1. Introduction
Due to the cultivation of high-yield varieties and the development of high-yield cultivation techniques, rice yields in China have increased. In 2017, rice production in China reached 221.76 million tons, accounting for 32.1% of total grain output. N is essential for rice production and has an important effect on rice growth and yield formation [
1]. However, excessive N fertilizer application and improper fertilization methods increase production costs, which not only restrict the production potential of rice but also negatively affect the environment by leaching, runoff, and volatilization [
2,
3]. Optimizing the application of N fertilizer to synchronize the supply of N with crop demand is of great significance for high-yield and high-efficiency rice production.
In rice production in China, N fertilizer is usually supplied via multiple applications as basal, tiller, and panicle fertilizer. Basal N is applied and mixed with soil before transplanting and tiller N is topdressed at 7–14 days after transplanting (DAT). Both basal and tiller N are used mainly to promote tillering during the early growth stage. Panicle N is topdressed at the panicle initiation (PI) stage and/or the initial spikelet differentiation (SD) stage. Panicle N is usually used to increase the number of differentiated spikelets, prevent differentiated spikelets from degeneration, increase dry matter accumulation during the middle and late growth stages, and increase the percentage of filled grains [
4,
5]. Generally, the basal:tiller:panicle fertilizer-N ratio is 40%–60%:20%–30%:20%–30%. The N-recovery efficiency (NRE) of panicle fertilizer is much greater than that of basal and tiller fertilizer. The NRE of basal and tiller fertilizer is approximately 10%–30%, while that of panicle fertilizer is 30%–80% [
6,
7]. In total, 35%–40% of applied fertilizer-N is absorbed by plants, approximately 20% to 26% is derived from panicle fertilizer, and basal and tiller fertilizers account for only approximately 7% to 10% [
8,
9,
10]. Given that the NRE of panicle N is significantly greater than that of basal and tiller N, to improve the N-use efficiency (NUE) and to reduce the loss of N, postponing N applications and heavy fertilization at the reproductive stage (i.e., reducing the amounts of basal and tillering N and increasing the amount of panicle N from 20%–30% to 40%–50%) is recommended as an effective measure to increase yields and has been adopted by farmers in parts of the Yangtze River Basin and the southern rice production area in China (e.g., Lin et al. [
11], Jiang et al. [
12], and Zhang et al. [
13]). Huang [
14] suggested the ratio of panicle N should be increased from 30% to 50% to control the development of ineffective tiller and increase the productive tiller percentage. Moreover, Sun et al. [
15] believed that increasing the ratio of panicle N to 40% was beneficial regarding the transport of nutrients from vegetative organs to grains during the grain-filling period.
However, some researchers have different opinions. Wu et al. [
16] reported that once the percentage of panicle N exceeded 35% in double-season early rice, the changing of rice leaf color from green to yellow during ripening was delayed, which was unfavorable to grain-filling and yield increase. Lin et al. [
8] reported that although increasing the ratio of panicle N from 30% to 50% increased the number of spikelets per panicle and the 1000-grain weight at a rate of 150 kgNha
−1, the yield at a rate of 300 kgNha
−1 decreased due to reductions in the seed-setting rate and the 1000-grain weight. Zhang et al. [
17] reported that the effectiveness of N applied at the PI and spikelet differentiation (SD) stages on grain yield and N-use efficiency (NUE) varied substantially with varieties with different panicle sizes and suggested the application of N at the PI stage to increase the sink capacity for varieties with a small panicle size, at the SD stage to increase filling efficiency for varieties with a large panicle size, and at either at the PI stage or the SD stage for varieties with a medium panicle size. Xu et al. [
18] reported that when the paddy field yield in control plots (no N fertilizer) was greater than 7000 kgha
−1, increasing the ratio of panicle N had no effect on rice grain yields. These results implied that the effects of increasing the ratio of panicle N on rice grain yield might be related to N rate, variety type, and soil fertility, but the relationship between these variables has been unclear until now.
