
Genetic Analysis of S5 Regulating the Hybrid Sterility between Indica and Japonica Subspecies in Rice

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Plasmid Construction and Plant Transformation
2.3. PCR Amplification and DNA Analysis
2.4. Fertility Examination
2.5. Agronomic Traits and Statistical Analysis
2.6. Sample Collection and Total RNA Extraction
2.7. Library Construction and RNA-Seq Analysis
2.8. DEG Identification and Analysis
2.9. Real-Time Quantitative PCR Analysis
3. Results
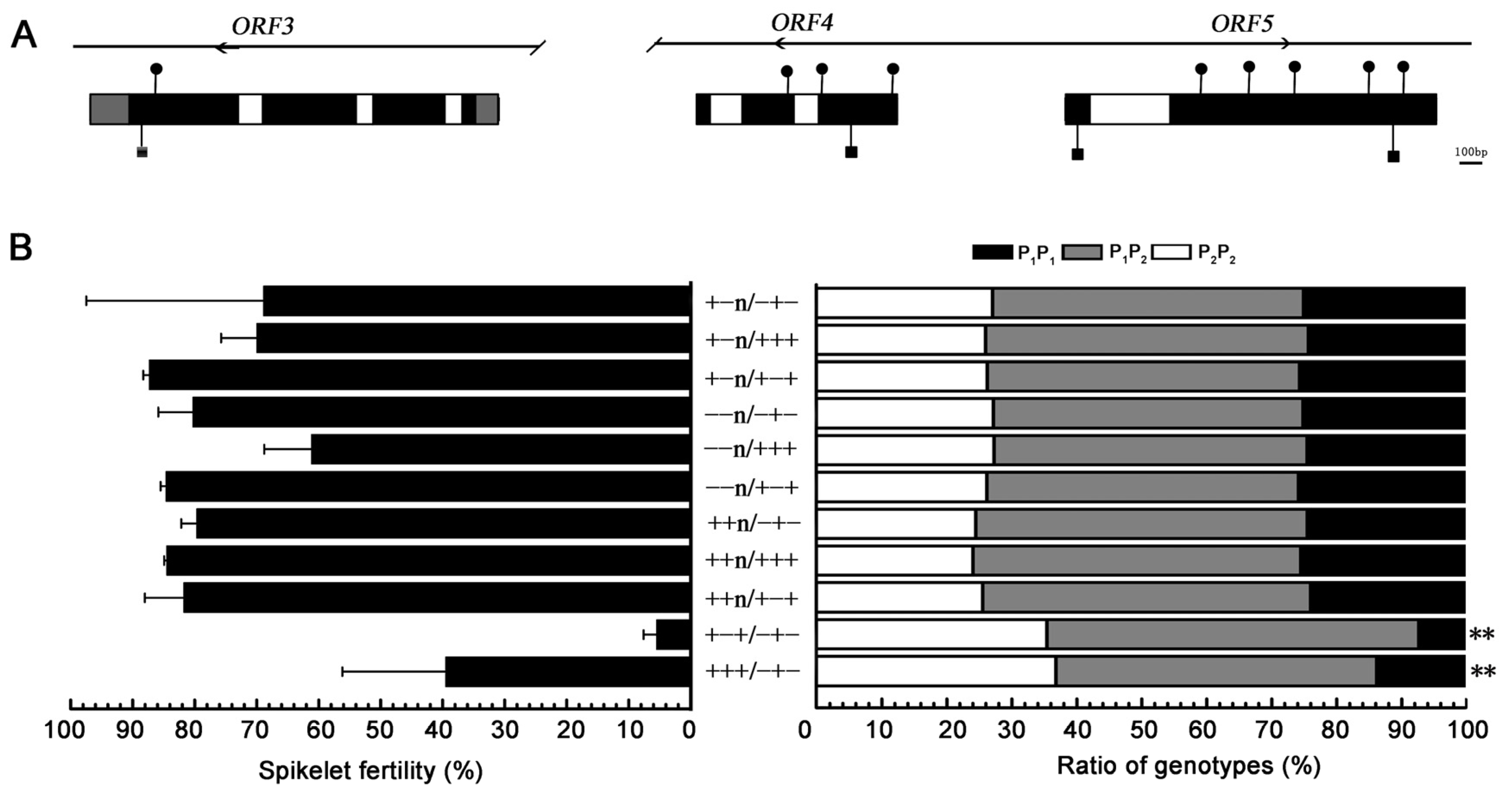
3.1. Nucleotide Variations and Genetic Effect of S5 Locus
3.2. Allelic Interactions of ORF4 in the HJX74-SSSLs
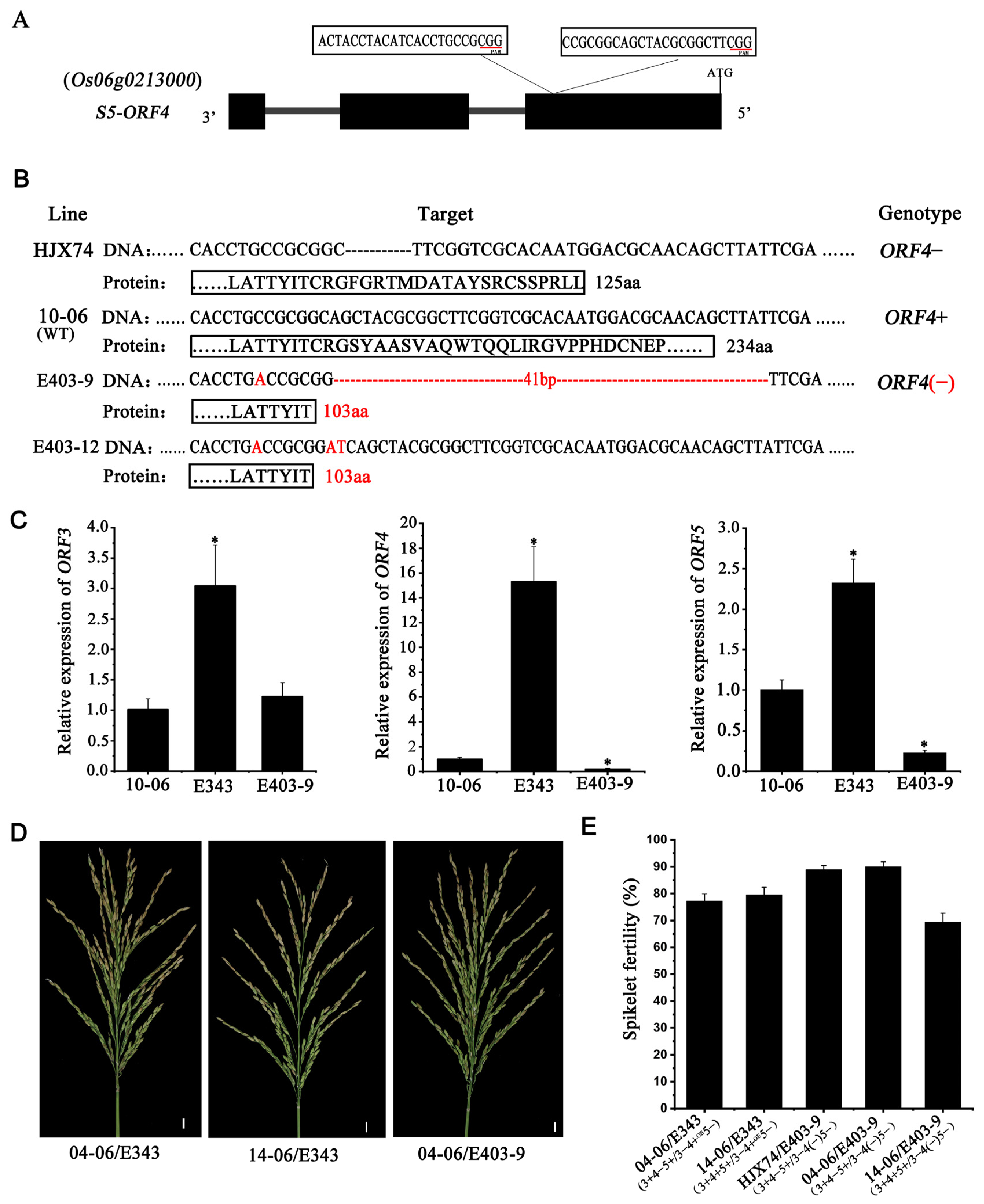
3.3. CRISPR/Cas9-Mediated Editing of ORF4 Gene in HJX74-SSSL
3.4. ORF4(−) Improved Indica/Japonica Hybrid Fertility in HJX74 Background
3.5. Overexpression of ORF4 Gene in HJX74-SSSL
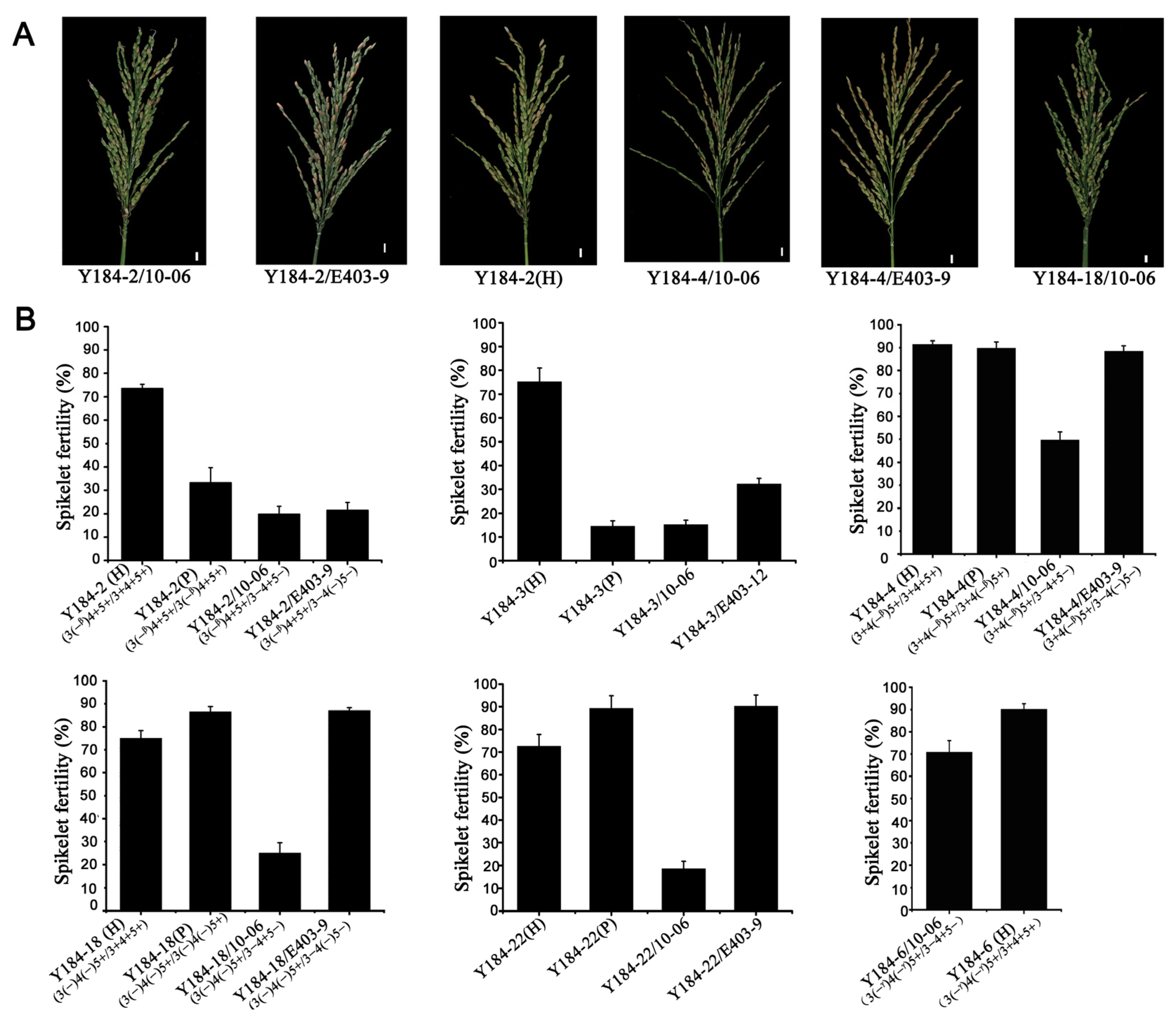
3.6. CRISPR/Cas9-Mediated Editing of ORF3 and ORF4 Gene in HJX74-SSSL
3.7. Strong Influence on Hybrid Sterility by Editing of ORF3 and ORF4 in HJX74 Background
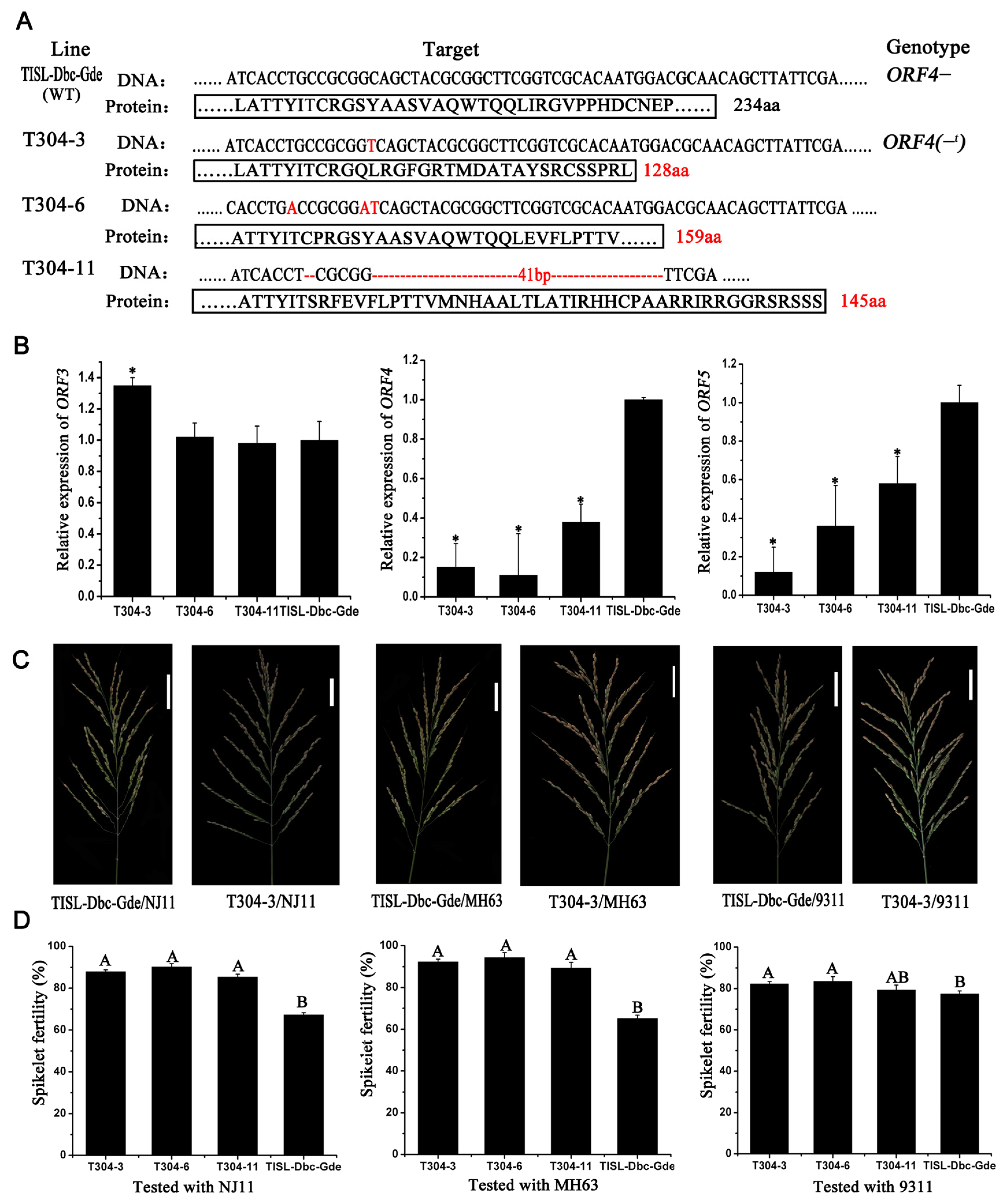
3.8. Konckout the ORF4 Gene to Overcome Hybrid Sterility Caused by S5-Dependent in Indica/Japonica Background
3.9. Comparative Transcriptome Analysis of Young Spikelet in Different Hybrids by RNA-Seq
3.10. Analysis of Genes Differentially Expressed in Hybrids F1 (ORF4+/ORF4+) Compare to Hybrids F1 (ORF4+/ORF4−)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khush, G.S.; Gupta, P. Strategies for increasing the yield potential of cereals: Case of rice as an example. Plant Breed. 2013, 132, 433–436. [Google Scholar] [CrossRef]
- Garris, A.J.; Tai, T.H.; Coburn, J.; Kresovich, S.; McCouch, S. Genetic structure and diversity in Oryza sativa L. Genetics 2005, 169, 1631–1638. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Guan, Z.; Gong, Z.; Yan, J.; Yang, G.; Liu, Y.G.; Yin, P. Crystal structure of WA352 provides insight into cytoplasmic male sterility in rice. Biochem. Biophys. Res. Commun 2018, 501, 898–904. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.P.; Maroof, M.A.; Xu, C.G.; Zhang, Q.; Biyashev, R.M. Comparative analysis of microsatellite DNA polymorphism in landraces and cultivars of rice. Mol. Gen. Genet. 