Microbial Consortia versus Single-Strain Inoculants: An Advantage in PGPM-Assisted Tomato Production?

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Case Study 1: Large-Scale Greenhouse Tomato Production in Timisoara, Romania 2016/2017
2.1.1. Tomato Cultivation and Fertilization
2.1.2. Microbial Inoculation
2.1.3. Plant Protection
2.1.4. Experimental Design and Statistical Evaluations
2.1.5. Pre- and Postharvest Analyses
2.2. Case study 2: Drip-Irrigated Field Production of Tomato (ARO Research Center), 2017
2.2.1. Tomato Cultivation and Fertilization
2.2.2. Microbial Inoculation
2.2.3. Plant Protection
2.2.4. Experimental Design and Statistical Evaluation
2.2.5. Pre- and Postharvest Analysis
2.2.6. Soil Microbiome Amplicon Sequencing
3. Results
3.1. Case Study 1: Greenhouse Tomato Production in Timisoara, Romania 2016 and 2017
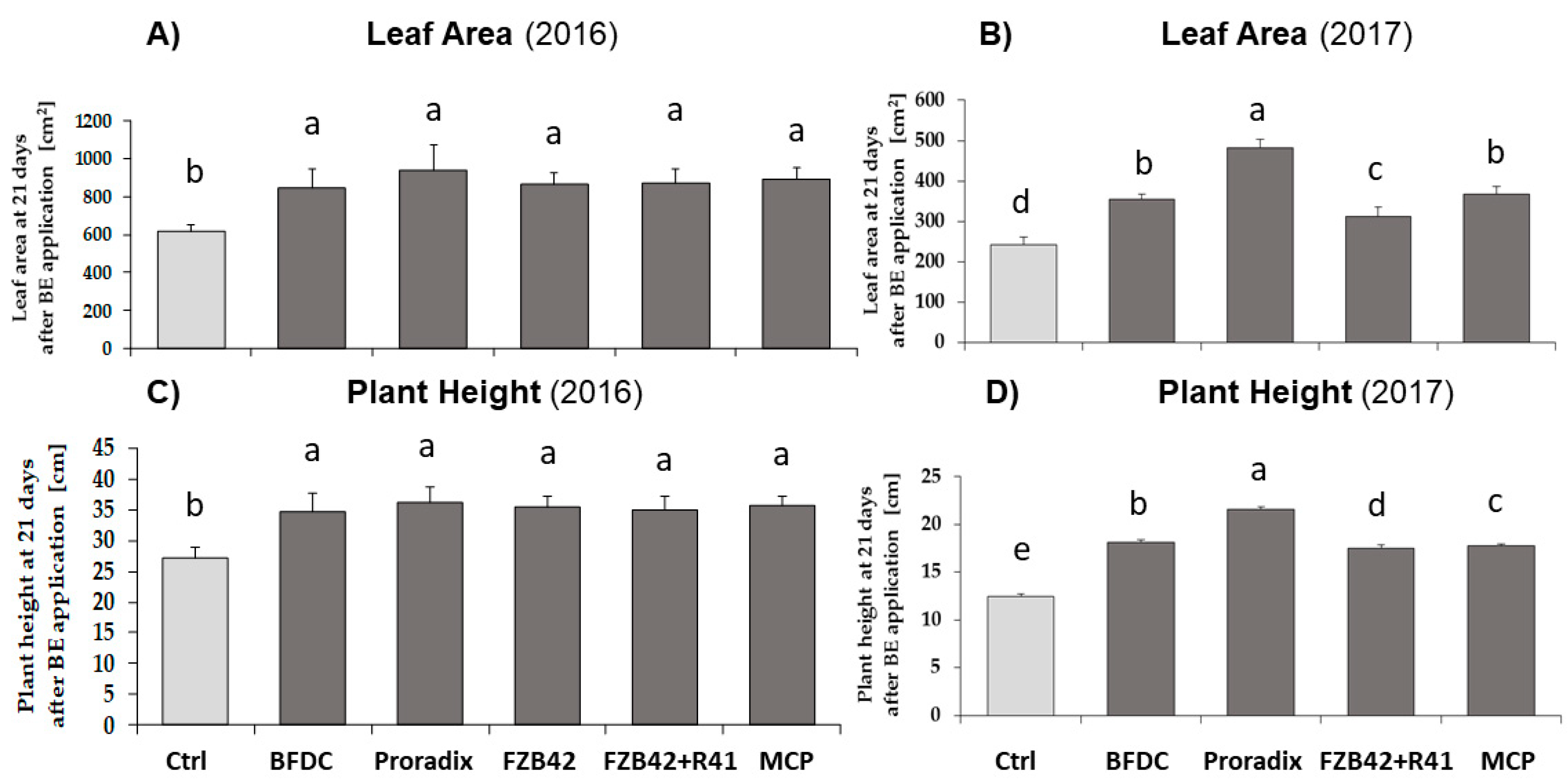
3.1.1. Growth of Nursery Plants
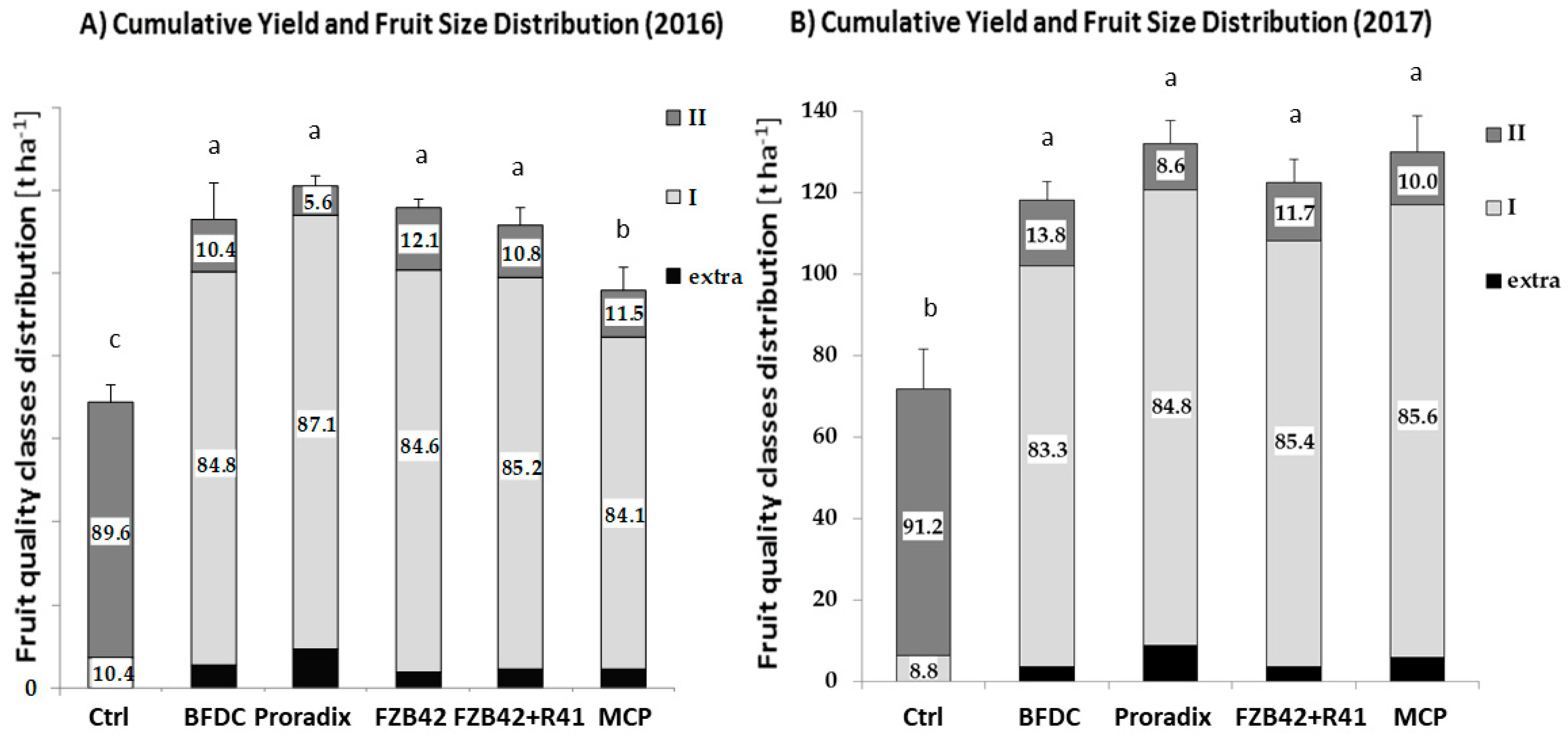
3.1.2. Cumulative Fruit Yield
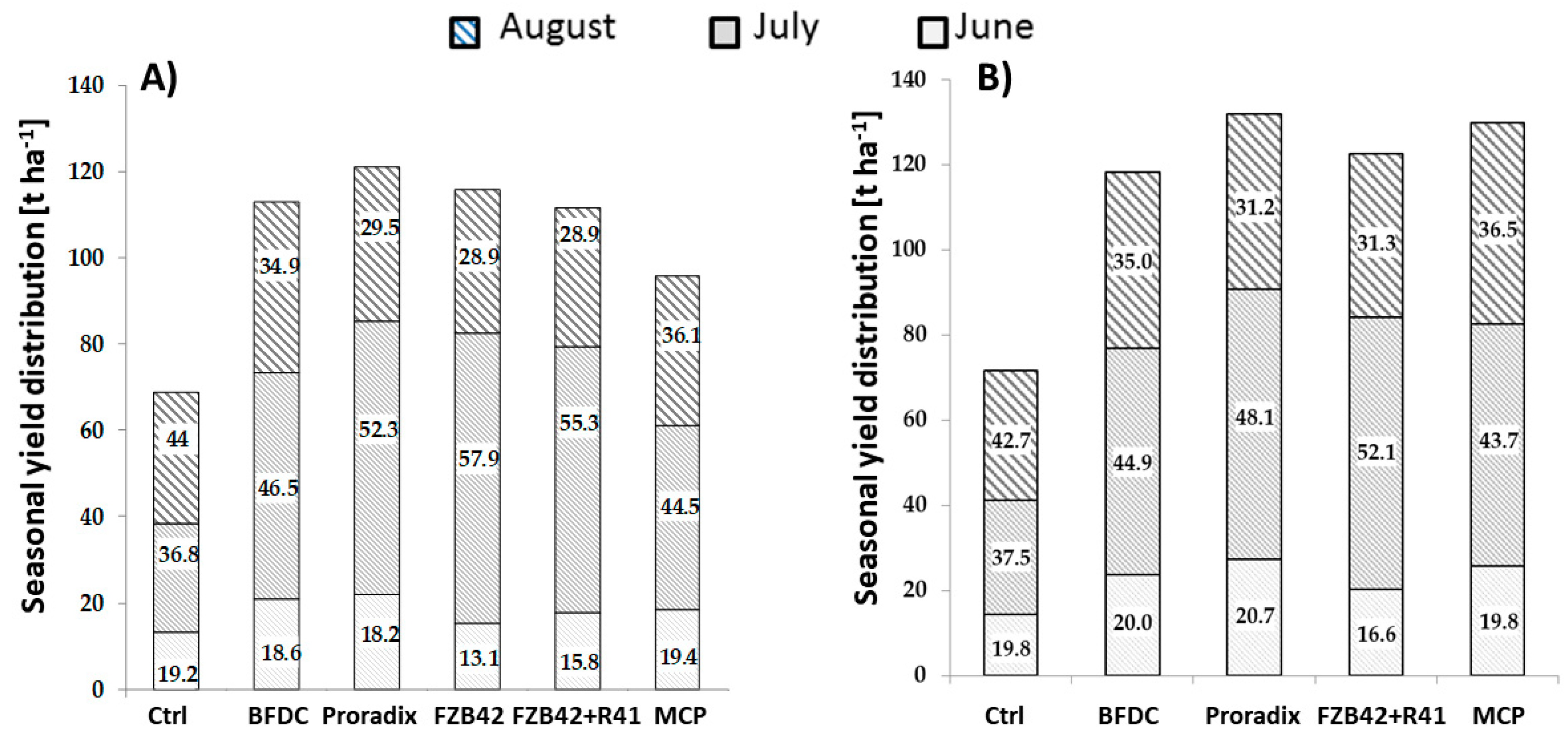
3.1.3. Distribution of Fruit Size and Seasonal Yield
3.2. Case Study 2: Open Field Tomato Production with Drip Fertigation and Fertilizer Placement, Ramat Negev Desert, Israel, 2017
3.2.1. Vegetative Growth and Phosphate Status
3.2.2. Fruit Yield
