
Three-Dimensional Reconstruction and Predictive Simulation Algorithm of Forest and Fruit Wood Borer Galleries Based on Two-Dimensional Images and Different Influencing Factors


Abstract
:1. Introduction
2. Data Collection and Preprocessing
2.1. Test Data Acquisition
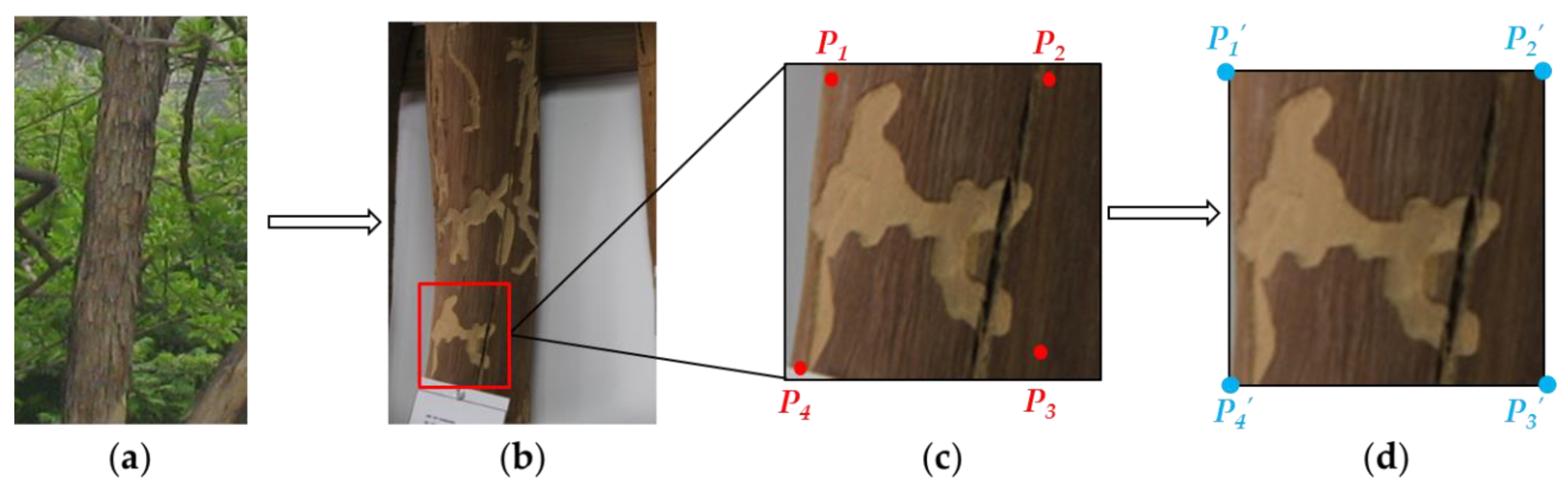
2.2. Image Correction
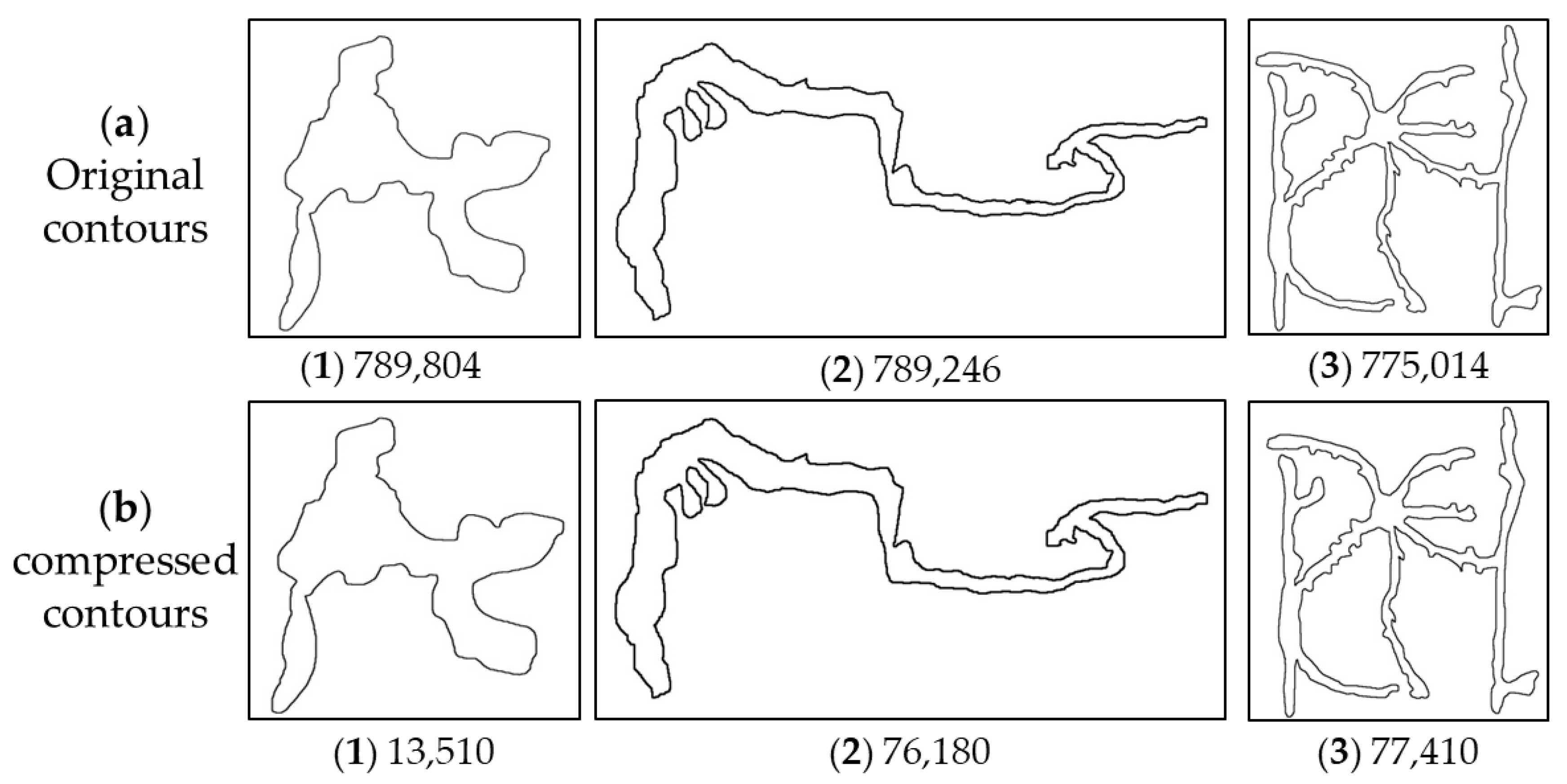
2.3. Contours Extraction and Compression
