
Comparison of Proximal Remote Sensing Devices of Vegetable Crops to Determine the Role of Grafting in Plant Resistance to Meloidogyne incognita

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
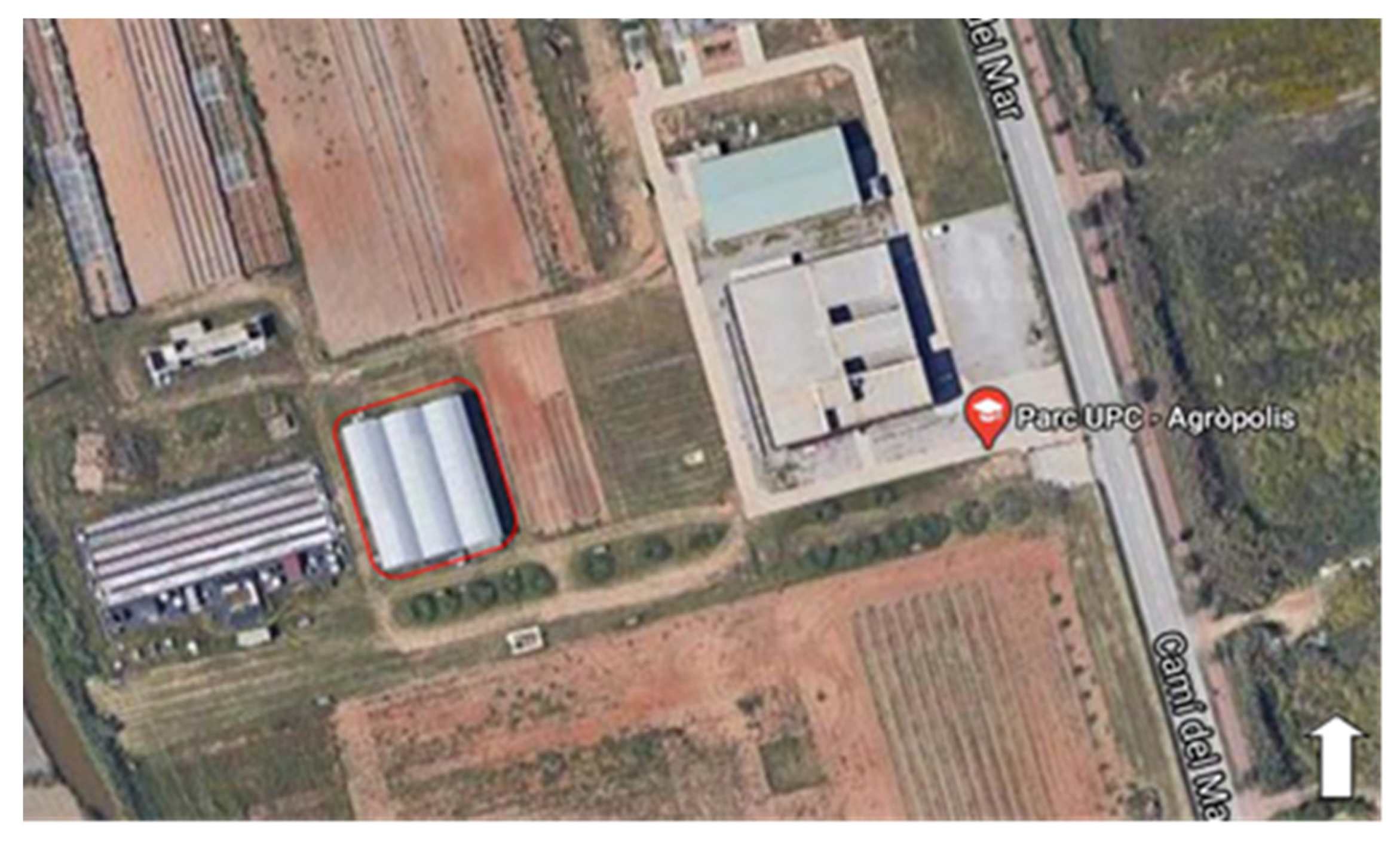
2.1. Study Site
2.2. Experimental Trial Designs
2.3. Sensors and Measurements
2.3.1. Determination of Leaf Level Pigments
SPAD
Dualex
2.3.2. Determination of Plant Health and Vigor
Trimble GreenSeeker NDVI
Red, Green, Blue (RGB) Images
2.3.3. Water Stress and Root Health
Porometer
Canopy Temperature
Determination of Stable Isotopes: 13C and 15N of the Soluble Fraction
2.4. Statistical Processing
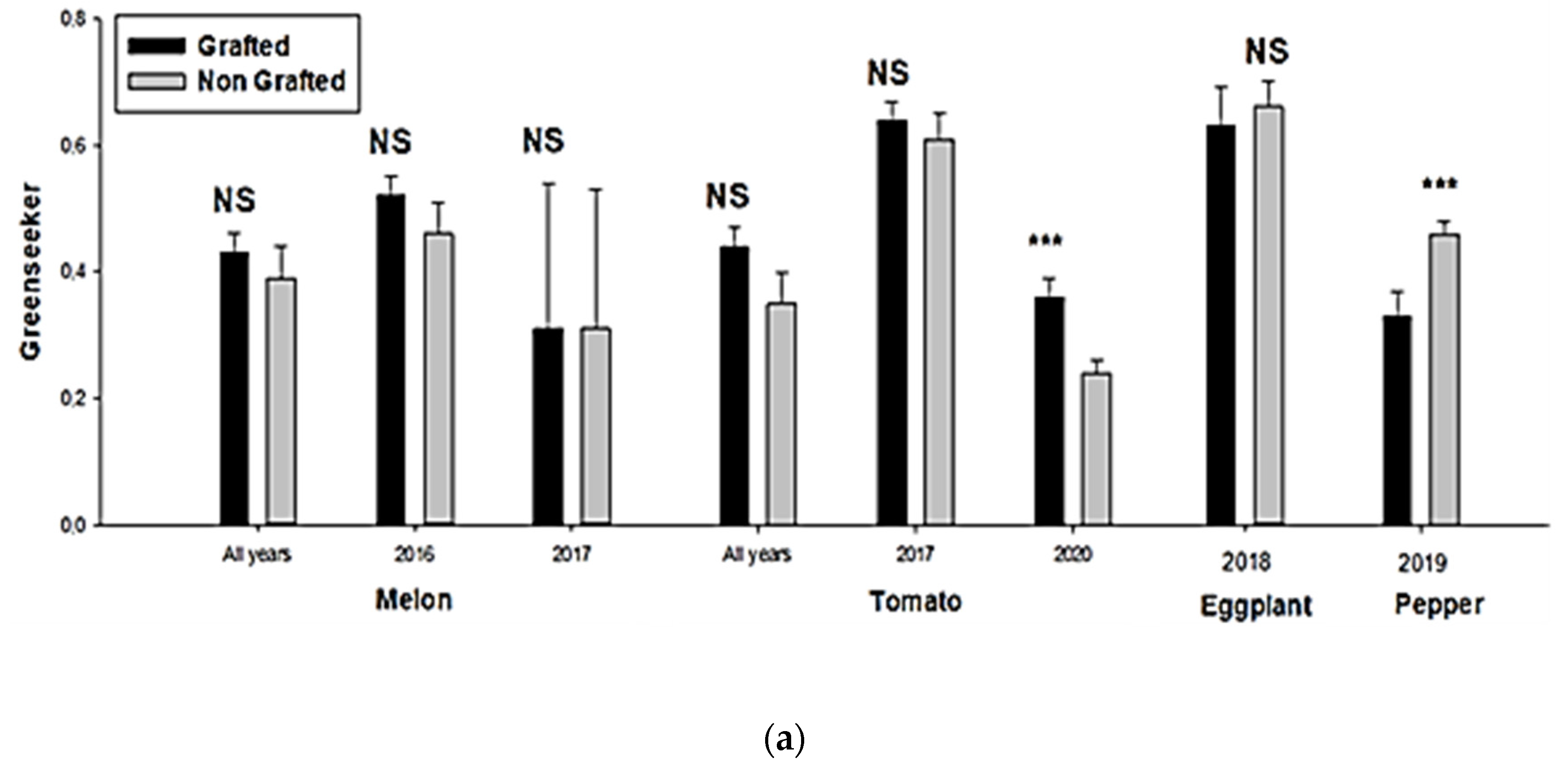
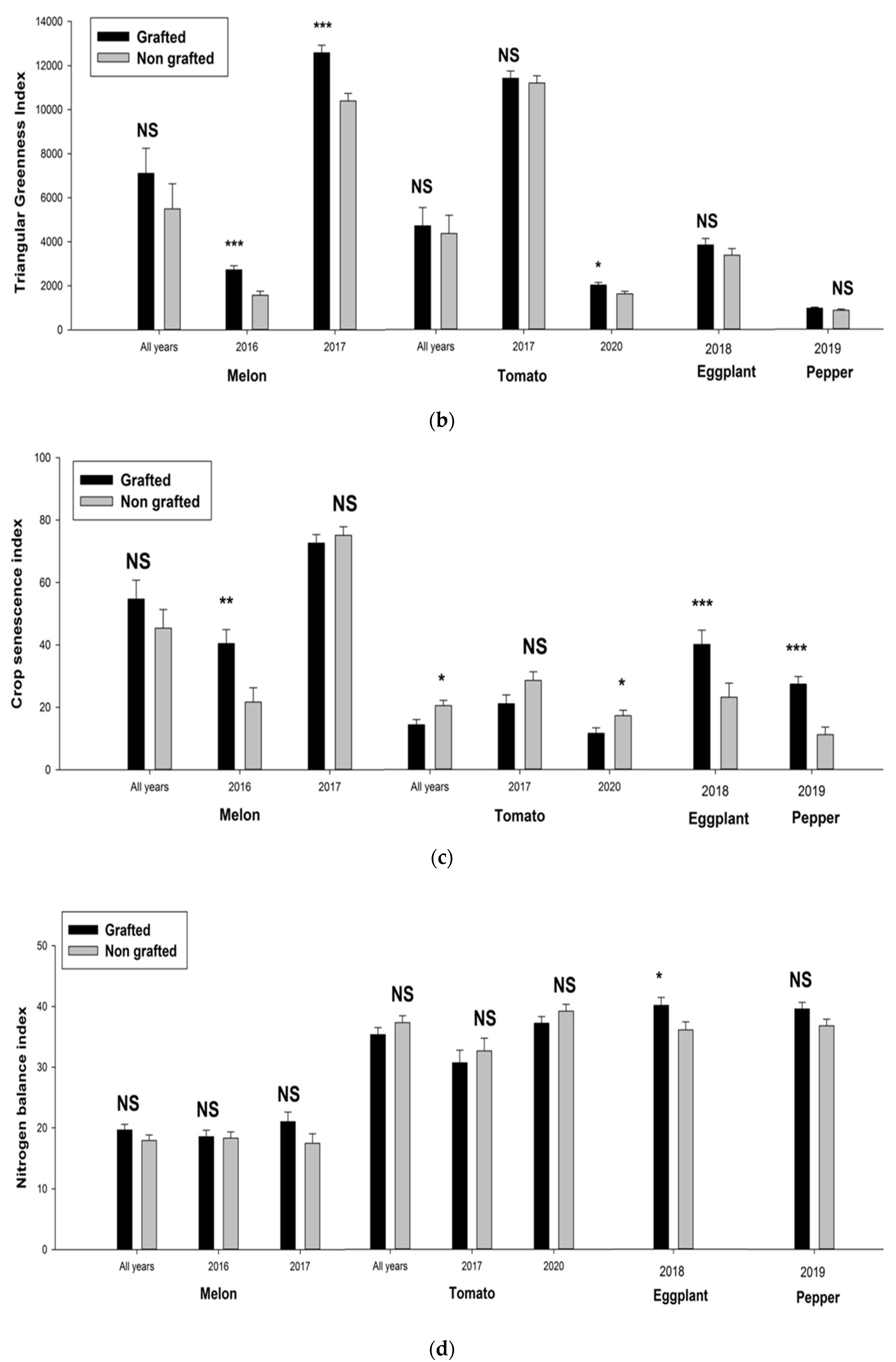
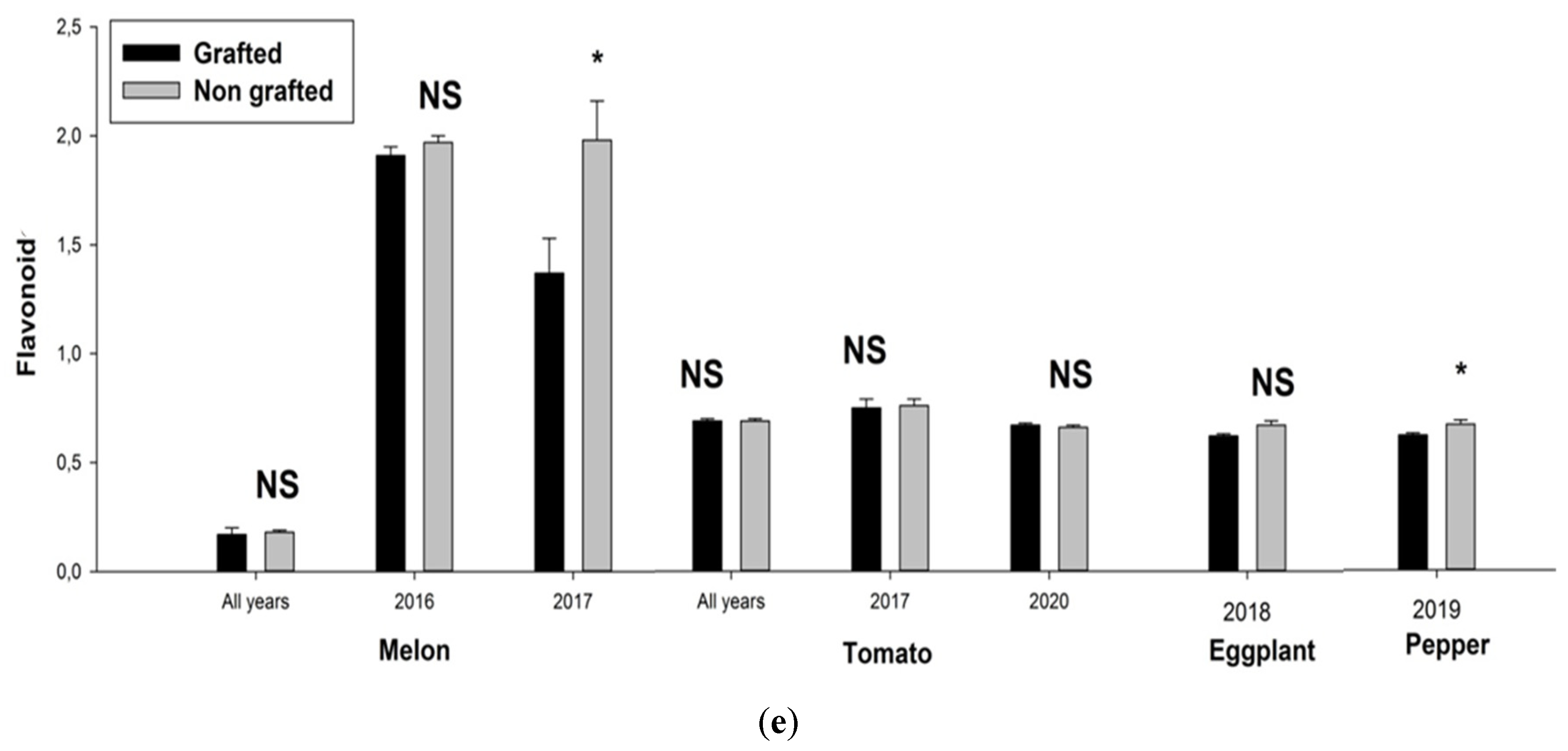
3. Results
3.1. Physiological Parameters
3.2. Crop Yield
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koenning, S.R.; Overstreet, C.; Noling, J.W.; Donald, P.A.; Becker, J.O.; Fortnum, B.A. Survey of crop losses in response to phytoparasitic nematodes in the United States for 1994. J. Nematol. 1999, 31, 587. [Google Scholar] [PubMed]
