Sensitivity Analysis of Alisma plantago-aquatica L., Cyperus difformis L. and Schoenoplectus mucronatus (L.) Palla to Penoxsulam



Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Collection
2.2. Plant Material and Growing Conditions
2.3. Preliminary Screenings
2.4. Dose-Response Experiments
3. Results
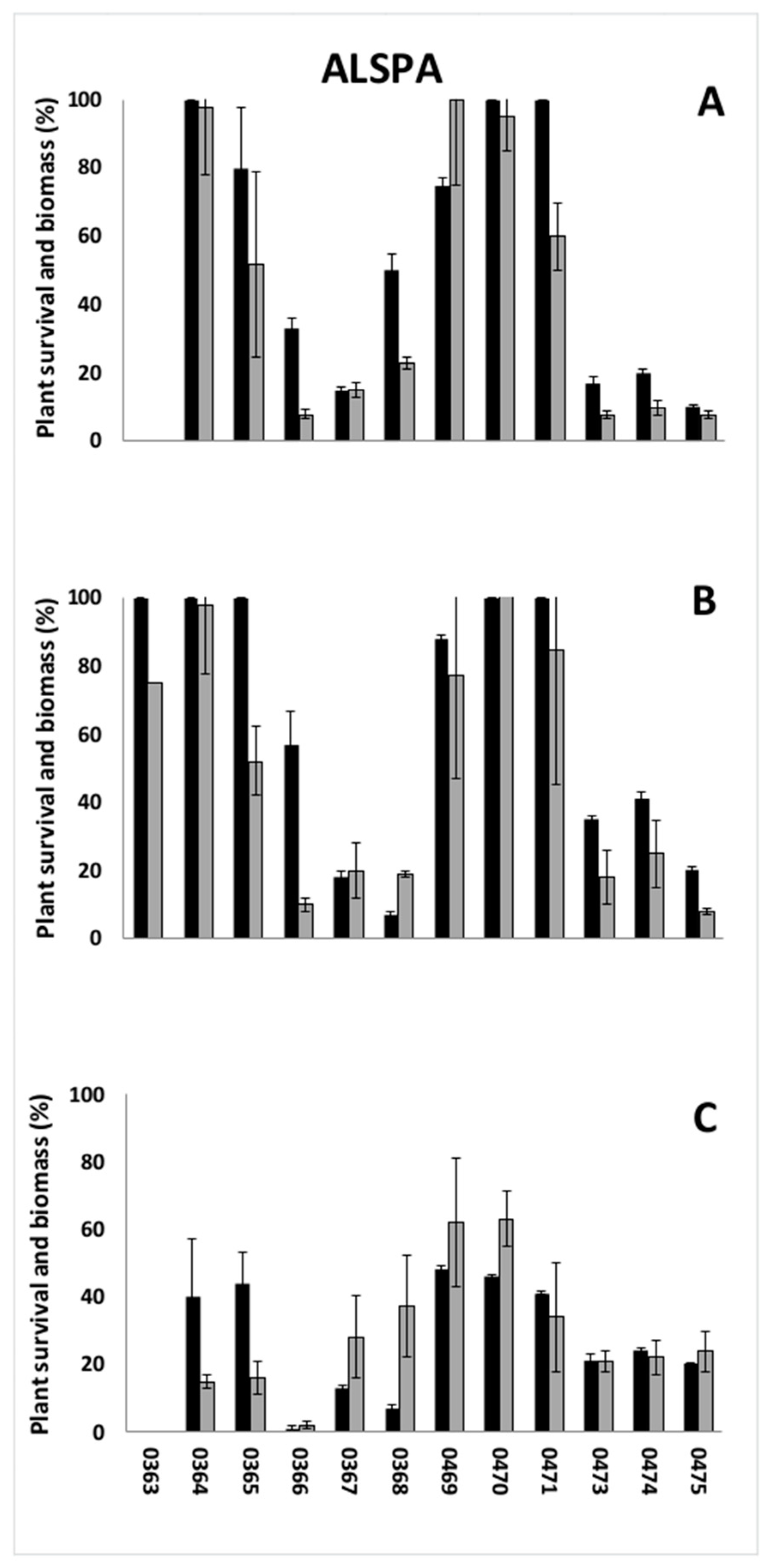
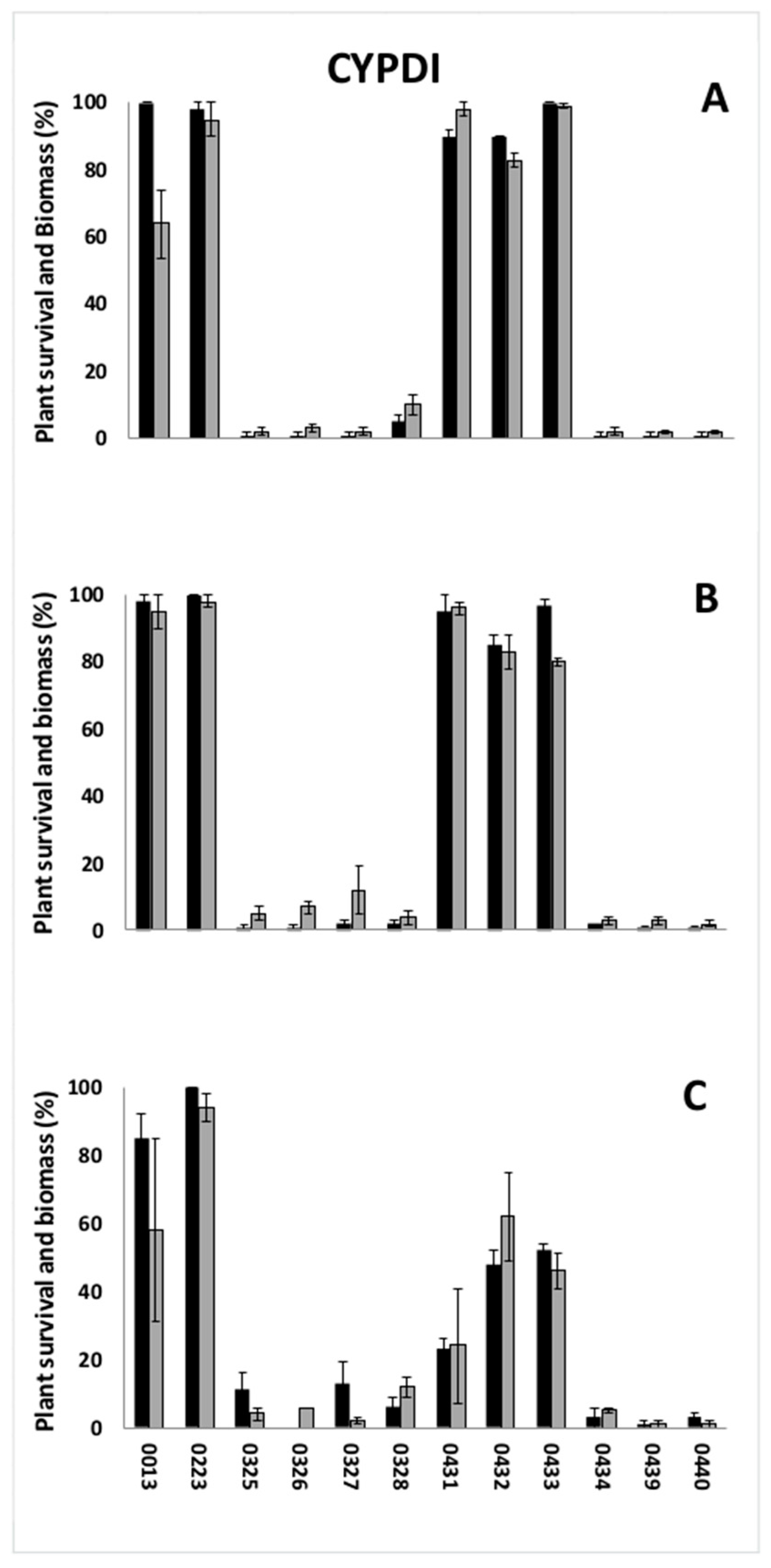
