
Boron Nutrition in Coffee Improves Drought Stress Resistance and, Together with Calcium, Improves Long-Term Productivity and Seed Composition


Abstract
:1. Introduction
2. Materials and Methods
2.1. Greenhouse Trial
2.2. Field Trial
3. Results
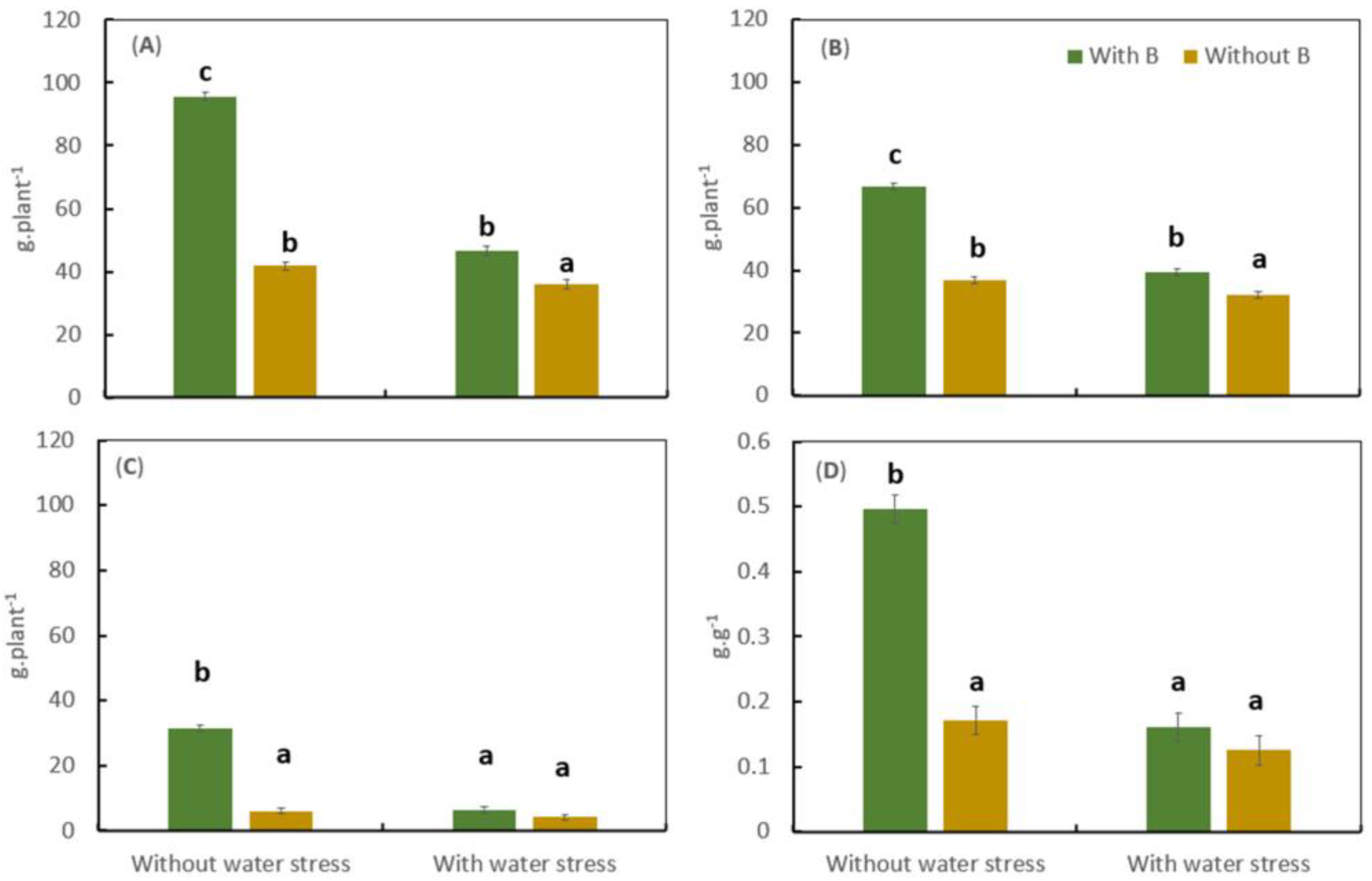
3.1. Effect of Water Stress and Boron on Biomass Accumulation and Boron and Nitrogen Uptake under Controlled Conditions
3.2. B Influence on the Coffee Yield under Field Conditions
3.3. B in the Soil after Five Years of Application in the Field Trial
3.4. B and the Biochemical Composition of the Green Coffee Beans Produced under Field Conditions
4. Discussion
4.1. Influence of the Iteraction of Water and B Deficits on the Biomass Accumulation, B and N Uptake under Controlled Conditions
4.2. Influence of B on Coffee Productivity
4.3. B and the Biochemical Composition of the Green Coffee Beans under Field Conditions
5. Conclusions
- −
- Boron deficits significantly reduce the growth of the young coffee plants, influencing mainly the shoot biomass accumulation and clearly indicating the strong susceptibility of the coffee plants to B deficit.