We previously found that the NRE of N applied both before transplanting and at tillering was significantly lower than the NRE applied at PI when fertilizer-N uptake only in plants was considered; however, when the distribution of fertilizer-N in both the plants and soil was compared, 70% of the basal-
15N, 29% of the tillering-
15N, and 46% of the panicle-
15N was recovered in the plants and soil [
7,
19]. Basal fertilizer is an effective long-term fertilizer; all of the fertilizer-
15N taken up by plants after heading was derived from basal N, and the residual basal fertilizer-N accounted for more than 90% of the total fertilizer-N that remained in paddy soil, which was available to subsequent crops via remineralization [
7]. Therefore, the low recovery efficiency of basal fertilizer-N in plants did not indicate high N loss, indicating that it is more reasonable to increase the ratio of N applied at PI by reducing the percentage of N applied at tillering while maintaining the ratio of basal N, that is, by postponing topdressing-N to increase the ratio of N applied at PI.
To determine the effect of postponing topdressing-N on the yield of different types of japonica rice and its relationship with soil fertility, we carried out field experiments in 2016 and 2017. Inbred japonica rice and hybrid japonica rice, which are mainly grown in the middle and lower reaches of the Yangtze River, were used as materials, and the effects of different ratios of panicle N on rice grain yield and dry matter accumulation in high- and low-fertility blue clayey paddy soils were analyzed.
4. Discussion
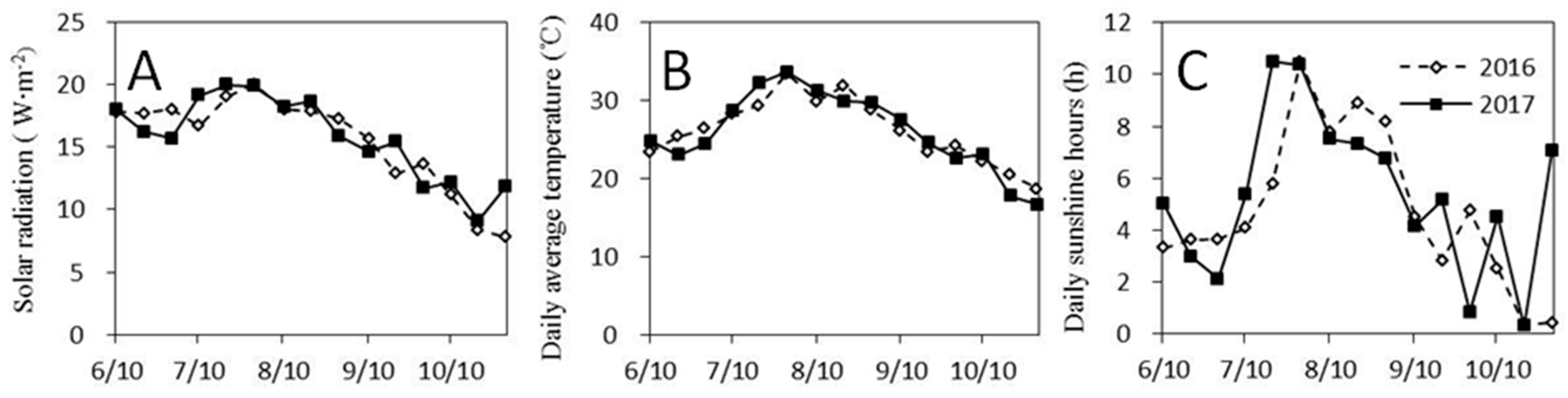
In this study, there was a significant difference in grain yield and dry matter accumulation between the years in both the high-fertility and low- fertility soils, with significantly lower values being detected in 2017 compared with 2016. This difference was likely due to the worse light and temperature conditions during the critical rice growth period in 2017. For example, the daily average temperature, solar radiation, and sunshine hours for two weeks after transplanting in 2016 were 26.1 °C, 18.2 Wm
−2, and 3.8 h, respectively, and 23.9 °C, 16.5 Wm
−2, and 2.8 h in 2017, respectively. Previous research showed that sunshine, temperature, water, and N affect tiller development [
21,
22]. Low temperature, low light intensity, and short sunshine duration after transplanting in 2017 delayed tiller development, and the number of effective panicles of the inbred and hybrid
japonica rice decreased from 2016 to 2017 by 18.6% and 17.5%, respectively. In addition, at the beginning of the panicle differentiation stage (2–8 August), the daily average temperature and the maximum and minimum daily temperatures in 2017 were 31.8 °C, 37.9 °C, and 28.4 °C, respectively, which were obviously greater than the 29.8 °C, 35.1 °C, and 26.2 °C values in 2016. It is well-known that the number of grains per panicle is determined at the PI stage, which is one of the most sensitive stages to high temperature and heat [
23]. Rice spikelets are prone to dysplasia when the average daily temperature is greater than 30 °C and when the daily maximum temperature reaches 35 °C [
24]. The high temperature at PI in 2017 in our experiment had a negative impact on the development of spikelets and resulted in an 8.2% and 5.6% decrease in the average grains per panicle of the inbred and hybrid
japonica varieties, respectively. Furthermore, the temperature decreased abruptly during the late grain-filling period in 2017 (12–22 October), and the daily average temperature and the daily minimum temperature in 2017 were 17.5 °C and 15.6 °C, respectively, which were significantly lower than the corresponding values (20.7 °C and 19.3 °C, respectively) in 2016. As the numbers of effective panicles and grains per panicle in 2017 were lower than those in 2016, the yield in 2017 was lower than that in 2016.