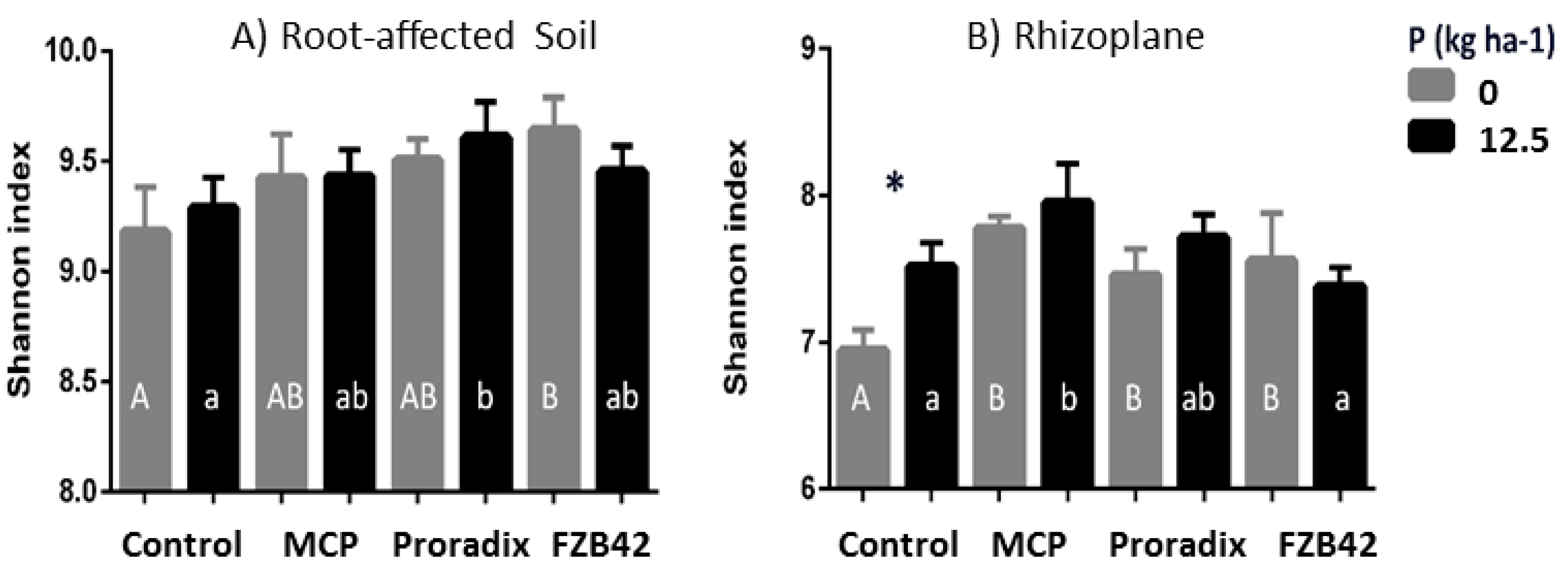
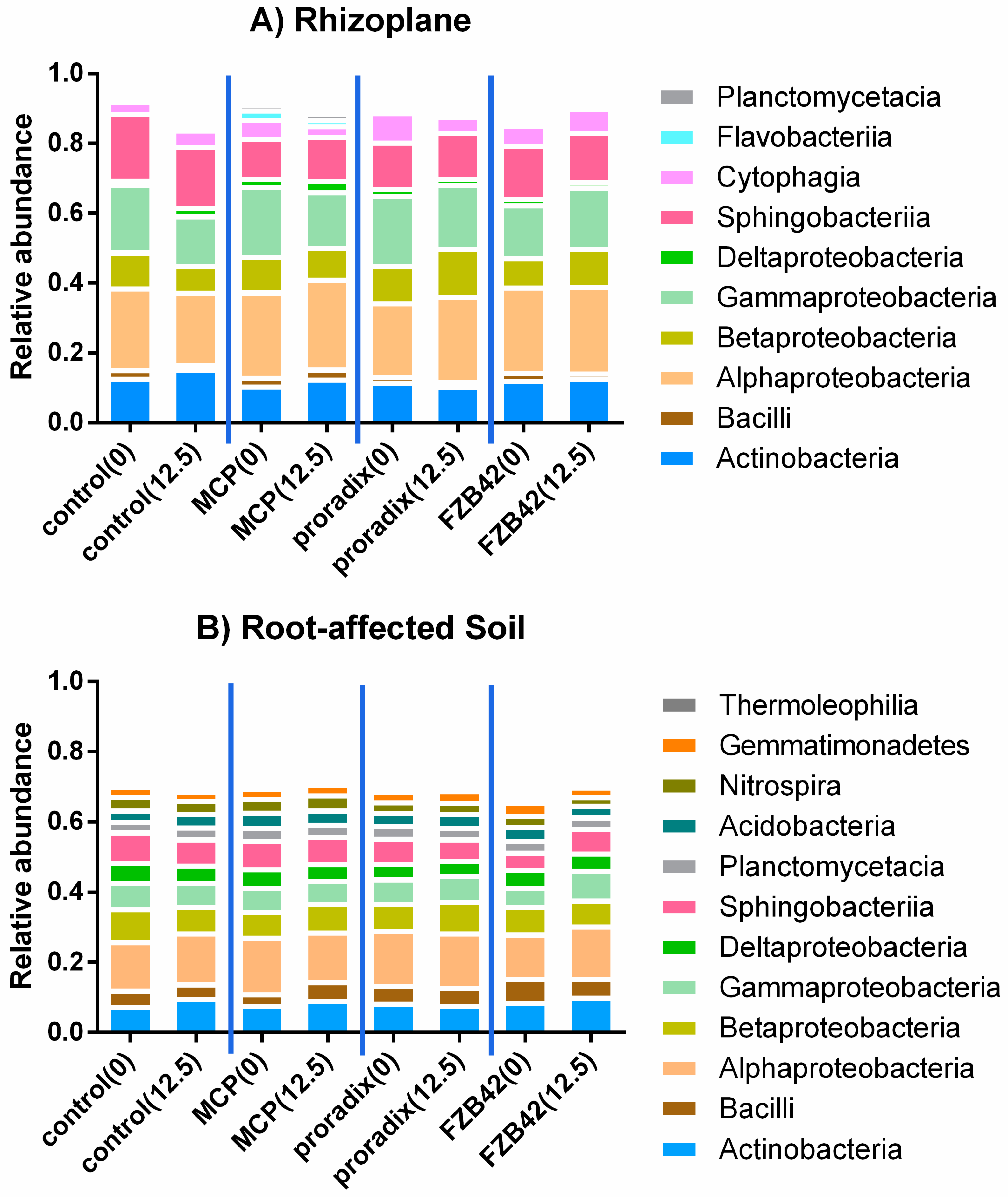
3.2.3. Microbiome Interactions
4. Discussion
4.1. Case Study I: Large-Scale Greenhouse Experiments Timisoara, Romania, 2016/2017
4.1.1. Nursery and Vegetative Growth
4.1.2. Generative Growth and Fruit Yield
Single Strains versus Microbial Consortia
4.2. Case Study II: Open Field Tomato prOduction with Drip Fertigation and Fertilizer Placement, Ramat Negev Desert, Israel, 2017
4.2.1. Vegetative Plant Growth and Yield Responses
4.2.2. Interactions with the Soil Microbiome
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nuti, M.; Giovannetti, G. Borderline Products between Bio-fertilizers/ Bio-effectors and Plant Protectants: The Role of Microbial Consortia. J. Agric. Sci. Technol. A 2015, 5, 305–315. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in Plant Science: A Global Perspective. Front. Plant Sci. 2017, 7, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Michalak, I.; Dmytryk, A.; Schroeder, G.; Chojnacka, K. The Application of Homogenate and Filtrate from Baltic Seaweeds in Seedling Growth Tests. Appl. Sci. 2017, 7, 230. [Google Scholar] [CrossRef]