- Kashaija, I.; Kizito, F.; McIntyre, B.; Sali, H. Spatial distribution of roots, nematode populations and root necrosis in highland banana in Uganda. Nematology 2004, 6, 7–12. [Google Scholar] [CrossRef]
- Djian Caporalino, C. Root knot nematodes (Meloidogyne spp.), a growing problem in French vegetable crops. EPPO Bull. 2012, 42, 127–137. [Google Scholar] [CrossRef]
- Sorribas, F.J.; Ornat, C.; Verdejo-Lucas, S.; Galeano, M.; Valero, J. Effectiveness and profitability of the Mi-resistant tomatoes to control root-knot nematodes. Eur. J. Plant Pathol. 2005, 111, 29–38. [Google Scholar] [CrossRef]
- Blok, V.C.; Jones, J.T.; Phillips, M.S.; Trudgill, D.L. Parasitism genes and host range disparities in biotrophic nematodes: The conundrum of polyphag versus specialisation. Bioessays 2008, 30, 249–259. [Google Scholar] [CrossRef]
- Davila, M.; Dickson, D.W. Base temperature and heat unit requirements for development of Meloidogyne arenaria and Meloidogyne javanica. J. Nematol. 2004, 36, 314. [Google Scholar]
- Araus, J.L.; Kefauver, S.C. Breeding to adapt agriculture to climate change: Affordable phenotyping solutions. Curr. Opin. Plant Biol. 2018, 45, 237–247. [Google Scholar] [CrossRef]
- Araus, J.L.; Cairns, J.E. Field high-throughput phenotyping: The new crop breeding frontier. Trends Plant Sci. 2014, 19, 52–61. [Google Scholar] [CrossRef]
- Condon, A.G.; Richards, R.A.; Rebetzke, G.J.; Farquhar, G.D. Breeding for high water-use efficiency. J. Exp. Bot. 2004, 55, 2447–2460. [Google Scholar] [CrossRef] [Green Version]
- Evans, R.D. Physiological mechanisms influencing plant nitrogen isotope composition. Trends Plant Sci. 2001, 6, 121–126. [Google Scholar] [CrossRef]
- Rossato, L. Nitrogen storage and remobilization in Brassicanapus L. during the growth cycle: Effects of methyl jasmonate on nitrate uptake, senescence, growth, and VSP accumulation. J. Exp. Bot. 2002, 53, 1131–1141. [Google Scholar] [CrossRef] [PubMed]
- Malagoli, P.; Laine, P.; Rossato, L.; Ourry, A. Dynamics of nitrogen uptake and mobilization in field-grown winter oil seed rape (Brassicanapus) from stem extension to harvest: I. Global N flows between vegetative and reproductive tissues in relation to leaf fall and their residual N. Ann. Bot. 2005, 95, 853–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, B.J.; Fry, B. Stable isotopes in ecosystem studies. Annu. Rev. Ecol. Syst. 1987, 18, 293–320. [Google Scholar] [CrossRef]
- United Nations Economic Commission for Europe (UNECE) Fresh Fruit and Vegetables-Standards. Available online: https://unece.org/trade/wp7/FFV-Standards (accessed on 20 April 2022).
- Konica, M.O. Chlorophyll Meter SPAD-502 Plus-A Lightweight Handheld Meter for Measuring the Chlorophyll Content of Leaves without Causing Damage to Plants. 2012. Available online: http://www.konikcaminolta.com/instrments/download/catalog/color/pdf/spad502plus_e1.pdf (accessed on 12 March 2019).