Berge et al. [
25] recommended that frequent, small doses of fertilizer be applied to synchronize with crop demand. In transplanted rice production systems, N fertilizer is applied several times, for example, as a basal fertilizer (applied before transplanting), revival (turning green) fertilizer (applied within one week after transplanting), tillering fertilizer, and panicle fertilizer. Tillering and panicle fertilizer can be topdressed more than once, i.e., at the initial and middle tillering/booting stages or even at or after heading [
26]. However, given increased labor costs and findings describing that more than three divided fertilizer-N applications had only small effects on grain yield, N fertilizer is usually applied to paddy fields three times, namely, before transplanting, at initial tillering, and at PI. Previous studies have clarified the function of N applied during different growth periods [
27,
28]. The major roles of basal and tillering fertilizer-N are to stimulate the production of tillers and to promote early growth [
27,
28]. It is believed that the leaves on the main culm emerge successively at definite intervals and that the individual leaves on the main culm develop simultaneously with certain tillers [
29]. A suitable N level in the plant shortens the leafing interval of the main culm and causes secondary, tertiary, and quaternary tillers to appear ahead of the synchronous pattern; however, when the N level decreases, the leafing interval becomes longer, and a delay in tiller development results [
30]. Therefore, the premise of “postponing N application and heavy fertilization at the booting stage” is that N supply is sufficient during the early growth stage; otherwise, it is easy to obtain insufficient numbers of effective panicles and reduced yields. The view of “postponing N application” management is agreed upon and supported by many researchers in China. Zhang et al. [
31] reported that postponing the application of N increased the panicle N, consequently constituting 50% of the total N to prevent excessive growth of the tillers and leaves at the vegetative stage, while both the dry matter accumulation during the grain-filling period and grain yield increased. Similar results were also reported by Lin et al. (2005), Li et al. (2010), and Sun et al. (2017). However, these studies only used limited varieties as materials and ignored the effects of soil N supply capacity on plant N demand.
It is well-known that a significant portion of N uptake by rice plants originates from soil N, and that the yield of rice growing in high-fertility soil is usually greater than that growing in low-fertility soil [
13,
32]. For example, Zhang et al. [
13] reported that more than 60% of N in plant tissue was derived from soil N supplied by N which was applied at rates ranging from 240–300 kgha
−1; [
33] reported that 80% of the N within a crop was derived from soil N supplied by N which was applied at a rate of 140 kgha
−1. Using the mid-season
indica hybrid rice Chuanxiang-you 9839 as a material, Xu et al. [
18] reported that the effect of an increased ratio of panicle N on rice grain yield was related to the basic fertility of the soil. If the yield of the soil not fertilized with N was more than 7000 kgha
−1, postponing the N application did not increase the yield, and even reduced the yield when 40% of the total N was applied at PI.
In this study, six inbred and four hybrid japonica rice varieties were used as materials, and the effects of postponing topdressing-N in both high-fertility and low-fertility blue clayey paddy soils were analyzed. In contrast to previous studies, our experiment showed that the effects of postponing topdressing-N on rice yield were related to variety, meteorological conditions, and soil fertility. Three general conclusions are outlined below.
