
13C Labelling of Litter Added to Tea (Camellia sinensis L.) Plantation Soil Reveals a Significant Positive Priming Effect That Leads to Less Soil Organic Carbon Accumulation

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Soil
2.2. 13C Labeling of Tea Plants
2.3. Experimental Design
2.4. CO2 and δ13C Measurement
2.5. β-glucosidase Activity Analysis
2.6. PLFA and δ13C Measurements
2.7. Statistical Analyses
3. Results
3.1. Changes in SOC
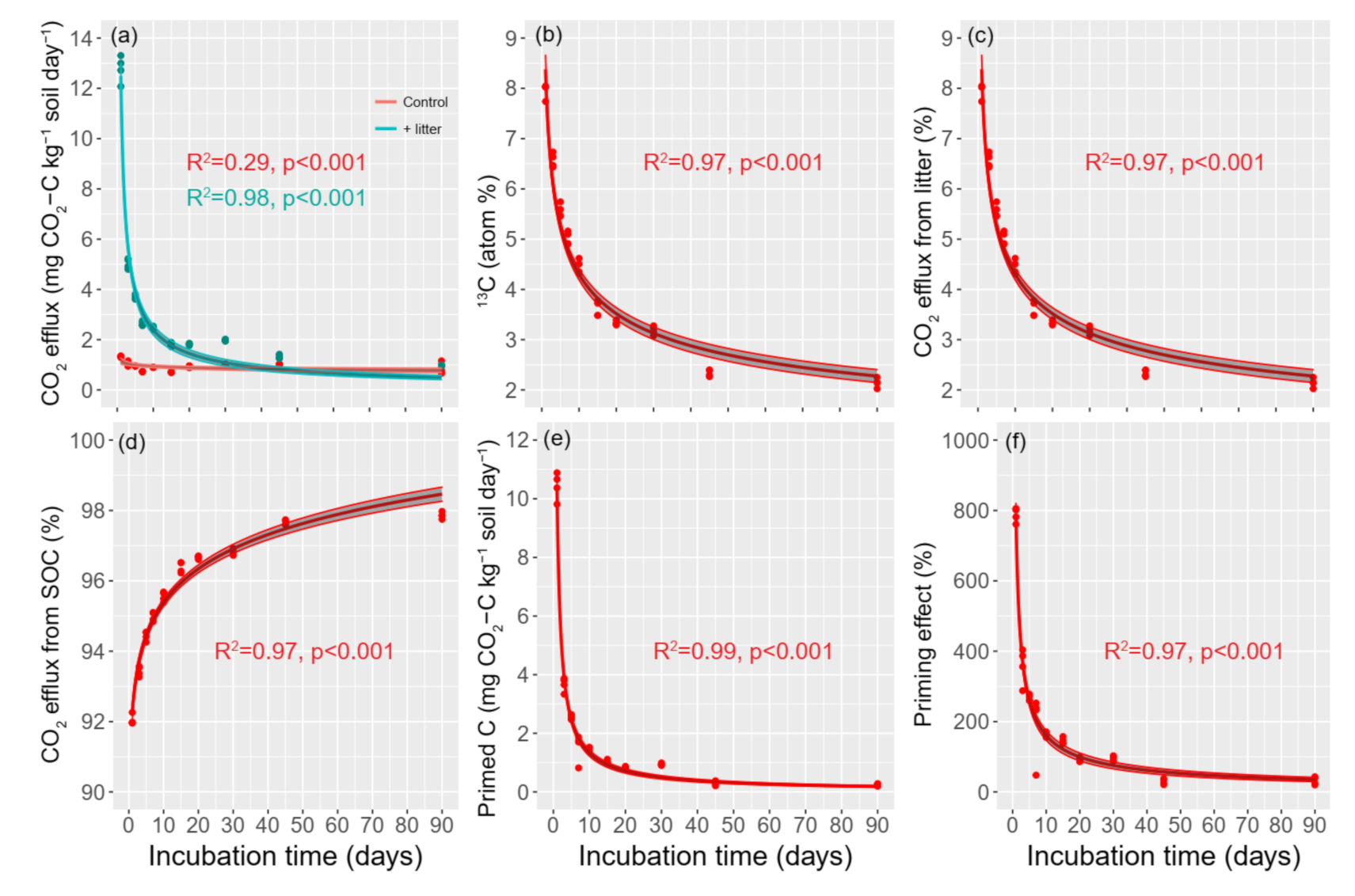
3.2. Changes in CO2 Efflux
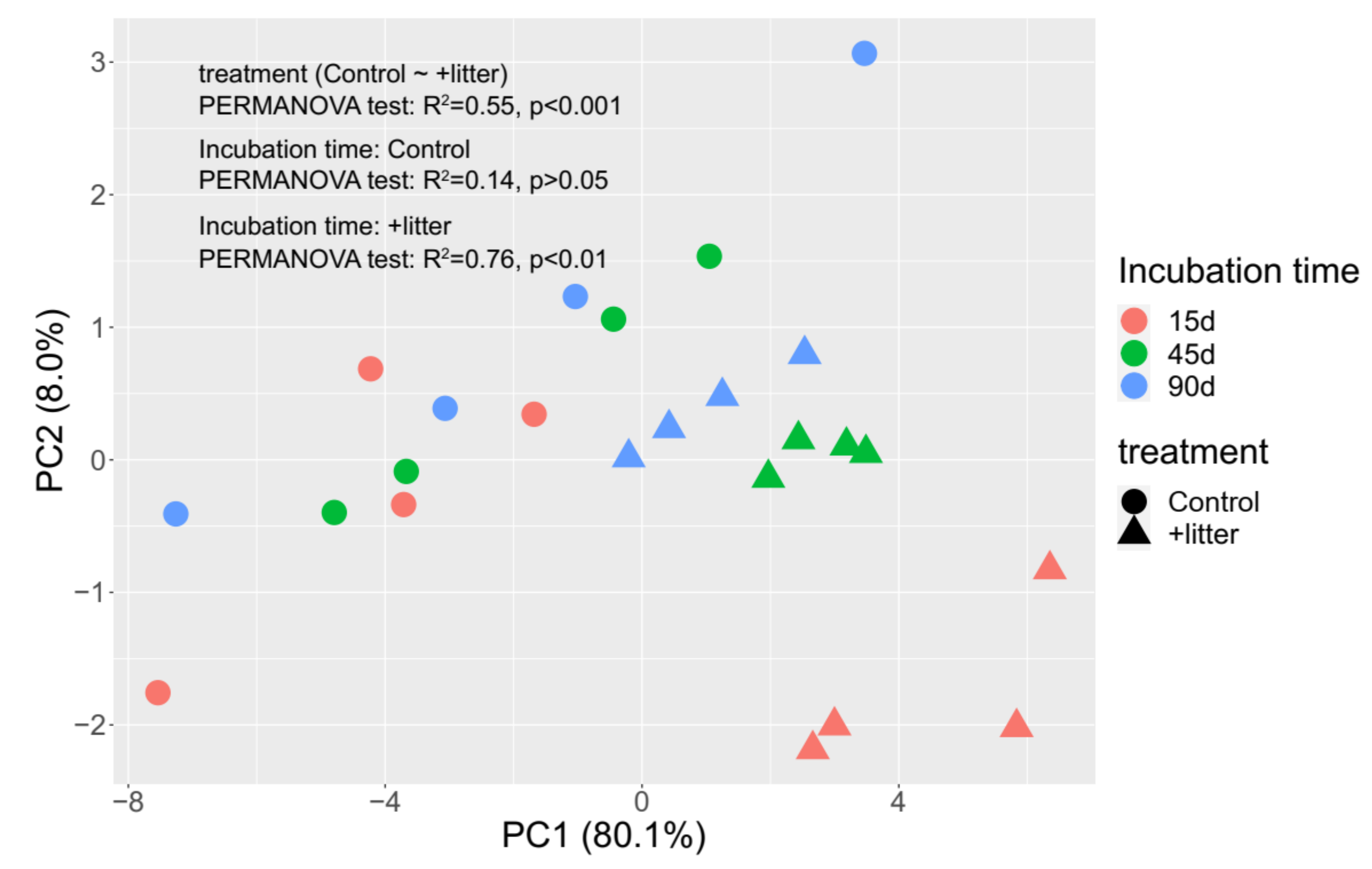
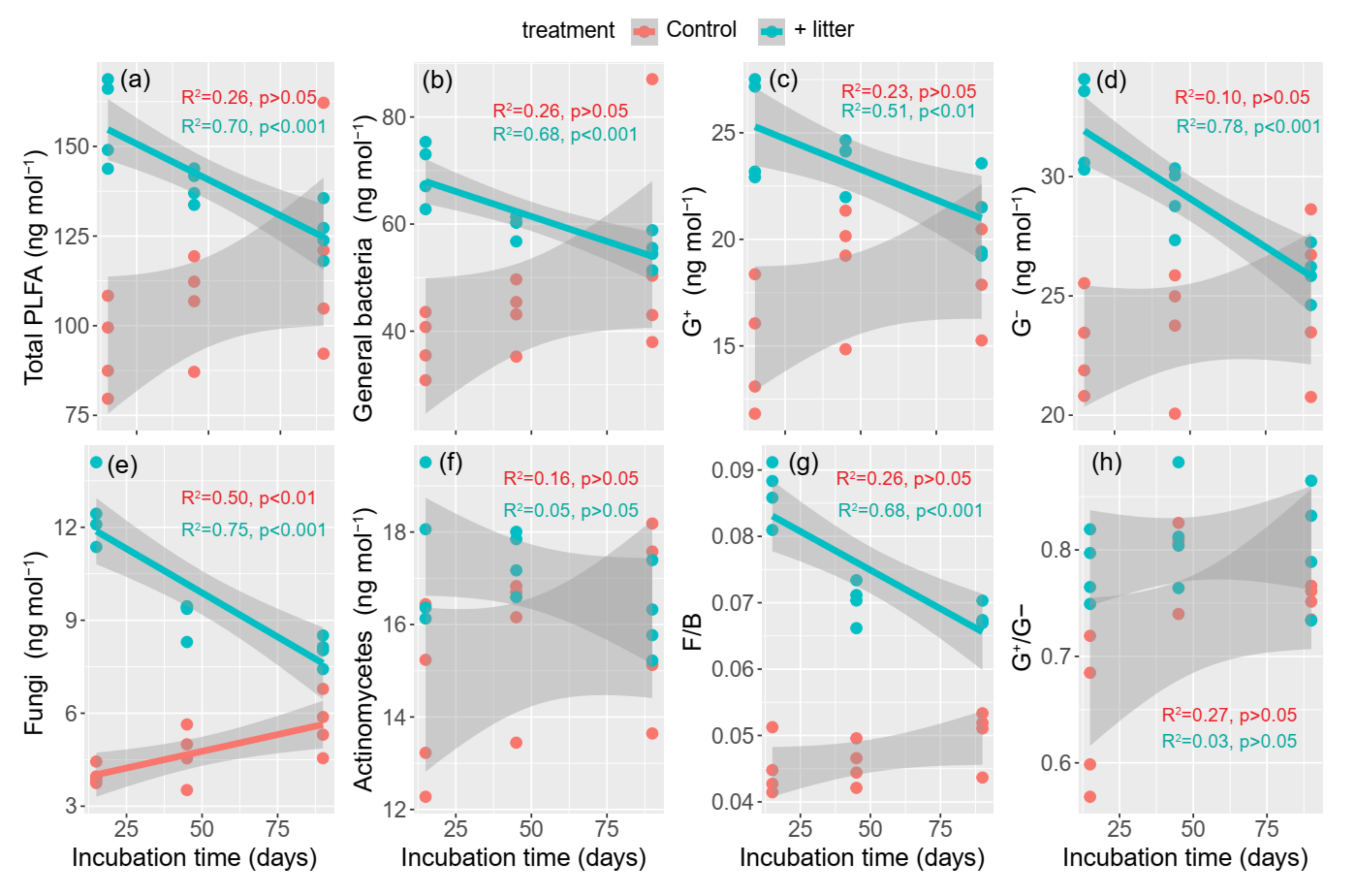
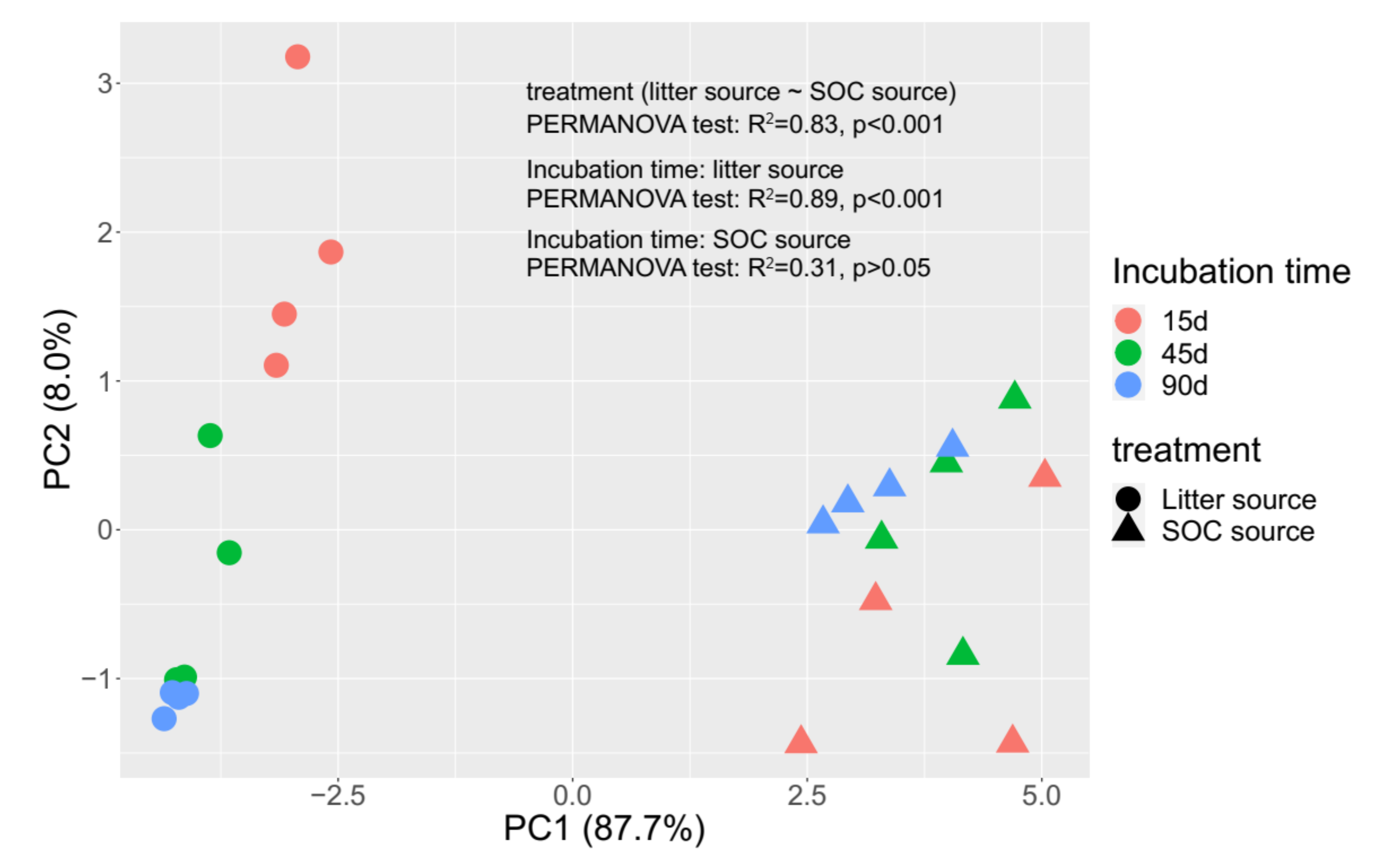
3.3. Microbial Community Composition
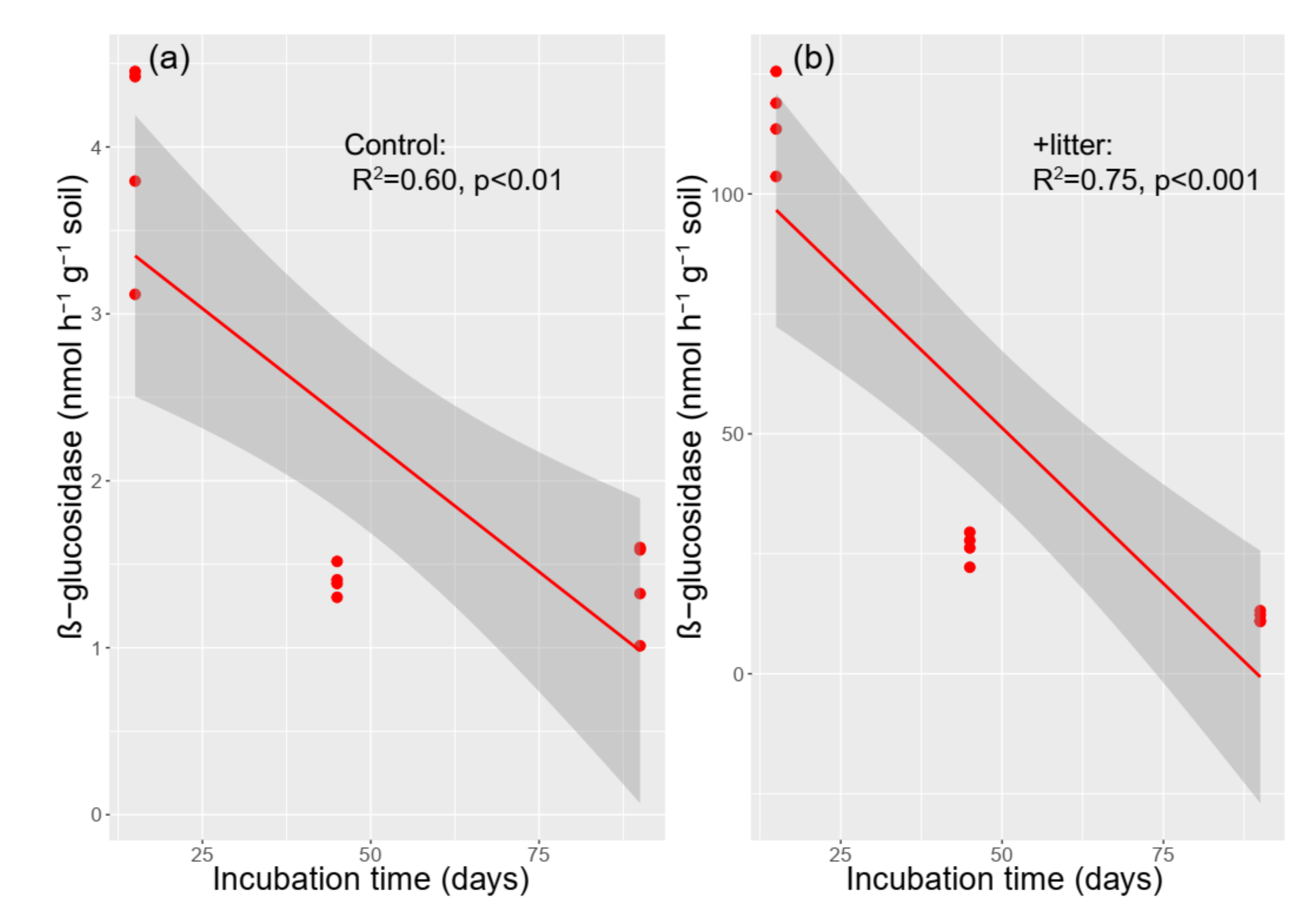
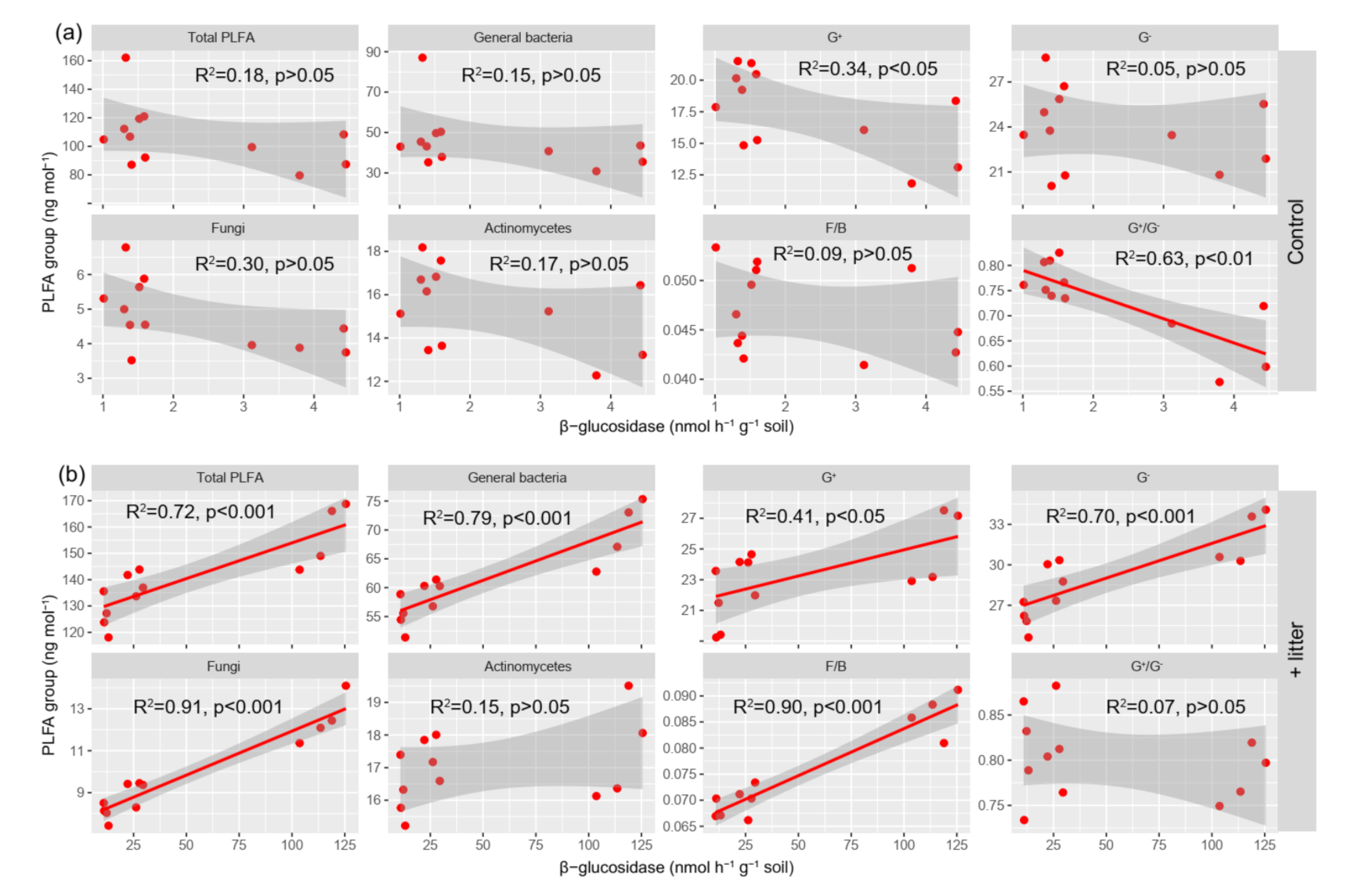
3.4. The Activity of β-glucosidase
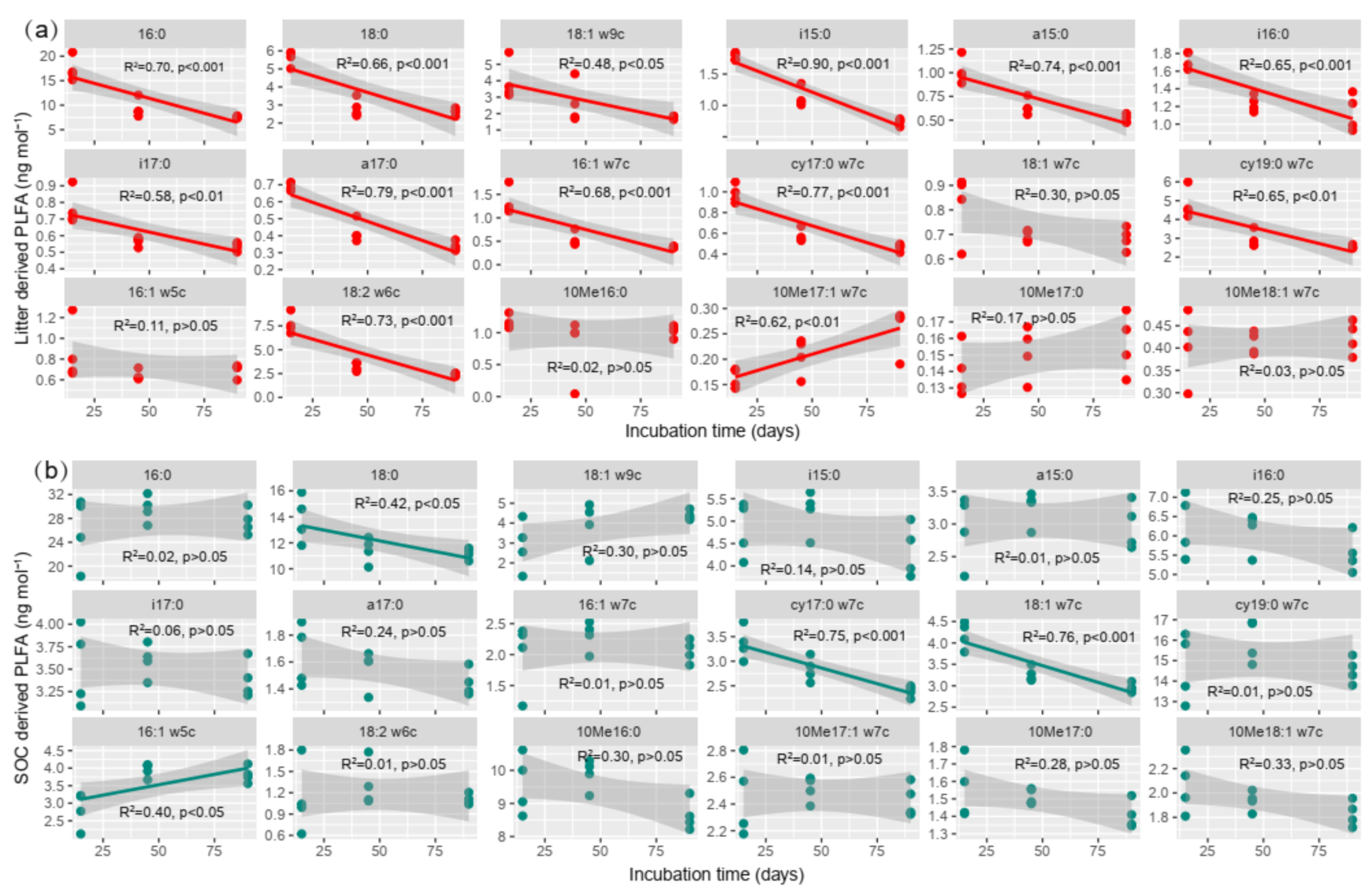
3.5. Microbial Communities Are Involved in Tea Litter Degradation
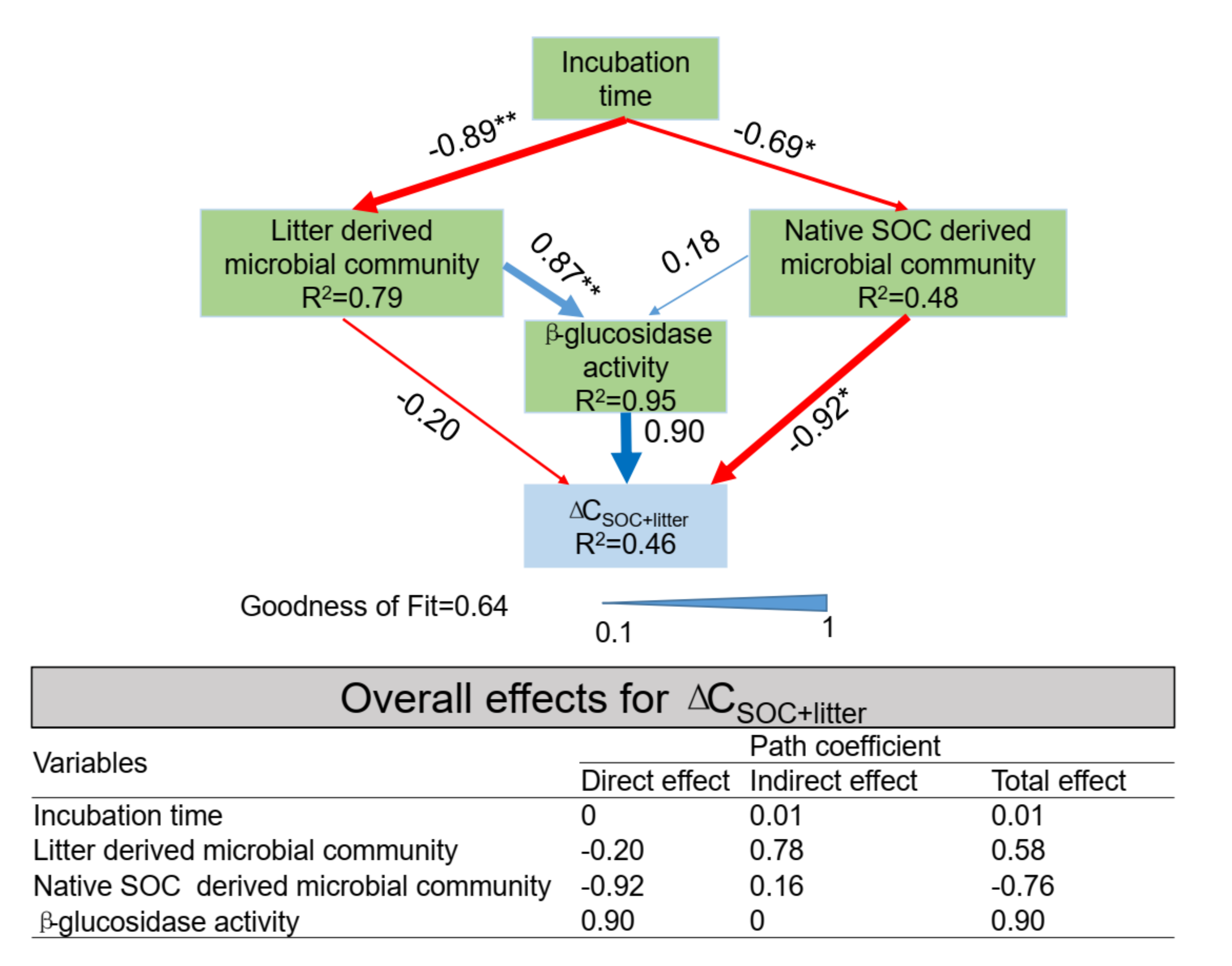
3.6. PLS-PM Analysis
4. Discussion
4.1. Effect of Litter Incorporation on CO2 Emission
4.2. Effect of Litter Incorporation on Soil Microbial Characteristics
4.3. The Relationship between Microbial Communities and SOC Accumulation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shehzadi, S.; Shah, Z.; Mohammad, W. Impact of organic amendments on soil carbon sequestration, water use efficiency and yield of irrigated wheat. Base 2017, 21, 36–49. [Google Scholar]
- Schlesinger, W.H.; Lichter, J. Limited carbon storage in soil and litter of experimental forest plots under increased atmospheric CO2. Nature 2001, 411, 466–469. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.J.; Li, A.Y.; Yang, Y.F.; Li, G.H.; Zhang, F. Soil organic carbon stability under natural and anthropogenic-induced perturbations. Earth Sci. Rev. 2020, 205, 103199. [Google Scholar] [CrossRef]