2.4. Three-Dimensional Models Reconstruction
3. Based on Influencing Factors for Predicting Forest and Fruit Wood Borer Galleries Spread Range
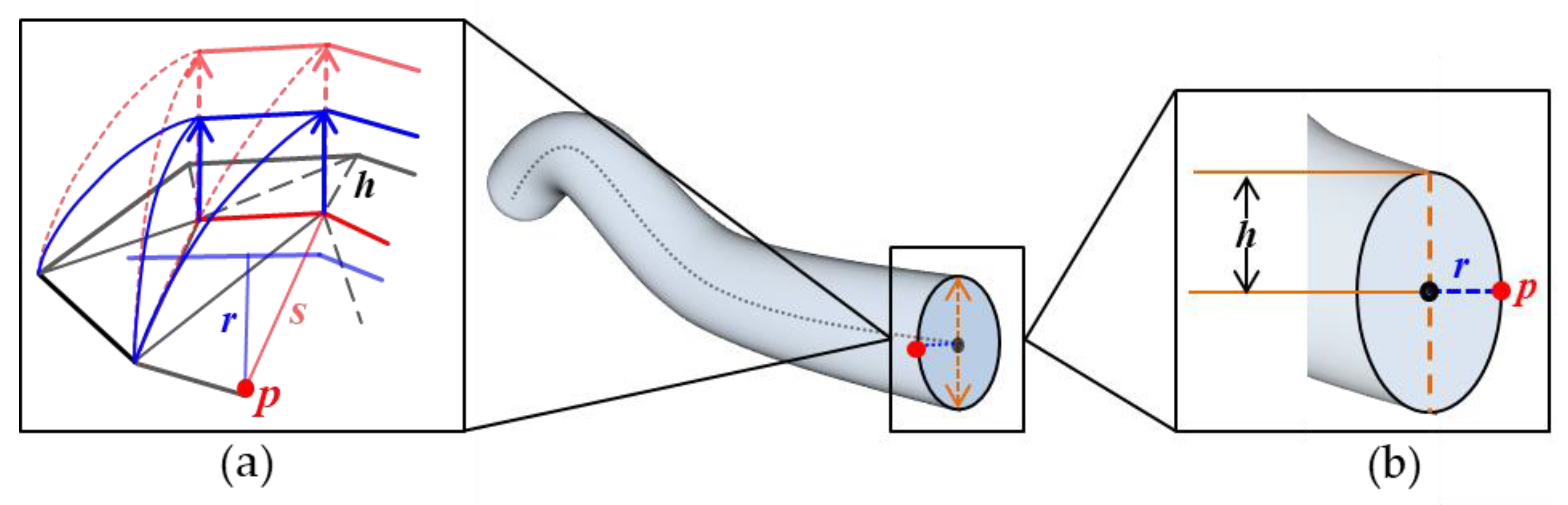
3.1. Three-Dimensional Skeleton
3.1.1. Three-Dimensional Skeleton Extraction
3.1.2. 3D Skeleton Description
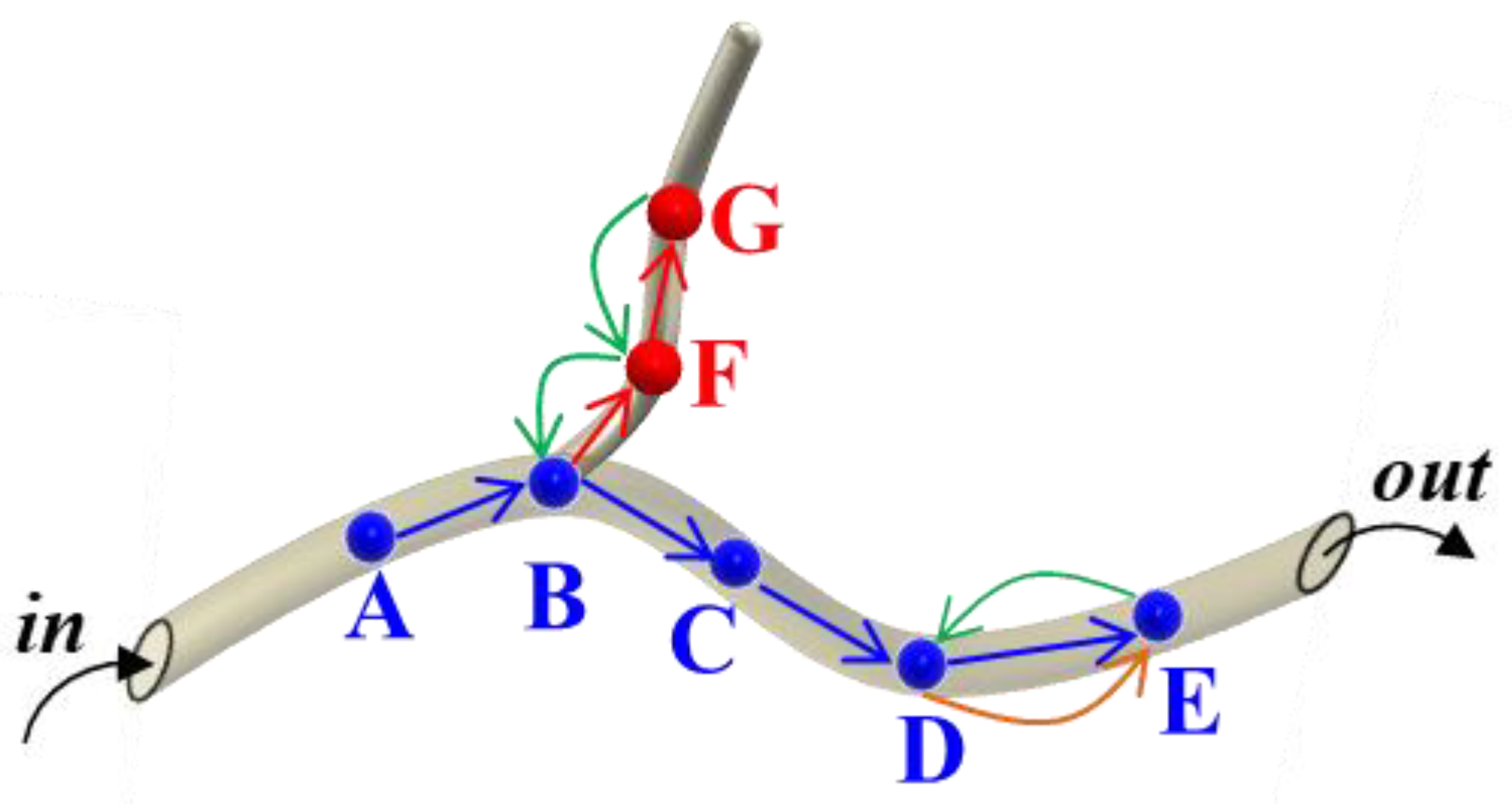
3.1.3. Description of Pest Movement
- Pioneering Movement
- 2.
- Forward Movement
- 3.