- Rouphael, Y.; Giordano, M.; Cardarelli, M.; Cozzolino, E.; Mori, M.; Kyriacou, M.C.; Bonini, P.; Colla, G. Plant- and Seaweed-Based Extracts Increase Yield but Differentially Modulate Nutritional Quality of Greenhouse Spinach through Biostimulant Action. Agronomy 2018, 8, 126. [Google Scholar] [CrossRef]
- Wilson, H.T.; Amirkhani, M.; Taylor, A.G. Evaluation of Gelatin as a Biostimulant Seed Treatment to Improve Plant Performance. Front. Plant Sci. 2018, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Amirkhani, M.; Netravali, A.N.; Huang, W.; Taylor, A.G. Investigation of Soy Protein—Based Biostimulant Seed Coating for Broccoli Seedling and Plant Growth Enhancement. HortScience 2016, 51, 1121–1126. [Google Scholar] [CrossRef]
- Tilman, D.; Fargione, J.; Wolff, B.; Antonio, C.D.; Dobson, A.; Howarth, R.; Schindler, D.; Schlesinger, W.H.; Simberloff, D.; Swackhamer, D. Forecasting Agriculturally Driven Global Environmental Change. Science 2001, 292, 281–284. [Google Scholar] [CrossRef]
- Xu, H.-L.; Wang, R.; Mridha, M.A.U. Effects of Organic Fertilizers and a Microbial Inoculant on Leaf Photosynthesis and Fruit Yield and Quality of Tomato Plants. J. Crop Prod. 2001, 3, 173–182. [Google Scholar] [CrossRef]
- Esitken, A.; Yildiz, H.E.; Ercisli, S.; Figen Donmez, M.; Turan, M.; Gunes, A. Effects of plant growth promoting bacteria (PGPB) on yield, growth and nutrient contents of organically grown strawberry. Sci. Hortic. 2010, 124, 62–66. [Google Scholar] [CrossRef] [Green Version]