- Liu, X.J.A.; Finley, B.K.; Mau, R.L.; Schwartz, E.; Dijkstra, P.; Bowker, M.A.; Hungate, B.A. The soil priming effect: Consistent across ecosystems, elusive mechanisms. Soil Biol. Biochem. 2020, 140, 107617. [Google Scholar] [CrossRef]
- Hollis, J.M.; Hannam, J.; Bellamy, P.H. Empirically-derived pedotransfer functions for predicting bulk density in European soils. Eur. J. Soil Sci. 2012, 63, 96–109. [Google Scholar] [CrossRef]
- Lehmann, J.; Kleber, M. The contentious nature of soil organic matter. Nature 2015, 528, 60–68. [Google Scholar] [CrossRef]
- Liu, E.K.; Yan, C.R.; Mei, X.R.; He, W.Q.; Bing, S.H.; Ding, L.P.; Liu, Q.; Liu, S.; Fan, T.L. Long-term effect of chemical fertilizer, straw, and manure on soil chemical and biological properties in northwest China. Geoderma 2010, 158, 173–180. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Friedel, J.K.; Stahr, K. Review of mechanisms and quantification of priming effects. Soil Biol. Biochem. 2000, 32, 1485–1498. [Google Scholar] [CrossRef]
- Fontaine, S.; Mariotti, A.; Abbadie, L. The priming effect of organic matter: A question of microbial competition? Soil Biol. Biochem. 2003, 35, 837–843. [Google Scholar] [CrossRef]
- Troy, S.M.; Lawlor, P.G.; O’Flynn, C.J.; Healy, M.G. Impact of biochar addition to soil on greenhouse gas emissions following pig manure application. Soil Biol. Biochem. 2013, 60, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Kuzyakov, Y. Priming effects: Interactions between living and dead organic matter. Soil Biol. Biochem. 2010, 42, 1363–1371. [Google Scholar] [CrossRef]
- Nottingham, A.T.; Griffiths, H.; Chamberlain, P.M.; Stott, A.W.; Tanner, E.V.J. Soil priming by sugar and leaf-litter substrates: A link to microbial groups. Appl. Soil Ecol. 2009, 42, 183–190. [Google Scholar] [CrossRef]