- Backward Movement
3.2. Influence Factors’ Establishment
3.2.1. Forest and Fruit Wood Borer Species
3.2.2. Host Tree Species
3.2.3. Density of Insect Population
3.2.4. Other Effects
3.3. Prediction of Forest and Fruit Wood Borer Galleries
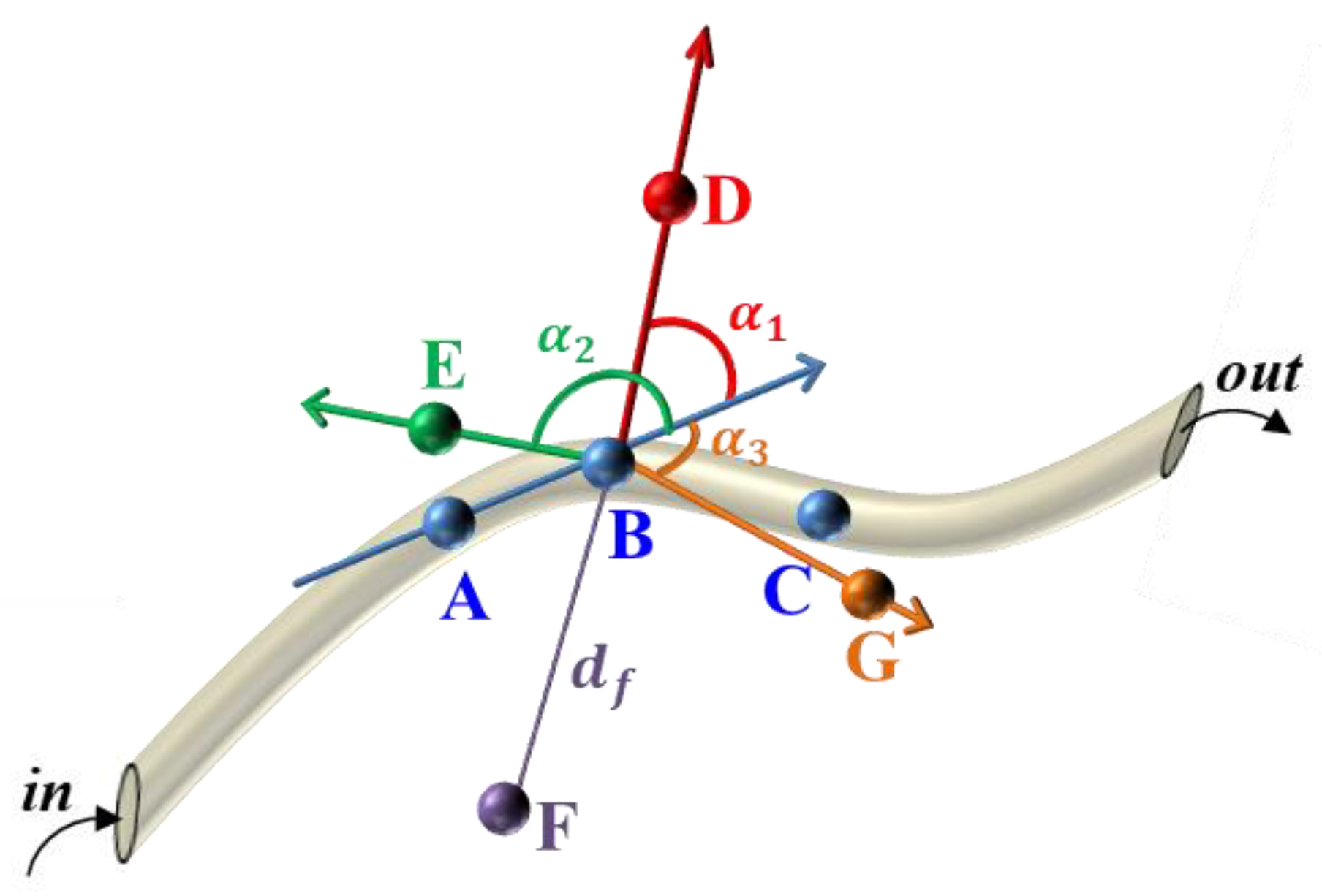
3.3.1. Define the Parameters
- The distance an insect moves per unit of time.
- 2.
- The angle of insect movement
- 3.
- The distance of the sample point
- 4.
- The angle of the sample point
3.3.2. Define the Prediction Point
- Pioneering Points
- 2.
- Forward Points
- 3.
- Backward Points
- 4.
- Wrong Points
3.3.3. Prediction Simulation Algorithm
| Algorithm 1. Simulation Algorithm for Prediction of Forest and Fruit Wood Borer Pest Galleries |
| Input: The three-dimensional skeleton points set , the set of edges between the points of the three-dimensional skeleton , the set of attributes of the points , the set of weights of edges , the defined movement distance and angle , and the set of possible points . |
| Output: The new 3D skeleton map obtained by the algorithm. |
| 1: for i in range (0, len()-1) do |
| 2: for j in range (0, len()-1) do 3: judge the distance between and 4: if , i = i + 1, j = 0 5: else judge the angle between and 6: if , insert into , update and |
| 7: else if , update according , update and 8: else update according to , update and 9: i = 0, j = 0 10: end if 11: end if 12: end for 13: return the new skeleton map |
4. Result and Analysis
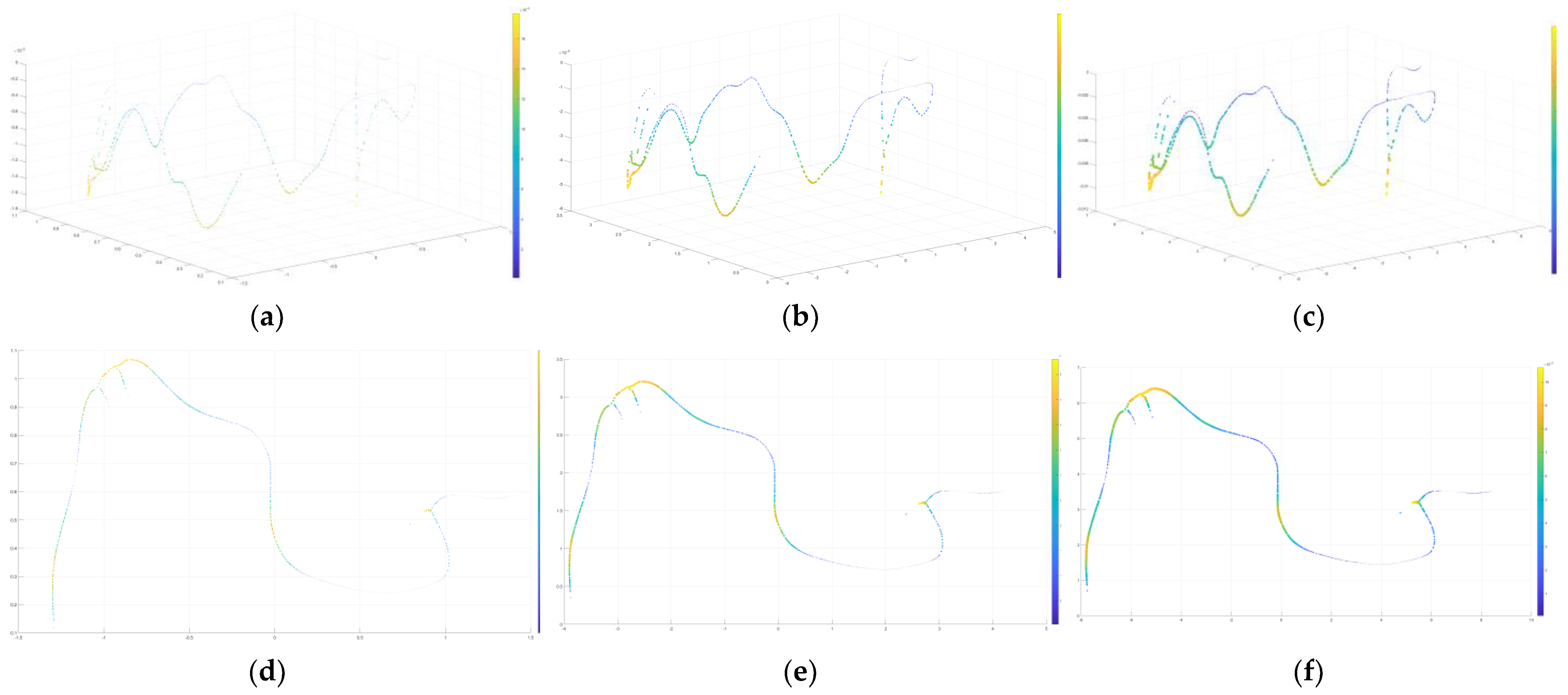
4.1. 3D Reconstruction
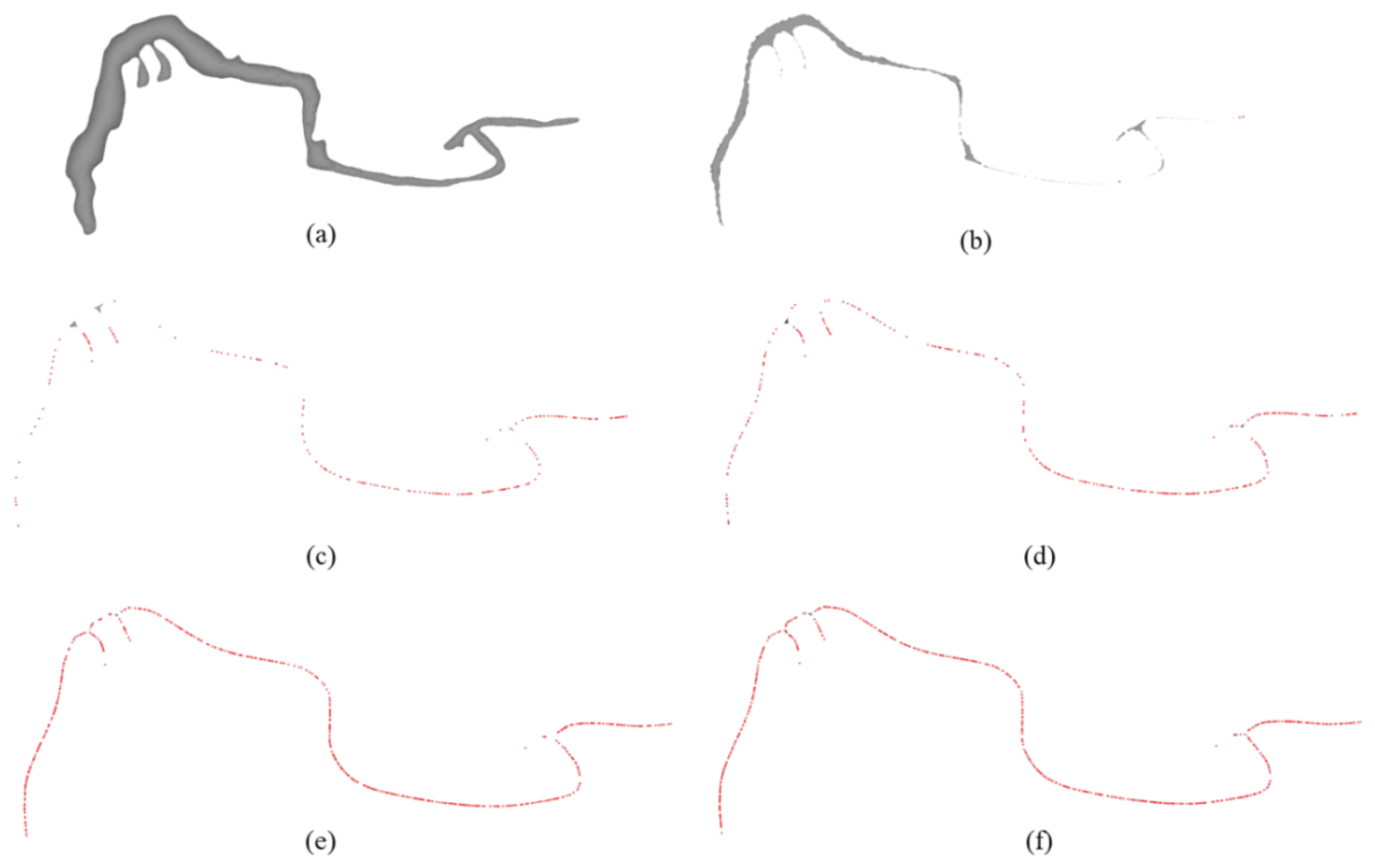
4.1.1. 3D Modeling Results of Wood Borer Galleries
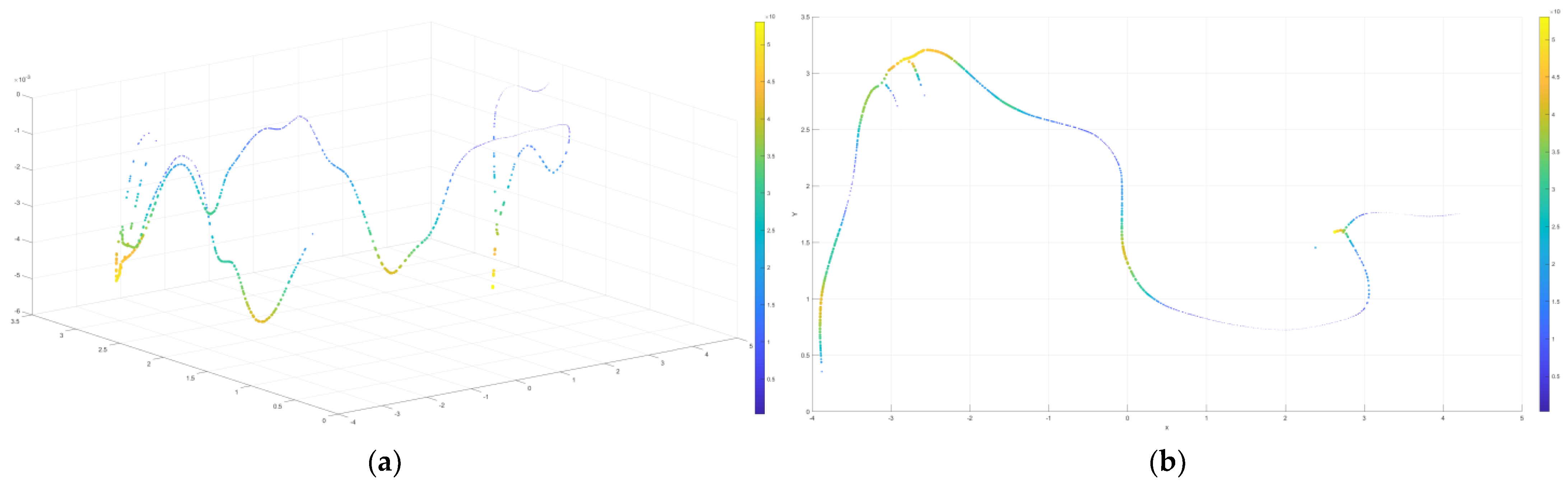
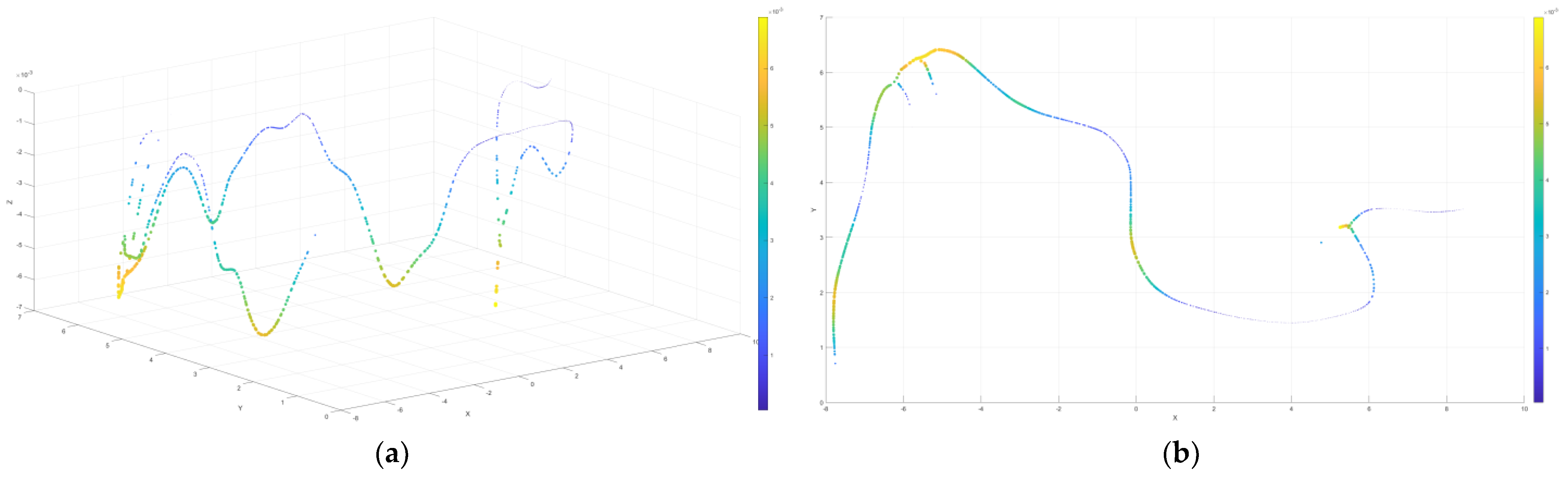
4.1.2. Visualization Results
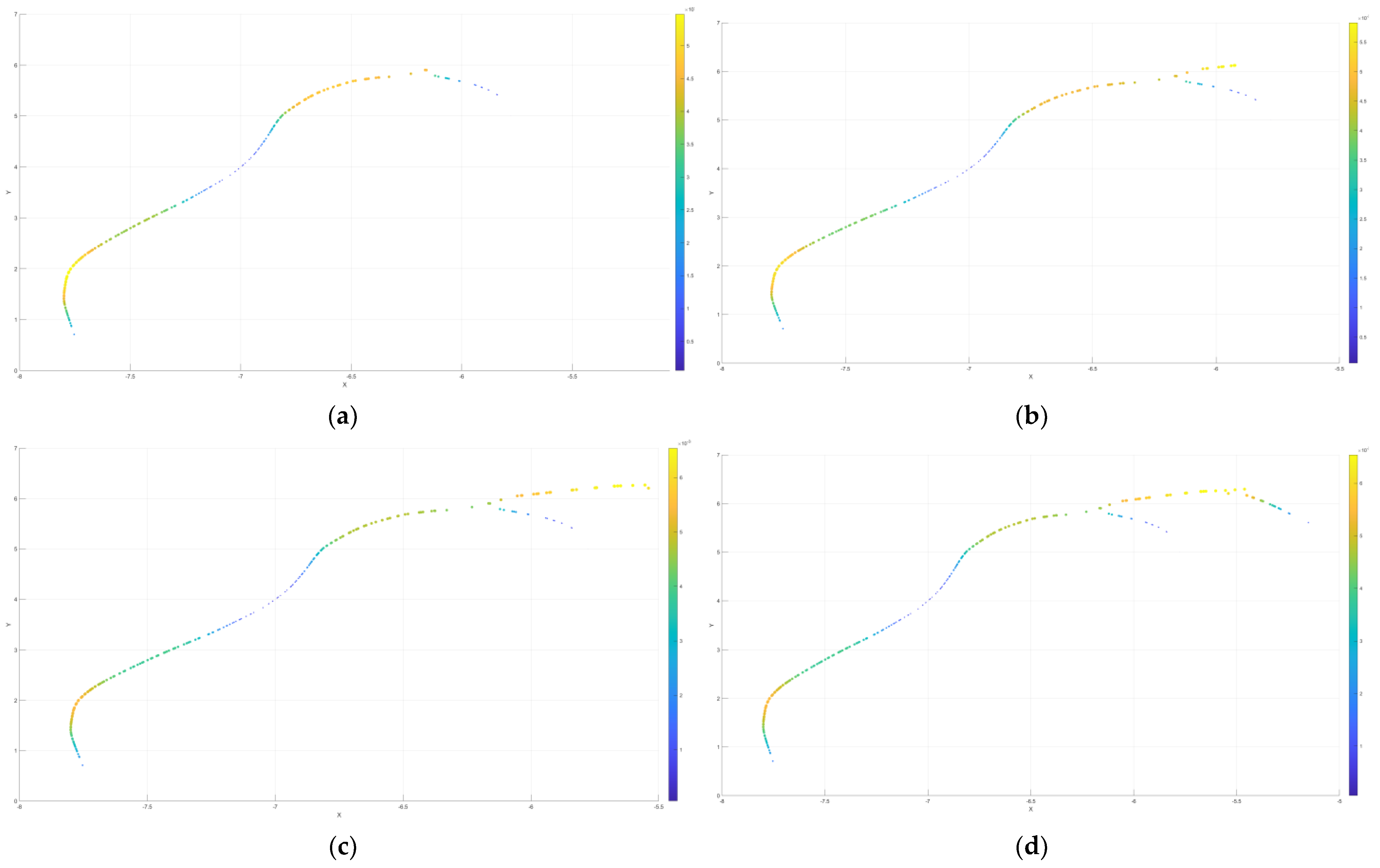
4.2. Predictive Simulation
4.2.1. Only Forward Points or Backward Points Exist
4.2.2. Only Pioneering Points Exist
4.2.3. Factors Influencing Regulation
5. Discussion
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, J.; Chang, G.B.; Qv, T.; Cui, Y.S.; Yan, J.; Song, Y.S. Hazard Assessment of Forester Pests in China. For. Pest Dis. 2019, 38, 11–17. [Google Scholar]
- Zhu, S.J. Study on the Occurrence Characteristics of Major Pests of Forestry and Fruit Trees under Climate Change Conditions in Xinjiang Region. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2013; p. 6. [Google Scholar]
- Guo, W.C.; Zhang, X.L.; Wu, W.; Zhang, W.; Fu, K.Y.; Tursun, A.; Ding, X.H.; Yimit, R. Current situation, trends and research progress of invasive alien species in agriculture and foresty in oases in Xinjiang, China. J. Biosaf. 2017, 26, 1–11. [Google Scholar]
- Zhou, M.J.; Nie, X.B.; Du, W.S.; Zhao, Y.X.; Li, J. Relationship between Sub-healthy Forest and Trunkborer-insect. J. Northeast. For. Univ. 2009, 37, 49–51. [Google Scholar]
- Yang, L.Y.; Liu, R.J.; Zhao, X.F.; Huang, S.; Hua, J.; Sun, S.H. Vertical Distribution of Wood-boring Pests and Its Parasitic Wasp in Pinus tabulaeformis. Chin. J. Biol. Control. 2021, 34, 701–708. [Google Scholar]