1994, 245, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Ying, Y.G.; Wang, B.X.; Chen, T.M.; Yang, B.; Liu, Y.; Xu, B.; Sun, Z.G.; Chi, M.; Li, J.; Li, J.M.; et al. Heterosis analysis of yield and quality characters of two-line indica-japonica hybrid combination. Jiangsu Agric. Sci. 2021, 49, 59–63. [Google Scholar]
- Cheng, S.H.; Zhuang, J.Y.; Fan, Y.Y.; Du, J.H.; Cao, L.Y. Progress in research and development on hybrid rice: A super-domesticate in China. Ann. Bot. 2007, 100, 959–966. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.Q. Prospects of utilization of inter-subspecific heterosis between indica and japonica rice. J. Integr. Agric. 2020, 19, 1–10. [Google Scholar] [CrossRef]
- Zhang, G.Q. The Next Generation of Rice: Inter-subspecific indica-japonica hybrid rice. Front. Plant Sci. 2022, 13, 857896. [Google Scholar] [CrossRef]
- Kato, A. On the affinity of rice varieties as shown by the fertility of rice plants. Centr. Agric. Inst. Kyushu Imp. Univ. 1928, 2, 241–276. [Google Scholar]
- Ikehashi, H.; Araki, H. Varietal screening of compatibility types revealed in F1 fertility of distant crosses in rice. Jpn. J. Breed. 1984, 34, 304–313. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, Y.D.; Zhang, Q.F. The molecular and evolutionary basis of reproductive isolation in plants. J. Genet. Genom. 2018, 45, 613–620. [Google Scholar] [CrossRef]
- Ouyang, Y.D.; Liu, Y.G.; Zhang, Q.F. Hybrid sterility in plant: Stories from rice. Curr. Opin. Plant Biol. 2010, 13, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Y.; Chen, J.J.; Ding, J.H.; Zhang, Q.F. Advances in the understanding of inter-subspecific hybrid sterility and wide-compatibility in rice. Chin. Sci. Bull. 2009, 54, 2332–2341. [Google Scholar] [CrossRef]
- Li, J.; Zhou, J.; Zhang, Y.; Yang, Y.; Tao, D. New insights into the nature of interspecific hybrid sterility in rice. Front. Plant Sci. 2020, 11, 555572. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Wang, Z.; Zhu, X.; Tang, Y.; Ye, L.; Yu, H.; Li, Y.; Zhang, N.; Liu, T.; Wang, T.; et al. A minimal genome design to maximally guarantee fertile inter-subspecific hybrid rice. Mol. Plant 2023, 16, 726–738. [Google Scholar] [CrossRef]