- Yan, J.F.; Wang, L.; Hu, Y.; Tsang, Y.F.; Zhang, Y.N.; Wu, J.H.; Fu, X.H.; Sun, Y. Plant litter composition selects different soil microbial structures and in turn drives different litter decomposition pattern and soil carbon sequestration capability. Geoderma 2018, 319, 194–203. [Google Scholar] [CrossRef]
- Malik, A.A.; Chowdhury, S.; Schlager, V.; Oliver, A.; Puissant, J.; Vazquez, P.G.M.; Jehmlich, N.; Bergen, M.V.; Griffiths, R.I.; Gleixner, G. Soil fungal: Bacterial ratios are linked to altered carbon cycling. Front. Microbiol. 2016, 7, 1247. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.; Senbayram, M.; Blagodatsky, S.; Myachina, O.; Ditter, T.K.; Lin, X.; Blagodatskaya, E.; Kuzyakov, Y. Soil C and N availability determine the priming effect: Microbial N mining and stoichiometric decomposition theories. Global Change Biol. 2014, 20, 2356–2367. [Google Scholar] [CrossRef]
- Pascault, N.; Ranjard, L.; Kaisermann, A.; Bachar, D.; Christen Richard Terrat, S.; Mathieu, O.; Lévêque, J.; Mougel, C.; Henault, C.; Lemanceau, P.; et al. Stimulation of different functional groups of bacteria by various plant residues as a driver of soil priming effect. Ecosystems 2013, 5, 810–822. [Google Scholar] [CrossRef]
- Yang, X.D.; Ma, L.F.; Ji, L.F.; Shi, Y.Z.; Yi, X.Y.; Yang, Q.L.; Ni, K.; Ruan, J.Y. Long-term nitrogen fertilization indirectly affects soil fungi community structure by changing soil and pruned litter in a subtropical tea (Camellia sinensis L.) plantation in China. Plant Soil 2019, 444, 409–426. [Google Scholar] [CrossRef]
- Li, L.J.; Zhu-Barker, X.; Ye, R.; Doane, T.A.; Horwath, W.R. Soil microbial biomass size and soil carbon influence the priming effect from carbon inputs depending on nitrogen availability. Soil Biol. Biochem. 2018, 119, 41–49. [Google Scholar] [CrossRef]
