
Characterization of VOCs Emitted by Foliage of Grapevine cv. Isabella for Prospecting Innovative Cropping Systems

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
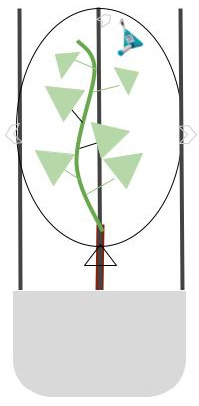
2.2. Collection of VOCs
2.3. First Experiment (Fall 2019)
2.4. Second Experiment (Spring 2020)
2.5. Analysis of Volatile Organic Compounds by GC-MS
2.6. Statistical Analysis
3. Results
3.1. First Experiment (Fall 2019)
3.2. Second Experiment (Spring 2020)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dudareva, N.; Pichersky, E.; Gershenzon, J. Biochemistry of Plant Volatiles. Plant Physiol. 2004, 135, 1893–1902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delory, B.M.; Delaplace, P.; Fauconnier, M.-L.; du Jardin, P. Root-Emitted Volatile Organic Compounds: Can They Mediate Belowground Plant-Plant Interactions? Plant Soil 2016, 402, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Loreto, F.; Schnitzler, J.-P. Abiotic Stresses and Induced BVOCs. Trends Plant Sci. 2010, 15, 154–166. [Google Scholar] [CrossRef]
- Loreto, F.; Dicke, M.; Schnitzler, J.-P.; Turlings, T.C.J. Plant Volatiles and the Environment. Plant Cell Environ. 2014, 37, 1905–1908. [Google Scholar] [CrossRef] [PubMed]
- Farré-Armengol, G.; Fernández-Martínez, M.; Filella, I.; Junker, R.R.; Peñuelas, J. Deciphering the Biotic and Climatic Factors that Influence Floral Scents: A Systematic Review of Floral Volatile Emissions. Front. Plant Sci. 2020, 11, 1154. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M.E. Sites of Synthesis, Biochemistry and Functional Role of Plant Volatiles. S. Afr. J. Bot. 2010, 76, 612–631. [Google Scholar] [CrossRef] [Green Version]
- Hammerbacher, A.; Coutinho, T.A.; Gershenzon, J. Roles of Plant Volatiles in Defence against Microbial Pathogens and Microbial Exploitation of Volatiles. Plant Cell Environ. 2019, 42, 2827–2843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Moraes, C.M.; Lewis, W.J.; Paré, P.W.; Alborn, H.T.; Tumlinson, J.H. Herbivore-Infested Plants Selectively Attract Parasitoids. Nature 1998, 393, 570–573. [Google Scholar] [CrossRef]
- Dicke, M.; Baldwin, I.T. The Evolutionary Context for Herbivore-Induced Plant Volatiles: Beyond the ‘Cry for Help’. Trends Plant Sci. 2010, 15, 167–175. [Google Scholar] [CrossRef]
- Conrath, U.; Beckers, G.J.M.; Langenbach, C.J.G.; Jaskiewicz, M.R. Priming for Enhanced Defense. Annu. Rev. Phytopathol. 2015, 53, 97–119. [Google Scholar] [CrossRef]
- Mauch-Mani, B.; Baccelli, I.; Luna, E.; Flors, V. Defense Priming: An Adaptive Part of Induced Resistance. Annu. Rev. Plant Biol. 2017, 68, 485–512. [Google Scholar] [CrossRef] [Green Version]
- Heil, M.; Karban, R. Explaining Evolution of Plant Communication by Airborne Signals. Trends Ecol. Evol. 2010, 25, 137–144. [Google Scholar] [CrossRef]
- Arimura, G.; Matsui, K.; Takabayashi, J. Chemical and Molecular Ecology of Herbivore-Induced Plant Volatiles: Proximate Factors and Their Ultimate Functions. Plant Cell Physiol. 2009, 50, 911–923. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Zhang, K.; Wu, Z.; Xu, J.; Erb, M. Plant Volatiles as Regulators of Plant Defense and Herbivore Immunity: Molecular Mechanisms and Unanswered Questions. Curr. Opin. Insect Sci. 2021, 44, 82–88. [Google Scholar] [CrossRef]
- Weeraddana, C.D.S.; Manolii, V.P.; Strelkov, S.E.; de la Mata, A.P.; Harynuk, J.J.; Evenden, M.L. Infection of Canola by the Root Pathogen Plasmodiophora brassicae Increases Resistance to Aboveground Herbivory by Bertha Armyworm, Mamestra configurata Walker (Lepidoptera: Noctuidae). Plant Sci. 2020, 300, 110625. [Google Scholar] [CrossRef]
- International Organisation of Vine and Wine (OIV, 2017). Available online: https://www.oiv.int/public/medias/5888/en-distribution-of-the-worlds-grapevine-varieties.pdf (accessed on 17 December 2021).
