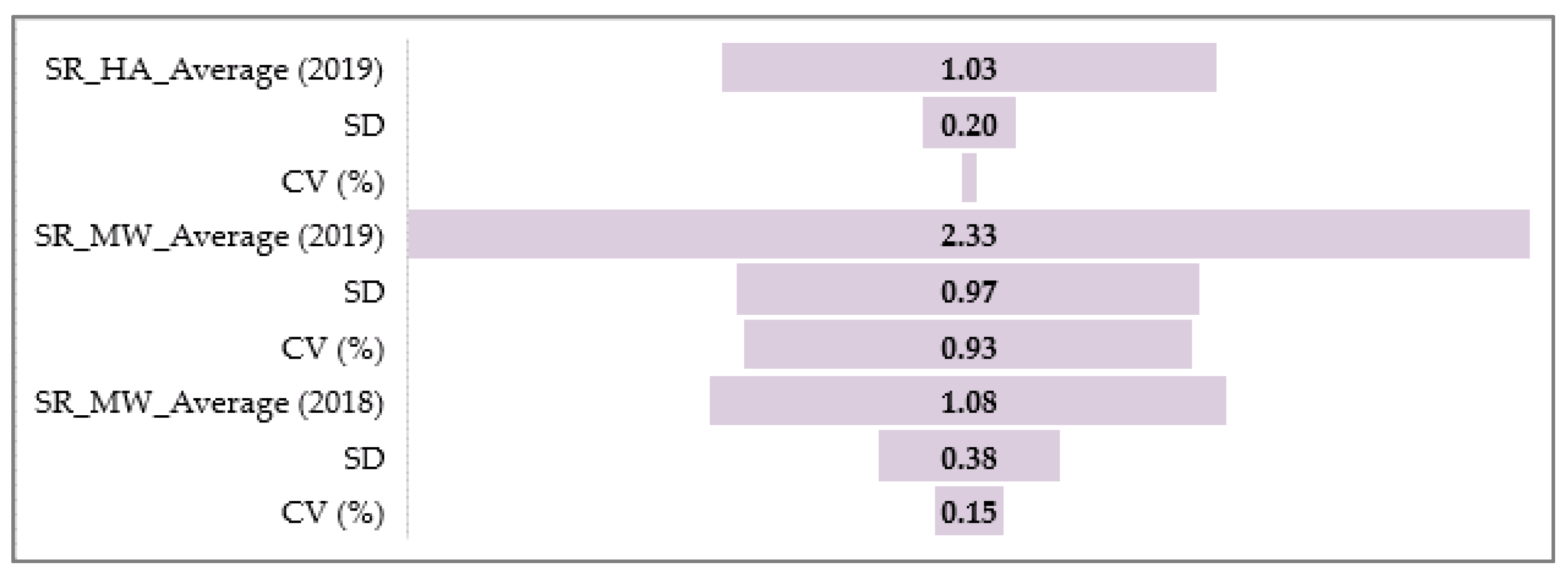
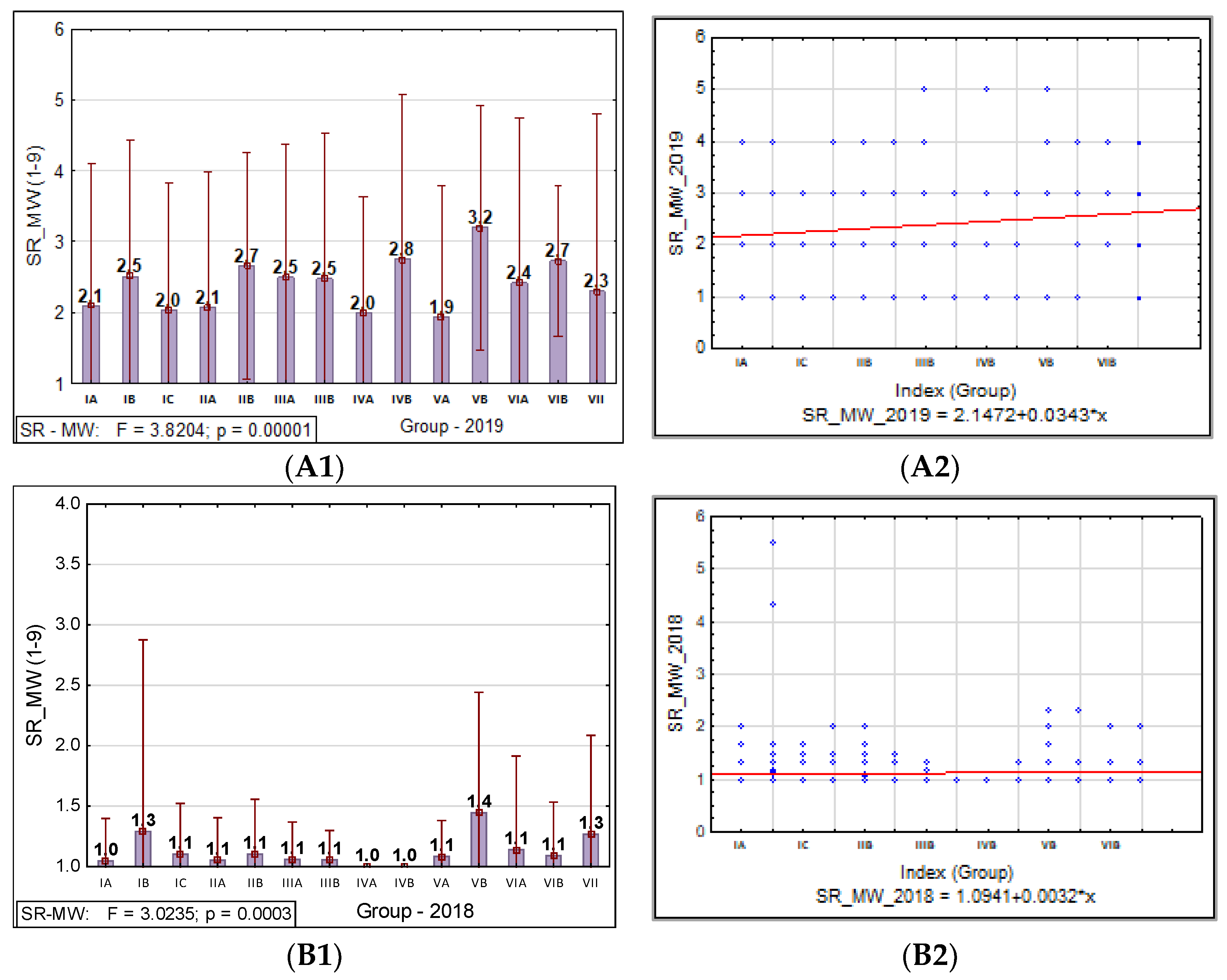
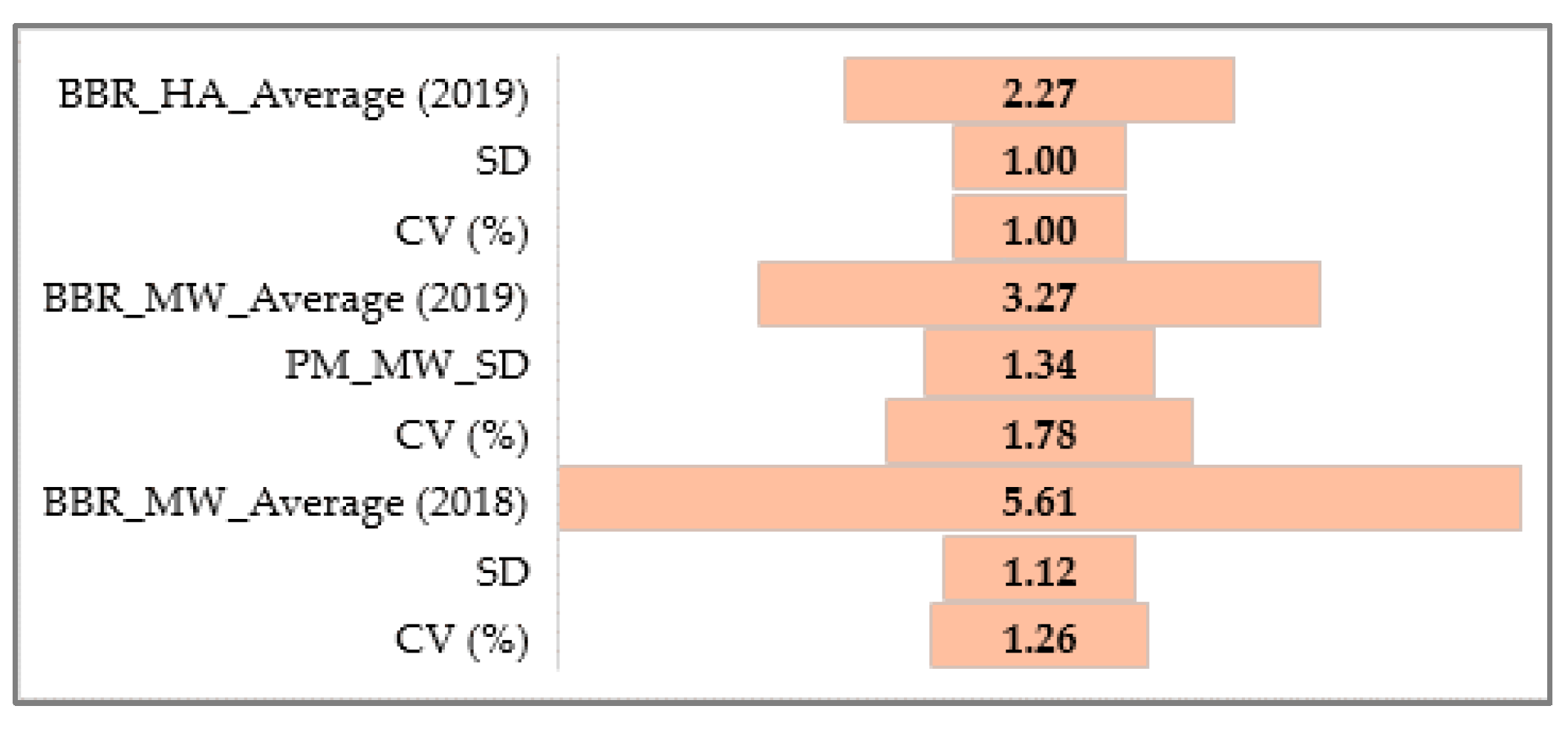
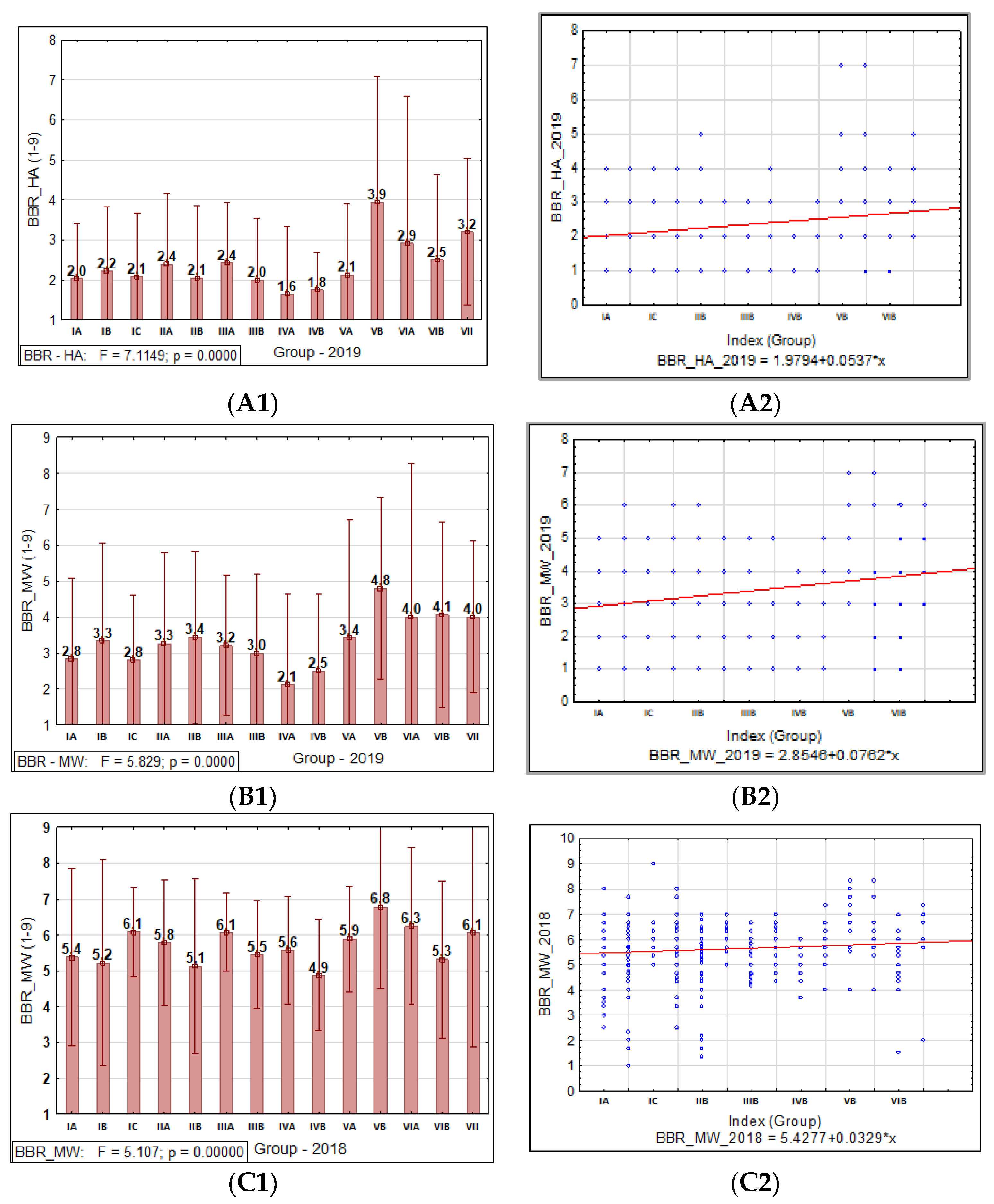
A statistical analysis of all the traits, PM, SR, and BBR resistance at the HA and MW stages, was conducted using Statistica software (version 13.31984-2017 TIBCO Software). It was used to obtain the range, mean, standard deviation (SD), coefficient of variation (CV), and analysis of variance (ANOVA test; α ≤ 0.05) values to confirm the significance of the differences in PM, SR, and BBR resistance between the accessions and the sub-collections. The sub-collections were created using two criteria: (1) cultivation period or registration of cultivars (group A—cultivated prior 1985, B—cultivated after 1985 and C—Polish landraces) and (2) country of origin or geographical region. For the sub-collections, the results are presented in the form of graphs with ANOVA and SD bars. Based on the differences between the sub-collections, it was possible draw conclusions about the progress (gain) in the breeding programs over the years.