- Abbasi, M.K.; Musa, N.; Manzoor, M. Mineralization of soluble P fertilizers and insoluble rock phosphate in response to phosphate-solubilizing bacteria and poultry manure and their effect on the growth and P utilization efficiency of chilli (Capsicum annuum L.). Biogeosciences 2015, 12, 4607–4619. [Google Scholar] [CrossRef]
- Thonar, C.; Duus, J.; Lekfeldt, S.; Cozzolino, V.; Kundel, D.; Kulhánek, M.; Mosimann, C.; Neumann, G.; Piccolo, A.; Rex, M.; et al. Potential of three microbial bio-effectors to promote maize growth and nutrient acquisition from alternative phosphorous fertilizers in contrasting soils. Chem. Biol. Technol. Agric. 2017, 1–16. [Google Scholar] [CrossRef]
- Hartmann, A.; Rothballer, M.; Schmid, M. Lorenz Hiltner, a pioneer in rhizosphere microbial ecology and soil bacteriology research. Plant Soil 2008, 312, 7–14. [Google Scholar] [CrossRef]
- Lopez-Cervantes, J.; Thorpe, D.T. Microbial Composition Comprising Liquid Fertilizer and Processes for Agricultural Use. Agrinos, AS. United States Patent Application Publication US 2013/0255338 A1, 3 October 2013. [Google Scholar]