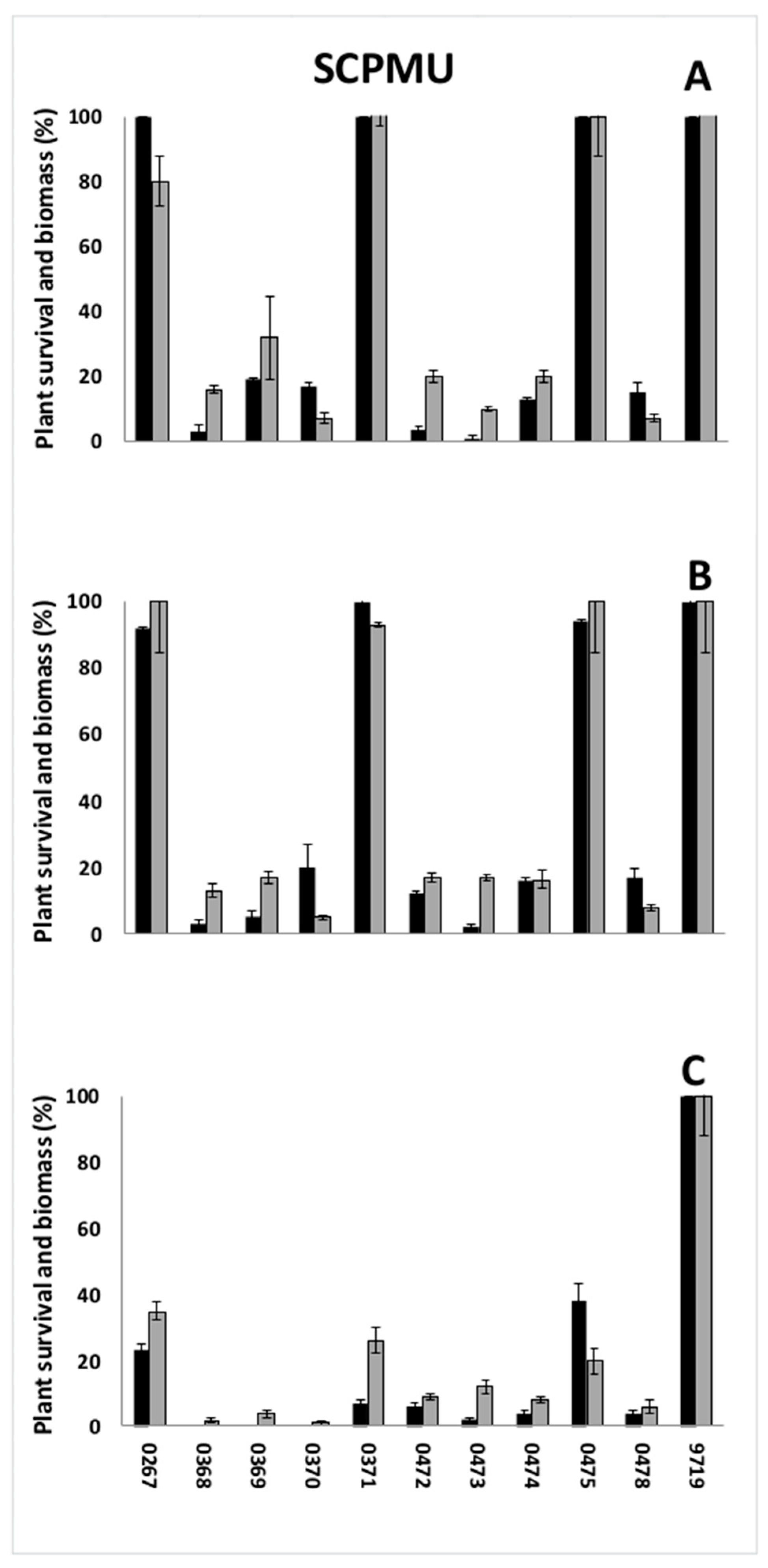
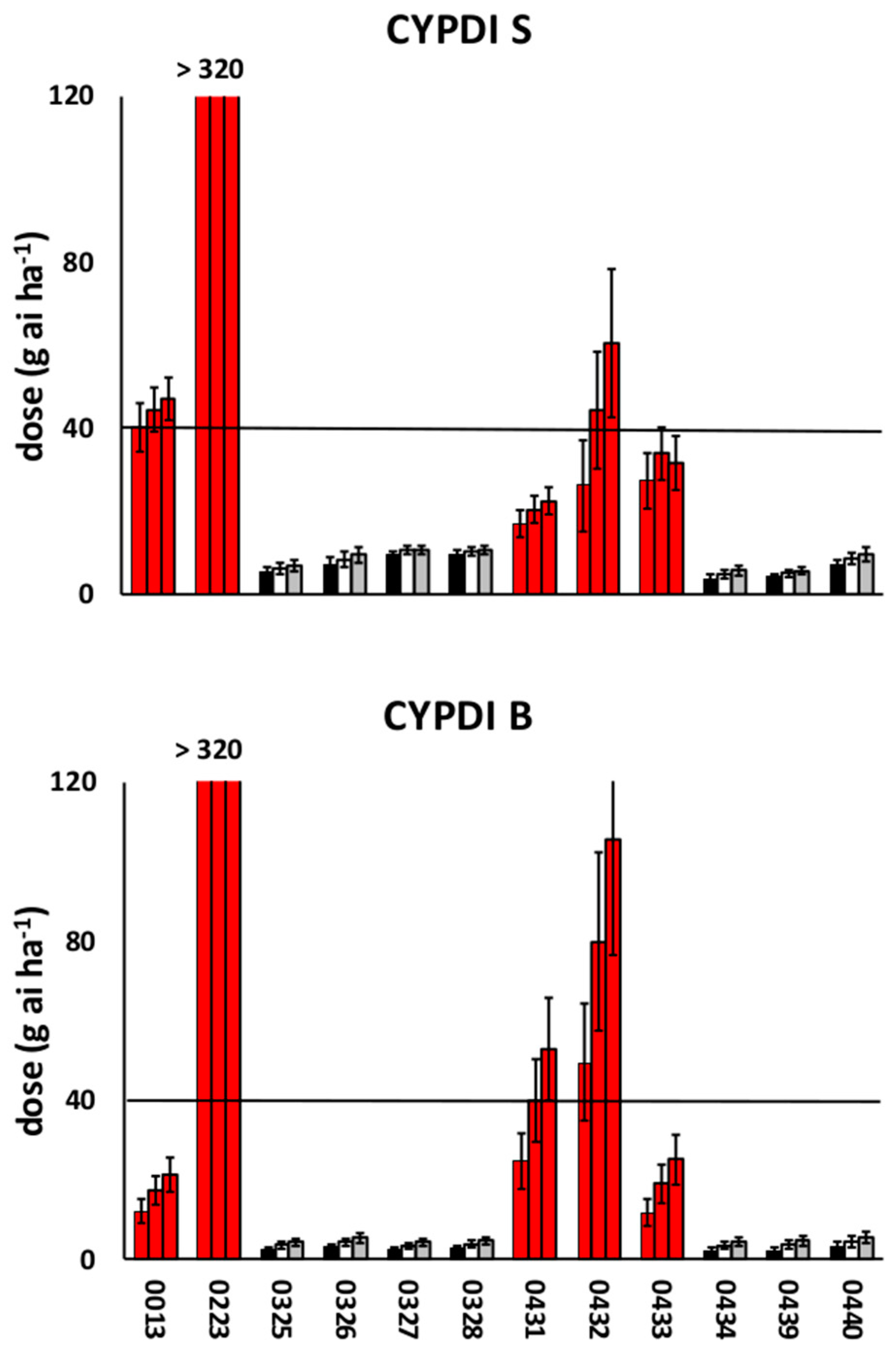
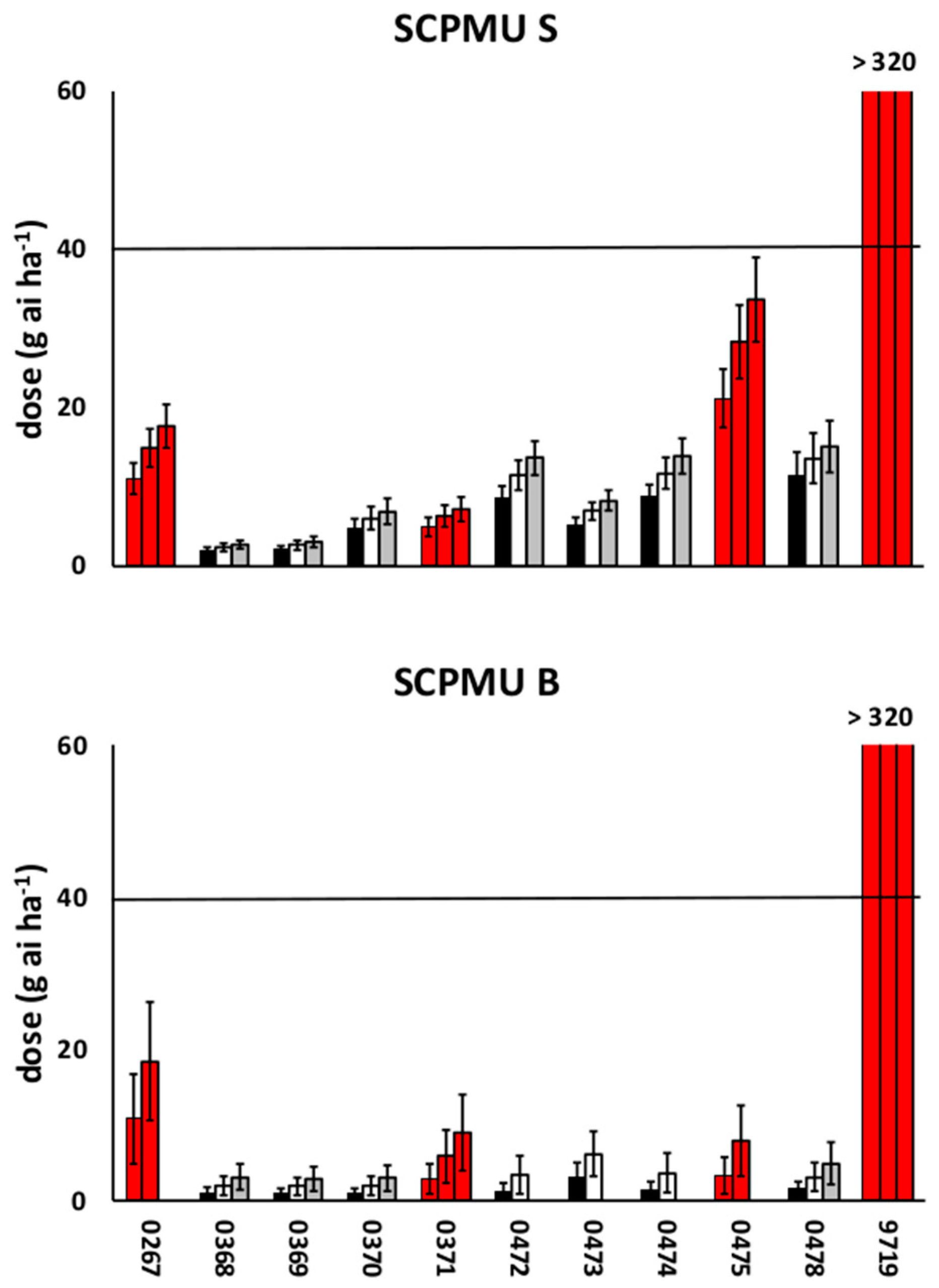
3.1. Preliminary Screenings
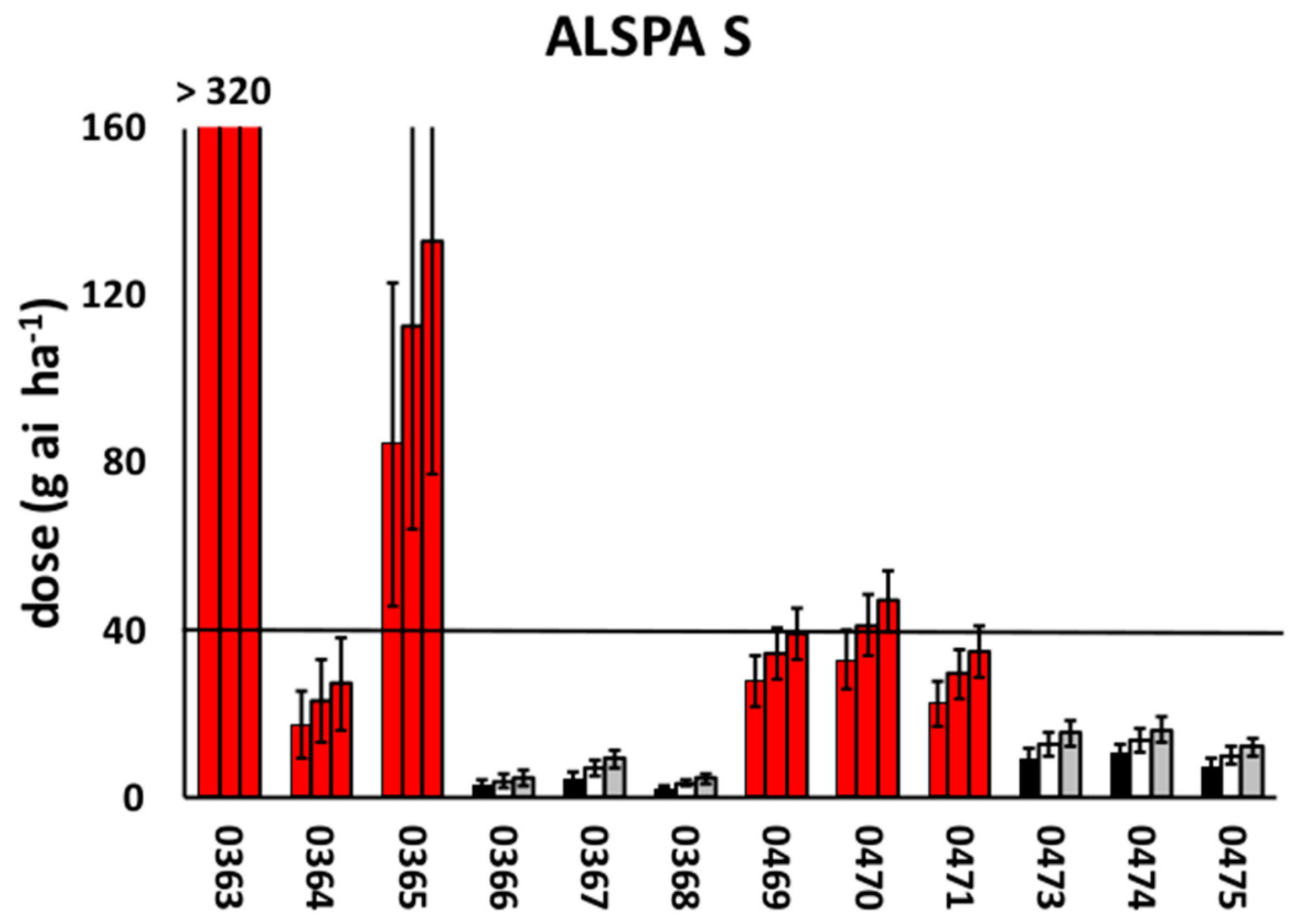
3.2. Dose-Response Experiments
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Heap, I. International Survey of Herbicide Resistant Weeds. Available online: www.weedscience.org (accessed on 30 July 2018).
- GIRE—Italian Herbicide Resistance Working Group—Database of Herbicide Resistance in Italy. Available online: www.resistenzaerbicidi.it (accessed on 30 July 2018).