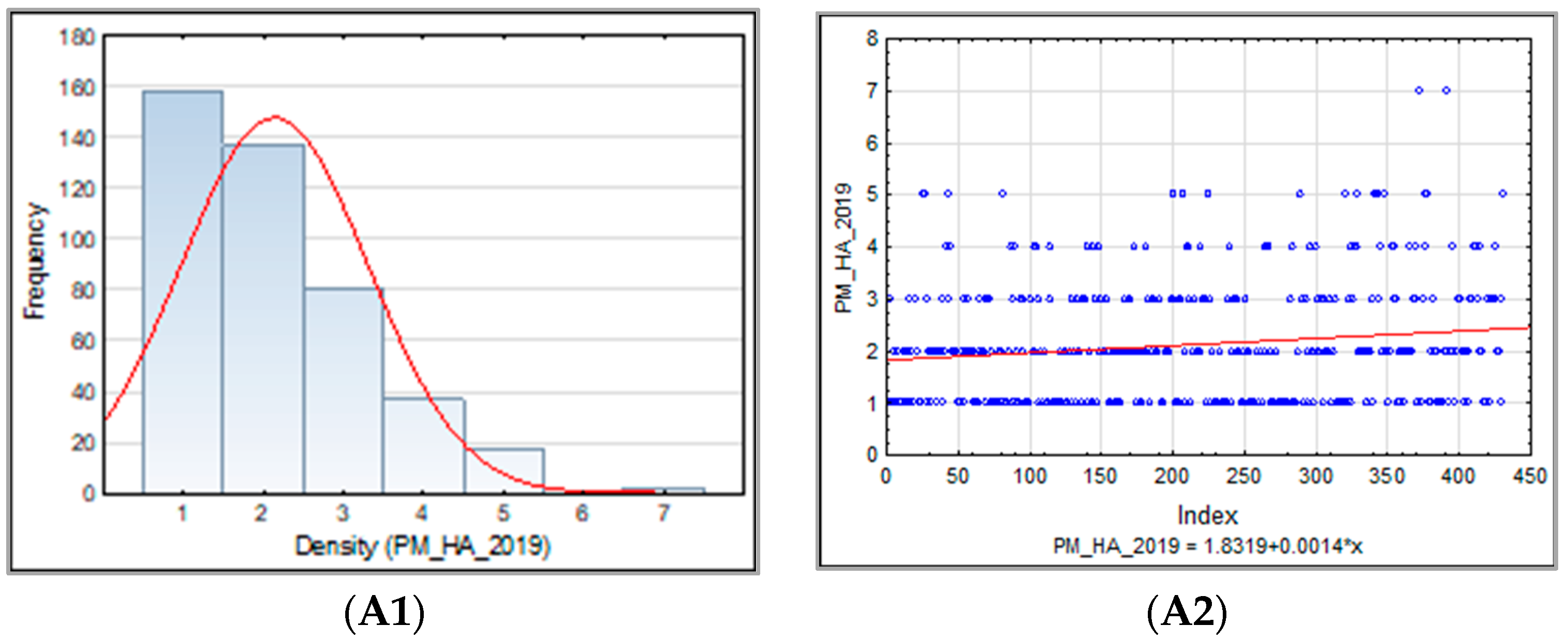
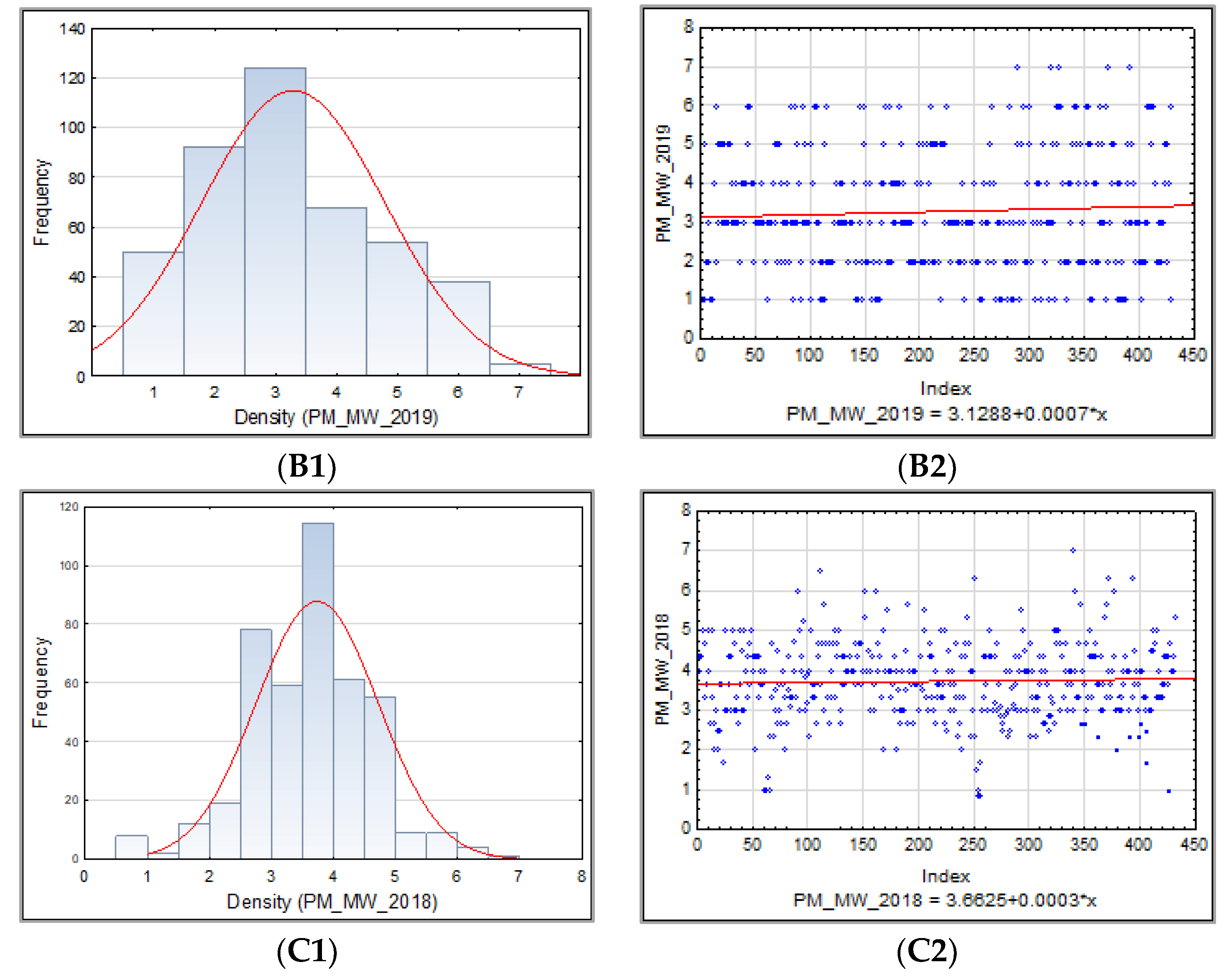
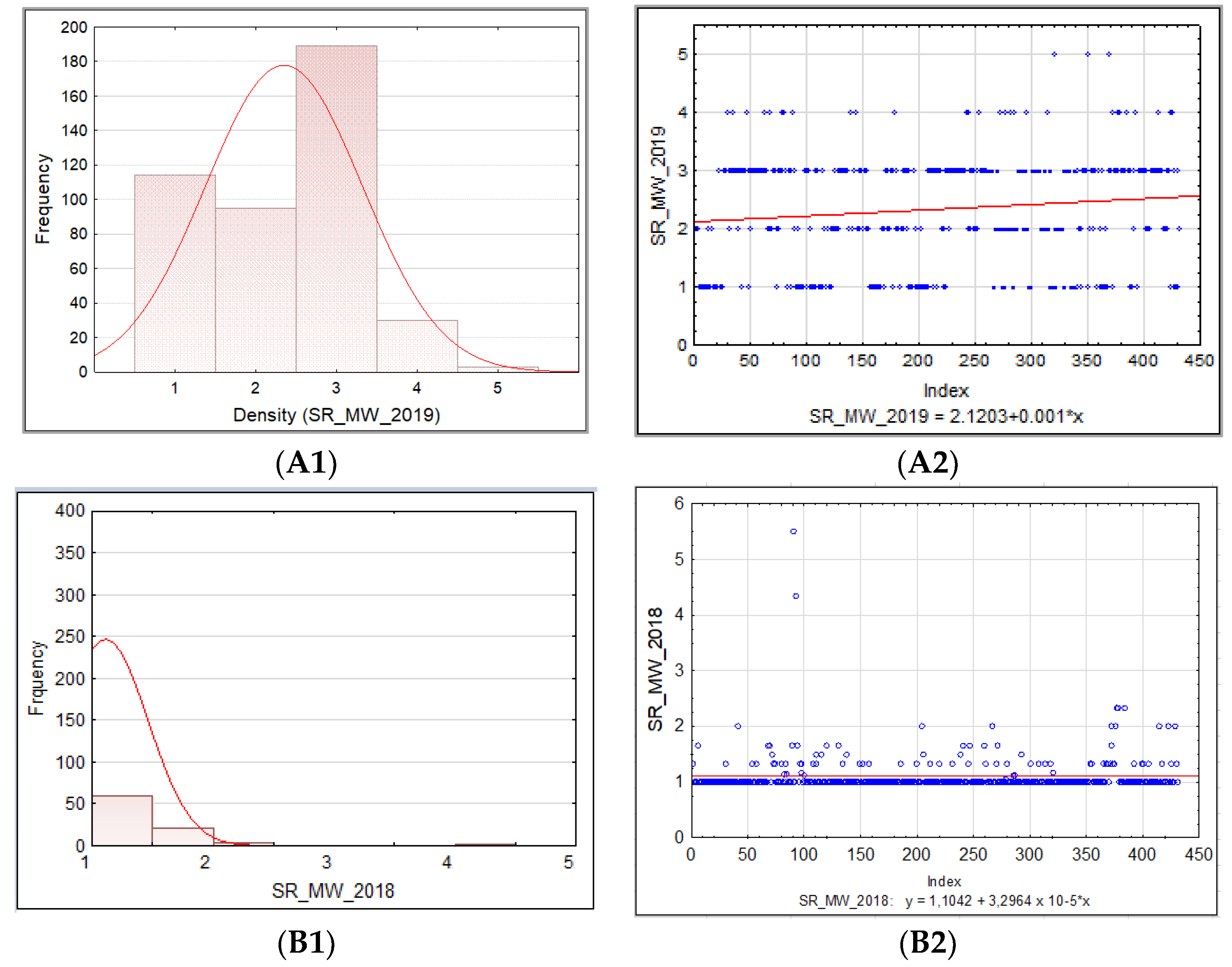
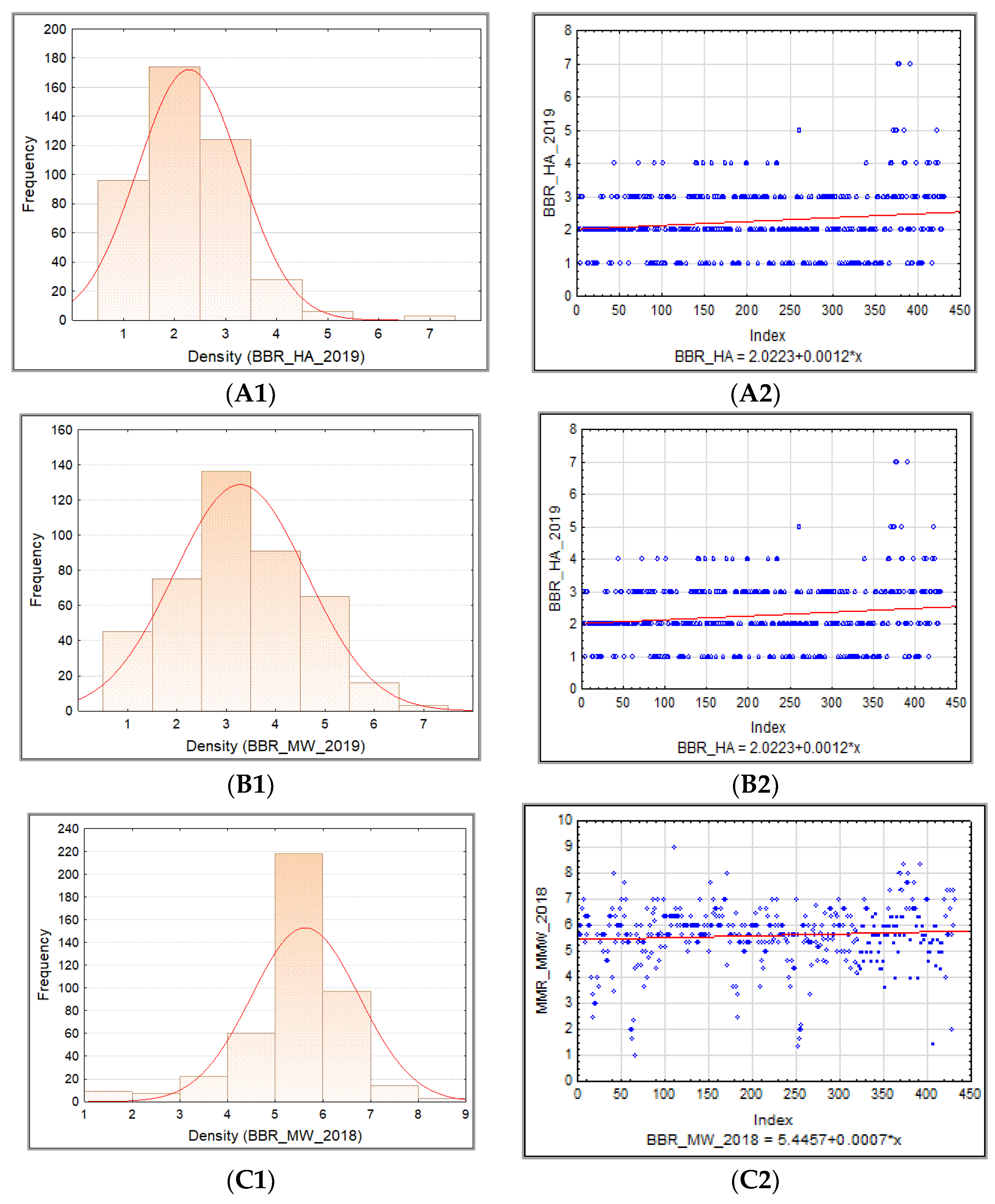
The frequency distribution of the barley accession scores for PM, BBR, and SR in 2018–2019 was presented for collection and for each sub-collection separately in the form of the normal Gauss frequency distribution and as a regression analysis model to estimate the relationship between the disease resistance scores (accessions) and the frequency index. Correlations between traits were also analyzed.
2.5. Genotyping