- Petretto, G.L.; Mercenaro, L.; Urgeghe, P.P.; Fadda, C.; Valentoni, A.; Del Caro, A. Grape and Wine Composition in Vitis vinifera L. cv. Cannonau Explored by GC-MS and Sensory Analysis. Foods 2021, 10, 101. [Google Scholar] [CrossRef]
- Previtali, P.; Dokoozlian, N.; Capone, D.L.; Wilkinson, K.L.; Ford, C.M. Exploratory Study of Sugar and C6 Compounds in Single Berries of Grapevine (Vitis vinifera L.) cv. Cabernet Sauvignon throughout Ripening. Aust. J. Grape Wine Res. 2021, 27, 194–205. [Google Scholar] [CrossRef]
- Rahman, F.U.; Nawaz, M.A.; Liu, R.; Sun, L.; Jiang, J.; Fan, X.; Liu, C.; Zhang, Y. Evaluation of Volatile Aroma Compounds from Chinese Wild Grape Berries by Headspace-SPME with GC-MS. Food Sci. Technol 2021. [Google Scholar] [CrossRef]
- Matarese, F.; Cuzzola, A.; Scalabrelli, G.; D’Onofrio, C. Expression of Terpene Synthase Genes Associated with the Formation of Volatiles in Different Organs of Vitis vinifera. Phytochem. 2014, 105, 12–24. [Google Scholar] [CrossRef]
- Lawo, N.C.; Weingart, G.J.F.; Schuhmacher, R.; Forneck, A. The Volatile Metabolome of Grapevine Roots: First Insights into the Metabolic Response upon Phylloxera Attack. Plant Physiol. Biochem. 2011, 49, 1059–1063. [Google Scholar] [CrossRef] [Green Version]
- Zito, P.; Scrima, A.; Sajeva, M.; Carimi, F.; Dötterl, S. Dimorphism in Inflorescence Scent of Dioecious Wild Grapevine. Biochem. Syst. Ecol. 2016, 66, 58–62. [Google Scholar] [CrossRef]
- Brantjes, N.B.M. Pollinator Attraction of Vitis vinifera subsp. Silvestris. VITIS J. Grapevine Res. 2016, 17, 229. [Google Scholar] [CrossRef]
- Zito, P.; Serraino, F.; Carimi, F.; Tavella, F.; Sajeva, M. Inflorescence-Visiting Insects of a Functionally Dioecious Wild Grapevine (Vitis vinifera subsp. sylvestris). Genet. Resour Crop. Evol. 2018, 65, 1329–1335. [Google Scholar] [CrossRef]
- Algarra Alarcon, A.; Lazazzara, V.; Cappellin, L.; Bianchedi, P.L.; Schuhmacher, R.; Wohlfahrt, G.; Pertot, I.; Biasioli, F.; Perazzolli, M. Emission of Volatile Sesquiterpenes and Monoterpenes in Grapevine Genotypes Following Plasmopara viticola Inoculation in Vitro: VOCs of Grapevines with Plasmopara viticola. J. Mass Spectrom. 2015, 50, 1013–1022. [Google Scholar] [CrossRef]
- Chitarrini, G.; Soini, E.; Riccadonna, S.; Franceschi, P.; Zulini, L.; Masuero, D.; Vecchione, A.; Stefanini, M.; Di Gaspero, G.; Mattivi, F.; et al. Identification of Biomarkers for Defense Response to Plasmopara viticola in a Resistant Grape Variety. Front. Plant Sci. 2017, 8, 1524. [Google Scholar] [CrossRef]
- Lazazzara, V.; Bueschl, C.; Parich, A.; Pertot, I.; Schuhmacher, R.; Perazzolli, M. Downy Mildew Symptoms on Grapevines Can Be Reduced by Volatile Organic Compounds of Resistant Genotypes. Sci. Rep. 2018, 8, 1618. [Google Scholar] [CrossRef] [Green Version]
- Ricciardi, V.; Marcianò, D.; Sargolzaei, M.; Maddalena, G.; Maghradze, D.; Tirelli, A.; Casati, P.; Bianco, P.A.; Failla, O.; Fracassetti, D.; et al. From Plant Resistance Response to the Discovery of Antimicrobial Compounds: The Role of Volatile Organic Compounds (VOCs) in Grapevine Downy Mildew Infection. Plant Physiol. Biochem. 2021, 160, 294–305. [Google Scholar] [CrossRef]