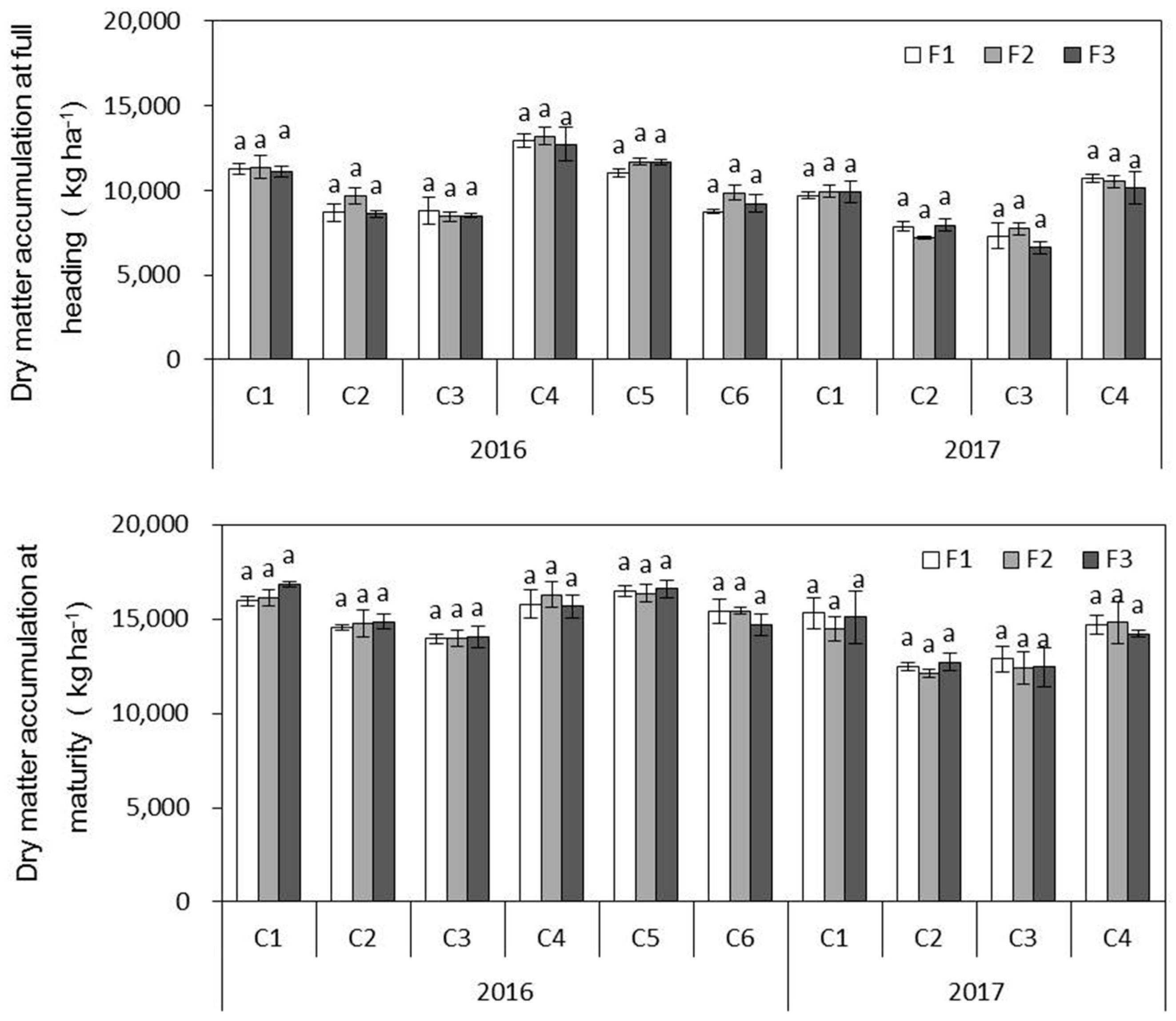
(1) In inbred japonica rice, increasing the ratio of panicle N had no effect on grain yield or dry matter accumulation in both high-fertility and low-fertility soils, although there were differences between years. The yield components in the different treatments in the same year were similar, as was the dry matter accumulation.
