
Function of Soybean miR159 Family Members in Plant Responses to Low Phosphorus, High Salinity, and Abscisic Acid Treatment

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Extraction of Total RNA, Reverse-Transcription, and Extraction of Genomic DNA
2.3. qRT-PCR Analysis
2.4. Measurement of Fresh Weight, Soluble Phosphate (Pi), and Total P Contents in Soybean Seedlings
2.5. Vector Construction and Soybean Transformation
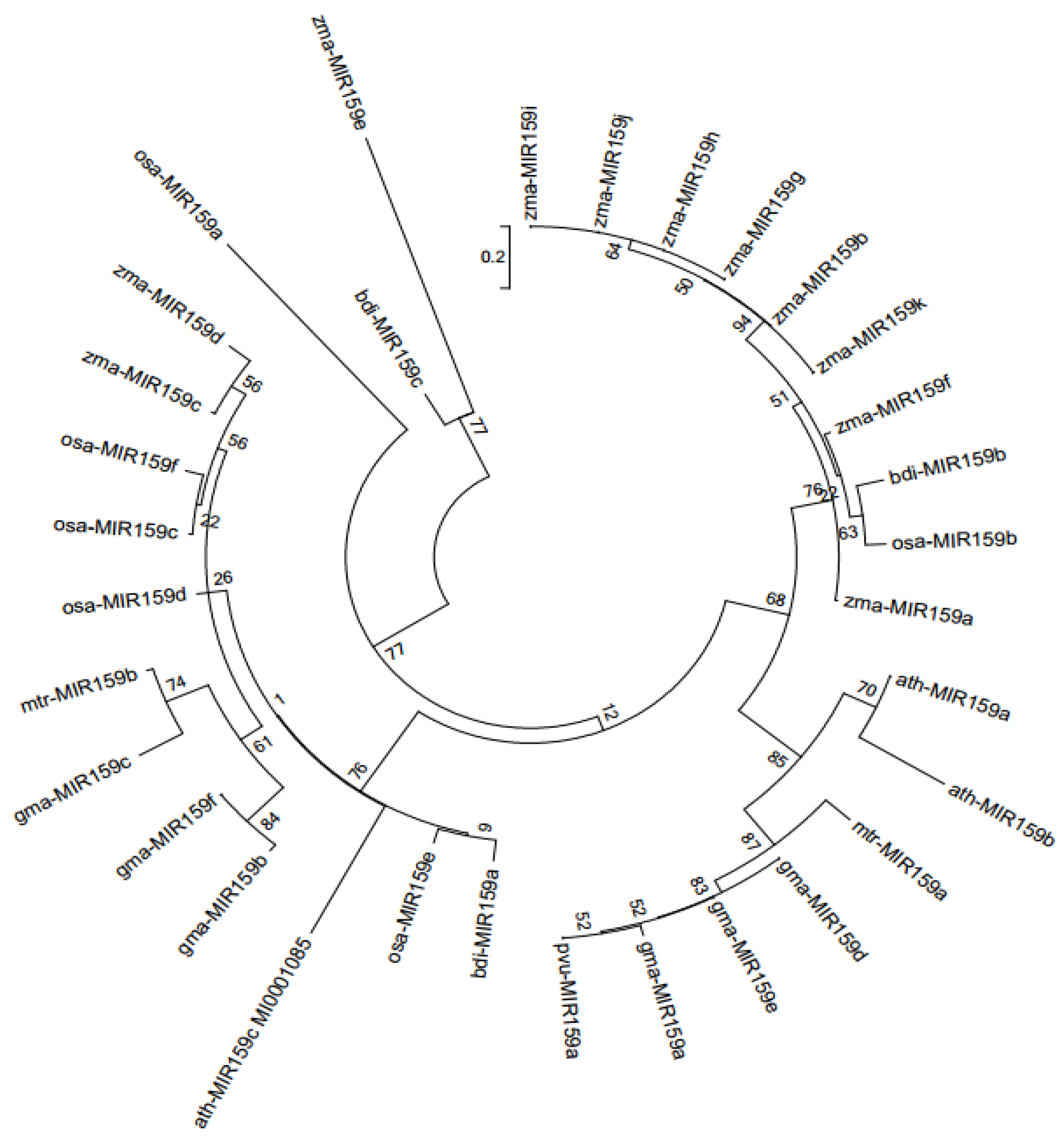
2.6. Phytogenetic Tree Reconstruction and Data Analysis
3. Results
3.1. Identification of MIR159 Gene Family Members in Soybean
3.2. Responses of the Soybean MIR159 Gene Family to Low-P Stress
3.3. Responses of the Soybean MIR159 Gene Family to Salt Stress
3.4. Responses of the Soybean MIR159 Gene Family to ABA
3.5. Overexpressing MIR159e Decrease Total Phosphorus Content in Soybean Leaves under LP Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jonesrhoades, M.W.; Bartel, D.P.; Bartel, B. MicroRNAs and their regulatory roles in plants. Annu. Rev. Plant Biol. 2006, 57, 19–53. [Google Scholar] [CrossRef] [PubMed]
- Rogers, K.; Chen, X. Biogenesis, turnover, and mode of action of plant microRNAs. Plant Cell 2013, 25, 2383–2399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achard, P.; Herr, A.; Baulcombe, D.C.; Harberd, N.P. Modulation of floral development by a gibberellin-regulated microRNA. Development 2004, 131, 3357–3365. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Zhang, X.; Cheng, Y.; Du, X.; Teotia, S.; Miao, C.; Sun, H.; Fan, G.; Tang, G.; Xue, H.; et al. The miR167-OsARF12 module regulates rice grain filling and grain size downstream of miR159. Plant Commun. 2023, 4, 100604. [Google Scholar] [CrossRef]
- Lei, P.; Qi, N.; Zhou, Y.; Wang, Y.; Zhu, X.; Xuan, Y.; Liu, X.; Fan, H.; Chen, L.; Duan, Y. Soybean miR159-GmMYB33 Regulatory network involved in gibberellin-modulated resistance to Heterodera glycines. Int J Mol Sci. 2021, 22, 13172. [Google Scholar] [CrossRef]
- Wang, M.; Wang, Q.; Zhang, B. Response of miRNAs and their targets to salt and drought stresses in cotton (Gossypium hirsutum L.). Gene 2013, 530, 26–32. [Google Scholar] [CrossRef]
- Zhao, P.; Wang, F.; Deng, Y.; Zhong, F.; Tian, P.; Lin, D.; Deng, J.; Zhang, Y.; Huang, T. Sly-miR159 regulates fruit morphology by modulating GA biosynthesis in tomato. Plant Biotechnol. J. 2022, 20, 833–845. [Google Scholar] [CrossRef] [PubMed]
- Kitazumi, A.; Kawahara, Y.; Onda, T.S.; De Koeyer, D.; de los Reyes, B.G. Implications of miR166 and miR159 induction to the basal response mechanisms of an andigena potato (Solanum tuberosum subsp. andigena). Genome 2015, 58, 13–24. [Google Scholar] [CrossRef]
- Csukasi, F.; Donaire, L.; Casañal, A.; Martínez-Priego, L.; Botella, M.A.; Medina-Escobar, N.; Llave, C.; Valpuesta, V. Two strawberry miR159 family members display developmental-specific expression patterns in the fruit receptacle and cooperatively regulate Fa-GAMYB. New Phytol. 2012, 195, 47–57. [Google Scholar] [CrossRef]