- Saiya-Cork, K.R.; Sinsabaugh, R.L.; Zak, D.R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- DeForest, J.L. The influence of time, storage temperature, and substrate age on potential soil enzyme activity in acidic forest soils using MUB-linked substrates and L-DOPA. Soil Biol. Biochem. 2009, 41, 1180–1186. [Google Scholar] [CrossRef]
- Ge, G.F.; Li, Z.J.; Fan, F.L.; Chu, G.X.; Hou, Z.N.; Liang, Y.C. Soil biological activity and their seasonal variations in response to long-term application of organic and inorganic fertilizers. Plant Soil 2010, 326, 31–44. [Google Scholar] [CrossRef]
- Thornton, B.; Zhang, Z.L.; Mayes, R.W.; Högberg, M.N.; Midwood, A.J. Can gas chromatography combustion isotope ratio mass spectrometry be used to quantify organic compound abundance? Rapid Commun. Mass Spectrom. 2011, 25, 2433–2438. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, G. PLS Path Modeling with R; Trowchez Editions: Berkeley, CA, USA, 2013. [Google Scholar]
- Fang, X.; Zhao, L.; Zhou, G.Y.; Huang, W.J.; Liu, J.X. Increased litter input increases litter decomposition and soil respiration but has minor effects on soil organic carbon in subtropical forests. Plant Soil 2015, 392, 139–153. [Google Scholar] [CrossRef]
- Prévost-Bouré, N.C.; Soudani, K.; Damesin, C.; Berveiller, D.; Lata, J.C.; Dufrêne, E. Increase in aboveground fresh litter quantity over-stimulates soil respiration in a temperate deciduous forest. Appl. Soil Ecol. 2010, 46, 26–34. [Google Scholar] [CrossRef]
- Xu, S.; Liu, L.L.; Sayer, E. Variability of above-ground litter inputs alters soil physicochemical and biological processes: A meta-analysis of litterfall-manipulation experiments. Biogeosciences 2013, 10, 7423–7433. [Google Scholar] [CrossRef] [Green Version]