- −
- Coffee plants are sensitive to long water stress periods, and B application significantly reduces the negative effect of this abiotic stress.
- −
- Under water stress conditions, it was possible to observe B remobilization in the phloem from mature to young leaves.
- −
- During a whole production cycle of five years, the mean application of 77 kg CaO and 0.6 kg B.ha−1.year−1 yielded 14% more than the control without B and Ca2+, while the increase in the mean B rate to 1.1 kg.ha−1.year−1 at the same Ca2+ rate increased the productivity by almost 37% compared to the control, indicating a long-term additive effect between B and Ca2+.
- −
- Unsaturated fatty acid and caffeine contents in the green coffee beans were good indicators of the stress conditions (water stress) under field conditions. These compounds were negatively correlated with the B content, indicating that B could reduce the formation of these compounds, which are considered as negative descriptors of coffee cup quality.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Samper, F.L.; Giovannucci, D.; Marques, V.L. The Powerful Role of Intangibles in the Coffee Value Chain; Economic Research Working Paper; World Intellectual Property Organization-WIPO: Geneva, Switzerland, 2017; No. 39; 79p. [Google Scholar]
- Cordes, Y.K.; Sagan, M.; Kennedy, S. Responsible Coffee Sourcing: Towards a Living Income for Producers; Columbia Center on Sustainable Investment, Columbia Law School and Earth Institute, Columbia University: New York, NY, USA, 2021; 72p. [Google Scholar]
- International Coffee Organization-ICO. Coffee Development Report. 2019. 98p. Available online: https://www.internationalcoffeecouncil.com/_files/ugd/0dd08e_b2c2768ae87045e383962ce14ef44925.pdf (accessed on 2 February 2023).
- DaMatta, M.F.; Ramalho, J.D.C. Impacts of drought and temperature stress on coffee physiology and production: A Review. Braz. J. Plant Physiol. 2006, 18, 55–81. [Google Scholar] [CrossRef]
- Ramirez, B.V.H.; Peña, Q.A.; Jaramillo, R.A.; Giraldo, E.; Suarez, A.; Duque, R. Agroclimatic risk for the Colombian coffee zone: Method to regionalize the climate variability. Cenicafé 2012, 63, 98–115. (In Spanish) [Google Scholar]
- Ramirez, B.V.H.; Jaramillo, R.A.; Arcila, P.J. The climate factors that influence the coffee crop productivity in Colombia. In Manual del Cafetero Colombiano-Cenicafé; Federación Nacional de Cafeteros de Colombia: Manizales, Colombia, 2013; Volume 1, pp. 205–238. (In Spanish) [Google Scholar]
- Ramalho, J.C.; Rodrigues, A.P.; Lidon, F.C.; Marques, L.M.C.; Leitao, A.E.; Fortunato, A.S.; Pais, J.P.; Silva, M.J.; Scotti-Campos, P.; Lopes, A.; et al. Stress cross-response of the antioxidative system promoted by superimposed drought and cold conditions in Coffee spp. PLoS ONE 2018, 16, e0198694. [Google Scholar] [CrossRef]
- Leon-Burgos, A.F.; Unigarro, C.; Balaguera-López, H.E. Can prolonged conditions of water deficit alter photosynthetic performance and water relations of coffee plants in central-west Colombia. S. Afr. J. Bot. 2022, 149, 366–375. [Google Scholar] [CrossRef]
- Bunn, C.; Läderach, P.; Ovalle, R.O.; Kirschke, D. A bitter cup: Climate change profile of global production of Arabica and Robusta coffee. Clim. Chang. 2015, 129, 89–101. [Google Scholar] [CrossRef]
- Grüter, R.; Traschel, T.; Laube, P.; Jaisli, I. Expected global suitability of coffee, cashew and avocado due to climate change. PLoS ONE 2022, 17, e0261976. [Google Scholar] [CrossRef] [PubMed]