- Chalal, M.; Winkler, J.B.; Gourrat, K.; Trouvelot, S.; Adrian, M.; Schnitzler, J.-P.; Jamois, F.; Daire, X. Sesquiterpene Volatile Organic Compounds (VOCs) Are Markers of Elicitation by Sulfated Laminarine in Grapevine. Front. Plant Sci. 2015, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Tasin, M.; Anfora, G.; Ioriatti, C.; Carlin, S.; De Cristofaro, A.; Schmidt, S.; Bengtsson, M.; Versini, G.; Witzgall, P. Antennal and Behavioral Responses of Grapevine Moth Lobesia botrana Females to Volatiles from Grapevine. J. Chem. Ecol. 2005, 31, 77–87. [Google Scholar] [CrossRef]
- Giacomuzzi, V.; Cappellin, L.; Nones, S.; Khomenko, I.; Biasioli, F.; Knight, A.L.; Angeli, S. Diel Rhythms in the Volatile Emission of Apple and Grape Foliage. Phytochemistry 2017, 138, 104–115. [Google Scholar] [CrossRef]
- Velásquez, A.; Valenzuela, M.; Carvajal, M.; Fiaschi, G.; Avio, L.; Giovannetti, M.; D’Onofrio, C.; Seeger, M. The Arbuscular Mycorrhizal Fungus Funneliformis mosseae Induces Changes and Increases the Concentration of Volatile Organic Compounds in Vitis vinifera cv. Sangiovese Leaf Tissue. Plant Physiol. Biochem. 2020, 155, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Loughrin, J.H.; Potter, D.A.; Hamilton-Kemp, T.R.; Byers, M.E. Diurnal Emission of Volatile Compounds by Japanese Beetle-Damaged Grape Leaves. Phytochemistry 1997, 45, 919–923. [Google Scholar] [CrossRef]
- Lucchi, A.; Sambado, P.; Royo, A.B.J.; Bagnoli, B.; Benelli, G. Lobesia botrana Males Mainly Fly at Dusk: Video Camera-Assisted Pheromone Traps and Implications for Mating Disruption. J. Pest. Sci. 2018, 91, 1327–1334. [Google Scholar] [CrossRef]
- Stellwaag, F. Neue Forschungen über Mottenflug, Eiablage und Dauer der Traubenwickler. Der Dtsch. Weinbau 1943, 22, 203–206. [Google Scholar]
- Gessler, C.; Pertot, I.; Perazzolli, M. Plasmopara viticola: A Review of Knowledge on Downy Mildew of Grapevine and Effective Disease Management. Phytopathol. Mediterr. 2011, 50, 3–44. [Google Scholar]
- Jordão, A.M.; Botelho, R.V. Vitis: Biology and Species, 1st ed.; Nova Science Publishers: Hauppauge, NY, USA, 2020. [Google Scholar]
- Pacifico, S.; D’Abrosca, B.; Scognamiglio, M.; Gallicchio, M.; Potenza, N.; Piccolella, S.; Russo, A.; Monaco, P.; Fiorentino, A. Metabolic Profiling of Strawberry Grape (Vitis × labruscana cv. ‘Isabella’) Components by Nuclear Magnetic Resonance (NMR) and Evaluation of Their Antioxidant and Antiproliferative Properties. J. Agric. Food Chem. 2011, 59, 7679–7687. [Google Scholar] [CrossRef]
- Hildebrandt, A.; Guillamón, M.; Lacorte, S.; Tauler, R.; Barceló, D. Impact of Pesticides Used in Agriculture and Vineyards to Surface and Groundwater Quality (North Spain). Water Res. 2008, 42, 3315–3326. [Google Scholar] [CrossRef]
- Fantke, P.; Friedrich, R.; Jolliet, O. Health Impact and Damage Cost Assessment of Pesticides in Europe. Environ. Intern. 2012, 49, 9–17. [Google Scholar] [CrossRef]
- Schusterova, D.; Hajslova, J.; Kocourek, V.; Pulkrabova, J. Pesticide Residues and Their Metabolites in Grapes and Wines from Conventional and Organic Farming System. Foods 2021, 10, 307. [Google Scholar] [CrossRef]