- Zhan, M.K. Studies on Population Dynamics of Woodborers and the Relationship with Pine Wilt Disease on Pinus massoniana, and Natural Enemies of Monochamus alternatus; Chinese Academy of Forestry: Beijing, China, 2014. [Google Scholar]
- Flower, C.E.; Knight, K.S.; Gonzalez-Meler, M.A. Using stable isotopes as a tool to investigate the impacts of EAB on tree physiology and EAB spread. In Proceedings of the Emerald Ash Borer Research and Technology Development Meeting, Pittsburgh, PA, USA, 21–22 October 2009. [Google Scholar]
- Flower, C.E.; Knight, K.S.; Rebbeck, J.; Gonzalez-Meler, M.A. The Relationship between the Emerald Ash Borer (Agrilus Planipennis) and Ash (Fraxinus spp.) Tree Decline: Using Visual Canopy Condition Assessments and Leaf Isotope Measurements to Assess Pest Damage. For. Ecol. Manag. 2013, 303, 143–147. [Google Scholar] [CrossRef]
- Crook, D.J.; Mastro, V.C. Chemical Ecology of the Emerald Ash Borer Agrilus planipennis. J. Chem. Ecol. 2010, 36, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Mccullough, D.G.; Siegert, N.W.; Poland, T.M.; Pierce, S.J.; Ahn, S.Z. Effects of Trap Type, Placement and Ash Distribution on Emerald Ash Borer Captures in a Low Density Site. Environ. Entomol. 2011, 40, 1239–1252. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Li, J.; Wang, J.; Hall, B. The Early Detection of the Emerald Ash Borer (EAB) Using Advanced Geospacial Technologies. Int. Arch. Photogramm. Remote Sens. Spatial Inf. Sci. 2014, XL-2, 213–219. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.W.; Hu, B.X.; Robinson, J. Early Detection of Emerald Ash Borer Infestation Using Multisourced Data: A Case Study in the Town of Oakville, Ontario, Canada. J. Appl. Remote Sens. 2014, 8, 083602. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhao, Z.G.; Wang, T.T.; Wu, W.Q.; Wang, X.; Ma, R.Y. Application of Sonic Signals for Fruit Damage Detection Produced by Grapholitha molesta Larval Feeding. J. Shanxi Agric. Univ. (Nat. Sci. Ed.) 2018, 38, 13. [Google Scholar]
- Lou, D.F.; Xu, X.F.; Li, J.; Zhou, H.S.; Liu, X.J.; Li, Q.F.; Jiao, Y.; Xu, L.; Chen, D.M.; Yu, D.J.; et al. Acoustic Characteristics and Their Comparison of Six Species of Wood Borers. Plant Quar. 2013, 27, 6–10. [Google Scholar]
- Al-Manie, M.A.; Alkanhal, M.I. Acoustic Detection of the Red Date Palm Weevil. Int. J. Electron. Commun. Eng. 2007, 1, 4. [Google Scholar]
- Mankin, R.W.; Brandhorst-Hubbard, J.; Flanders, K.L.; Zhang, M.; Crocker, R.L.; Lapointe, S.L.; McCoy, C.W.; Fisher, J.R.; Weaver, D.K. Eavesdropping on Insects Hidden in Soil and Interior Structures of Plants. Ecol. Behav. 2000, 93, 1173–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mankin, R.W.; Hubbard, J.L.; Flanders, K.L. Acoustic Indicators for Mapping Infestation Probabilities of Soil Invertebrates. J. Econ. Entomol. 2007, 100, 11. [Google Scholar] [CrossRef]