- Sekar, J.; Raj, R.; Prabavathy, V.R. Microbial Consortial Products for Sustainable Agriculture: Commercialization and Regulatory Issues in India. In Agriculturally Important Microorganisms; Singh, H.B., Sarma, B.K., Keswani, C., Eds.; Springer Science+Business Media: Singapore, 2016; pp. 107–131. [Google Scholar]
- Higa, T.; Parr, J.F. Beneficial and Effective Microorganisms for a Sustainable Agriculture and Environment; International Nature Farming Research Center Atami: Okinawa, Japan, 1994; pp. 1–16. [Google Scholar]
- Hadar, Y. Suppressive compost: When plant pathology met microbial ecology. Phytoparasitica 2011, 39, 311–314. [Google Scholar] [CrossRef]
- Carvalhais, L.C.; Muzzi, F.; Tan, C.-H.; Hsien-Choo, J.; Schenk, P.M. Plant growth in Arabidopsis is assisted by compost soil-derived microbial communities. Front. Plant Sci. 2013, 4, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Bashan, Y. Inoculants of plant growth-promoting bacteria for use in agriculture. Biotechnol. Adv. 1998, 16, 729–770. [Google Scholar] [CrossRef]
- Edathil, T.T.; Manian, S.; Udaiyan, K. Interaction of multiple VAM fungal species on root colonization, plant growth and nutrient status of tomato seedlings. Agric. Ecosyst. Environ. 1996, 59, 63–68. [Google Scholar] [CrossRef]