- Blagodatskaya, E.; Khomyakov, N.; Myachina, O.; Bogomolova, I.; Blagodatsky, S.; Kuzyakov, Y. Microbial interactions affect sources of priming induced by cellulose. Soil Biol. Biochem. 2014, 74, 39–49. [Google Scholar] [CrossRef]
- Fontaine, S.; Bardoux, G.; Abbadie, L.; Mariotti, A. Carbon input to soil may decrease soil carbon content. Ecol. Lett. 2004, 7, 314–320. [Google Scholar] [CrossRef]
- Shahzad, T.; Chenu, C.; Repinçay, C.; Mougin, C.; Ollier, J.L.; Fontaine, S. Plant clipping decelerates the mineralization of recalcitrant soil organic matter under multiple grassland species. Soil Biol. Biochem. 2012, 51, 73–80. [Google Scholar] [CrossRef]
- Tian, J.; Pausch, J.; Yu, G.R.; Blagodatskaya, E.; Kuzyakov, Y. Aggregate size and glucose level affect priming sources: A three source-partitioning study. Soil Biol. Biochem. 2016, 97, 199–210. [Google Scholar] [CrossRef]
- Bell, J.M.; Smith, J.L.; Bailey, V.L.; Bolton, H. Priming effect and C storage in semi-arid no-till spring crop rotations. Biol. Fertil. Soils 2003, 37, 237–244. [Google Scholar] [CrossRef]
- Lu, W.W.; Ding, W.X.; Zhang, J.H.; Li, Y.; Luo, J.F.; Bolan, N.; Xie, Z.B. Biochar suppressed the decomposition of organic carbon in a cultivated sandy loam soil: A negative priming effect. Soil Biol. Biochem. 2014, 76, 12–21. [Google Scholar] [CrossRef]
- Maestrini, B.; Herrmann, A.M.; Nannipieri, P.; Schmidt, M.W.; Abiven, S. Ryegrass-derived pyrogenic organic matter changes organic carbon and nitrogen mineralization in a temperate forest soil. Soil Biol. Biochem. 2014, 69, 291–301. [Google Scholar] [CrossRef]
- Luo, Z.K.; Wang, E.L.; Sun, O.J. A meta-analysis of the temporal dynamics of priming soil carbon decomposition by fresh carbon inputs across ecosystems. Soil Biol. Biochem. 2016, 101, 96–103. [Google Scholar] [CrossRef]