- Brilli, F.; Loreto, F.; Baccelli, I. Exploiting Plant Volatile Organic Compounds (VOCs) in Agriculture to Improve Sustainable Defense Strategies and Productivity of Crops. Front. Plant Sci. 2019, 10, 264. [Google Scholar] [CrossRef]
- Kigathi, R.N.; Weisser, W.W.; Reichelt, M.; Gershenzon, J.; Unsicker, S.B. Plant Volatile Emission Depends on the Species Composition of the Neighboring Plant Community. BMC Plant Biol. 2019, 19, 58. [Google Scholar] [CrossRef]
- Dini, I.; Marra, R.; Cavallo, P.; Pironti, A.; Sepe, I.; Troisi, J.; Scala, G.; Lombari, P.; Vinale, F. Trichoderma Strains and Metabolites Selectively Increase the Production of Volatile Organic Compounds (VOCs) in Olive Trees. Metabolites 2021, 11, 213. [Google Scholar] [CrossRef]
- Joos, P.E.; Godoi, A.F.L.; De Jong, R.; de Zeeuw, J.; Van Grieken, R. Trace Analysis of Benzene, Toluene, Ethylbenzene and Xylene Isomers in Environmental Samples by Low-Pressure Gas Chromatography–Ion Trap Mass Spectrometry. J. Chromatogr. A 2003, 985, 191–196. [Google Scholar] [CrossRef]
- Castro-Marín, A.; Buglia, A.G.; Riponi, C.; Chinnici, F. Volatile and Fixed Composition of Sulphite-Free White Wines Obtained after Fermentation in the Presence of Chitosan. LWT 2018, 93, 174–180. [Google Scholar] [CrossRef]
- Rid, M.; Markheiser, A.; Hoffmann, C.; Gross, J. Waxy Bloom on Grape Berry Surface Is One Important Factor for Oviposition of European Grapevine Moths. J. Pest. Sci. 2018, 91, 1225–1239. [Google Scholar] [CrossRef]
- Portillo-Estrada, M.; Ariza-Carricondo, C.; Ceulemans, R. Outburst of Senescence-Related VOC Emissions from a Bioenergy Poplar Plantation. Plant Physiol. Biochem. 2020, 148, 324–332. [Google Scholar] [CrossRef]
- Alex, A.A.; Dommun, D.F.; Kubmarawa, D.; Okechukwu, J.O.; Victor, E.I. Antioxidant Activities And Phytochemical Screening Of Peanut (Arachis hypogea) Leaves. Afr. J. Environ. Nat. Sci. Res. 2020, 3, 28–37. [Google Scholar]
- Junior, S.B.; Henrique Março, P.; Valderrama, P.; Cardoso Damasceno, F.; Silvana Aranda, M.; Alcaraz Zini, C.; Bastos Caramão, E.; Melo, A.M.T.; Teixiera Filho, J.; Teixeira Godoy, H. Analysis of Volatile Compounds in Capsicum spp. by Headspace Solid-Phase Microextraction and GC × GC-TOFMS. Anal. Methods 2015, 7, 521–529. [Google Scholar] [CrossRef] [Green Version]
- Schade, F.; Legge, R.L.; Thompson, J.E. Fragrance Volatiles of Developing and Senescing Carnation Flowers. Phytochemistry 2001, 56, 703–710. [Google Scholar] [CrossRef]
- Gfrerer, E.; Laina, D.; Gibernau, M.; Fuchs, R.; Happ, M.; Tolasch, T.; Trutschnig, W.; Hörger, A.C.; Comes, H.P.; Dötterl, S. Floral Scents of a Deceptive Plant Are Hyperdiverse and Under Population-Specific Phenotypic Selection. Front. Plant Sci. 2021, 12, 1910. [Google Scholar] [CrossRef]
- Tabanca, N.; Kürkcüoglu, M.; Baser, K.H.C.; Tümen, G.; Duman, H. Composition of the Essential Oils of Satureja spinosa L. J. Essent. Oil Res. 2004, 16, 127–128. [Google Scholar] [CrossRef]