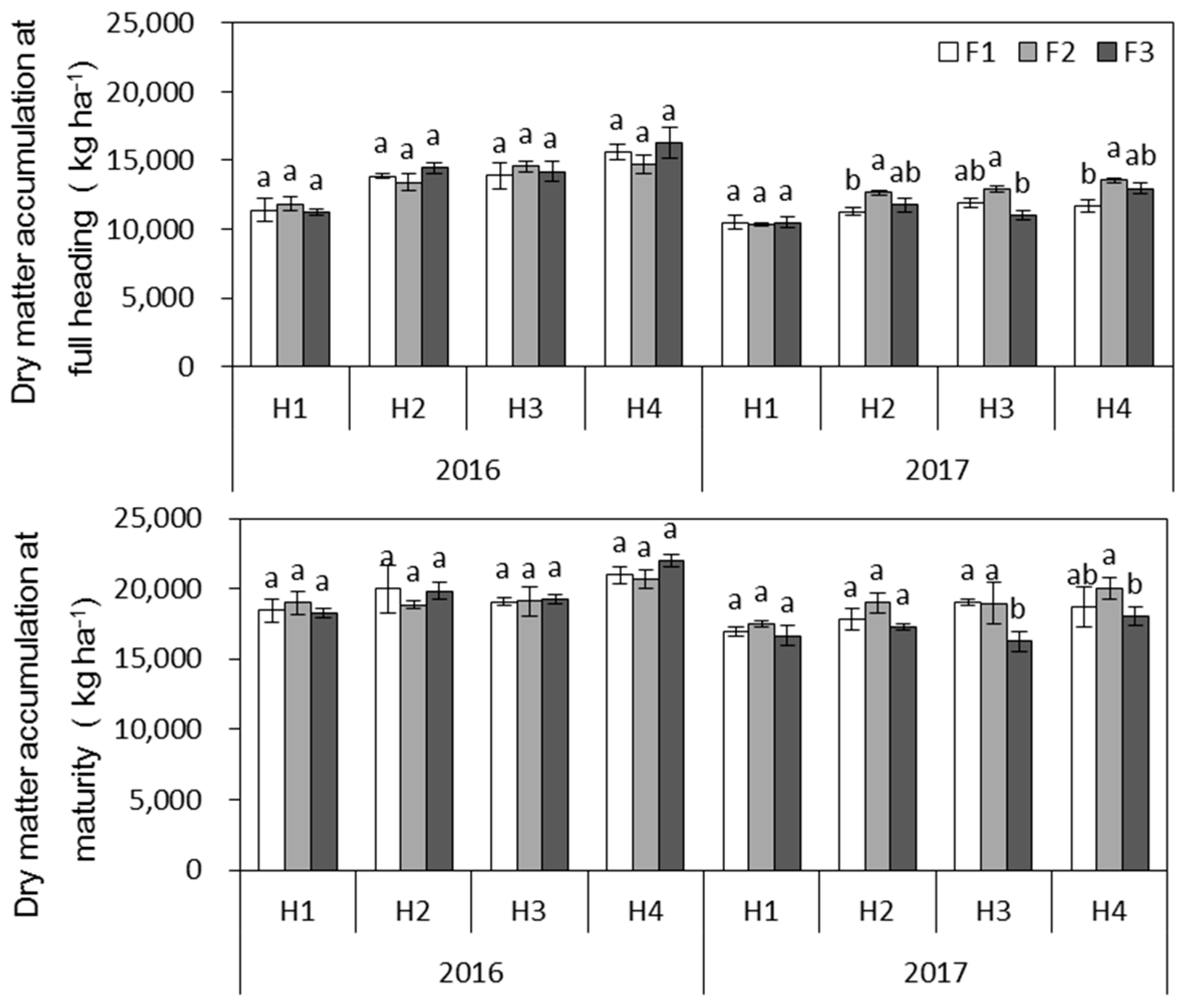
(2) With respect to hybrid
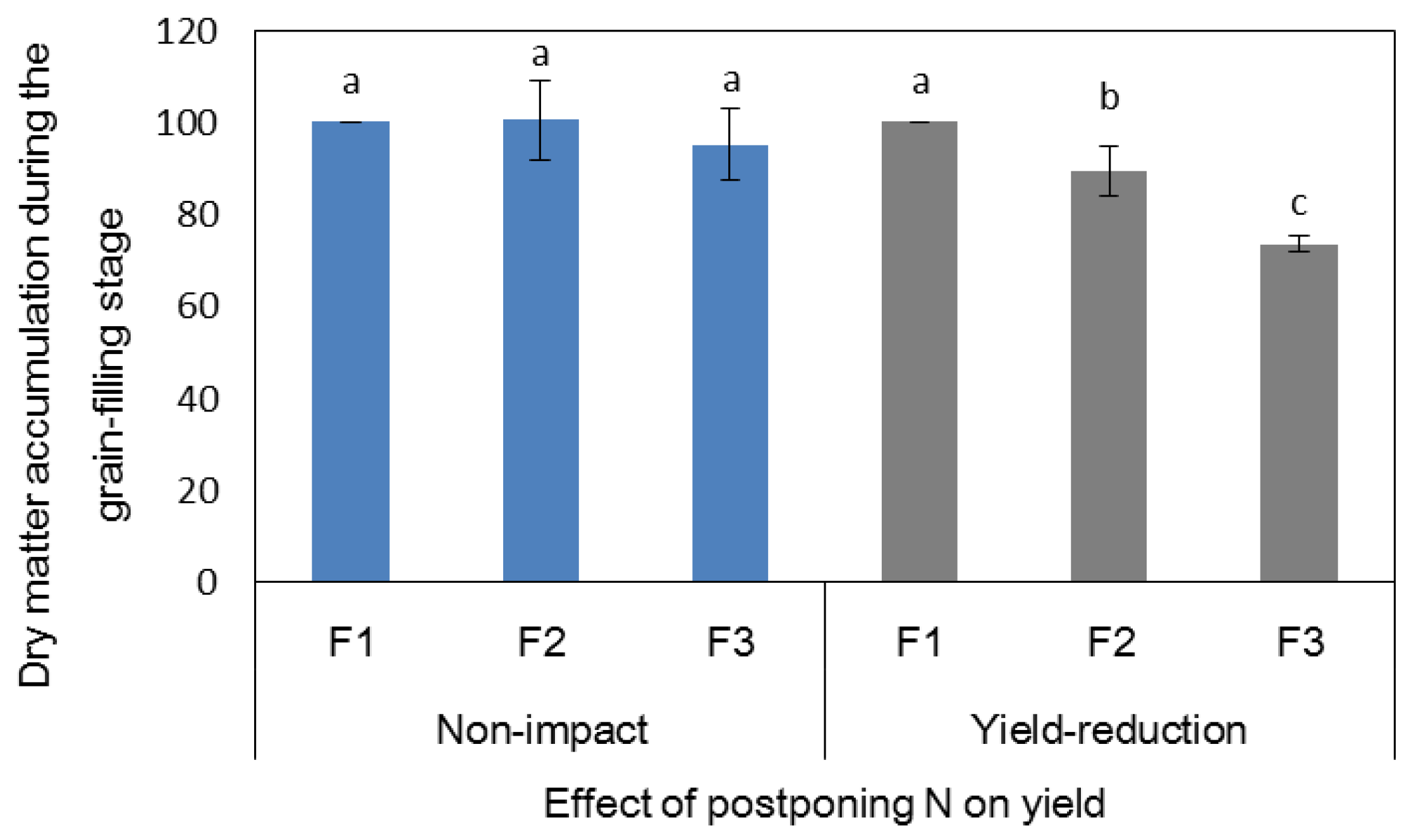
japonica rice grown in high-fertility soil, increasing the ratio of panicle N had no effect on the number of effective panicles, the number of spikelets per panicle, or the 1000-grain weight in either year, while the effect on the seed-setting rate differed between 2016 and 2017. In 2016, which presented good temperature conditions during the grain-filling period, there was no significant difference in seed-setting rate between the treatments for any variety; however, in 2017, during which the temperature sharply declined in mid-October, the seed-setting rate of only H1 and H2 remained unchanged, while that of H3 and H4 significantly decreased when 40% of the total N was applied as a single application at PI. Grain-filling is a process involving starch accumulation. The sucrose used for starch synthesis in the grain originates from the degradation of nonstructural carbohydrates (NSCs) assimilated in the leaf sheaths and stems before heading and from the translocation of carbon assimilates from the leaves after heading [
34]. Zhang et al. [
5] reported that NSCs translocated more slowly in response to high levels of panicle N compared with moderate levels. Only 37.9% of
13C translocated after 20 days after heading under high-panicle N treatment in IIyou 107, while 63.8% translocated under moderate levels. Weather conditions also affect NSC translocation; the grain weight and seed-setting rate of
japonica rice are maximal when the average daily temperature during grain-filling is 23.5–24 °C [
35]. However, these parameters are severely affected when the daily average temperature is lower than 20 °C or when the lowest temperature is lower than 15 °C [
36]. When the growth periods of the four hybrid rice varieties were compared, H1 and H2 were heading at the end of August and were mature on 9 and 16 October in 2017, respectively, while H3 and H4 were heading during 7–11 September and were mature at the end of October. Moreover, the low temperature during mid-October in 2017 had a short-term effect on H1 and H2 but had a long-term and severe effect on the grain-filling of H3 and H4. Therefore, in high-fertility soil, the effect of postponing topdressing-N on the yield of hybrid rice was related to the growth periods of the varieties and the weather conditions. Postponing topdressing-N application slowed the translocation of NSCs, which increased the probability of low-temperature stress on varieties that had longer growth periods.