- Kaufmann, H.; Segl, K.; Itzerott, S.; Bach, H.; Wagner, A.; Hill, J.; Müller, A. Hyperspectral Algorithms: Report in the Frame of EnMAP Preparation Activities; Potsdam: Darst, Germany, 2010. [Google Scholar]
- Cerovic, Z.G.; Masdoumier, G.; Ghozlen, N.B.; Latouche, G. A new optical leaf-clip meter for simultaneous non-destructive assessment of leaf chlorophyll and epidermal flavonoids. Physiol. Plant. 2012, 146, 251–260. [Google Scholar] [CrossRef]
- Cerovic, Z.G.; Ghozlen, N.B.; Milhade, C.; Obert, M.; Debuisson, S.; Le Moigne, M. Nondestructive Diagnostic Test for Nitrogen Nutrition of Grapevine (Vitis vinifera L.) Based on Dualex Leaf-Clip Measurements in the Field. J. Agric. Food Chem. 2015, 63, 3669–3680. [Google Scholar] [CrossRef]
- Gracia-Romero, A.; Kefauver, S.C.; Fernandez-Gallego, J.A.; Vergara-Díaz, O.; Nieto-Taladriz, M.T.; Araus, J.L. UAV and ground image-based phenotyping: A proof of concept with Durum wheat. Remote Sens. 2019, 11, 1244. [Google Scholar] [CrossRef] [Green Version]
- Kefauver, S.; Kerfal, S.; Fernandez Gallego, J.A.; El-Haddad, G. CerealScanner Gitlab. Available online: https://gitlab.com/sckefauver/cerealscanner (accessed on 14 March 2019).
- Zaman-Allah, M.; Vergara, O.; Araus, J.L.; Tarekegne, A.; Magorokosho, C.; Zarco-Tejada, P.J.; Hornero, A.; Albà, A.H.; Das, B.; Craufurd, P.; et al. Unmanned aerial platform-based multi-spectral imaging for field phenotyping of maize. Plant Methods 2015, 11, 35. [Google Scholar] [CrossRef] [Green Version]
- Hunt, E.R.; Cavigelli, M.; Daughtry, C.S.; Mcmurtrey, J.E.; Walthall, C.L. Evaluation of digital photography from model aircraft for remote sensing of crop biomass and nitrogen status. Precis. Agric. 2005, 6, 359–378. [Google Scholar] [CrossRef]
- Hunt, E.R.; Doraiswamy, P.C.; Mcmurtrey, J.E.; Daughtry, C.S.T.; Perry, E.M.; Akhmedov, B. A visible band index for remote sensing leaf chlorophyll content at the canopy scale. Int. J. Appl. Earth Obser. Geoinf. 2013, 21, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Vergara-Díaz, O.; Zaman-Allah, M.A.; Masuka, B.; Hornero, A.; Zarco-Tejada, P.; Prasanna, B.M.; Araus, J.L. A novel remote sensing approach for prediction of maize yield under different conditions of nitrogen fertilization. Front. Plant Sci. 2016, 7, 666. [Google Scholar] [CrossRef] [Green Version]
- Stern, A.; Doraiswamy, P.C.; Hunt Jr, E.R. Changes of crop rotation in Iowa determined from the United States Department of Agriculture, National Agricultural Statistics Service cropland data layer product. J. Appl. Remote Sens. 2012, 6, 063590. [Google Scholar] [CrossRef]
- Hunt, E.R.; Daughtry, C.S.T.; Eitel, J.U.; Long, D.S. Remote sensing leaf chlorophyll content using a visible band index. Agron. J. 2011, 103, 1090–1099. [Google Scholar] [CrossRef] [Green Version]
- Montague, T.; Hellman, E.; Krawitzky, M. Comparison of greenhouse grown, containerized grapevine stomatal conductance measurements using two differing porometers. In Proceedings of the 2nd Annual National Viticulture Research Conference, Davis, CA, USA, 9–11 July 2008; pp. 58–61. [Google Scholar]
- Silva-Sánchez, A.; Buil-Salafranca, J.; Cabral, A.C.; Uriz-Ezcaray, N.; García-Mendívil, H.A.; Sorribas, F.J.; Gracia-Romero, A. Comparison of proximal remote sensing devices for estimating physiological responses of eggplants to root-knot nematodes. Proceedings 2019, 18, 9. [Google Scholar]
- Duncan, G.A.; Gates, R.; Montross, M.D. Measuring Relative Humidity in Agricultural Environments; Agricultural Engineering Extension Publications-Uknowledge: Lexington, KY, USA, 2005. [Google Scholar]
- Cabrera-Bosquet, L.; Albrizio, R.; Nogues, S.; Araus, J.L. Dual Delta 13C/delta 18O response to water and nitrogen availability and its relationship with yield in field-grown durum wheat. Plant Cell Environ. 2011, 34, 418–433. [Google Scholar] [CrossRef] [PubMed]