- Forlani, G.; Occhipinti, A.; Bossi, S.; Bertea, C.M.; Varese, C.; Maffei, M.E. Magnaporthe oryzae Cell Wall Hydrolysate Induces ROS and Fungistatic VOCs in Rice Cell Cultures. J. Plant Physiol. 2011, 168, 2041–2047. [Google Scholar] [CrossRef]
- Germinara, G.S.; Pistillo, M.; Griffo, R.; Garonna, A.P.; Di Palma, A. Electroantennographic Responses of Aromia bungii (Faldermann, 1835) (Coleoptera, Cerambycidae) to a Range of Volatile Compounds. Insects 2019, 10, 274. [Google Scholar] [CrossRef] [Green Version]
- Schorkopf, D.L.P. Male Meliponine Bees (Scaptotrigona aff. depilis) Produce Alarm Pheromones to Which Workers Respond with Fight and Males with Flight. J. Comp. Physiol. A 2016, 202, 667–678. [Google Scholar] [CrossRef]
- Wang, Y.; Li, H.-Q.; Gao, X.-T.; Lu, H.-C.; Peng, W.-T.; Chen, W.; Li, S.-D.; Li, S.-P.; Duan, C.-Q.; Wang, J. Influence of Attenuated Reflected Solar Radiation from the Vineyard Floor on Volatile Compounds in Cabernet Sauvignon Grapes and Wines of the North Foot of Mt. Tianshan. Food Res. Int. 2020, 137, 109688. [Google Scholar] [CrossRef]
- Arimura, G.; Shiojiri, K.; Karban, R. Acquired Immunity to Herbivory and Allelopathy Caused by Airborne Plant Emissions. Phytochemistry 2010, 71, 1642–1649. [Google Scholar] [CrossRef]
- Weeraddana, C.; Wise, I.; Lamb, R.; Wolfe, S.; Wist, T.; McCartney, C.; Costamagna, A. A Laboratory Method for Mass Rearing the Orange Wheat Blossom Midge, Sitodiplosis mosellana (Diptera: Cecidomyiidae). Can. Entomol. 2021, 153, 828–836. [Google Scholar] [CrossRef]
- Birkett, M.A.; Bruce, T.J.; Martin, J.L.; Smart, L.E.; Oakley, J.; Wadhams, L.J. Responses of Female Orange Wheat Blossom Midge, Sitodiplosis mosellana, to Wheat Panicle Volatiles. J. Chem. Ecol. 2004, 30, 1319–1328. [Google Scholar] [CrossRef]
- Zeng, L.; Wang, X.; Kang, M.; Dong, F.; Yang, Z. Regulation of the Rhythmic Emission of Plant Volatiles by the Circadian Clock. Int. J. Mol. Sci. 2017, 18, 2408. [Google Scholar] [CrossRef] [Green Version]
- Tingey, D.T.; Manning, M.; Grothaus, L.C.; Burns, W.F. The Influence of Light and Temperature on Isoprene Emission Rates from Live Oak. Physiol. Plant 1979, 47, 112–118. [Google Scholar] [CrossRef]
- Schuh, G.; Heiden, A.C.; Hoffmann, T.; Kahl, J.; Rockel, P.; Rudolph, J.; Wildt, J. Emissions of Volatile Organic Compounds from Sunflower and Beech: Dependence on Temperature and Light Intensity. J. Atmos. Chem. 1997, 27, 291–318. [Google Scholar] [CrossRef]
- Ghirardo, A.; Koch, K.; Taipale, R.; Zimmer, I.; Schnitzler, J.-P.; Rinne, J. Determination of de Novo and Pool Emissions of Terpenes from Four Common Boreal/Alpine Trees by 13CO2 Labelling and PTR-MS Analysis. Plant Cell Environ. 2010. [Google Scholar] [CrossRef] [PubMed]
- Kono, A.; Shimizu, T. Leaf Trichomes as an Effective Structure for Disease Resistance: The Case of Grapevine Downy Mildew. Jpn. Agric. Res. Q. JARQ 2020, 54, 293–298. [Google Scholar] [CrossRef]
- Özer, N.; Şabudak, T.; Özer, C.; Gindro, K.; Schnee, S.; Solak, E. Investigations on the Role of Cuticular Wax in Resistance to Powdery Mildew in Grapevine. J. Gen. Plant Pathol. 2017, 83, 316–328. [Google Scholar] [CrossRef]