In total, 454 barley accessions were genotyped by using Diversity Arrays Technology (DArT) Pty Ltd., Monana, Australia, using DArTseq [
59]. The SNP decisions were taken using IBSC Barley Morex v2 assembly [
60]. The Barley GBS 1.0 platform DArT genotyping service returned 28,530 in-silico DArT-seq markers.
2.6. Data Filtering Process
The DArT data were handled in the same manner as described previously for soybean [
61]. That is, we used the dartR v1.1.11 package [
62] in the R programming language. SNPs and genotypes were removed if SNP markers contained >5% missing data and genotypes contained >10% missing data, respectively. SNPs with a reproducibility score (RepAvg) <100% were removed. Where SNPs originated from the same fragment, a random SNP was retained while the others were discarded. Non-informative monomorphic SNPs were removed, as were rare SNPs with a minor allele frequency of <1%. After filtering, 453 (as well as 1 individual, which was removed due to having >10% missing calls) and 10,153 SNPs were retained for further analysis.
2.7. Genome-Wide Association Study (GWAS)
A GWAS analysis was conducted using the GAPIT v2018.08.18 R package [
63,
64]. We used the recently developed Bayesian information and Linkage disequilibrium Iteratively Nested Keyway (BLINK) model, which has been shown to produce fewer false positives, identify more true positives and scale very large data sets [
63,
64,
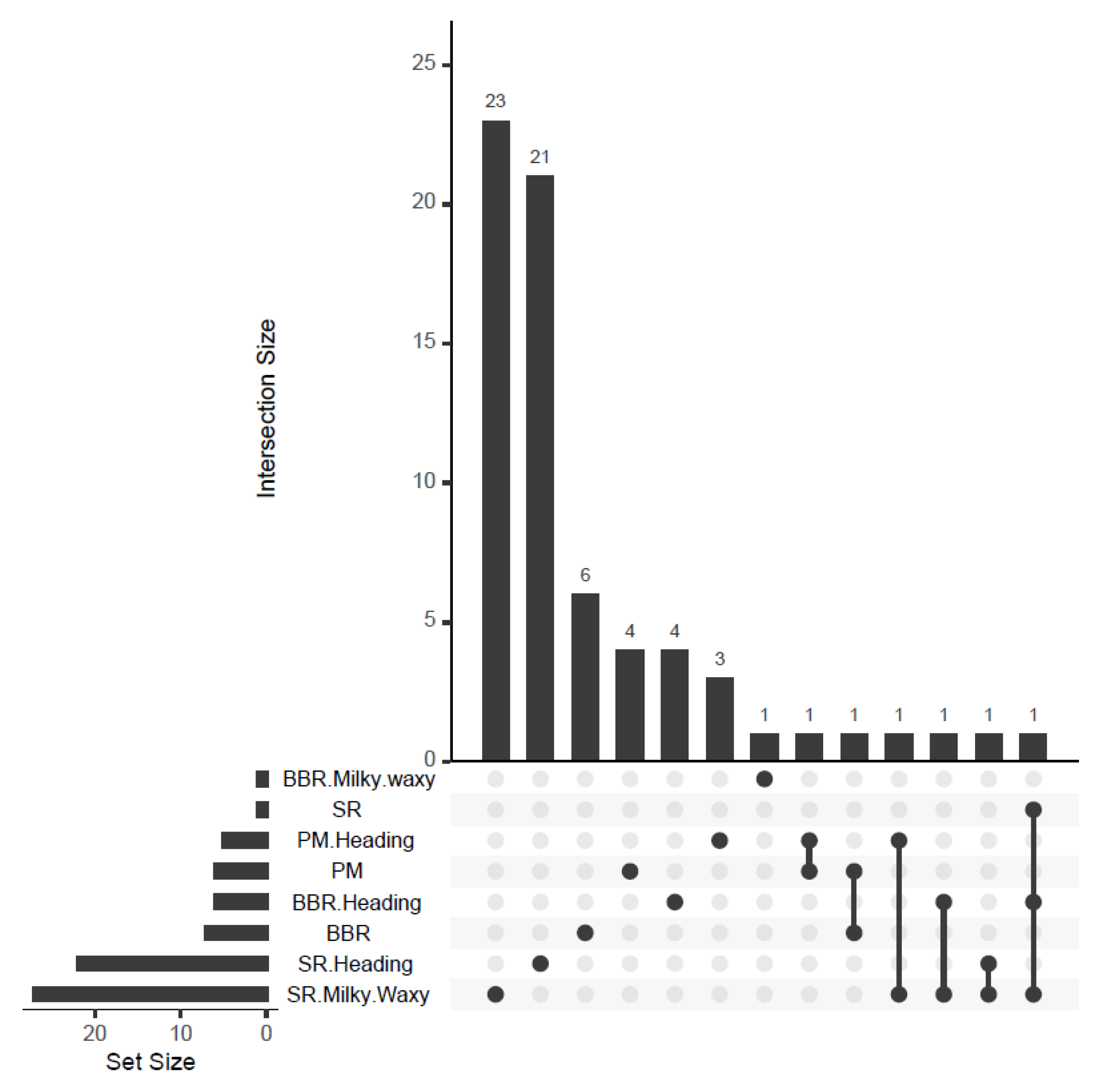
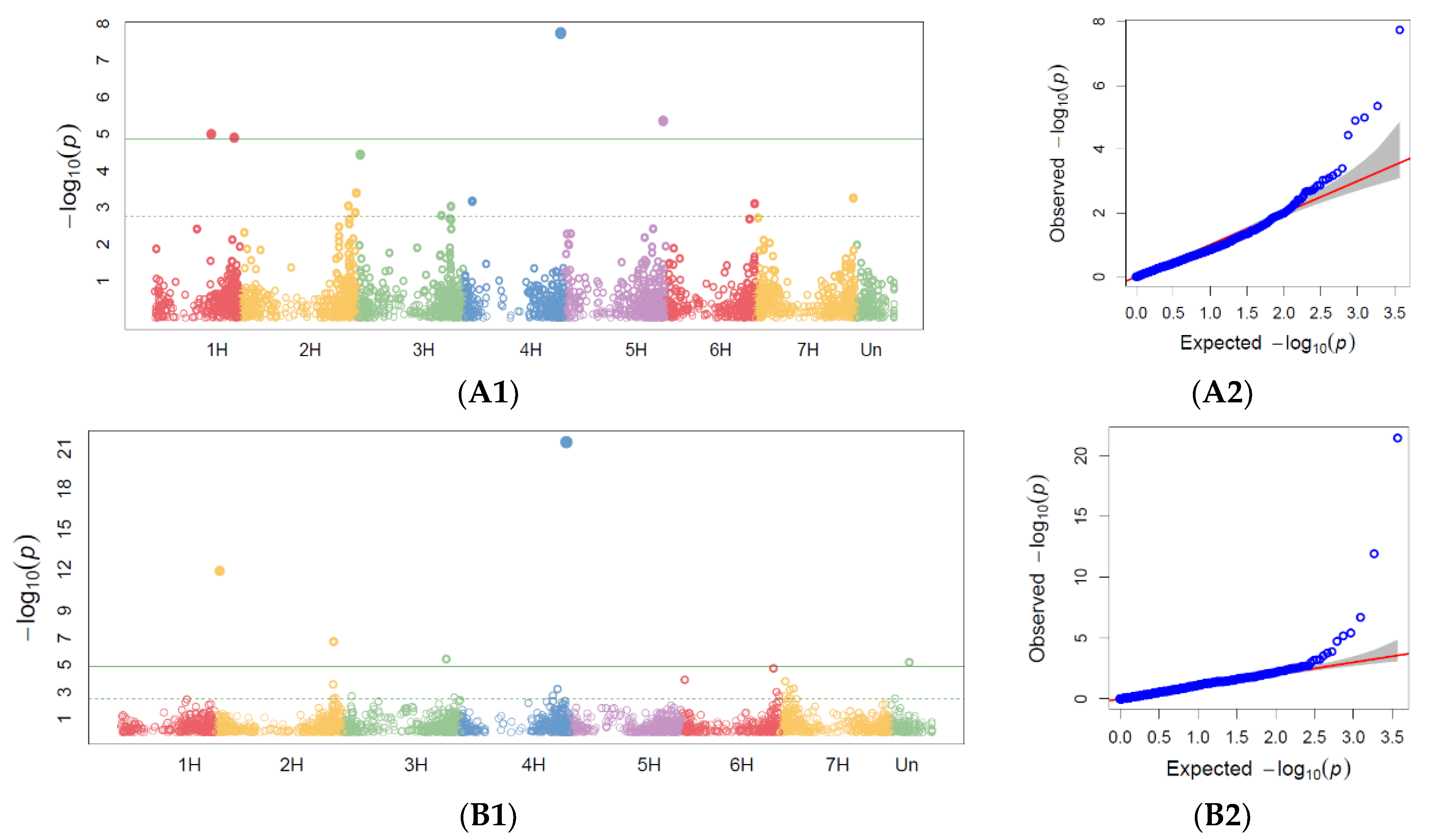
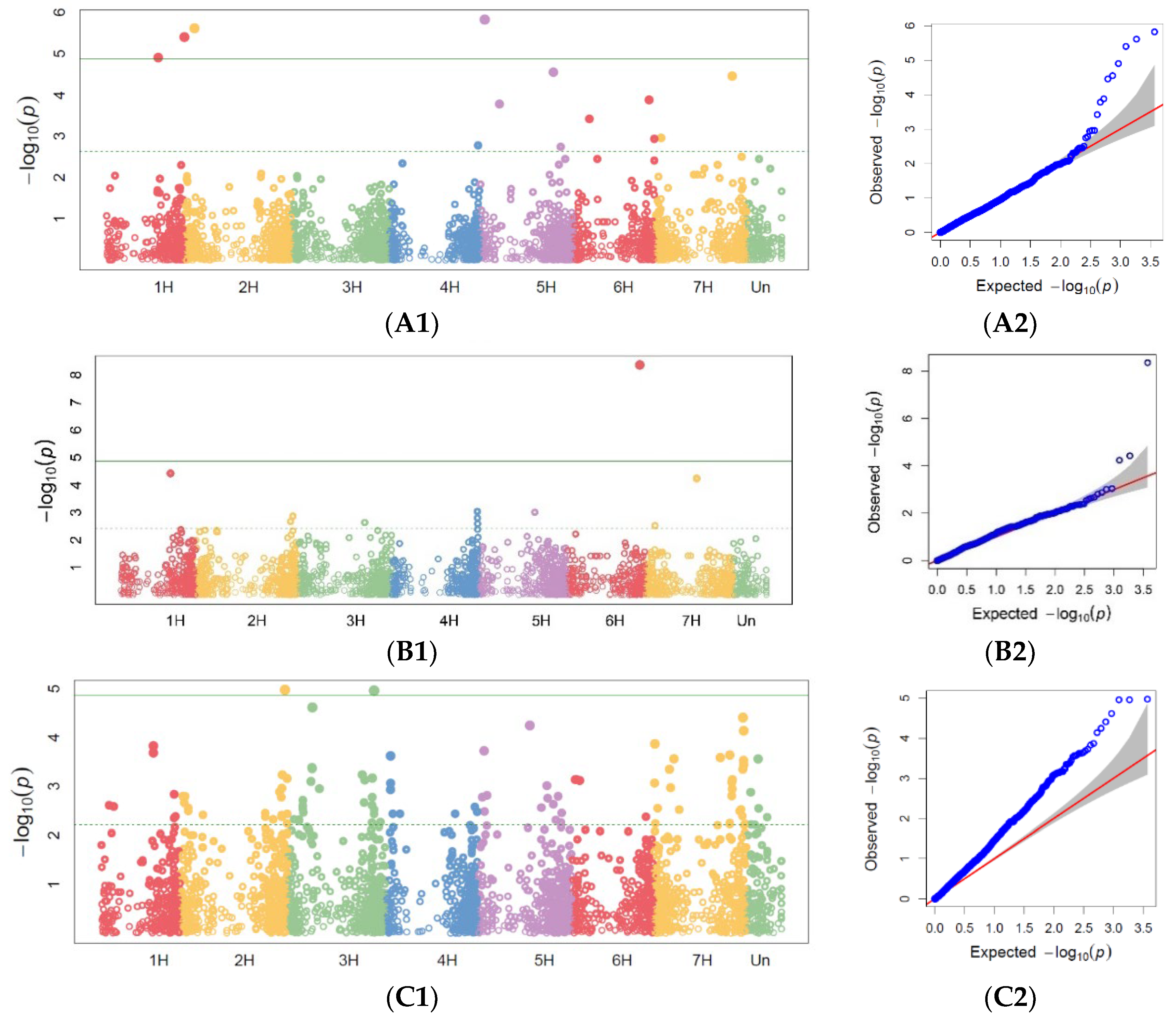
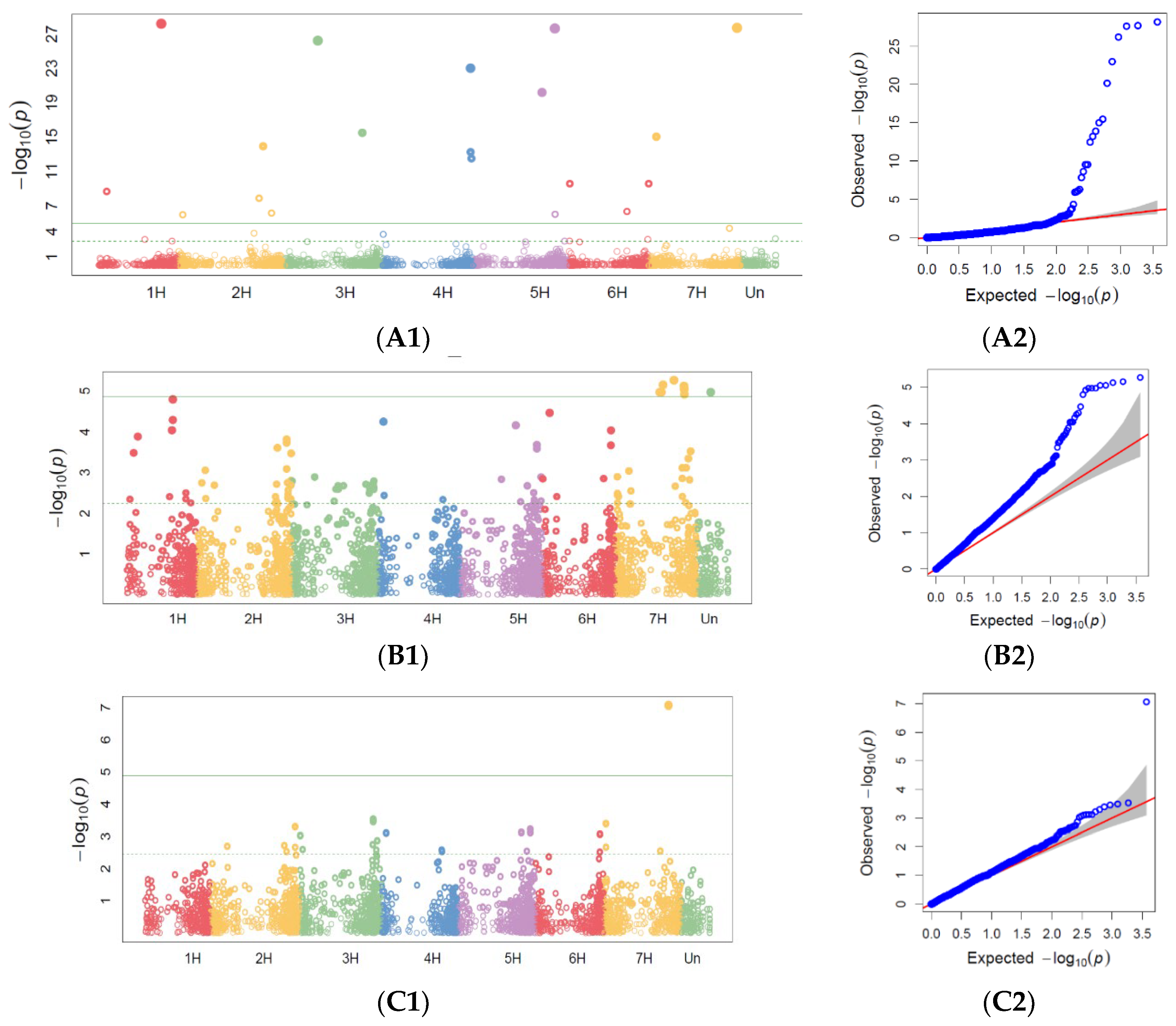
65]. The physical genome positions of the markers were derived from the DArTseq SNP genotype file. Only markers with a physical position on one of the chromosomes and zero missing data were used as inputs to the GWAS analysis. GWAS for PM, SR, and BBR was conducted for disease resistance scoring at the heading and milky-waxy seed stages in 2019 and for maximum scores across all replicates in 2018–2019 based on the fact that, because of drought (temperatures were relatively very high during spring, while precipitation was at a low level), on average, the disease severity observed in the accessions was scored at a low level. In order to show the distribution of SNPs over the chromosome, Manhattan plots were also generated.