- Fröhlich, A.; Buddrus-Schiemann, K.; Durner, J.; Hartmann, A.; Rad, U. Von Response of barley to root colonization by Pseudomonas sp. DSMZ 13134 under laboratory, greenhouse, and field conditions. J. Plant Interact. 2012, 7, 1–9. [Google Scholar] [CrossRef]
- Borris, R. Towards a New Generation of Commercial Microbial Disease Control and Plant Growth Promotion Products. In Principles of Plant-Microbe Interactions; Lugtenberg, B., Ed.; Springer International Publishing: Basel, Switzerland, 2015; pp. 329–337. [Google Scholar]
- Nkebiwe, P.M.; Weinmann, M.; Müller, T. Improving fertilizer—Depot exploitation and maize growth by inoculation with plant growth-romoting bacteria: From lab to field. Chem. Biol. Technol. Agric. 2016, 3, 1–16. [Google Scholar] [CrossRef]
- Mpanga, I.K.; Dapaah, H.K.; Geistlinger, J.; Ludewig, U. Soil Type-Dependent Interactions of P-Solubilizing Microorganisms with Organic and Inorganic Fertilizers Mediate Plant Growth Promotion in Tomato. Agronomy 2018, 8, 213. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornischer, U.; Koller, M. Biologischer Anbau von Tomaten. Bioland Beratung GmbH Kompetenzzentrum Ökolandbau Niedersachsen FiBl/KÖN/Bioland, Germany. 2005. Available online: https://www.bioland.de/fileadmin/dateien/HP_Dokumente/Verlag/MB_Tomaten.pdf (accessed on 20 December 2018).