- Gupta, U.C. Introduction. In Boron and Its Role in Crops Production; Gupta, U.C., Ed.; CRC Press: Boca Raton, FL, USA, 1993; 256p. [Google Scholar]
- Welch, R.M.; Allaway, W.H.; House, W.A.; Kubota, J. Geographic distribution of trace elements problems. In Micronutrients in Agriculture, 2nd ed.; Montvelt, J.J., Cox, F.R., Shuman, L.M., Welch, R.M., Eds.; SSSA Book Series 4; SSSA: Madision, WI, USA, 1991; pp. 31–57. [Google Scholar]
- Sarkar, D.; Mandal, B.; Mazumdar, D. Plant availability of boron in acid soils as assessed by different extractants. J. Plant Nutr. Soil Sci. 2008, 171, 249–254. [Google Scholar] [CrossRef]
- Antoniadis, V.; Chatzissavvidis, C.; Paparnakis, A. Boron behavior in apple plants in acidic and limed soil. J. Plant Nutr. Soil Sci. 2013, 176, 267–272. [Google Scholar] [CrossRef]
- Brown, H.P.; Shelp, J.B. Boron mobility in plants. Plant Soil 1997, 193, 85–101. [Google Scholar] [CrossRef]
- Malavolta, E.; Lima-Filho, O.F. Mineral nutrition and fertilization of traditional, high plant density irrigated and organic coffee plantation. In I Simposio de Pesquisa dos Cafés do Brasil; Federal University of Lavras: Lavras, MG, Brazil, 2000; pp. 331–353. (In Portuguese) [Google Scholar]
- Ramirez, F.; Bertsch, F.; Mora, L. Nutrients consumption by fruits and branches of coffee during development and maturation in Aquiares, Turrialba, Costa Rica. Agron. Costaricence 2002, 26, 33–42. (In Spanish) [Google Scholar]
- Laviola, B.G.; Martines, H.E.P.; Salomão, L.C.C.; Cruz, C.D.; Mendoza, S.M. Nutrients accumulation by the coffee beans in two elevations: Micronutrients. Rev. Bras. Ciênc. Solo 2007, 31, 1439–1449. (In Portuguese) [Google Scholar] [CrossRef]
- Sadeghian, K.S.; Salamanca, J.A. Micronutrients in coffee fruits and leaves. Cenicafé 2015, 66, 73–87. (In Spanish) [Google Scholar]
- Lima-Filho, O.F.; Malavolta, E. Evaluation of extraction procedure on determination of critical soil and foliar level of boron and zinc in coffee plants. Comm. Soil Sci. Plant Anal. 1998, 29, 825–833. [Google Scholar] [CrossRef]
- Malavolta, E. Manual of Plant Nutrition; Editorial Agronomica Ceres Ltda: Sao Paulo, Brasil, 2006; 631p. (In Portuguese) [Google Scholar]
- Favarin, J.L.; Teles de Souza, L.; Mazzafera, P.; Dimenstein, L. Soil correction and fertilization of irrigated coffee trees in production. In Cafeicultura do Cerrado; Carvalho, G.R., Ferreira, A.D., Andrade, V.T., Botelho, C.E., Carvalho, J.P.F., Eds.; EPAMIG: Belo Horizonte, Brazil, 2021; 564p. (In Portuguese) [Google Scholar]
- Gonzales, F.A. Why boron is an essential element for vascular plants. New Phytol. 2020, 226, 1228–1230. [Google Scholar] [CrossRef] [PubMed]
- Brown, H.P.; Bellaloui, N.; Wimmer, A.M.; Bassil, E.S.; Ruiz, J.; Hu, H.; Pfeffer, H.; Dannel, F.; Römheld, V. Boron in plant biology. Plant Biol. 2002, 4, 205–223. [Google Scholar] [CrossRef]
- Goldbach, E.H.; Wimmer, A.M. Boron in plants and animals: Is there a role beyond cell-wall structure? J. Plant. Nutr. Soil Sci. 2007, 170, 39–48. [Google Scholar] [CrossRef]
- Huang, L.; Ye, S.; Bell, W.R.; Dell, B. Boron nutrition and chilling tolerance of warm climate crop species. Ann. Bot. 2005, 96, 755–767. [Google Scholar] [CrossRef]
- Golbach, H.E.; Wimmer, M.A.; Chaumont, F.; Matoh, T.; Volkmann, D.; Baluska, F. Rapid response of plants to boron deprivation. In Boron in Plant and Animal Nutrition; Goldbach, H.E., Rerkasem, B., Wimmer, M., Brown, P.H., Thellier, M., Bell, R.W., Eds.; Kluwer Academic: New York, NY, USA, 2002; pp. 167–180. [Google Scholar]