- Jiang, J.; Xu, P.; Li, Y.; Li, Y.; Zhou, X.; Jiang, M.; Zhang, J.; Zhu, J.; Wang, W.; Yang, L. Identification of miRNAs and their target genes in genic male sterility lines in Brassica napus by small RNA sequencing. BMC Plant Biol. 2021, 9, 520. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, F.; Cao, H.; Peng, H.; Ni, Z.; Sun, Q.; Yao, Y. TamiR159 directed wheat TaGAMYB cleavage and its involvement in anther development and heat response. PLoS ONE 2012, 7, e48445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garighan, J.; Dvorak, E.; Estevan, J.; Loridon, K.; Huettel, B.; Sarah, G.; Farrera, I.; Leclercq, J.; Grynberg, P.; Coiti Togawa, R.; et al. The identification of small RNAs differentially expressed in apple buds reveals a potential role of the Mir159-MYB regulatory module during dormancy. Plants 2021, 3, 2665. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Zhang, Q.; Zhao, Y.Q.; Yang, J.; He, H.B.; Jia, G.X. The lre-miR159a-LrGAMYB pathway mediates resistance to grey mould infection in Lilium regale. Mol. Plant Pathol. 2020, 21, 749–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Z.; Wang, N.; Jalajakumari, M.; Blackman, L.; Shen, E.; Verma, S.; Wang, M.B.; Millar, A.A. miR159 represses a constitutive pathogen defense response in tobacco. Plant Physiol. 2020, 182, 2182–2198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millar, A.A.; Lohe, A.; Wong, G. Biology and function of miR159 in plants. Plants 2019, 8, 255. [Google Scholar] [CrossRef] [Green Version]
- Reyes, J.L.; Chua, N. ABA induction of miR159 controls transcript levels of two MYB factors during Arabidopsis seed germination. Plant J. 2007, 49, 592–606. [Google Scholar] [CrossRef]
- Millar, A.A.; Gubler, F. The Arabidopsis GAMYB-Like genes, MYB33 and MYB65, are microRNA-regulated genes that redundantly facilitate anther development. Plant Cell 2005, 17, 705–721. [Google Scholar] [CrossRef] [Green Version]
- Allen, R.S.; Li, J.; Stahle, M.I. Genetic analysis reveals functional redundancy and the major target genes of the Arabidopsis miR159 family. Proc. Natl. Acad. Sci. USA 2007, 104, 16371–16376. [Google Scholar] [CrossRef]
- Xue, T.; Liu, Z.; Dai, X.; Xiang, F. Primary root growth in Arabidopsis thaliana is inhibited by the miR159 mediated repression of MYB33, MYB65 and MYB101. Plant Sci. 2017, 262, 182–189. [Google Scholar] [CrossRef]
- Mishra, S.; Chaudhary, R.; Sharma, P. Temporal expression analysis of microRNAs and their target GRAS genes induced by osmotic stress in two contrasting wheat genotypes. Mol Biol Rep. 2023, 13, 1–13. [Google Scholar] [CrossRef]
- Zeng, H.Q.; Zhu, Y.Y.; Huang, S.Q.; Yang, Z.M. Analysis of phosphorus-deficient responsive miRNAs and cis-elements from soybean (Glycine max L.). J. Plant Physiol. 2010, 167, 1289–1297. [Google Scholar] [CrossRef]