4. Discussion
This study is a part of a larger effort focused on developing and implementing a national management system for major crop plant genetic resources stored in the Polish Gene Bank, which incorporates phenotypic and genotypic data (
https://agrobank.cdr.gov.pl/index.php (accessed on 29 October 2021)). This management system, with data sets for major crops, will facilitate more effective breeding of new cultivars well adapted to changing climate conditions.
There are many examples of the utility of landraces or old cultivars as potential sources of new genes and alleles for crop breeding. In barley, the genetic variability of old cultivars or landraces was not fully exploited at the beginning of modern breeding.
Among barley diseases, the most relevant in Europe, including Poland, as well as Australia, Asia, and the US, are Blumeria graminis f. sp. hordei (Bgh), which causes powdery mildew; Puccinia hordei (Ph), which causes barley brown rust; P. striiformis f. sp. hordei (Psh), which causes stripe rust; and Puccinia graminis f. sp. tritici (Pgt), which causes stem rust. Seedling tests using sets of isolates virulent or avirulent to known resistance genes provide a means to differentiate resistance to such pathogens. However, the use of specific resistance genes in barley quickly results in the selection of virulent races of Rbgh, Rpg, and Rph.
Adult plant resistance (APR) is considered to be potentially more durable for controlling barley disease development and yield losses. This is complicated due to many factors influencing the final yield losses caused by disease. For instance, during the development stage at which infection first occurs, agroclimatic conditions favorable for pathogen development and the plant’s resistance to the infection all play a role. Because of this, there is a need for developing genome-assisted breeding strategies, in parallel with genomic studies, to understand these traits. The most important stages of plant development connected with final yield are the heading and milky-waxy seed stages [
66].
In this study, we aimed to determine the markers for a qualitative trait such as APR to PM, BBR, and SR in a collection of 431 accessions covering a large range of origins and years. The accessions were stored as a seed sample at the Polish Gene Bank (National Centre for Plant Genetic Resources—NCPGR) at the Plant Breeding and Acclimatization Institute—National Research Institute (PBAI-NIR).
Like other gene banks, the Polish Gene Bank, not only plays an important role in the conservation of plant genetic resources, but also as a source of new genetic alleles. Gene banks are invaluable sources of genetic material for important traits in breeding programs [
40]. Without well curated genetic collections, material stored in gene banks will continue to be underutilized. Therefore, the long-term development plans of traditional gene banks should pivot to becoming biological resource centers and provide access to the wealth of metadata connected with their accessions. This should include phenotypic data (e.g., stress tolerance).
The introgression of new alleles into elite cultivars is performed more simply and effectively with old cultivars and landraces than with wild relatives [
17,
67,
68]. For this reason, old barley cultivars and landraces collected in European countries should be thoroughly mined for new genes [
12,
69,
70].
Based on our study, we found that accessions originating in Northern Europe were more susceptible to PM, SR, and BBR than genotypes originating in other European countries. The genetic gain obtained during breeding programs for PM in the sub-collection originating in Great Britain was the highest. This is consistent with the breeding history in this region, since PM was responsible for significant yield losses during the 1960s and 1970s. This resulted in breeders paying particular attention to this disease in subsequent years.
Previous studies have identified multiple markers associated with powdery mildew resistance on chromosome 7H, which explained 8.9% of the total genetic variance [
71]. Other reports have described PM resistance markers on 3H, 4H, and 5H [
72] as well as novel major-effect APR genes for PM (
Rbgh1,
Rbgh3, and
Rbgh3) on chromosomes 5HL, 7HS, and 1HS, respectively, in landraces [
34]. Our findings suggest that of the five markers associated with the maximum PM disease score, two are on chromosome 2H and may represent novel sources of resistance. APR to PM is very important since the resistance of most high-yielding European spring barley cultivars is determined by
mlo [
20].
Previous studies have reported markers for BBR resistance on chromosome 5H, which correspond to a region containing the APR gene
Rph20 [
33,
37], on chromosome 2H [
37] and the APR-to-BBR resistance gene
Rph23, located on chromosome 7HS [
38]. We confirmed markers localized on the chromosome 5H and 7H and identified additional markers on chromosome 1H and 3H.
However, symptoms of the SR occurred on the barley plants later than the HA; resistance to this disease is among the most serious problems preventing barley grain yield [
73,
74,
75,
76].
Rpg1 is the resistance gene deployed in many barley varieties and provides remarkably durable resistance to most races of this pathogen. The
Rpg4 and
Rpg5 genes were well characterized [
30]. In our study, the frequency of the accessions with disease symptoms at the HA stage was very low, but it was possible to find some markers associated with this disease. This should be confirmed during our next study. Of the 48 markers identified as being associated with SR resistance, 12 were on chromosome 7H, 1 was in the telomeric region of the short arm, and 7 were in the telomeric region of the long arm.
Rpg1 was previously mapped to 7HS [
76].
This study confirmed that old barley cultivars and landraces are an important source of genetic variation and valuable sources of resistance to biotic stresses, including powdery mildew and rusts [
2,
23,
48,
49,
50,
51,
52,
53,
54]. The method of GWAS with DArT data used in this study was proven to be very efficient at identifying markers of APR to PM, BBR, and SR.
This will help plant breeders to use characterized germplasm for APR to PM, BBR, and SR in their breeding programs.
Moreover, the APR markers identified in this study can be used in combination with major R-genes in the same cultivar or in different cultivars with different R-genes, both in spring and winter barley. APR exerts less selective pressure for pathogens on developing plants than R-gene resistance and it can serve as an additional and important resistance factor in case of the breakdown in effectiveness of specific R-genes. Such diverse deployment of many sources of resistance to both APR and R-genes will result in more durable and efficient genetic control of PM, BBR, and SR [
34,
76].
Our study provided genotypic and phenotypic information on a diverse set of previously un-characterized Polish Gene Bank accessions. This work was conducted to develop and implement a national management system for crop plant genetic resources as part of the AGROBANK project at the Polish Gene Bank (NCPGR) (
https://agrobank.cdr.gov.pl/index.php (accessed on 29 October 2021)). It will play a leading role in incorporating the phenotypic and genotypic data of crop plants of agronomic importance to Polish agriculture and food production, such as wheat and barley (
http://dane.agrobank.pcss.pl/jbrowse/ (accessed on 29 October 2021)).





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}