(3) With respect to the hybrid japonica rice grown in low-fertility soil, a significant decrease in the number of effective panicles was observed when the ratio of tillering N decreased from 30% to 20%, indicating that postponing topdressing-N resulted in a N shortage during the early growth stage.
All of the above results showed that postponing topdressing-N had no obvious effect on increasing the grain yield of
japonica rice in either high- or low-fertility soils. The results of these experiments therefore challenge the findings of Xu, who reported that increasing the ratio of panicle N had no effect on the yield of hybrid rice grown in high-fertility soil but could increase the grain yield of hybrid rice grown in low-fertility soil [
18]. This discrepancy might be due to the different N application methods and soil type/texture in the two experiments. In contrast to other methods, increasing the ratio of panicle N by reducing both basal and tiller N decreased the proportion of only tiller N, while the ratio of basal N remained unchanged. Our previous research indicated that basal fertilizer was an effective long-term fertilizer and that the proportion of total N at the full heading stage was much greater from the basal N than from the tiller N and panicle N [
7], especially in clayey paddy soils. Organic matter can act as a reservoir of plant nutrients [
37] and supplies nutrients via mineralization, desorption, and binding to reduce nutrient leaching. Soil organic matter (SOM) decomposition and accumulation are influenced by soil texture [
38], clay mineralogy [
39], temperature, water content, and microbial activity [
40,
41]. Clay minerals can bind organic matter [
42] and protect SOM from decomposition via physical protection by the formation of macroaggregates [
43] and organo-mineral complexes [
44]. Compared with sandy soils, the blue clayey soil in this experiment presented slower SOM decomposition, especially when the soil temperature was low in the early stage of growth; postponing topdressing-N by reducing the ratio of tillering N therefore adversely affected tiller development and decreased the number of effective panicles in the low-fertility soil. However, clayey soils have a strong fertility conservation capability, and our previous research showed that basal fertilizer-N, tillering fertilizer-N, and panicle fertilizer-N comprised 1.3%, 0.6%, and 0.4% of the soil N at the heading stage when 210 kgNha
−1 was applied at a ratio of 40%:30%:30% [
7]. With an increase in soil temperature at the reproduction stage, the release of soil nutrients accelerated. Compared with the hybrid rice grown in the high-fertility soil, the rice grown in the low-fertility soil had 17.4% fewer effective panicles but 20.4% more grains per panicle, and no significant difference in grain yield was detected between the high- and low-fertility soils in 2016. Thus, the experiments carried out in the blue clayey paddy soil revealed that soil fertility affected mainly the occurrence of tillers at the early growth stage. Moreover, the adverse effects on grain yield were reduced by increasing the number of spikelets per panicle when there was no inclement weather during grain-filling, indicating that the N supply at the booting stage met the needs of rice growth and was not the yield-limiting factor in this study, even in the low-fertility soil. Therefore, in this blue clayey soil, the proportion of panicle N applied to
japonica rice should not exceed 30%; otherwise, excessive fertilizer at PI will aggravate the instability of rice yields. However, due to the lack of relevant soil information support, the relationships between soil type, texture, and the effect of postponing N applications remain unexplored.






{kind=link}
{kind=link}
{kind=link}
{kind=link}