- Markus Lange, B.; Turner, G.W. Terpenoid Biosynthesis in Trichomes—Current Status and Future Opportunities. Plant Biotechnol. J. 2013, 11, 2–22. [Google Scholar] [CrossRef]
- Tissier, A.; Morgan, J.A.; Dudareva, N. Plant Volatiles: Going ‘In’ but Not ‘Out’ of Trichome Cavities. Trends Plant Sci. 2017, 22, 930–938. [Google Scholar] [CrossRef]
- Gershenzon, J.; Dudareva, N. The Function of Terpene Natural Products in the Natural World. Nat. Chem. Biol. 2007, 3, 408–414. [Google Scholar] [CrossRef]
- Yazaki, K.; Arimura, G.; Ohnishi, T. ‘Hidden’ Terpenoids in Plants: Their Biosynthesis, Localization and Ecological Roles. Plant Cell Physiol. 2017, 58, 1615–1621. [Google Scholar] [CrossRef] [Green Version]
- Rosenkranz, M.; Chen, Y.; Zhu, P.; Vlot, A.C. Volatile Terpenes—Mediators of Plant-to-Plant Communication. Plant J. 2021, 108, 617–631. [Google Scholar] [CrossRef]
- Mofikoya, A.O.; Bui, T.N.T.; Kivimäenpää, M.; Holopainen, J.K.; Himanen, S.J.; Blande, J.D. Foliar Behaviour of Biogenic Semi-Volatiles: Potential Applications in Sustainable Pest Management. Arthropod-Plant Interact. 2019, 13, 193–212. [Google Scholar] [CrossRef] [Green Version]
- Copolovici, L.; Niinemets, Ü. Temperature Dependencies of Henry’s Law Constants for Different Plant Sesquiterpenes. Chemosphere 2015, 138, 751–757. [Google Scholar] [CrossRef] [Green Version]
- Vallat, A.; Gu, H.; Dorn, S. How Rainfall, Relative Humidity and Temperature Influence Volatile Emissions from Apple Trees in Situ. Phytochemistry 2005, 66, 1540–1550. [Google Scholar] [CrossRef]
- Wang, X.; Tao, Y.; Wu, Y.; An, R.; Yue, Z. Aroma Compounds and Characteristics of Noble-Rot Wines of Chardonnay Grapes Artificially Botrytized in the Vineyard. Food Chem. 2017, 226, 41–50. [Google Scholar] [CrossRef]
- Wilson, C.L.; Solar, J.M.; El Ghaouth, A.; Wisniewski, M.E. Rapid Evaluation of Plant Extracts and Essential Oils for Antifungal Activity Against Botrytis cinerea. Plant Dis. 1997, 81, 204–210. [Google Scholar] [CrossRef] [Green Version]
- Santino, A.; Poltronieri, P.; Mita, G. Advances on Plant Products with Potential to Control Toxigenic Fungi: A Review. Food Addit. Contam. 2005, 22, 389–395. [Google Scholar] [CrossRef]
- Frost, C.J.; Mescher, M.C.; Dervinis, C.; Davis, J.M.; Carlson, J.E.; De Moraes, C.M. Priming Defense Genes and Metabolites in Hybrid Poplar by the Green Leaf Volatile Cis-3-hexenyl Acetate. New Phytol. 2008, 180, 722–734. [Google Scholar] [CrossRef]
- Rodov, V.; Nafussi, B.; Ben-Yehoshua, S. Essential oil components as potential means to control Penicillium digitatum Pers. (Sacc.) and other postharvest pathogens of citrus fruit. Fresh Prod. 2011, 5, 43–50. [Google Scholar]
- Ameye, M.; Audenaert, K.; De Zutter, N.; Steppe, K.; Van Meulebroek, L.; Vanhaecke, L.; De Vleesschauwer, D.; Haesaert, G.; Smagghe, G. Priming of Wheat with the Green Leaf Volatile Z-3-Hexenyl Acetate Enhances Defense against Fusarium graminearum But Boosts Deoxynivalenol Production. Plant Physiol. 2015, 167, 1671–1684. [Google Scholar] [CrossRef] [Green Version]
- Hachlafi, N.E.; Aanniz, T.; Menyiy, N.E.; Baaboua, A.E.; Omari, N.E.; Balahbib, A.; Shariati, M.A.; Zengin, G.; Fikri-Benbrahim, K.; Bouyahya, A. In Vitro and in Vivo Biological Investigations of Camphene and Its Mechanism Insights: A Review. Food Rev. Int. 2021, 1–28. [Google Scholar] [CrossRef]