- Sattin, M.; Berto, D.; Zanin, G.; Tabacchi, M. Resistance to ALS inhibitors in rice in north-western Italy. In Proceedings of the British Crop Protection Conference—Weeds, Brighton, UK, 15–18 November 1999; pp. 783–790. [Google Scholar]
- Sattin, M. Herbicide resistance in Europe: An overview. In Proceedings of the BCPC International Congress—Crop Science & Technology, Glasgow, UK, 31 October–2 November 2005; pp. 131–138. [Google Scholar]
- Osuna, M.D.; Vidotto, F.; Fischer, A.J.; Bayer, D.E.; de Prado, R.; Ferrero, A. Cross-resistance to bispyribac-sodium and bensulfuron-methyl in Echinochloa phyllopogon and Cyperus difformis. Pest. Biochem. Physiol. 2002, 73, 9–17. [Google Scholar] [CrossRef]
- Kuk, Y.I.; Kim, K.H.; Kwon, O.D.; Lee, D.J.; Burgos, N.R.; Jung, S.; Guh, J.O. Cross-resistance pattern and alternative herbicides for Cyperus difformis resistant to sulfonylurea herbicides in Korea. Pest. Manag. Sci. 2004, 60, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Merotto, A., Jr.; Jasieniuk, M.; Osuna, M.D.; Vidotto, F.; Ferrero, A.; Fischer, A.J. Cross-resistance to herbicides of five ALS-inhibiting groups and sequencing of the ALS gene in Cyperus difformis L. J. Agric. Food Chem. 2009, 57, 1389–1398. [Google Scholar] [CrossRef] [PubMed]
- Scarabel, L.; Locascio, A.; Furini, A.; Sattin, M.; Varotto, S. Characterisation of ALS genes in the polyploid species Schoenoplectus mucronatus and implications for resistance management. Pest. Manag. Sci. 2010, 6, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Panozzo, S.; Scarabel, L.; Tranel, P.J.; Sattin, M. Target-site resistance to ALS inhibitors in the polyploid species Echinochloa crus-galli. Pest. Biochem. Physiol. 2013, 105, 93–101. [Google Scholar] [CrossRef]
- Scarabel, L.; Cenghialta, C.; Manuello, D.; Sattin, M. Monitoring and management of imidazolinone-resistant red rice (Oryza sativa L., var. sylvatica) in Clearfield® Italian paddy rice. Agronomy 2012, 2, 371–383. [Google Scholar] [CrossRef]
- EPPO (European and Mediterranean Plant Protection Organization). Efficacy evaluation of plant protection products. Resistance risk analysis. PP1/213 (3). EPPO Bull. 2015, 45, 371–387. [Google Scholar] [CrossRef]
- Espeby, L.A.; Fogelfors, H.; Milberg, P. Susceptibility variation to new and established herbicides: Examples of inter-population sensitivity of grass weeds. Crop Prot 2011, 30, 429–435. [Google Scholar] [CrossRef] [Green Version]
- Beckie, H.J.; Tardif, F.J. Herbicide cross resistance in weeds. Crop Prot. 2012, 35, 15–28. [Google Scholar] [CrossRef]
- De Mol, F.; Gerowitt, B.; Kaczmarek, S.; Matysiak, K.; Sønderskov, M.; Mathiassen, S.K. Intraregional and inter-regional variability of herbicide sensitivity in common arable weed populations. Weed Res. 2015, 55, 370–379. [Google Scholar] [CrossRef]
- Duke, S.O. Why have no new herbicide modes of action appeared in recent years? Pest. Manag. Sci. 2011, 68, 505–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotteveel, T.; Jorgensen, L.N.; Heimbach, U. Resistance management in Europe: A preliminary proposal for the determination of a minimum number of active substances necessary to manage resistance. OEPP/EPPO Bull. 2011, 41, 432–438. [Google Scholar] [CrossRef]
- Kudsk, P. Optimising herbicide dose: A straightforward approach to reduce the risk of side effects of herbicides. Environmentalist 2008, 28, 49–55. [Google Scholar] [CrossRef]
- McDougall, P. The Global Agrochemical Market Trends by Crop. In Proceedings of the 11th China International Forum on Development of Pesticide Industry, Shanghai, China, 28 February 2017; Available online: http://www.cac-conference.com/Uploads/Editor/2017-03-07/58be2c387de29.pdf (accessed on 15 June 2018).