- Shen, R.; Wang, L.; Liu, X.; Wu, J.; Jin, W.; Zhao, X.; Xie, X.; Zhu, Q.; Tang, H.; Li, Q.; et al. Genomic structural variation-mediated allelic suppression causes hybrid male sterility in rice. Nat. Commun. 2017, 8, 1310. [Google Scholar] [CrossRef] [Green Version]
- Mizuta, Y.; Harushima, Y.; Kurata, N. Rice pollen hybrid incompatibility caused by reciprocal gene loss of duplicated genes. Proc. Natl. Acad. Sci. USA 2010, 107, 20417–20422. [Google Scholar] [CrossRef] [Green Version]
- Long, Y.; Zhao, L.; Niu, B.; Su, J.; Wu, H.; Chen, Y.; Zhang, Q.; Guo, J.; Zhuang, C.; Mei, M.; et al. Hybrid male sterility in rice controlled by interaction between divergent alleles of two adjacent genes. Proc. Natl. Acad. Sci. USA 2008, 105, 18871–18876. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Ding, J.; Ouyang, Y.; Du, H.; Yang, J.; Cheng, K.; Zhao, J.; Qiu, S.; Zhang, X.; Yao, J.; et al. A triallelic system of S5 is a major regulator of the reproductive barrier and compatibility of indica-japonica hybrids in rice. Proc. Natl. Acad. Sci. USA 2008, 105, 11436–11441. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Zhao, Z.; Shi, Y.; Tian, H.; Liu, L.; Bian, X.; Xu, Y.; Zheng, X.; Gan, L.; Shen, Y.; et al. Hybrid sterility in rice (Oryza sativa L.) involves the tetratricopeptide repeat domain containing protein. Genetics 2016, 203, 1439–1451. [Google Scholar] [CrossRef] [Green Version]
- Kubo, T.; Takashi, T.; Ashikari, M.; Yoshimura, A.; Kurata, N. Two tightly linked genes at the hsa1 locus cause both F1 and F2 hybrid sterility in rice. Mol. Plant 2016, 9, 221–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitamura, E. Genetic studies on sterility observed n hybrids between distantly related varieties of rice, Oryza sativa L. Rep. Chugoku Exp. Stat. 1962, 8, 141–205. [Google Scholar]
- Oka, H.I. Genic analysis for the sterility of hybrids between distantly related varieties of cultivated rice. J. Genet. 1957, 55, 397–409. [Google Scholar] [CrossRef]
- Oka, H.I. Analysis of genes controlling F1 sterility in rice by the use of isogenic lines. Genetics 1974, 77, 521–534. [Google Scholar] [CrossRef]