- Di Francesco, A.; Ugolini, L.; Lazzeri, L.; Mari, M. Production of Volatile Organic Compounds by Aureobasidium pullulans as a Potential Mechanism of Action against Postharvest Fruit Pathogens. Biol. Control 2015, 81, 8–14. [Google Scholar] [CrossRef]
- Quintana-Rodriguez, E.; Morales-Vargas, A.T.; Molina-Torres, J.; Ádame-Alvarez, R.M.; Acosta-Gallegos, J.A.; Heil, M. Plant Volatiles Cause Direct, Induced and Associational Resistance in Common Bean to the Fungal Pathogen Colletotrichum lindemuthianum. J. Ecol. 2015, 103, 250–260. [Google Scholar] [CrossRef]
- Ninkovic, V.; Dahlin, I.; Vucetic, A.; Petrovic-Obradovic, O.; Glinwood, R.; Webster, B. Volatile Exchange between Undamaged Plants - a New Mechanism Affecting Insect Orientation in Intercropping. PLoS ONE 2013, 8, e69431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Mallik, A.; Zeng, R.S. Control of Panama Disease of Banana by Rotating and Intercropping with Chinese Chive (Allium tuberosum Rottler): Role of Plant Volatiles. J. Chem. Ecol. 2013, 39, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; de Boer, W.; Zhang, Y.; Ding, C.; Zhang, T.; Wang, X. Suppression of Soil-Borne Fusarium Pathogens of Peanut by Intercropping with the Medicinal Herb Atractylodes lancea. Soil Biol. Biochem. 2018, 116, 120–130. [Google Scholar] [CrossRef]
- Lv, J.; Dong, Y.; Dong, K.; Zhao, Q.; Yang, Z.; Chen, L. Intercropping with Wheat Suppressed Fusarium Wilt in Faba Bean and Modulated the Composition of Root Exudates. Plant Soil 2020, 448, 153–164. [Google Scholar] [CrossRef]
- Luo, C.; Ma, L.; Zhu, J.; Guo, Z.; Dong, K.; Dong, Y. Effects of Nitrogen and Intercropping on the Occurrence of Wheat Powdery Mildew and Stripe Rust and the Relationship with Crop Yield. Front. Plant Sci. 2021, 12, 637393. [Google Scholar] [CrossRef]
- Kulakiotu, E.K.; Thanassoulopoulos, C.C.; Sfakiotakis, E.M. Biological Control of Botrytis cinerea by Volatiles of ‘Isabella’ Grapes. Phytopathology 2004, 94, 924–931. [Google Scholar] [CrossRef] [Green Version]
- Kulakiotu, E.K.; Thanassoulopoulos, C.C.; Sfakiotakis, E.M. Postharvest Biological Control of Botrytis cinerea on Kiwifruit by Volatiles of [Isabella] Grapes. Phytopathology 2004, 94, 1280–1285. [Google Scholar] [CrossRef] [Green Version]
- Tessarin, P.; Parpinello, G.P.; Rombolà, A.D. Physiological and Enological Implications of Postveraison Trimming in an Organically-Managed Sangiovese Vineyard. Am. J. Enol. Vitic. 2018, 69, 59–69. [Google Scholar] [CrossRef]
- Zeng, W.; Melotto, M.; He, S.Y. Plant Stomata: A Checkpoint of Host Immunity and Pathogen Virulence. Curr. Opin. Biotechnol. 2010, 21, 599–603. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| No. | RI | Compound | Diurnal | Nocturnal | Diurnal | Nocturnal | ||
|---|---|---|---|---|---|---|---|---|
| Fall 2019 | Spring 2020 | |||||||
| Terpenes | ||||||||
| 1 | 1078 | Camphene | 0.0014 ± 0.0008 | 0.0012 ± 0.0005 | 0.0058 ± 0.0022 | 0.0011 ± 0.0006 | * | |