- Marschner, P. Marschner’s Mineral Nutrition of Higher Plants Third Edition; Elsevier Academic Press: San Diego, CA, USA, 2012; ISBN 9780123849052. [Google Scholar]
- Nielsen, K.L.; Thorup-Kristensen, K. Growing media for organic tomato plantlet production Archived at http://orgprints.org/00001606. Acta Hortic. 2001, 664, 183–188. [Google Scholar] [CrossRef]
- Tiquia, S.M.; Tam, N.F.Y.; Hodgkiss, I.J. Effects of composting on phytotoxicity of spent pig-manure sawdust litter. Environ. Pollut. 1996, 93, 249–256. [Google Scholar] [CrossRef]
- Loecke, T.D.; Liebman, M.; Cambardella, C.A.; Richard, T.L. Corn Growth Responses to Composted and Fresh Solid Swine Manures. Crop Sci. 2004, 44, 177–184. [Google Scholar] [CrossRef]
- Gulden, R.H.; Vessey, J.K. Penicillium bilaii inoculation increases root-hair production in field pea. Can. J. Plant Sci. 2000, 80, 801–804. [Google Scholar] [CrossRef] [Green Version]
- Leggett, M.; Newlands, N.; Greenshields, D.; West, L.; Inman, S.; Koivunen, M. Maize yield response to a phosphorus-solubilizing microbial inoculant in field trials Maize yield response to a phosphorus-solubilizing microbial inoculant in field trials. J. Agric. Sci. 2014, 1–15. [Google Scholar] [CrossRef]
- Nkebiwe, P.M.; Neumann, G.; Müller, T. Densely rooted rhizosphere hotspots induced around subsurface NH4+-fertilizer depots: A home for soil PGPMs? Chem. Biol. Technol. Agric. 2017, 4, 1–16. [Google Scholar] [CrossRef]
- Sánchez-Esteva, S.; Muñoz, B.G.; Jensen, L.S.; De Neergaard, A.; Magid, J. The effect of Penicillium bilaii on wheat growth and phosphorus uptake as affected by soil pH, soil P and application of sewage sludge. Chem. Biol. Technol. Agric. 2016, 3, 1–11. [Google Scholar] [CrossRef]
- Von Rad, U.; Mueller, M.J.; Durner, J. Evaluation of natural and synthetic stimulants of plant immunity by microarray technology. New Phytol. 2005, 165, 191–202. [Google Scholar] [CrossRef]
- Meng, Q. Characterization of Bacillus amyloliquefaciens Strain BAC03 in Disease Control and Plant Growth Promotion. Ph.D. Thesis, Institute of Plant Pathology, Michigan State University, East Lansing, MI, USA, 2014. [Google Scholar]
- Xie, S.; Jiang, H.; Ding, T.; Xu, Q.; Chai, W.; Cheng, B. Bacillus amyloliquefaciens FZB42 represses plant miR846 to induce systemic resistance via a jasmonic acid-dependent signalling pathway. Mol. Plant Pathol. 2018, 19, 1612–1623. [Google Scholar] [CrossRef]
- Suen, P.-K.; Zhang, S.; Sun, S.S.M. Molecular characterization of a tomato purple acid phosphatase during seed germination and seedling growth under phosphate stress. Plant Cell Rep. 2015, 34, 981–992. [Google Scholar] [CrossRef] [PubMed]
- Pilbeam, D.J.; Cakmak, I.; Marschner, H.; Kirkby, E.A. Effect of withdrawal of phosphorus on nitrate assimilation and PEP carboxylase activity in tomato. Plant Soil 1993, 154, 111–117. [Google Scholar] [CrossRef]
- Neumann, G.; Römheld, V. Root excretion of carboxylic acids and protons in phosphorus-deficient plants. Plant Soil 1999, 211, 121–130. [Google Scholar] [CrossRef]
- Yusran, Y.; Roemheld, V.; Mueller, T. Effects of Pseudomonas sp. ”Proradix” and Bacillus amyloliquefaciens FZB42 on the Establishment of AMF Infection, Nutrient Acquisition and Growth of Tomato Affected by Fusarium oxysporum Schlecht f.sp. radicis-lycopersici Jarvis and Shoemaker. In Proceedings of the International Plant Nutrition Colloquium XVI, California Digital Library, University of California, Davis, CA, USA, 31 July 2009. [Google Scholar]
- Chen, J. The Combined Use of Chemical and Organic Fertilizers and/or Biofertilizer for Crop Growth and Soil Fertility. In Proceedings of the International Workshop on Sustained Management of the Soil-Rhizosphere System for Efficient Crop Production and Fertilizer Use, Land Development Department, Bangkok, Thailand, 16–20 October 2006; pp. 1–11. [Google Scholar]