- Ahanger, A.M.; Taleb, M.N.; Allah, A.E.F.; Ahmad, P.; Hajiboland, R. Plant growth under drought stress: Significance of mineral nutrients. In Water Stress and Crop Plants: A Sustainable Approach; Ahmad, P., Ed.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2016; Volume 2, pp. 649–667. [Google Scholar]
- Reguera, M.; Camacho-Cristóbal, J.J. Molecular, Metabolic and Physiological Responses to Boron Stress in Higher Plants. Plants 2023, 12, 2136. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Smith, M.S.; Shabala, S.; Yu, M. Phytohormones in plant responses to boron deficiency and toxicity. J. Exp. Bot. 2023, 74, 743–754. [Google Scholar] [CrossRef]
- Lima de Souza, T.; Pádua, O.D.; Ferreira, S.C.; Pereira, R.T.H.; Campos, C.J.P.; da Silva Resende, R.E.; Fernandes, T.J.; de Souza, T.R.; Ramirez, B.V.; Guelfi, D. Nitrogen fertilizer technologies: Opportunities to improve nutrient use efficiency towards sustainable coffee production systems. Agric. Ecosyst. Environ. 2023, 345, 108317. [Google Scholar] [CrossRef]
- Rosolem, C.A.; Leite, M.V. Coffee leaf and stem anatomy under boron deficiency. R. Bras. Ciênc. Solo 2007, 31, 477–483. [Google Scholar] [CrossRef]
- Malavolta, E.; Fernandes, D.R.; Casale, H.; Peres, R.J. Be the doctor of your coffee plantation. In Informaciones Agronomicas; Brazilian Association for Potash and Phosphorus: Piracicaba, Brazil, 1993; Volume 64, 36p. (In Portuguese) [Google Scholar]
- Nyomora, A.M.S.; Brown, P.H.; Pinney, K.; Polito, V.S. Foliar Application of Boron to Almond Trees Affects Pollen Quality. J. Am. Soc. Hort. Sci. 2000, 125, 265–270. [Google Scholar] [CrossRef]
- Ramirez, B.V.H.; Küsters, J.; de Souza, T.R.; Simmes, C. Calcium nutrition in coffee and its influences on growth, stress tolerance, cations uptake, and productivity. Front. Agron. 2020, 2, 590892. [Google Scholar] [CrossRef]
- Arcila, P.J.; Buhr, L.; Bleiholder, H.; Hack, H.; Meier, U.; Wicke, H. Application of the extended BBCH scale for the description of the growing stages of the coffee (Coffea spp.). Ann. Appl. Biol. 2002, 141, 19–27. [Google Scholar] [CrossRef]
- Baumgarten, A.; Dachler, M. The assessment of nutrient availability in soils using the CaCl2/DTPA (CAT) extraction method. Acta Hortic. 2000, 511, 35–42. [Google Scholar] [CrossRef]
- González, O.H.; Salamanca, J.A. Representative Soil Units of the Colombian Coffee Zone; Chinchiná Cenicafé: Caldas, Colombia, 2008; 25p. (In Spanish) [Google Scholar]
- Ramirez, B.V.H. Coffee phenology is a useful tool to support decision-making. Av. Técnicos Cenicafé 2014, 441, 1–8. (In Spanish) [Google Scholar]
- Puerta, Q.G.I.; Echeverry, M.J.G. Controlled coffee fermentation: Technology to add value to the quality. Av. Técnicos Cenicafé 2015, 454, 1–12. (In Spanish) [Google Scholar] [CrossRef]
- Ashoor, H.S.; Seperich, J.G.; Woodrow, C.; Monte, C.; Welty, J. High-Performance liquid chromatography determination of caffeine in decaffeinated coffee, tea, and beverage Products. J. Assoc. Off. Anal. Chem. 1983, 66, 606–609. [Google Scholar] [PubMed]
- Villarreal, D.; Laffargue, A.; Posada, H.; Bertands, B.; Lashermes, P.; Dussert, S. Genotypic and environmental effects on Coffee (Coffea arabica L.) bean fatty acid profile: Impact on variety and origen chemometric determination. J. Agric. Food Chem. 2009, 57, 11321–11327. [Google Scholar] [CrossRef]
- Villarreal, D.; Baena, L.; Posada, H. Lipids and Fatty acids analysis in green coffee beans in advanced lines of Coffea arabica planted in Colombia. Cenicafé 2012, 63, 19–40. (In Spanish) [Google Scholar]