- Larelle, D.; Mann, R.K.; Cavanna, S.; Bernes, R.; Duriatti, A.; Mavrotas, C. Penoxsulam, a new broad spectrum rice herbicide for weed control in European Union paddies. In Proceedings of the BCPC International Congress—Crop Science & Technology, Glasgow, UK, 10–12 November 2003; Volume 1, pp. 75–80. [Google Scholar]
- Yasuor, H.; Osuna, M.D.; Ortiz, A.; Saldain, N.E.; Eckert, J.W.; Fischer, A.J. Mechanism of resistance to penoxsulam in late watergrass [Echinochloa phyllopogon (Stapf) Koss.]. J. Agric. Food Chem. 2009, 57, 3653–3660. [Google Scholar] [CrossRef] [PubMed]
- Scarabel, L.; Berto, D.; Sattin, M. Dormancy breaking and germination of Alisma plantago-aquatica and Scirpus mucronatus. Asp. Appl. Biol. 2003, 69, 285–292. [Google Scholar]
- Sattin, M.; Gasparetto, M.A.; Campagna, C. Situation and management of Avena sterilis ssp ludoviciana and Phalaris paradoxa resistant to ACCase inhibitors in Italy. In Proceedings of the BCPC Conference: Weeds, Brighton, UK, 13–15 November 2001; pp. 755–762. [Google Scholar]
- Hess, M.; Barralis, G.; Bleiholder, H.; Buhr, L.; Eggers, T.; Hack, H.; Stauss, R. Use of the extended BBCH scale—General for the descriptions of the growth stages of mono- and dicotyledonous weed species. Weed Res. 1997, 37, 433–441. [Google Scholar] [CrossRef]
- Panozzo, S.; Scarabel, L.; Collavo, A.; Sattin, M. Protocols for robust herbicide resistance testing in different weed species. J. Vis. Exp. 2015, 101, e52923. [Google Scholar] [CrossRef] [PubMed]
- Ritz, C. Toward a unified approach to dose-response modeling in ecotoxicology. Environ. Toxicol. Chem. 2010, 29, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Efron, B. The bootstrap and modern statistics. J. Am. Stat. Assoc. 2000, 95, 1293–1296. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, W.; Chen, X.; Liu, W.; Zhang, S.; Shu, Y. Studies on shapes and germination characters of Alisma plantago-aquatica seeds. Zhongguo Zhongyao Zazhi 2009, 34, 26–29. [Google Scholar] [PubMed]
- Tabacchi, M.; Scarabel, L.; Sattin, M. Herbicide resistance in Italian rice crops: A late-developing but a fast-evolving story. In Proceedings of the Conference Challenges and Opportunities for Sustainable Rice-Based Production Systems, Torino, Italy, 13–15 September 2004; pp. 227–238. [Google Scholar]
- Tranel, P.J.; Wright, T.R.; Heap, I.M. Mutations in Herbicide-Resistant Weeds to ALS Inhibitors. Available online: http://www.weedscience.com (accessed on 30 July 2018).
- Calha, I.M.; Osuna, M.D.; Serra, C.; Moreira, I.; De Prado, R.; Rocha, F. Mechanism of resistance to bensulfuron-methyl in Alisma plantago-aquatica biotypes from Portuguese rice paddy fields. Weed Res. 2007, 47, 231–240. [Google Scholar] [CrossRef]
- Busi, R.; Vidotto, F.; Fischer, A.J.; Osuna, M.D.; De Prado, R.; Ferrero, A. Patterns of resistance to ALS herbicides in smallflower umbrella sedge (Cyperus difformis) and ricefield bulrush (Schoenoplectus mucronatus). Weed Technol. 2006, 20, 1004–1014. [Google Scholar] [CrossRef]
- Tehranchian, P.; Riar, D.S.; Norsworthy, J.K.; Nandula, V.; McElroy, S.; Chen, S.; Scott, R.C. ALS-resistant smallflower umbrella sedge (Cyperus difformis) in Arkansas rice: Physiological and molecular basis of resistance. Weed Sci. 2015, 63, 561–568. [Google Scholar] [CrossRef]
- Cruz-Hipolito, H.; Osuna, M.D.; Vidal, R.A.; de Prado, R. Resistance mechanism to bensulfuron-methyl in biotypes of Scirpus mucronatus L. collected in Chilean rice fields. J. Agric. Food Chem. 2009, 57, 4273–4278. [Google Scholar] [CrossRef] [PubMed]
- Uchino, A.; Ogata, S.; Kohara, H.; Yoshida, S.; Yoshioka, T.; Watanabe, H. Molecular basis of diverse responses to acetolactate synthase-inhibiting herbicides in sulfonylurea-resistant biotypes of Schoenoplectus juncoides. Weed Biol. Manag. 2007, 7, 89–96. [Google Scholar] [CrossRef]
- Sada, Y.; Ikeda, H.; Kizawa, S. Resistance levels of sulfonylurea-resistant Schoenoplectus juncoides [Roxb.] Palla having various Pro197 mutations in acetolactate synthase to imazosulfuron, bensulfuron-methyl, metsulfuronmethyl and imazaquin-ammonium. Weed Biol. Manag. 2013, 13, 53–61. [Google Scholar] [CrossRef]