- Zimmerman, A.R.; Gao, B.; Ahn, M.Y. Positive and negative carbon mineralization priming effects among a variety of biochar-amended soils. Soil Biol. Biochem. 2011, 43, 1169–1179. [Google Scholar] [CrossRef]
- Maestrini, B.; Nannipieri, P.; Abiven, S. A meta-analysis on pyrogenic organic matter induced priming effect. GCB Bioenergy 2015, 7, 577–590. [Google Scholar] [CrossRef]
- Smith, J.L.; Collins, H.P.; Bailey, V.L. The effect of young biochar on soil respiration. Soil Biol. Biochem. 2010, 42, 2345–2347. [Google Scholar] [CrossRef]
- Hamer, U.; Marschner, B. Priming effects in soils after combined and repeated substrate additions. Geoderma 2005, 128, 38–51. [Google Scholar] [CrossRef]
- Adamczyk, M.; Perez-Mon, C.; Gunz, S.; Frey, B. Strong shifts in microbial community structure are associated with increased litter input rather than temperature in high Arctic soils. Soil Biol. Biochem. 2020, 151, 108054. [Google Scholar] [CrossRef]
- Che, R.; Liu, D.; Qin, J.; Wang, F.; Wang, W.; Xu, Z.; Cui, X. Increased litter input significantly changed the total and active microbial communities in degraded grassland soils. J. Soils Sediments 2020, 20, 2804–2816. [Google Scholar] [CrossRef]
- Bradford, M.A.; Keiser, A.D.; Davies, C.A.; Mersmann, C.A.; Strickland, M.S. Empirical evidence that soil carbon formation from plant inputs is positively related to microbial growth. Biogeochemistry 2013, 113, 271–281. [Google Scholar] [CrossRef]
- Wang, J.; Chapman, S.J.; Yao, H.Y. Incorporation of 13C-labelled rice rhizodeposition into soil microbial communities under different fertilizer applications. Appl. Soil Ecol. 2016, 101, 11–19. [Google Scholar] [CrossRef]