- Ramirez, B.V.H.; Küsters, J. Calcium and potassium nutrition increase the water use efficiency in coffee: A promising strategy to adapt to climate change. Hydrology 2021, 8, 75. [Google Scholar] [CrossRef]
- Brown, H.P.; Hu, H. Phloem mobility of boron in species dependent. Evidences for phloem mobility in sorbitol-rich species. Ann. Bot. 1996, 77, 497–505. [Google Scholar] [CrossRef]
- Leite, M.V.; Brown, H.P.; Rosolem, A.C. Boron translocation in coffee tress. Plant Soil 2007, 290, 221–229. [Google Scholar] [CrossRef]
- Du, W.; Pan, Z.Y.; Hussain, S.B.; Han, Z.X.; Peng, S.A.; Liu, Y.Z. Foliar supplied boron can be transported to roots as a boron-sucrose complex via phloem in citrus trees. Front. Plant Sci. 2020, 11, 250. [Google Scholar] [CrossRef] [PubMed]
- Brown, H.P.; Hu, H. Phloem boron mobility in diverse plant species. Bot. Acta 1998, 111, 331–335. [Google Scholar] [CrossRef]
- Noiraud, N.; Maurousset, L.; Lemonie, R. Transport of polyols in higher plants. Plant Phys. Bio. 2001, 39, 717–728. [Google Scholar] [CrossRef]
- Olivera-Silva, S.L.; Prado, R.d.M.; Abreu, C.H., Jr.; Silva, G.P.d.; Silva, G.B.d., Jr.; Silva, J.L.F.d. 10Boron is mobile in cowpea plants. Front. Plant Sci. 2021, 12, 717219. [Google Scholar] [CrossRef]
- Balleto, S.A.C.; Menegario, A.A.; Giné, M.F. Boron isotope dilution in cellular fractions of coffee leaves evaluated by inductively coupled plasma mass spectrometry with direct injection nebulization (DIN-ICP-MS). J. Braz. Chem. Soc. 2003, 14, 269–273. [Google Scholar] [CrossRef]
- Boaretto, R.M.; Quaggio, J.A.; Mourao Filho, F.A.A.; Giné, M.F.; Boaretto, A.E. Absorption and mobility of boton in young citrus plants. Commun. Soil Sci. Plant Anal. 2008, 39, 2501–2514. [Google Scholar] [CrossRef]
- Boaretto, R.M.; Quaggio, J.A.; Mattos, D., Jr.; Muraoka, T.; Boaretto, A.E. Boron Uptake and Distribution in Field Grown Citrus Trees. J. Plant Nutr. 2011, 34, 839–849. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, Z.; Wang, S.; Shi, L.; Xu, F. Genotype differences in the synergy effect of nitrogen and boron on the seed yield and nitrogen uses efficiency of Brasicca napus. J. Sci. Food Agric. 2021, 102, 3563–3571. [Google Scholar] [CrossRef] [PubMed]
- Karim, M.; Zhang, Y.Q.; Zhao, R.R.; Chen, X.P.; Zhang, F.S.; Zou, C.Q. Alleviation of drought stress in winter wheat by late foliar application of zinc, boron, and manganese. J. Plant Nutr. Soil Sci. 2012, 175, 142–151. [Google Scholar] [CrossRef]
- Cakmak, I.; Römheld, V. Boron deficiency-induced impairments of cellular functions in plants. Plant Soil 1997, 193, 71–83. [Google Scholar] [CrossRef]
- Yan, L.; Du, C.; Riaz, M.; Jiang, C. Boron mitigates citrus root injuries by regulating intracellular pH and reactive oxygen species to resist H+-toxicity. Environ. Pollut. 2019, 255, 113254. [Google Scholar] [CrossRef] [PubMed]
- DaMatta, M.F.; Ronchi, P.C.; Maestri, M.; Barros, S.R. Ecophysiology of coffee growth and production: A Review. Braz. J. Plant Physiol. 2007, 19, 485–510. [Google Scholar] [CrossRef]
- Santinato, F.; de Oliveira, T.T.; de Mello, P.R.; Caione, G.; de Albuquerque, S.V.; Santinato, R. Boron doses applied to soil during coffee development. Comun. Sci. 2016, 7, 49–55. [Google Scholar] [CrossRef]
- Trenberth, E.K. The Definition of El Niño. Bull. Am. Meteorol. Soc. 1997, 78, 2771–2777. [Google Scholar] [CrossRef]
- National Oceanic and Atmospheric Administration-NOAA/National Weather Service/Climate Prediction Center. ONI Data Base. Available online: https://origin.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ONI_v5.php (accessed on 1 January 2023).