- Mankin, R.W.; Mizrach, A.; Hetzroni, A.; Levsky, S.; Nakache, Y.; Soroker, V. Temporal and Spectral Features of Sounds of Wood-Boring Beetle Larvae: Identifiable Patterns of Activity Enable Improved Discrimination from Background Noise. Fla. Entomol. 2008, 91, 241–248. [Google Scholar] [CrossRef]
- Mankin, R.W.; Smith, M.T.; Tropp, J.M.; Atkinson, E.B.; Jong, D.Y. Detection of Anoplophora glabripennis (Coleoptera: Cerambycidae) Larvae in Different Host Trees and Tissues by Automated Analyses of Sound-Impulse Frequency and Temporal Patterns. J. Econ. Entomol. 2008, 101, 12. [Google Scholar] [CrossRef]
- Mankin, R.W.; Burman, H.; Menocal, O.; Carrillo, D. Acoustic Detection of Mallodon dasystomus (Coleoptera: Cerambycidae) in Persea Americana (Laurales: Lauraceae) Branch Stumps. Fla. Entomol. 2018, 101, 321–323. [Google Scholar] [CrossRef] [Green Version]
- Li, W.J.; Ren, L.L.; Li, C.; Luo, Y.Q. Detailed observations on bark damage caused by the grey tiger long-horned beetle (Xylotrechus rusticus L.). Chin. J. Appl. Entomol. 2013, 50, 1270–1273. [Google Scholar]
- Zhang, Y.K.; Long, J.M.; Zhao, C.P.; Ma, Z.L.; Zhu, G.Y.; Zhang, Z.B. Study on spatial distribution patterns of the eggs of Batocera rufomaculata and sampling techniques. China Plant Prot. 2019, 39, 25–30. [Google Scholar]
- Liu, A.Q. Studies on Larval Tunnels and Adult Antennal Sensilla of Xylotrechus Quadripes Chevrolat. Master’s Thesis, Yunnan University, Kunming, China, 2020; p. 4. [Google Scholar]
- Castel, T.; Beaudoin, A.; Floury, N.; Le Toan, T.; Caraglio, Y.; Barczi, J.F. Deriving forest canopy parameters for backscatter models using the AMAP architectural plant model. IEEE Trans. Geosci. Remote Sens. 2001, 39, 571–583. [Google Scholar] [CrossRef]
- Ijiri, T.; Owada, S.; Okabe, M.; Igarashi, T. Floral Diagrams and Inflorescences: Interactive Flower Modeling Using Botanical Structural Constraints. ACM Trans. Graph. 2005, 24, 720–726. [Google Scholar] [CrossRef]
- Lintermann, B.; Deussen, O. Interactive Modeling of Plants. IEEE Comput. Grap. Appl. 1999, 19, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Ren, L.L.; Pan, L.; Luo, Y.Q. Clarification of the species name of Agrilus subrobustus Saunders, a description of its morphological characteristics and a three-dimensional reconstruction of the larval galleries of this pest. Chin. J. Appl. Entomol. 2016, 53, 864–873. [Google Scholar]
- Hen, N.N. 3D Reconstruction Algorithm for Medical Images. Master’s Thesis, Henan University of Technology, Zhengzhou, China, 2018; p. 5. [Google Scholar]
- Gao, F.; Fu, Z.L. Fast marching cubes algorithm for 3D reconstruction for medical images. J. Comput. Appl. 2013, 33, 201–203, 213. [Google Scholar]
- Pi, Y.F. Development of Computational Human Phantoms and Applications to Automated CT Image Segmentation. Ph.D. Thesis, University of Science and Technology of China, Hefei, China, 2018; p. 5. [Google Scholar]