- Zhao, M.; Ding, H.; Zhu, J.K.; Zhang, F.; Li, W.X. Involvement of miR169 in the nitrogen-starvation responses in Arabidopsis. New Phytol. 2011, 190, 906–915. [Google Scholar] [CrossRef] [Green Version]
- Vidal, E.A.; Araus, V.; Lu, C.; Parry, G.; Green, P.J.; Coruzzi, G.M.; Gutiérrez, R.A. Nitrate-responsive miR393/AFB3 regulatory module controls root system architecture in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 4477–4482. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, P.; Liu, Q.; Li, G.; Di, D.; Xia, G.; Kronzucker, H.J.; Fang, S.; Chu, J.; Shi, W. TaANR1-TaBG1 and TaWabi5-TaNRT2s/NARs link ABA metabolism and nitrate acquisition in wheat roots. Plant Physiol. 2020, 182, 1440–1453. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Jiang, Z.; Wang, W.; Qiu, Y.; Zhang, Z.; Liu, Y.; Li, A.; Gao, X.; Liu, L.; Qian, Y.; et al. Nitrate-NRT1.1B-SPX4 cascade integrates nitrogen and phosphorus signalling networks in plants. Nat. Plants. 2019, 5, 401–413. [Google Scholar] [CrossRef]
- Zhang, T.T.; Yan, C.X.; Zhao, X.B.; Li, C.J.; Shi, C.R.; Yu, J.J.; Chan, S.H.; Li, R.G. Identification and functional characterization of salt tolerance related microRNAs in roots of peanut (Arachis hypogaea L.). J. Peanut Sci. 2016, 45, 36–42, (In Chinese with English Abstract). [Google Scholar]
- Tang, M.N.; Liu, Q.; Zhang, Y. Response of salt-tolerant miRNAs to high-salt habitats in mangrove companion plant hippocampal teeth. MPB 2017, 15, 1137–1142, (In Chinese with English Abstract). [Google Scholar]
- Lu, C.; Fedoroff, N. A Mutation in the Arabidopsis HYL1 gene encoding a dsRNA binding protein affects responses to abscisic acid, auxin, and cytokinin. Plant Cell 2000, 12, 2351–2366. [Google Scholar] [CrossRef] [Green Version]
- López-Galiano, M.J.; García-Robles, I.; González-Hernández, A.I.; Camañes, G.; Vicedo, B.; Real, M.D.; Rausell, C. Expression of mir159 is altered in tomato plants undergoing drought stress. Plants 2019, 8, 201. [Google Scholar] [CrossRef] [Green Version]
- AAbe, H.; Urao, T.; Ito, T.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) Function as Transcriptional Activators in Abscisic Acid Signaling. Plant Cell 2003, 15, 63–78. [Google Scholar] [CrossRef] [Green Version]
- Huo, W.G.; Li, B.D.; Kuang, J.B. Functional characterization of the steroid reductase genes GmDET2 and GmDET2b from Glycine max. Int J Mol Sci. 2018, 19, 726. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Liu, Q.; Chen, L.; Kuang, J.B.; Liao, H. Genome-wide identification of soybean microRNAs and their targets reveals their organ-specificity and responses to phosphate starvation. BMC Genom. 2013, 14, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Jiao, F.; Wu, Z.; Li, Y.; Wang, X.; He, X.; Zhong, W.; Wu, P. OsPHR2 is involved in phosphate-starvation signaling and excessive phosphate accumulation in shoots of plants. Plant Physiol. 