- Ramírez, B.V.H.; Arcila, P.J.; Jaramillo, R.A.; Rendón, S.J.S.; Cuesta, G.G.; Menza, F.H.D.; Mejia, M.C.G.; Montoya, D.F.; Mejia, M.J.W.; Torres, N.J.C.; et al. The Coffee crop flowering and its relationship with water, thermal, and energy availability. Cenicafé 2010, 61, 132–158. (In Spanish) [Google Scholar]
- Peña, Q.A.; Ramirez, B.V.H.; Jaramillo, R.A.; Rendon, S.J.R.; Arcila, P.J. Effects of the daylength and soil humidity on the Flowering of Coffea arabica L. in Colombia. Rev. Fac. Nal. Agr. Medellin 2011, 64, 5745–5754. [Google Scholar]
- Sheng, O.; Zhou, G.; Wie, Q.; Peng, S.; Deng, X. Effects of excess boron on growth, gas exchange, and boron status of four orange scion-rootstock combinations. J. Plant Nutr. Soil Sci. 2010, 173, 469–476. [Google Scholar] [CrossRef]
- Da Silva, C.R.; Baird, R.; Degryse, F.; McLaughlin, J.M. Slow and fast-release boron sources in potash fertilizers: Spatial variability, nutrient dissolution, and plant uptake. Soil Sci. Soc. Am. J. 2018, 82, 1437–1448. [Google Scholar] [CrossRef]
- Terraza, P.M.F.; Summer, E.M.; Cabrera, L.M.; Thomson, A. Boron adsorption and desorption on volcanic ash-derived soils. Soil Sci. Soc. Am. J. 2018, 82, 66–75. [Google Scholar] [CrossRef]
- Cong, T.T. Study on supplying boron to coffee on basaltic soil in Central Highlands of Vietnam. J. Fertil. Pestic. 2017, 8, 000179. [Google Scholar] [CrossRef]
- Mühling, K.H.; Wimmer, M.; Goldbach, H.E. Apoplastic and membrane-associated Ca+2 in leaves and roots as affected by boron deficiency. Physiol. Plant. 1998, 102, 179–184. [Google Scholar] [CrossRef]
- DeMoranville, C.; Deubert, K. Effect of commercial calcium-boron and manganese-zinc formulations on fruit set of cranberries. J. Hortic. Sci. 1987, 62, 163–169. [Google Scholar] [CrossRef]
- Galeriani, T.M.; Nieves, G.O.; Santos Fereira, J.H.; Oliveira, R.N.; Calonego, J.C.; Crusciol, C.A.C. Calcium and boron fertilization improves soybean photosynthetic efficiency and grain yield. Plants 2022, 11, 2937. [Google Scholar] [CrossRef] [PubMed]
- Lambot, C.; Herrera, J.C.; Bertrand, B.; Sadeghian, S.; Benavides, P.; Gaitan, A. Cultivating coffee-quality—Terrior and Agro-Ecosystem. In The Craft and Science of Coffee; Folmer, B., Ed.; Academic Press/Elsevier: Amsterdam, The Netherlands, 2017; pp. 17–45. [Google Scholar]
- Buffo, A.R.; Cardelli, F.C. Coffee flavour: An overview. Flavour Fragr. J. 2004, 19, 99–104. [Google Scholar] [CrossRef]
- Campiño, L.I.; Villegas, H.A.M.; Arana, V.; Posada, H. Characterization of Chlorogenic acids (CGA) and nine isomers in a F2 population derived from Coffea arabica L. Agron. Colomb. 2020, 31, 19–28. [Google Scholar] [CrossRef]
- Dessalegn, Y.; Labuschagne, T.M.; Osthoff, G.; Herselman, L. Genetic diviersity and correlation of bean caffeine content with cup quality and green bean physical characteristics in coffee (Coffea arabica L.). J. Sci. Food Agric. 2008, 88, 1726–1730. [Google Scholar] [CrossRef]
- Chen, B.; Furtado, A.; Smyth, E.H.; Henry, J.R. Influence of genotype and environment on coffee quality. Trends Food Sci. Technol. 2016, 57, 20–30. [Google Scholar] [CrossRef]