- Irnstorfer, N.; Unger, E.; Hojreh, A.; Homolka, P. An anthropomorphic phantom representing a prematurely born neonate for digital x-ray imaging using 3D printing: Proof of concept and comparison of image quality from different systems. Sci. Rep. 2019, 9, 14375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, K.B. Soft Tissue Modeling Based on Medical Images and Its Application in Virtual Surgery. Master’s Thesis, Nanchang University, Nanchang, China, 2019; p. 5. [Google Scholar]
- Zou, H.; Zhu, C.Z.; Wang, C.; Tian, G.Y.; Zhang, H.F.; Sun, C.D. Application of three-dimensional reconstruction combined with augmented reality technology in precise hepatectomy performed by robot. Chin. J. Robot. Surg. 2020, 1, 141–147. [Google Scholar]
- Ross, R.J.; McDonald, K.A.; Green, D.W.; Schad, K.C. Relationship Between Log and Lumber Modulus of Elasticity. Wood Eng. 1997, 47, 89–92. [Google Scholar]
- Beall, F.C. Overview of the Use of Ultrasonic Technologies in Research on Wood Properties. Wood Sci. Technol. 2002, 36, 197–212. [Google Scholar] [CrossRef]
- Rinn, F. Intact-decay transitions in profiles of density-calibratable resistance drilling devices using long thin needles. Arboric. J. 2016, 38, 204–217. [Google Scholar] [CrossRef]
- Ge, Z.; Chen, L.; Luo, R.; Wang, Y.; Zhou, Y. The Detection of Structure in Wood by X-ray CT Imaging Technique. BioResources 2018, 13, 3674–3685. [Google Scholar] [CrossRef]
- Butnor, J.R.; Pruyn, M.L.; Shaw, D.C.; Harmon, M.E.; Mucciardi, A.N.; Ryan, M.G. Detecting Defects in Conifers with Ground Penetrating Radar: Applications and Challenges. For. Pathol. 2009, 39, 309–322. [Google Scholar] [CrossRef]
- Du, X.; Feng, H.; Hu, M.; Fang, Y.; Chen, S. Three-Dimensional Stress Wave Imaging of Wood Internal Defects Using TKriging Method. Comput. Electron. Agric. 2018, 148, 63–71. [Google Scholar] [CrossRef]
- Ge, Z.D.; Qi, Y.H.; Luo, R.; Chen, L.X.; Wang, Y.W.; Zhou, Y.C. Correction of Rotation Center for the Wood CT Imaging System. Sci. Silvae Sin. 2018, 54, 164–171. [Google Scholar]
- Okabe, M.; Owada, S.; Igarashi, T. Interactive Design of Botanical Trees Using Freehand Sketches and Example-Based Editing. Eurographics 2005, 24, 3. [Google Scholar] [CrossRef]
- Anastacio, F.; Prusinkiewicz, P.; Sousa, M.C. Sketch-Based Parameterization of L-Systems Using Illustration-Inspired Construction Lines and Depth Modulation. Comput. Graph. 2009, 33, 440–451. [Google Scholar] [CrossRef]
- Longay, S.; Runions, A.; Boudon, F.; Prusinkiewicz, P. TreeSketch: Interactive Procedural Modeling of Trees on a Tablet. In Proceedings of the International Symposium on Sketch-Based Interfaces and Modeling, Annecy, France, 4–6 June 2012; pp. 107–120. [Google Scholar] [CrossRef]
- Zhang, H. Three-Dimensional Reconstruction of the Galleries of Several Wood Borer Species. Master’s Thesis, Beijing Forestry University, Beijing, China, 2017; p. 6. [Google Scholar]
- Ehrlich, P.R.; Raven, P.H. Butterflies and Plants: A Study in Coevolution. Evolution 1964, 18, 586–608. [Google Scholar] [CrossRef]