- Wei, C.M.; Wang, L.; Yang, Y.X.; Chen, Z.X.; Shahid, M.Q.; Li, J.Q.; Liu, X.D.; Lu, Y.G. Identification of an S5n allele in Oryza rufipogon Griff. and its effect on embryo sac fertility. Chin. Sci. Bull. 2010, 55, 1255–1262. [Google Scholar] [CrossRef]
- Du, H.; Ouyang, Y.; Zhang, C.; Zhang, Q. Complex evolution of S5, a major reproductive barrier regulator, in the cultivated rice Oryza sativa and its wild relatives. New Phytol. 2011, 191, 275–287. [Google Scholar] [CrossRef]
- Tong, J.F.; Li, Y.H.; Yang, Y.X.; Shahid, M.Q.; Lu, Y.G. Molecular evolution of rice S5n and functional comparison among different sequences. Chin. Sci. Bull. 2011, 56, 2016–2024. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.; Shahid, M.Q.; Li, Y.H.; Tong, J.F.; Zhou, X.W.; Liu, X.D.; Lu, Y.G. Molecular evolution of S5 locus and large differences in its coding region revealed insignificant effect on indica/japonica embryo sac fertility. Plant Syst. Evol. 2015, 301, 639–655. [Google Scholar] [CrossRef]
- Yang, J.; Zhao, X.; Cheng, K.; Du, H.; Ouyang, Y.; Chen, J.; Qiu, S.; Huang, J.; Jiang, Y.; Jiang, L.; et al. A killer-protector system regulates both hybrid sterility and segregation distortion in rice. Science 2012, 337, 1336–1340. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.F.; Yu, Y.M.; Cheng, K.; Ouyang, Y.D.; Wang, J.; Gong, L.; Zhang, Q.H.; Li, X.H.; Xiao, J.H.; Zhang, Q. Processes underlying a reproductive barrier in indica-japonica rice hybrids revealed by transcriptome analysis. Plant Physiol. 2017, 174, 1683–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mi, J.; Li, G.; Xu, C.; Yang, J.; Yu, H.; Wang, G.; Li, X.; Xiao, J.; Song, H.; Zhang, Q.; et al. Artificial selection in domestication and breeding prevents speciation in rice. Mol. Plant 2020, 13, 650–657. [Google Scholar] [CrossRef]
- Mi, J.; Li, G.; Huang, J.; Yu, H.; Zhou, F.; Zhang, Q.; Ouyang, Y.; Mou, T. Stacking S5-n and f5-n to overcome sterility in indica-japonica hybrid rice. Theor. Appl. Genet. 2016, 129, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Wei, X.; Gao, G.; Jiang, H. Development and evaluation of improved lines based on an elite rice variety 9311 for overcoming hybrid sterility in rice. Mol. Breed. 2020, 40, 102. [Google Scholar] [CrossRef]