- Colin, Y.; Goberna, M.; Verdú, M.; Navarro-Cano, J.A. Successional trajectories of soil bacterial communities in mine tailings: The role of plant functional traits. J. Environ. Manage. 2019, 241, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Wadman, W.P.; De Haan, S. Decomposition of organic matter from 36 soils in a long-term pot experiment. Plant Soil 1997, 189, 289–301. [Google Scholar] [CrossRef]
- Kéraval, B.; Lehours, A.C.; Colombet, J.; Amblard, C.; Alvarez, G.; Fontaine, S. Soil carbon dioxide emissions controlled by an extracellular oxidative metabolism identifiable by its isotope signature. Biogeosciences 2016, 13, 6353–6362. [Google Scholar] [CrossRef] [Green Version]
- Maire, V.; Alvarez, G.; Colombet, J.; Comby, A.; Despinasse, R.; Dubreucq, E.; Fontaine, S. An unknown oxidative metabolism substantially contributes to soil CO2 emissions. Biogeosciences 2013, 10, 1155–1167. [Google Scholar] [CrossRef] [Green Version]
- Whitman, T.; DeCiucies, S.; Hanley, K.; Enders, A.; Woolet, J.; Lehmann, J. Microbial community shifts reflect losses of native soil carbon with pyrogenic and fresh organic matter additions and are greatest in low-carbon soils. Appl. Environ. Microbiol. 2021, 87, e02555-20. [Google Scholar] [CrossRef]
- Riggs, C.E.; Hobbie, S.E.; Bach, E.M.; Hofmockel, K.S.; Kazanski, C.E. Nitrogen addition changes grassland soil organic matter decomposition. Biogeochemistry 2015, 125, 203–219. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Global Change Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef] [Green Version]
- Schimel, J.P.; Weintraub, M.N. The implications of exoenzyme activity on microbial carbon and nitrogen limitation in soil: A theoretical model. Soil Biol. Biochem. 2003, 35, 549–563. [Google Scholar] [CrossRef]