- Figueiredo, P.L.; Borém, M.F.; Ribeiro, C.F.; Giomao, S.G.; Taveira, S.J.H.; Malta, R.M. Fatty acids profiles and parameters of quality of specialty coffees produced in different Brazilian regions. Afr. J. Agr. Res. 2015, 10, 3484–3493. [Google Scholar] [CrossRef]
- Toci, T.A.; Neto, J.M.F.; Torres, G.A.; Farah, A. Changes in triacylglycerols and free fatty acids composition during storage of roasted coffee. Food Sci. Technol. 2013, 50, 581–590. [Google Scholar] [CrossRef]
- Bellaloui, N. Effect of water stress and foliar boron application on seed protein, oil, fatty acids, and nitrogen metabolism in soybean. Am. J. Plant Sci. 2011, 2, 692–701. [Google Scholar] [CrossRef]
- He, M.; Ding, N.Z. Plant unsaturated fatty acids: Multiple roles in stress response. Front. Plant Sci. 2020, 11, 562785. [Google Scholar] [CrossRef] [PubMed]
- Upchurch, G.R. Fatty acids unsaturation, mobilization, and regulation in the response of coffee plants to stress. Biotechnol. Lett. 2008, 30, 967–977. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Muhammad, R.; Yan, L.; Zhang, Z.; Jian, C. How the cells were injured and the secondary metabolites in the shikimate pathway were changed by boron deficiency in trifoliate orange root. Plant Physiol. Biochem. 2020, 151, 630–639. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xia, F.; Wang, M.; Mao, P. Metabolomic analyses of alfalfa (Medicago sativa L. cv. Aohan) reproductive organs under boron deficit and surplus conditions. Ecotoxicol. Environ. Saf. 2020, 202, 111011. [Google Scholar] [CrossRef]
- Quiles, P.C.; Navarro, G.M.T.; Herrera, R.M.B.; Camacho, C.J.J.; Gonzales, F.A.; Rexach, J. Boron deficits increase cytosolic Ca+2 levels mainly via Ca+2 influx from the apoplast in Arabidopsis thaliana roots. Int. J. Mol. Sci. 2019, 20, 2297. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | T. min (°C) | T. max (°C) | T. med (°C) | R.H (%) | Rainfall (mm) | Sunshine (h) |
|---|---|---|---|---|---|---|
| 2014 | 15.7 | 23.6 | 19.1 | 74.6 | 1741.3 | 1233.1 |
| 2015 | 15.8 | 24.2 | 19.5 | 72.3 | 1319.6 | 1243.1 |
| 2016 | 16.1 | 24.1 | 19.6 | 73.4 | 1625.3 | 1241.4 |
| 2017 | 15.7 | 23.6 | 19.1 | 70.5 | 1976.3 | 1211.2 |
| 2018 | 15.7 | 23.5 | 19.0 | 75.3 | 1761.9 | |
| 2019 | 1482.1 | |||||
| Mean | 15.8 | 23.8 | 19.3 | 73.2 | 1651.1 | 1231.2 |
| Season | N | P2O5 | K2O | MgO | CaO | S |
|---|---|---|---|---|---|---|
| kg.ha−1 | ||||||
| 2014–2016 ¥ | 251 | 84 | 186 | 38 | 109 | 35 |
| 2016–2017 | 150 | 77 | 119 | 26 | 59 | 23 |
| 2017–2018 | 250 | 90 | 240 | 49 | 52 | 56 |
| 2018–2019 | 250 | 118 | 290 | 37 | 86 | 48 |
| Average | 225 | 92 | 209 | 38 | 77 | 41 |
| B Treatments/Season | 2014–2016 ¥ | 2016–2017 | 2017–2018 | 2018–2019 | Average |
|---|---|---|---|---|---|
| kg. B ha−1 | |||||
| High B rate | 0.8 | 1.1 | 1.05 | 1.39 | 1.1 |
| Reduced B rate (50% less) | 0.6 | 0.76 | 0.76 | 0.40 | 0.6 |
| Water Level (WL) | B Level (BL) | Nutrient Content | Nutrient Uptake by Plants | ||||
|---|---|---|---|---|---|---|---|
| Young Leaves | Mature Leaves | N | B | ||||
| N (%) | B (mg.kg−1) | N (%) | B (mg.kg−1) | mg. plant−1 | |||
| Without Water Stress | +B | 3.12 a | 63.72 b | 2.41 a | 265.07 c | 1873.4 b | 11.63 c |
| −B | 3.70 b | 12.73 a | 3.72 b | 9.56 a | 1553.6 a | 6.32 b | |