2008, 146, 1673–1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wang, Y.; Tian, J.; Lim, B.L.; Yan, X.; Liao, H. Overexpressing AtPAP15 enhances phosphorus efficiency in soybean. Plant Physiol. 2009, 151, 233–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Peterson, D.; Tamura, K. MEGA-CC: Computing core of molecular evolutionary genetics analysis program for automated and iterative data analysis. Bioinformatics 2012, 28, 2685–2686. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server. Nucleic Acids Res. 2011, 39, W155–W159. [Google Scholar] [CrossRef] [Green Version]
- Figueredo, M.S.; Formey, D.; Rodriguez, J.; Ibanez, F.; Hernandez, G.; Fabra, A. Identification of miRNAs linked to peanut nodule functional processes. J Biosci. 2020, 45, 62. [Google Scholar] [CrossRef]
- Khraiwesh, B.; Zhu, J.K.; Zhu, J. Role of miRNAs and siRNAs in biotic and abiotic stress responses of plants. Biochim. Biophys. Acta (BBA)-Gene Regul. Mech. 2012, 1819, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Jin, Y.X. Evolution of MIR159/319 microRNA genes and their post-transcriptional regulatory link to siRNA pathways. BMC Evol. Biol. 2011, 11, 122. [Google Scholar] [CrossRef] [Green Version]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.X.; Mitros, T.; Nelson, W.; Hyten, D. Genome sequence of the paleopolyploid soybean (Glycine max (L.) Merr.). Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [Green Version]
- Bustos, R.; Castrillo, G.; Linhares, F.; Puga, M.I.; Rubio, V.; P’erez-Perez, J.; Solano, R.; Leyva, A.; Paz-Ares, J. A central regulatory system largely controls transcriptional activation and repression responses to phosphate starvation in Arabidopsis. PLoS Genet 2010, 6, e1001102. [Google Scholar] [CrossRef] [Green Version]
- Bian, X.Y.; Song, D.F.; Han, S.Y.; Wang, J.H.; Zhu, M.Y. Flowering time control in ornamental gloxinia (Sinningia speciosa) by manipulation of miR159 expression. Ann. Bot. 2013, 111, 791–799. [Google Scholar]
- Nord, E.A.; Lynch, J.P. Delayed reproduction in Arabidopsis thaliana improves fitness in soil with suboptimal phosphorus availability. Plant Cell Environ. 2008, 31, 1432–1441. [Google Scholar] [CrossRef]
- Olivera, M.; Tejera, N.; Iribarne, C.; Ocana, A.; Lluch, C. Growth, nitrogen fixation and ammonium assimilation in common bean (Phaseolus vulgaris): Effect of phosphorus. Physiol Plant. 2004, 121, 498–505. [Google Scholar] [CrossRef]
- Schulze, J.; Temple, G.; Temple, S.J.; Beschow, H.; Vance, C.P. Nitrogen fixation by white lupin under phosphorus deficiency. Ann. Bot. 2006, 98, 731–740. [Google Scholar] [CrossRef] [Green Version]
- Pokoo, R.; Ren, S.; Wang, Q.; Motes, C.M.; Hernandez, T.D.; Ahmadi, S.; Monteros, M.J.; Zheng, Y.; Sunkar, R. Genotype- and tissue-specific miRNA profiles and their targets in three alfalfa (Medicago sativa L.) genotypes. BMC Genom. 2018, 19, 913. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.K.; Sunkar, R. Nutrient- and other stress-responsive microRNAs in plants: Role for thiol-based redox signaling. Plant Signal. Behav. 2015, 10, e1010916. [Google Scholar] [PubMed]