- Gervais, D.J.; Green, D.F.; Work, T.T. Causes of variation in wood-boring beetle damage in fire-killed black spruce (Picea mariana) forests in the central boreal forest of Quebec. Ecoscience 2012, 19, 398–403. [Google Scholar] [CrossRef]
- Thibault, M.; Moreau, G. Enhancing Bark- and Wood-Boring Beetle Colonization and Survival in Vertical Deadwood during Thinning Entries. J. Insect Conserv. 2016, 20, 789–796. [Google Scholar] [CrossRef]
- Yang, X.F. Effect of Host Plant Factors on Occurrence and Growing Development and Oviposition Preference of Grapholita molesta Busck. Master’s Thesis, Hebei Agricultural University, Baoding, China, 2013; p. 6. [Google Scholar]
- Feng, Y. Modeling, Evolution and Application of Big Data of Forest Pest in Changbai Mountains by Means of Network Science. Ph.D. Thesis, Northeastern University, Shenyang, China, 2016; p. 4. [Google Scholar]
- Buma, B.; Pugh, E.T.; Wessman, C.A. Effect of the Current Major Insect Outbreaks on Decadal Phenological and LAI Trends in Southern Rocky Mountain Forests. Int. J. Remote Sens. 2013, 34, 7249–7274. [Google Scholar] [CrossRef]
- Guo, Z.W.; Wu, C.Y.; Wang, X.Y. Forest insect-disease monitoring and estimation based on satellite remote sensing data. Geogr. Res. 2019, 38, 831–843. [Google Scholar]
- Feng, L.; Jia, Z.; Li, Q.; Zhao, A.; Zhao, Y.; Zhang, Z. Spatiotemporal Change of Sparse Vegetation Coverage in Northern China. J. Indian Soc. Remote Sens. 2019, 47, 359–366. [Google Scholar] [CrossRef]
- Cheng, Z.J.; Sun, W.; Gao, Y.B.; He, S.C.; Zhou, J.C. Damage to maize crops in Northeastern China caused by the third generation of the armyworm Mythimna separata (Walker). Chin. J. Appl. Entomol. 2018, 55, 849–856. [Google Scholar]
- Gao, F.L.; Guo, S.M.; Yan, M.; Li, J.M.; Yu, F.H. Effects of simulated insect herbivory on the growth and chemical defense of Alternanthera philoxeroides in different habitats. Acta Ecol. Sin. 2018, 38, 2344–2352. [Google Scholar]
- Cao, Q.J.; Yang, F.T.; Liang, Y.; Jiang, X.L.; Lamine, D.; Li, G. Simulation in Harm Degree of Mythimna seperata in Filling Stage and Its Effect on Maize Yield and Quality. Acta Agric. Boreali-Sin. 2015, 30, 180–185. [Google Scholar]
- Schat, M.; Blossey, B. Influence of Natural and Simulated Leaf Beetle Herbivory on Biomass Allocation and Plant Architecture of Purple Loosestrife (Lythrum salicaria L.). Environ. Entomol. 2005, 34, 906–914. [Google Scholar] [CrossRef] [Green Version]
- Su, X.Y.; Lin, C.S. Computer Simulation of Population Dynamics of the Oriental Armyworm (Mythimna separta). Acta Ecol. Sin. 1986, 6, 65–73. [Google Scholar]
- Su, X.Y.; Lin, C.S. A Dynamic Econmic Threshold (ET) for Controlling the Oriental Armyworm (Mythimna separata W.) on Wheat. Acta Ecol. Sin. 1987, 7, 322–330. [Google Scholar]
- Casas, J.; Aluja, M. The Geometry of Search Movements of Insects in Plant Canopies. Behav. Ecol. 1997, 8, 37–45. [Google Scholar] [CrossRef]