- Tehranchian, P.; Norsworthy, J.K.; Nandula, V.; McElroy, S.; Chenc, S.; Scott, R.C. First report of resistance to acetolactate-synthase-inhibiting herbicides in yellow nutsedge (Cyperus esculentus): Confirmation and characterization. Pest. Manag. Sci. 2015, 71, 1274–1280. [Google Scholar] [CrossRef] [PubMed]
- Tejavathi, D.H.; Nijalingappa, B.H.M. Cytological studies in some members of Cyperaceae. Cytologia 1990, 55, 363–372. [Google Scholar] [CrossRef]
- Cheema, P.; Bir, S.S. Cytomorphological studies in Cyperus Linn. V. Section Cyperus Linn. From Punjab, NW India; Indian Science Congress Association: Kolkata, India, 1995; Volume 82, pp. 82–83. [Google Scholar]
- Merotto, A., Jr.; Jasieniuk, M.; Fischer, A.J. Distribution and cross-resistance patterns of ALS-inhibiting herbicide resistance in smallflower umbrella sedge (Cyperus difformis). Weed Sci. 2010, 58, 22–29. [Google Scholar] [CrossRef]
- Sada, Y.; Kizawa, S.; Ikeda, H. Varied occurrence of diverse sulfonylurea-resistant biotypes of Schoenoplectus juncoides [Roxb.] Palla in Japan, as classified by an acetolactate synthase gene mutation. Weed Biol. Manag. 2012, 12, 168–176. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Population Code | Origin | Collection Site | Previous Direct Exposure to ALS Inhibitors |
|---|---|---|---|---|
| ALSPA | 0363 | Italy | Rice field | - |
| 0364 | Spain | Rice field | - | |
| 0365 | Italy | Rice field | - | |
| 0366 | Spain | Rice field | - | |
| 0367 | Italy | Ditch | No | |
| 0368 | Italy | Rice field | No or sporadic | |
| 0469 | Spain | Rice field | Yes | |
| 0470 | Italy | Rice field | - | |
| 0471 | Portugal | Rice field | - | |
| 0473 | Italy | Uncultivate area | No or sporadic | |
| 0474 | Italy | Ditch | No | |
| 0475 | Italy | Rice field | No or sporadic | |
| CYPDI | 0013 | Italy | Rice field | - |
| 0325 | Italy | Rice field | - | |
| 0326 | Spain | Rice field | Yes | |
| 0327 | Italy | Rice field | - | |
| 0328 | Spain | Rice field | Yes | |
| 0223 | Italy | Rice field | Yes | |
| 0431 | Spain | Rice field | Yes | |
| 0432 | Spain | Rice field | Yes | |
| 0433 | Italy | Rice field | Yes | |
| 0434 | Greece | Rice field | - | |
| 0439 | Italy | Rice field | No or sporadic | |
| 0440 | Italy | Rice field | No or sporadic | |
| SCPMU | 9719 | Italy | Rice field | Yes |
| 0368 | Italy | Rice field | - | |
| 0369 | Italy | Rice field | - | |
| 0370 | Italy | Rice field | - | |
| 0371 | Italy | Rice field | - | |
| 0267 | Italy | Rice field | Yes | |
| 0472 | Italy | Rice field | - | |
| 0473 | Italy | Rice field | - | |
| 0474 | Italy | Rice field | - | |
| 0475 | Italy | Rice field | - | |
| 0478 | Italy | Rice field | No or sporadic |
| ED50 or GR50 g ai ha−1 | ED80 or GR80 g ai ha−1 | |||||
|---|---|---|---|---|---|---|
| Average | Confidence Interval | Average | Confidence Interval | |||
| ALSPA | ||||||
| Plant survival | 6.0 | 3.6 | 8.4 | 8.4 | 5.2 | 11.6 |
| CYPDI | ||||||
| Plant survival | 6.5 | 4.9 | 8.1 | 7.6 | 6.0 | 9.2 |
| Fresh biomass | 2.6 | 2.4 | 2.8 | 3.8 | 3.5 | 4.1 |
| SCPMU | ||||||
| Plant survival | 6.1 | 3.7 | 8.5 | 7.8 | 4.8 | 10.8 |
| Fresh biomass | 1.4 | 0.9 | 1.9 | 3.1 | 2.1 | 4.1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loddo, D.; Kudsk, P.; Costa, B.; Dalla Valle, N.; Sattin, M. Sensitivity Analysis of Alisma plantago-aquatica L., Cyperus difformis L. and Schoenoplectus mucronatus (L.) Palla to Penoxsulam. Agronomy 2018, 8, 220. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy8100220
Loddo D, Kudsk P, Costa B, Dalla Valle N, Sattin M. Sensitivity Analysis of Alisma plantago-aquatica L., Cyperus difformis L. and Schoenoplectus mucronatus (L.) Palla to Penoxsulam. Agronomy. 2018; 8(10):220. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy8100220
Chicago/Turabian StyleLoddo, Donato, Per Kudsk, Bruno Costa, Natalino Dalla Valle, and Maurizio Sattin. 2018. "Sensitivity Analysis of Alisma plantago-aquatica L., Cyperus difformis L. and Schoenoplectus mucronatus (L.) Palla to Penoxsulam" Agronomy 8, no. 10: 220. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy8100220