- Guo, J.; Li, Y.; Xiong, L.; Yan, T.; Zou, J.; Dai, Z.; Tang, G.; Sun, K.; Luan, X.; Yang, W.; et al. Development of wide-compatible indica lines by pyramiding multiple neutral alleles of indica-japonica hybrid sterility loci. Front. Plant Sci. 2022, 13, 890568. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhao, Z.; Liu, X.; Liu, L.; Jiang, L.; Liu, S.; Zhang, W.; Wang, Y.; Liu, Y.; Wan, J. Marker-assisted breeding of a photoperiod-sensitive male sterile japonica rice with high cross-compatibility with indica rice. Mol. Breed. 2011, 27, 247–258. [Google Scholar] [CrossRef]
- Guo, J.; Xu, X.; Li, W.; Zhu, W.; Zhu, H.; Liu, Z.; Luan, X.; Dai, Z.; Liu, G.; Zhang, Z.; et al. Overcoming inter-subspecific hybrid sterility in rice by developing indica-compatible japonica lines. Sci. Rep. 2016, 6, 26878. [Google Scholar] [CrossRef] [Green Version]
- Afzal, S.; Sirohi, P.; Singh, N.K. A review of CRISPR associated genome engineering: Application, advances and future prospects of genome targeting tool for crop improvement. Biotechnol. Lett. 2020, 42, 1611–1632. [Google Scholar] [CrossRef]
- Gao, C. Genome engineering for crop improvement and future agriculture. Cell 2021, 184, 1621–1635. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.X.; Li, B.B.; Yang, Z.G.; Huang, J.Q.; Sun, W.H.; Bhanbhro, N.; Liu, W.T.; Chen, K.M. Dissecting plant gene functions using CRISPR toolsets for crop improvement. J. Agric. Food Chem. 2022, 70, 7343–7359. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Niu, B.; Long, Y.; Li, G.; Tang, J.; Zhang, Y.; Ren, D.; Liu, Y.G.; Chen, L. Suppression or knockout of SaF/SaM overcomes the Sa-mediated hybrid male sterility in rice. J. Integr. Plant Biol. 2017, 59, 669–679. [Google Scholar] [CrossRef] [Green Version]
- Hiei, Y.; Komari, T.; Kubo, T. Transformation of rice mediated by Agrobacterium tumefaciens. Plant Mol Biol 1997, 35, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.F.; Zhang, H.; Xu, N.F.; Zhang, B.T.; Gou, F.; Zhu, J.K. Application of the CRISPR-Cas system for efficient genome engineering in plants. Mol. Plant 2013, 6, 2008–2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Zhang, Q.; Zhu, Q.; Liu, W.; Chen, Y.; Qiu, R.; Wang, B.; Yang, Z.; Li, H.; Lin, Y.; et al. A robust CRISPR/Cas9 system for convenient, high-efficiency multiplex genome editing in monocot and dicot plants. Mol. Plant 2015, 8, 1274–1284. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Ning, H.; Bennett, J.; Khush, G.S. PCR-based marker-assisted selection in rice breeding. IRRI Discuss. Pap. 1995, 12. [Google Scholar]
- Panaud, O.; Chen, X.; McCouch, S.R. Development of microsatellite markers and characterization of simple sequence length polymorphism (SSLP) in rice (Oryza sativa L.). Mol. Gen. Genet. 1996, 252, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Li, R.Q.; Li, Y.R.; Kristiansen, K.; Wang, J. SOAP: Short oligonucleotide alignment program. Bioinformatics 2008, 24, 713–714. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Abdi, H. The Bonferonni and Šidák Corrections for Multiple Comparisons. Encycl. Meas. Stat. 2007, 3, 2007. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ji, Q.; Zhang, M.; Lu, J.; Wang, H.; Lin, B.; Liu, Q.; Chao, Q.; Zhang, Y.; Liu, C.; Gu, M.; et al. Molecular basis underlying the S5-dependent reproductive isolation and compatibility of indica/japonica rice hybrids. Plant Physiol. 2012, 158, 1319–1328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, S.; Wakasa, Y.; Takahashi, H.; Kawakatsu, T.; Takaiwa, F. Signal transduction by IRE1-mediated splicing of bZIP50 and other stress sensors in the endoplasmic reticulum stress response of rice. Plant J. 2012, 69, 946–956. [Google Scholar] [CrossRef] [PubMed]
- Niu, N.; Liang, W.; Yang, X.; Jin, W.; Wilson, Z.A.; Hu, J.; Zhang, D. EAT1 promotes tapetal cell death by regulating aspartic proteases during male reproductive development in rice. Nat. Commun. 2013, 4, 1445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef]
- Mao, C.; Ding, J.; Zhang, B.; Xi, D.; Ming, F. OsNAC2 positively affects salt-induced cell death and binds to the OsAP37 and OsCOX11 promoters. Plant J. 2018, 94, 454–468. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, S.; Suzuki, Y.; Yoshizawa, R.; Kanno, K.; Makino, A. Effect of individual suppression of RBCS multigene family on Rubisco contents in rice leaves. Plant Cell Environ. 