| 2 | 1105 | β-pinene a | 0.0056 ± 0.0014 | 0.0050 ± 0.0011 | 0.0795 ± 0.0181 | 0.0034 ± 0.0010 | ** | |
| Hydrocarbons | ||||||||
| 3 | 1126 | Ethylbenzene | nd | 0.0014 ± 0.0008 | * | nd | nd | |
| 4 | 1202 | Dodecane a | nd | nd | 0.5596 ± 0.2272 | 0.3431 ± 0.0352 | * | |
| 5 | 1240 | 1-hexadecene | nd | nd | nd | 0.0205 ± 0.0246 | ||
| 6 | 1257 | trans-3-dodecene | 0.0018 ± 0.0014 | 0.0017 ± 0.0004 | nd | nd | ||
| 7 | 1298 | Tridecane a | nd | nd | 0.3837 ± 0.2009 | 0.1641 ± 0.0514 | * | |
| 8 | 1314 | Butylbenzene | nd | 0.0010 ± 0.0014 | nd | nd | ||
| 9 | 1392 | 5,5 dibutylnonane | nd | nd | 0.0748 ± 0.0773 | 0.0360 ± 0.0106 | ||
| 10 | 1402 | Tetradecane a | nd | nd | 0.5657 ± 0.2668 | 0.2092 ± 0.1280 | * | |
| Green leaf volatiles (GLVs) | ||||||||
| 11 | 1321 | cis-3-hexenyl acetate a | 0.0003 ± 0.0003 | 0.0016 ± 0.0024 | nd | 0.0132 ± 0.0209 | ||
| Esters | ||||||||
| 12 | 1453 | 2-butoxyethyl acetate | nd | nd | 0.0636 ± 0.0143 | 0.0417 ± 0.0147 | ||
| 13 | 1495 | Ethyl 2-methyllactate | nd | 0.0065 ± 0.0030 | * | nd | nd | |
| Ketones | ||||||||
| 14 | 1364 | 4-hydroxy-4-methyl-2-pentanone a | nd | nd | 0.2647 ± 0.2095 | nd | ** | |
| 15 | 1393 | 2-nonanone | nd | 0.0004 ± 0.0006 | nd | nd | ||
| Alcohols | ||||||||
| 16 | 1311 | 2-hexanola | 0.0167 ± 0.0289 | 0.0137 ± 0.0051 | 0.0120 ± 0.0123 | nd | * | |
| 17 | 1450 | 1-heptanola | 0.0183 ± 0.0048 | 0.0074 ± 0.0026 | * | 0.0822 ± 0.0273 | 0.0227 ± 0.0184 | * |
| 18 | 1455 | 2-butoxyethanol a | nd | 0.0079 ± 0.0029 | ** | nd | nd | |
| 19 | 1460 | 3-ethyl-2-heptanol | nd | 0.0026 ± 0.0006 | * | nd | nd | |
| 20 | 1479 | 2-ethylhexanol a | nd | 0.0015 ± 0.0014 | nd | nd | ||
| 21 | 1481 | 3-nonanol | nd | nd | 0.0246 ± 0.0084 | 0.0067 ± 0.0084 | ** | |
| 22 | 1483 | 2-hexadecanol | 0.0092 ± 0.0045 | nd | * | nd | nd | |
| 23 | 1511 | 2-nonanol a | 0.0133 ± 0.0050 | 0.0028 ± 0.0002 | * | 0.0112 ± 0.0058 | nd | *** |
| 24 | 1557 | 1-octanol a | 0.0019 ± 0.0020 | 0.0010 ± 0.0006 | 0.0132 ± 0.0111 | nd | * | |
| 25 | 1939 | Phenethyl alcohol a | 0.0014 ± 0.0025 | nd | 0.0135 ± 0.0167 | nd | ||
| Total Sum | 0.0698 ± 0.0325 | 0.0556 ± 0.0235 | 2.1542 ± 0.7873 | 0.8619 ± 0.1718 | ** | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Declet, A.; Castro-Marín, A.; Moretti Conti, L.; Lombini, A.; Chinnici, F.; Rombolà, A.D. Characterization of VOCs Emitted by Foliage of Grapevine cv. Isabella for Prospecting Innovative Cropping Systems. Agronomy 2022, 12, 272. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020272
Rodríguez-Declet A, Castro-Marín A, Moretti Conti L, Lombini A, Chinnici F, Rombolà AD. Characterization of VOCs Emitted by Foliage of Grapevine cv. Isabella for Prospecting Innovative Cropping Systems. Agronomy. 2022; 12(2):272. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020272
Chicago/Turabian StyleRodríguez-Declet, Arleen, Antonio Castro-Marín, Luca Moretti Conti, Alessandra Lombini, Fabio Chinnici, and Adamo Domenico Rombolà. 2022. "Characterization of VOCs Emitted by Foliage of Grapevine cv. Isabella for Prospecting Innovative Cropping Systems" Agronomy 12, no. 2: 272. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020272