- Yousfi, S.; Serret, M.D.; Araus, J.L. Comparative response of d13C, d18O and d15N in durum wheat exposed to salinity at the vegetative and reproductive stages. Plant Cell Environ. 2013, 36, 1214–1227. [Google Scholar] [CrossRef]
- Fernandez-Gallego, J.A.; Kefauver, S.C.; Gutiérrez, N.A.; Nieto-Taladriz, M.T.; Araus, J.L. Wheat ear counting in-field conditions: High throughput and low-cost approach using RGB images. Plant Methods 2018, 14, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Miguel, A.; Maroto, J.V.; San Bautista, A.; Baixauli, C.; Cebolla, V.; Pascual, B.; Guardiola, J.L. The grafting of triploid watermelon is an advantageous alternative to soil fumigation by methyl bromide for control of Fusarium wilt. Sci. Hortic. 2004, 103, 9–17. [Google Scholar] [CrossRef]
- Casadesus, J.; Kaya, Y.; Bort, J.; Nachit, M.M.; Araus, J.L.; Amor, S.; Ferrazzano, G.; Maalouf, F. Using vegetation indices derived from conventional digital cameras as selection criteria for wheat breeding in water-limited environments. Ann. Appl. Biol. 2007, 150, 227–236. [Google Scholar] [CrossRef]
- De Guiran, G. Protection des Cultures Maraîchères et Fruitières Face Aux Capacités D’adaptation des Nématodes Meloidogyne; Comptes Rendus de l’Académie d’agriculture: Paris, France, 1983. [Google Scholar]
- Feng, X.; Zhan, Y.; Wang, Q.; Yang, X.; Yu, C.; Wang, H.; He, Y. Hyperspectral imaging combined with machine learning as a tool to obtain high-throughput plant salt-stress phenotyping. Plant J. 2020, 101, 1448–1461. [Google Scholar] [CrossRef]
- Yang, M.D.; Tseng, H.H.; Hsu, Y.C.; Tsai, H.P. Semantic segmentation using deep learning with vegetation indices for rice lodging identification in multi-date UAV visible images. Remote Sens. 2020, 12, 633. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, T.R.; Rufty, T.W. Nitrogen and water resources commonly limit crop yield increases, not necessarily plant genetics. Glob. Food Secur. 2012, 1, 94–98. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Queval, G.; Foyer, C.H. The impact of global change factors on redox signaling underpinning stress tolerance. Plant Physiol. 2013, 161, 5–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giné, A.; González, C.; Serrano, L.; Sorribas, F.J. Population dynamics of Meloidogyne incognita on cucumber grafted onto the Cucurbita hybrid RS841 or non-grafted and yield losses under protected cultivation. Eur. J. Plant Pathol. 2017, 148, 795–805. [Google Scholar] [CrossRef] [Green Version]
- Expósito, A.; Pujolà, M.; Achaerandio, I.; Giné, A.; Escudero, N.; Fullana, A.M.; Cunquero, M.; Loza-Alvarez, P.; Sorribas, F.J. Tomato and melon Meloidogyne resistant rootstocks improve crop yield but melon fruit quality is influenced by the cropping season. Front. Plant Sci. 2020, 1742. [Google Scholar] [CrossRef] [PubMed]
- Goverse, A.; Smant, G. The activation and suppression of plant innate immunity by parasitic nematodes. Ann Rev. Phytopath. 2014, 52, 243–265. [Google Scholar]
- Haverkort, A.J.; Valkenburg, G.W. The influence of cyst nematodes and drought on potato growth. 3. Effects on carbon isotope fractionation. Neth. J. Plant Pathol. 1992, 98, 12–20. [Google Scholar] [CrossRef]
- Rivard, C.L.; Louws, F.J. Grafting to manage soilborne diseases in heirloom tomato production. Hort Sci. 2008, 43, 2104–2111. [Google Scholar] [CrossRef] [Green Version]
- Guan, W.; Zhao, X.; Dickson, D.W.; Mendes, M.L.; Thies, J. Root-knot nematode resistance, yield, and fruit quality of specialty melons grafted onto Cucumis metulifer. Hort Sci. 2014, 49, 1046–1051. [Google Scholar] [CrossRef] [Green Version]
- Sigüenza, C.; Schochow, M.; Turini, T.; Ploeg, A. Use of Cucumis metuliferus as a rootstock for melon to manage Meloidogyne incognita. J. Nematol. 2005, 37, 276. [Google Scholar]