2012, 35, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Makino, A. Availability of rubisco small subunit up-regulates the transcript levels of large subunit for stoichiometric assembly of its holoenzyme in rice. Plant Physiol. 2012, 160, 533–540. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Sun, B.; Shi, Z.; Miao, X.; Li, H. Identification of the rice genes and metabolites involved in dual resistance against brown planthopper and rice blast fungus. Plant Cell Environ. 2022, 45, 1914–1929. [Google Scholar] [CrossRef]
- Reddy, I. Rice flavonoid pathway genes, OsDfr and OsAns, are induced by dehydration, high salt and ABA, and contain stress responsive promoter elements that interact with the transcription activator, OsC1-MYB. Plant Sci. 2004, 166, 1505–1513. [Google Scholar]
- Zhang, B.; Deng, L.; Qian, Q.; Xiong, G.; Zeng, D.; Li, R.; Guo, L.; Li, J.; Zhou, Y. A missense mutation in the transmembrane domain of CESA4 affects protein abundance in the plasma membrane and results in abnormal cell wall biosynthesis in rice. Plant Mol. Biol. 2009, 71, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Qiu, S.Q.; Xu, C.G.; Li, X.H.; Zhang, Q. Genetic dissection of embryo sac fertility, pollen fertility, and their contributions to spikelet fertility of intersubspecific hybrids in rice. Theor. Appl. Genet. 2005, 110, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.W.; He, Y.Q.; Xu, C.G.; Zhang, Q. Identification and confirmation of three neutral alleles conferring wide compatibility in inter-subspecific hybrids of rice (Oryza sativa L.) using near-isogenic lines. Theor. Appl. Genet. 2005, 111, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Li, X.; Wang, Y.; Mi, J.; Xing, F.; Zhang, D.; Dong, Q.; Li, X.; Xiao, J.; Zhang, Q.; et al. Three representative inter and intra-subspecific crosses reveal the genetic architecture of reproductive isolation in rice. Plant J. 2017, 92, 349–362. [Google Scholar] [CrossRef] [Green Version]
- Ikehashi, H.; Araki, H. Genetics of F1 sterility in remote crosses of rice. In Rice Genetics I; IRRI: Manila, Philippines, 1986; pp. 119–130. [Google Scholar]
- Ouyang, Y.; Li, G.; Mi, J.; Xu, C.; Du, H.; Zhang, C.; Xie, W.; Li, X.; Xiao, J.; Song, H.; et al. Origination and establishment of a trigenic reproductive isolation system in rice. Mol. Plant 2016, 9, 1542–1545. [Google Scholar] [CrossRef] [Green Version]