- Zucchero, J.C.; Caspi, M.; Dunn, K. ngl9: A third MADS box gene expressed in alfalfa root nodules. Mol. Plant-Microbe Interact. 2001, 14, 1463–1467. [Google Scholar] [CrossRef] [Green Version]
- Mega, R.; Abe, F.; Kim, J.S.; Tsuboi, Y.; Tanaka, K.; Kobayashi, H.; Sakata, Y.; Hanada, K.; Tsujimoto, H.; Kikuchi, J.; et al. Tuning water-use efficiency and drought tolerance in wheat using abscisic acid receptors. Nat. Plants 2019, 5, 153–159. [Google Scholar] [CrossRef]
- Jiang, Y.; Wu, X.; Shi, M.; Yu, J.; Guo, C. The miR159-MYB33-ABI5 module regulates seed germination in Arabidopsis. Physiol. Plantarum. 2022, 174, e13659. [Google Scholar] [CrossRef]
- Manish, T. Blessing in disguise: A loss of miR159 makes plant drought tolerant and ABA sensitive. Physiol. Plantarum. 2022, 174, e13763. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Mature miRNA Sequence | Position on Chromosome |
|---|---|---|
| MIR159a | GAGCUCCUUGAAGUCCAAUUG | Gm09: 40,266,722–40,266,935 + |
| MIR159b | GAGUUCCCUGCACUCCAAGUC | Gm07: 5,424,789–5,424,974 − |
| MIR159c | AUUGGAGUGAAGGGAGCUCCG | Gm16: 2,830,034–2,830,218 − |
| MIR159d | AGCUGCUUAGCUAUGGAUCCC | Gm09: 40,267,077–40,267,097 + |
| MIR159e | GAGCUCCUUGAAGUCCAAUU | Gm07: 9,561,934–9,562,144 − |
| MIR159f | GAGUUCCCUGCACUCCAAGUC | Gm16: 2,819,636–2,819,815 − |
| Low-P Responsive Elements | NaCl-Responsive Elements | ABA-Responsive Elements | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TATA -Box | TATA -Box Like | W-box | PHR1 Element | ABRE-Like | ACGT Sequence | rd22 | AtMYB2 | MYC2 | ABRE | DPBF | RY Elements | |
| MIR159a | 4 | 1 | 4 | 1 | 1 | 1 | 2 | 1 | 1 | 1 | ||
| MIR159b | 4 | 1 | 3 | 1 | 1 | 1 | 1 | 1 | 1 | |||
| MIR159c | 4 | 1 | 4 | 1 | 1 | 1 | 2 | 2 | 1 | |||
| MIR159d | 4 | 1 | 4 | 1 | 1 | 1 | 3 | 1 | 1 | 1 | ||
| MIR159e | 4 | 4 | 1 | 1 | 1 | 4 | 3 | 1 | 1 | 1 | ||
| MIR159f | 4 | 1 | 3 | 1 | 1 | 1 | 1 | 1 | 1 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, B.; Tao, P.; Xu, F.; He, P.; Wang, J. Function of Soybean miR159 Family Members in Plant Responses to Low Phosphorus, High Salinity, and Abscisic Acid Treatment. Agronomy 2023, 13, 1798. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13071798
Li B, Tao P, Xu F, He P, Wang J. Function of Soybean miR159 Family Members in Plant Responses to Low Phosphorus, High Salinity, and Abscisic Acid Treatment. Agronomy. 2023; 13(7):1798. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13071798
Chicago/Turabian StyleLi, Bodi, Ping Tao, Feng Xu, Pingan He, and Jinxiang Wang. 2023. "Function of Soybean miR159 Family Members in Plant Responses to Low Phosphorus, High Salinity, and Abscisic Acid Treatment" Agronomy 13, no. 7: 1798. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13071798