- Expósito, A.; Munera, M.; Giné, A.; López-Gómez, M.; Cáceres, A.; Picó, B.; Gisbert, C.; Medina, V.; Sorribas, F.J. Cucumis metuliferus is resistant to root-knot nematode Mi1.2 gene (a)virulent isolates and a promising melon rootstock. Plant Pathol. 2018, 67, 1161–1167. [Google Scholar] [CrossRef] [Green Version]
- Expósito, A.; García, S.; Giné, A.; Escudero, N.; Sorribas, F.J. Cucumis metuliferus reduces Meloidogyne incognita virulence against the Mi1.2 resistance gene in a tomato–melon rotation sequence. Pest Manag. Sci. 2019, 75, 1902–1910. [Google Scholar] [CrossRef]
- Bletsos, F.A. Use of grafting and calcium cyanamide as alternatives to methyl bromide soil fumigation and their effects on growth, yield, quality, and fusarium wilt control in melon. J. Phytopathol. 2005, 153, 155–161. [Google Scholar] [CrossRef]
- Tao, M.Q.; Jahan, M.S.; Hou, K.; Shu, S.; Wang, Y.; Sun, J.; Guo, S.-R. Bitter Melon (Momordica charantia L.) Rootstock Improves the Heat Tolerance of Cucumber by Regulating Photosynthetic and Antioxidant Defense Pathways. Plants 2020, 9, 692. [Google Scholar] [CrossRef] [PubMed]
- Abd-Elgawad, M.M. Biological control agents in the integrated nematode management of potato in Egypt. Egypt. J. Biol. Pest Control. 2020, 30, 1–13. [Google Scholar] [CrossRef]
- Gassmann, A.J.; Stock, S.P.; Sisterson, M.S.; Carrière, Y.; Tabashnik, B.E. Synergism between entomopathogenic nematodes and Bacillus thuringiensis crops: Integrating biological control and resistance management. J. Appl. Ecol. 2008, 45, 957–966. [Google Scholar] [CrossRef]
- Nisha, M.S.; Sheela, M.S. Bio-Management of Meloidogyne incognita on Coleus, Solenostemon rotundifolius by Integrating Solarization, Paecilomyces lilacinus, Bacillus macerans and Neemcake. Indian J. Nematol. 2006, 36, 136–138. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Size and Description | Microscopic Soil Worm Measuring 0.3 mm Long (2nd Grade Juvenile Free Stage in the Soil) at 0.7 mm (Female Obese Pear Shaped in the Root). Oral Perforator Stylus [6]. |
|---|---|
| Reproduction | Sexual or asexual (parthenogenesis) [1]. |
| Life cycle | 3 weeks to 3 months (depending on temperature). Mandatory endoparasite (inside the root). Eggs/juveniles/adults: 4 successive molts –evolution, Female [1]. |
| Multiplication | Lays 300 to 1000 eggs per cycle, Several possible cycles per year = 300 to 200,000 eggs per year [1]. |
| Conservation | In the form of eggs in the soil, between 5 and 30 cm deep [1]. |
| Survival | Juveniles live at least up to 15 days, depending on environmental conditions (pH, temperature, soil moisture, presence or not of plants). Eggs > 1 year, under certain conditions. Dispersion can be by humans (shoes, tools, machines) and by water at stage J2 [6] |
| Wales | Damage to roots (gall index from 0 to 10). Wilting, withering or even death of plants [1]. |
| Main hosts | Vegetables: asparagus, eggplant, vegetable beet, carrot, celery, chicory, cucumber, melon, pumpkin, zucchini, spinach, beans, lettuce, onion, pepper, tomato, potato, leek; rapeseed; cereals; fruit trees; flower crops; weeds including Rumex spp., amaranth, nightshade [6]. |
| Protection | Prophylaxis: cleaning, disinfection of tools, no spreading potentially by waste or sludge. Physical protection: solarization, steam disinfection, soil flood. Biological protection: organic matter, bacteria, mushrooms, mycorrhizae. Chemical protection: pre- and post-planting, treatment seeds, plant extract Crop protection: rotation, trap plant, green manure “nematicide”, black fallow, bio-fumigation, anaerobic bio-disinfection Varietal protection: resistance, grafting [6]. |
| Target Group | Index | Formula |
|---|---|---|
| Vegetation Cover | Green Area (GA) | 60 < Hue < 180 [20] |
| Greener Area (GGA) | 80 < Hue <180 [20] | |
| Greenness | Crop Senescence Index (CSI) | (GA-GGA)/GA [24] |