- Kallugudi, J.; Singh, V.J.; Vinod, K.K.; Krishnan, S.G.; Nandakumar, S.; Dixit, B.K.; Ellur, R.K.; Bollinedi, H.; Nagarajan, M.; Kumar, A.; et al. Population dynamics of wide compatibility system and evaluation of intersubspecific hybrids by indica-japonica hybridization in rice. Plants 2022, 11, 1930. [Google Scholar] [CrossRef]
- Priyadarshi, R.; Arremsetty, H.P.S.; Singh, A.K.; Khandekar, D.; Ulaganathan, K.; Shenoy, V.; Sinha, P.; Singh, V.K. Marker-assisted improvement of the elite maintainer line of rice, IR 58025B for wide compatibility (S5(n)) gene. Front. Plant Sci. 2018, 9, 1051. [Google Scholar] [CrossRef] [Green Version]
- Oono, Y.; Wakasa, Y.; Hirose, S.; Yang, L.; Sakuta, C.; Takaiwa, F. Analysis of ER stress in developing rice endosperm accumulating beta-amyloid peptide. Plant Biotechnol. J. 2010, 8, 691–718. [Google Scholar] [CrossRef]
- Ohta, M.; Wakasa, Y.; Takahashi, H.; Hayashi, S.; Kudo, K.; Takaiwa, F. Analysis of rice ER-resident J-proteins reveals diversity and functional differentiation of the ER-resident Hsp70 system in plants. J. Exp. Bot. 2013, 64, 5429–5441. [Google Scholar] [CrossRef] [Green Version]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, J.; Zhou, X.; Chen, K.; Ye, C.; Liu, J.; Sun, K.; Tang, G.; Wang, S.; Zhang, G.; Chen, Y.; et al. Genetic Analysis of S5 Regulating the Hybrid Sterility between Indica and Japonica Subspecies in Rice. Agronomy 2023, 13, 1094. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13041094
Guo J, Zhou X, Chen K, Ye C, Liu J, Sun K, Tang G, Wang S, Zhang G, Chen Y, et al. Genetic Analysis of S5 Regulating the Hybrid Sterility between Indica and Japonica Subspecies in Rice. Agronomy. 2023; 13(4):1094. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13041094
Chicago/Turabian StyleGuo, Jie, Xinqiao Zhou, Ke Chen, Chanjuan Ye, Juan Liu, Kangli Sun, Guang Tang, Shaokui Wang, Guiquan Zhang, Youding Chen, and et al. 2023. "Genetic Analysis of S5 Regulating the Hybrid Sterility between Indica and Japonica Subspecies in Rice" Agronomy 13, no. 4: 1094. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13041094