- Liu, W.X.; Qiao, C.L.; Yang, S.; Bai, W.M.; Liu, L.L. Microbial carbon use efficiency and priming effect regulate soil carbon storage under nitrogen deposition by slowing soil organic matter decomposition. Geoderma 2018, 332, 37–44. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Treatment | Day 0 | Day 15 | Day 45 | Day 90 |
|---|---|---|---|---|---|
| SOC (mg g−1) | Control | 43.85 (0.38) a | 36.57 (0.62) b | 32.19 (1.29) c | 34.19 (0.80) bc |
| +litter | 44.97 (0.13) a | 36.48 (2.89) b | 37.06 (2.23) b | 37.57 (2.03) b | |
| TN (mg g−1) | Control | 4.30 (0.04) a | 3.54 (0.06) b | 3.16 (0.10) b | 3.33 (0.09) bc |
| +litter | 4.05 (0.03) a | 3.53 (0.23) ab | 3.48 (0.17) ab | 3.44 (0.16) b | |
| SOC/TN | Control | 10.21 (0.05) ns | 10.32 (0.01) | 10.19 (0.09) | 10.27 (0.05) |
| +litter | 11.11 (0.07) a | 10.31 (0.18) c | 10.65 (0.12) bc | 10.90 (0.13) ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Ni, K.; Ma, L.; Shi, Y.; Yi, X.; Ji, L.; Ruan, J. 13C Labelling of Litter Added to Tea (Camellia sinensis L.) Plantation Soil Reveals a Significant Positive Priming Effect That Leads to Less Soil Organic Carbon Accumulation. Agronomy 2022, 12, 293. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020293
Yang X, Ni K, Ma L, Shi Y, Yi X, Ji L, Ruan J. 13C Labelling of Litter Added to Tea (Camellia sinensis L.) Plantation Soil Reveals a Significant Positive Priming Effect That Leads to Less Soil Organic Carbon Accumulation. Agronomy. 2022; 12(2):293. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020293
Chicago/Turabian StyleYang, Xiangde, Kang Ni, Lifeng Ma, Yuanzhi Shi, Xiaoyun Yi, Lingfei Ji, and Jianyun Ruan. 2022. "13C Labelling of Litter Added to Tea (Camellia sinensis L.) Plantation Soil Reveals a Significant Positive Priming Effect That Leads to Less Soil Organic Carbon Accumulation" Agronomy 12, no. 2: 293. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020293