| Normalized Green-Red Difference Index (NGRDI) | (R550-R670)/(R550 + R670) [25] | |
| Triangular Greenness Index (TGI) | −0.5[190(R670 − R550) −120(R670 − R480) [26] |
| Parameters | Mean Melon 2016 + 2017 n = 36 (20 + 16) | Mean Tomato 2017 + 2020 n = 56 (16 + 40) | Mean Eggplant 2018 n = 20 | Mean pepper 2019 n = 40 | |
| Leaf level pigments (SPAD, Dualex) | SPAD | 39.42 ± 1.16 | 44.58 ± 0.93 | 40.16 ± 1.56 | 55.03 ± 1.10 |
| Chl | 33.20 ± 0.67 | 24.20 ± 0.54 | 24.04 ± 0.90 | 24.76 ± 0.64 | |
| Flav | 1.82 ± 0.04 | 0.69 ± 0.05 | 0.65 ± 0.04 | 0.66 ± 0.03 | |
| Anth | 0.17 ± 0.01 | 0.05 ± 0.02 | 0.02 ± 0.04 | 0.02 ± 0.02 | |
| NBI | 18.79 ± 0.83 | 36.31 ± 0.67 | 38.09 ± 1.12 | 38.17 ± 0.79 | |
| Canopy vigor, biomass (GA and GGA up to RGB) | NDVI | 0.41 ± 0.02 | 0.39 ± 0.04 | 0.64 ± 0.03 | 0.39 ± 0.02 |
| GA | 0.43 ± 0.03 | 0.37 ± 0.02 | 0.68 ± 0.04 | 0.07 ± 0.03 | |
| GGA | 0.22 ± 0.03 | 0.49 ± 0.02 | 0.48 ± 0.03 | 0.06 ± 0.02 | |
| NGRDI | −0.60 ± 0.36 | 1.46 ± 0.29 | 0.05 ± 0.48 | −0.13 ± 0.35 | |
| TGI | 6300.72 ± 603.65 | 4525.35 ± 484 | 3601.41 ± 809.89 | 895.58 ± 588.37 | |
| CSI | 50.09 ± 2.58 | 17.45 ± 2.07 | 31.63 ± 3.46 | 19.29 ± 2.45 | |
| Water stress and root health | Porometer | 102.60 ± 5.89 | 123.21 ± 4.72 | ||
| Temp | 25.18 ± 0.27 | 24.36 ± 0.22 | 25.53 ± 0.36 | 26.67 ± 0.26 | |
| δ13C | 14.16 ± 1.13 | −30.57 ± 0.89 | −30.19 ± 1.54 | −29.58 ± 1.06 | |
| Percent C | 21.12 ± 1.35 | 36.81 ± 1.07 | 37.94 ± 1.84 | 26.63 ± 1.26 | |
| δ15N | 4.02 ± 0.30 | 4.63 ± 0.24 | 6.05 ± 0.41 | 7.12 ± 0.28 | |
| Percent N | 1.16 ± 0.13 | 3.31 ± 0.10 | 2.96 ± 0.18 | 1.96 ± 0.12 |
| Parameters | p Value Treatments (Grafted, Non-Grafted) | p Value Interaction (Crop * Treatments) | |
|---|---|---|---|
| Leaf level pigments (SPAD and Dualex) | SPAD | 0.110 | 0.380 |
| Chl | 0.140 | 0.066 | |
| Flav | 0.003 | 0.002 | |
| Anth | 0.607 | 0.565 | |
| NBI | 0.060 | 0.044 | |
| Canopy vigor, biomass (GA and GGA up to RGB) | NDVI | 0.768 | 0.004 |
| GA | 0.725 | 0.606 | |
| GGA | 0.102 | 0.810 | |
| NGRDI | 0.474 | 0.889 | |
| TGI | 0.322 | 0.810 | |
| CSI | 0.001 | 0.002 | |
| Water stress and root health | Porometer | 0.724 | 0.267 |
| Temperature | 0.178 | 0.654 | |
| δ13C | 0.819 | 0.984 | |
| Percent C | 0.702 | 0.925 | |
| δ15N | 0.335 | 0.121 | |
| Percent N | 0.793 | 0.210 |
| Crop Yield (kg/plant) | |||
|---|---|---|---|
| Year | Crop | Grafted | Non-grafted |
| 2016 | Melon | 0.8 ± 0.2 | 0.1 ± 0.1 * |
| 2017 | Melon | 3.1 ± 0.3 | 0.3 ± 0.1 * |
| Tomato | 2.9 ± 0.2 | 2.0 ± 0.2 * | |
| 2018 | Eggplant | 4.0 ± 0.4 | 1.9 ± 0.2 * |
| 2019 | Pepper | 0.4 ± 0.002 | 0.2 ± 0.05 |
| 2020 | Tomato | Resistant cv. Caramba | Susceptible cv. Durinta |
| 1.6 ± 0.01 | 2.2 ± 0.2 * | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamdane, Y.; Gracia-Romero, A.; Buchaillot, M.L.; Sanchez-Bragado, R.; Fullana, A.M.; Sorribas, F.J.; Araus, J.L.; Kefauver, S.C. Comparison of Proximal Remote Sensing Devices of Vegetable Crops to Determine the Role of Grafting in Plant Resistance to Meloidogyne incognita. Agronomy 2022, 12, 1098. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051098
Hamdane Y, Gracia-Romero A, Buchaillot ML, Sanchez-Bragado R, Fullana AM, Sorribas FJ, Araus JL, Kefauver SC. Comparison of Proximal Remote Sensing Devices of Vegetable Crops to Determine the Role of Grafting in Plant Resistance to Meloidogyne incognita. Agronomy. 2022; 12(5):1098. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051098
Chicago/Turabian StyleHamdane, Yassine, Adrian Gracia-Romero, Maria Luisa Buchaillot, Rut Sanchez-Bragado, Aida Magdalena Fullana, Francisco Javier Sorribas, José Luis Araus, and Shawn C. Kefauver. 2022. "Comparison of Proximal Remote Sensing Devices of Vegetable Crops to Determine the Role of Grafting in Plant Resistance to Meloidogyne incognita" Agronomy 12, no. 5: 1098. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051098