| With Water Stress | +B | 3.37 ab | 249.14 c | 3.86 b | 80.54 b | 1413.3 a | 0.38 a |
| −B | 4.69 c | 11.44 a | 4.27 c | 11.93 a | 1402.8 a | 0.37 a | |
| WL | ** | ** | ** | ** | ** | ** | |
| BL | ** | ** | ** | ** | ** | ** | |
| WL × BL | * | ** | ** | ** | * | ** | |
| Period | Harvest Year | Net Rainfall | Crop Evapotranspiration (ETc) |
|---|---|---|---|
| mm | |||
| 2015–2016 | 2016 | 529.0 | 565.0 |
| 2016–2017 | 2017 | 1096.0 | 980.0 |
| 2017–2018 | 2018 | 925.0 | 901.0 |
| 2018–2019 | 2019 | 837.0 | 833.0 |
| Treatment | Year | Caffeine | C14:0 Myristic | C16:0 Palmitic | C18:0 Stearic | C18:1 Oleate | C18:2 Linoleate | C18:3 Linolenate | C20:0 Arachidic | C22:0 Behenic |
|---|---|---|---|---|---|---|---|---|---|---|
| g.100g−1 | ||||||||||
| 77 kg CaO-1.1 kg B | 2016 | 1.35 c | 0.06 c | 19.44 | 3.86 | 4.94 c | 44.56 c | 2.84 c | 1.27 | 0.35 |
| 77 kg CaO-0.6 kg B | 1.36 c | 0.06 c | 20.47 | 3.98 | 5.09 c | 44.85 c | 2.88 c | 1.32 | 0.36 | |
| 0 kg CaO-0 kg B | 1.40 c | 0.05 c | 18.61 | 3.65 | 4.67 c | 41.32 bc | 2.66 c | 1.21 | 0.34 | |
| Average | 1.37 B | 0.06 B | 19.5 | 3.83 | 4.90 B | 43.58 B | 2.79 B | 1.27 | 0.35 | |
| 77 kg CaO-1.1 kg B | 2019 | 1.11 b | 0.01 a | 16.00 | 3.15 | 0.47 a | 27.11 a | 2.11 b | 1.24 | 0.32 |
| 77 kg CaO-0.6 kg B | 1.12 b | 0.03 b | 16.45 | 3.32 | 2.74 b | 29.12 a | 1.49 a | 1.29 | 0.34 | |
| 0 kg CaO-0 kg B | 1.02 a | 0.03 b | 19.78 | 3.89 | 2.19 b | 33.78 ab | 2.02 b | 1.53 | 0.41 | |
| Average | 1.09 A | 0.02 A | 17.4 | 3.45 | 1.80 A | 30.0A | 1.88 A | 1.35 | 0.36 | |
| Significance | ||||||||||
| Treatment | NS | NS | NS | NS | * | NS | NS | NS | NS | |
| Year | ** | ** | NS | NS | ** | ** | ** | NS | NS | |
| Treatment × Year | * | * | NS | NS | * | NS | * | NS | NS | |
| Pearson Correlation | Caffeine | C14:0 Myristic | C16:0 Palmitic | C18:0 Stearic | C18:1 Oleate | C18:2 Linoleate | C18:3 Linolenate | C20:0 Arachidic | C22:0 Behenic |
|---|---|---|---|---|---|---|---|---|---|
| g.100g−1 | |||||||||
| Correlation coefficient | −0.5684 | −0.6944 | −0.3401 | −0.3362 | −0.6298 | −0.6344 | −0.5923 | −0.0197 | −0.148 |
| p-value | 0.0047 ** | 0.0002 *** | 0.1123 ns | 0.1167 ns | 0.0013 ** | 0.0012 ** | 0.0029 ** | 0.9288 ns | 0.5004 ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramirez-Builes, V.H.; Küsters, J.; Thiele, E.; Leal-Varon, L.A. Boron Nutrition in Coffee Improves Drought Stress Resistance and, Together with Calcium, Improves Long-Term Productivity and Seed Composition. Agronomy 2024, 14, 474. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14030474
Ramirez-Builes VH, Küsters J, Thiele E, Leal-Varon LA. Boron Nutrition in Coffee Improves Drought Stress Resistance and, Together with Calcium, Improves Long-Term Productivity and Seed Composition. Agronomy. 2024; 14(3):474. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14030474
Chicago/Turabian StyleRamirez-Builes, Victor Hugo, Jürgen Küsters, Ellen Thiele, and Luis Alfredo Leal-Varon. 2024. "Boron Nutrition in Coffee Improves Drought Stress Resistance and, Together with Calcium, Improves Long-Term Productivity and Seed Composition" Agronomy 14, no. 3: 474. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14030474