- Hanan, J.; Prusinkiewicz, P.; Zalucki, M.; Skirvin, D. Simulation of Insect Movement with Respect to Plant Architecture and Morphogenesis. Comput. Electron. Agric. 2002, 35, 255–269. [Google Scholar] [CrossRef]
- Wang, M.; Cribb, B.; Clarke, A.R.; Hanan, J. A Generic Individual-Based Spatially Explicit Model as a Novel Tool for Investigating Insect-Plant Interactions: A Case Study of the Behavioural Ecology of Frugivorous Tephritidae. PLoS ONE 2016, 11, e0151777. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.Y.; Han, W.; Lin, D.; Chen, C.C.; Chen, X.L.; Jiang, F. Visual simulation of maize growth responding to armyworm (Mythimna separata) attack. Trans. Chin. Soc. Agric. Eng. 2019, 35, 191–198. [Google Scholar]
- Igarashi, T.; Matsuoka, S.; Tanaka, H. Teddy: A Sketching Interface for 3D Freeform Design//SIGGRAPH 07; ACM: San Diego, CA, USA, 1999. [Google Scholar] [CrossRef]
- Douglas, D.A.; Peucker, T.K. Algorithms for the reduction of the number of points required to represent a digitized line or its caricature. Can. Cartogr. 1973, 10, 112–122. [Google Scholar] [CrossRef] [Green Version]
- Tagliasacchi, A.; Alhashim, I.; Olson, M.; Zhang, H. Mean Curvature Skeletons. Comput. Graph. Forum 2012, 31, 1735–1744. [Google Scholar] [CrossRef] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Camera Parameters | Parameter Specification |
|---|---|
| Sensor Type | CMOS |
| Sensor size | 22.3 × 14.9 mm |
| Effective Pixels | 18 million |
| Image Processor | DIGIC 4 |
| Image resolution | Approx. 17.9 million pixels (5184 × 3456) |
| Image | Species | Model | Vertexes * | Faces * | Times (s) | Skeleton Points * |
|---|---|---|---|---|---|---|
| Figure 8(a1) | Eucryptorrhynchus brandti | Figure 8(b1) | 3812 | 7620 | 0.54 | 1720 |
| Figure 8(a2) | Cerambycidae | Figure 8(b2) | 6678 | 13,352 | 1.04 | 2074 |
| Figure 8(a3) | Ips shangrilla | Figure 8(b3) | 12,302 | 24,600 | 1.33 | 4644 |
| Figure 8(a4) | Cerambycidae | Figure 8(b4) | 5196 | 10,388 | 1.12 | 2057 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Yang, M.; Li, W. Three-Dimensional Reconstruction and Predictive Simulation Algorithm of Forest and Fruit Wood Borer Galleries Based on Two-Dimensional Images and Different Influencing Factors. Agronomy 2022, 12, 1087. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051087
Li X, Yang M, Li W. Three-Dimensional Reconstruction and Predictive Simulation Algorithm of Forest and Fruit Wood Borer Galleries Based on Two-Dimensional Images and Different Influencing Factors. Agronomy. 2022; 12(5):1087. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051087
Chicago/Turabian StyleLi, Xiao, Meng Yang, and Wanlu Li. 2022. "Three-Dimensional Reconstruction and Predictive Simulation Algorithm of Forest and Fruit Wood Borer Galleries Based on Two-Dimensional Images and Different Influencing Factors" Agronomy 12, no. 5: 1087. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051087