- Xu, H.; Wang, R.; Mridha, A.U. Effects of Organic Fertilizers and a Microbial Inoculant on Leaf Photosynthesis and Fruit Yield and Quality of Tomato Plants Effects of Organic Fertilizers and a Microbial Inoculant on Leaf Photosynthesis and Fruit Yield and Quality of Tomato Plants. J. Crop Prod. 2008, 3, 173–182. [Google Scholar] [CrossRef]
- Chowdhury, S.P.; Dietel, K.; Rändler, M.; Schmid, M.; Junge, H.; Borriss, R.; Hartmann, A.; Grosch, R. Effects of Bacillus amyloliquefaciens FZB42 on Lettuce Growth and Health under Pathogen Pressure and Its Impact on the Rhizosphere Bacterial Community. PLoS ONE 2013, 8, e68818. [Google Scholar] [CrossRef]
- Srivastava, A.; Handa, A.K. Hormonal Regulation of Tomato Fruit Development: A Molecular Perspective. J. Plant Growth Regul. 2005, 24, 67–82. [Google Scholar] [CrossRef]
- Alam, S.M. Fruit Yield of Tomato as Affected by NAA Spray. Asian J. Plant Sci. 2002, 1, 1–5. [Google Scholar] [CrossRef]
- Sarkar, M.D.; Jahan, M.S.; Kabir, M.H.; Kabir, K.; Rojoni, R.N. Flower and Fruit Setting of Summer Tomato Regulated by Plant Hormones. Appl. Sci. Rep. 2014, 7, 117–120. [Google Scholar] [CrossRef]
- Buddrus-Schiemann, K.E.M. Wirkung des biologischen Pflanzenstärkungsmittels Proradix® (Pseudomonas fluorescens) auf das Wachstum von Gerste (Hordeum vulgare L. cv. Barke) und auf die bakterielle Gemeinschaft in der Rhizosphäre. Ph.D. Thesis, Ludwig-Maximilians-University, Munich, Germany, 2008. [Google Scholar]
- Oancea, F.; Raut, I.; Zamfiropol-Cristea, V. Influence of soil treatment with microbial plant biostimulant on tomato yield and quality. Agric. Food 2017, 156–165. [Google Scholar]
- Ownley, B.H.; Seth, D.; Hamilton, C.; Dee, M. Effects of Plant-Growth-Promoting-Rhizobacteria on Biomass, Flowering, and YIELD of Field Tomatoes. In Extension—Vegetable Production; University of Tennesee, Institute of Agriculture: Knoxville, TN, USA, 1999. [Google Scholar]
- Murphy, J.F.; Reddy, M.S.; Ryu, C.; Kloepper, J.W.; Li, R. Rhizobacteria-Mediated Growth Promotion of Tomato Leads to Protection Against Cucumber mosaic virus. Phytopathology 2003, 93, 1301–1307. [Google Scholar] [CrossRef]
- Karakurt, H.; Kotan, R.; Dadaşoğlu, F.; Aslantaş, R.; Şahİn, F. Effects of plant growth promoting rhizobacteria on fruit set, pomological and chemical characteristics, color values, and vegetative growth of sour cherry (Prunus cerasus cv. Kütahya ). Turk. J. Biol. 2011, 35, 283–291. [Google Scholar] [CrossRef]
- Jing, J.; Rui, Y.; Zhang, F.; Rengel, Z.; Shen, J. Localized application of phosphorus and ammonium improves growth of maize seedlings by stimulating root proliferation and rhizosphere acidification. Field Crops Res. 2010, 119, 355–364. [Google Scholar] [CrossRef]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. Springer Plus 2013, 2, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H.; Schropp, A. Vergleichende Untersuchungen über die Empfindlichkeit von 6 Unterlagensorten der Weinrebe gegenüber Phosphat- induziertem Zink-Mangel. Vitis 1977, 16, 79–88. [Google Scholar]
- Cakmak, I.; Marschner, H. Mechanism of phosphorus-induced zinc deficiency in cotton. III. Changes in physiological availability of zinc in plants. Physiol. Plant. 1987, 70, 13–20. [Google Scholar] [CrossRef]
- Marilley, L.; Vogt, G.; Blanc, M.; Aragno, M. Bacterial diversity in the bulk soil and rhizosphere fractions of Lolium perenne and Trifolium repens as revealed by PCR restriction analysis of 16S rDNA. Plant Soil 1998, 198, 219–224. [Google Scholar] [CrossRef]
- Berg, G.; Zachow, C.; Lottmann, J.; Go, M.; Costa, R.; Icrobiol, A.P.P.L.E.N.M. Impact of Plant Species and Site on Rhizosphere-Associated Fungi Antagonistic to Verticillium dahliae Kleb. Appl. Environ. Microbiol. 2005, 71, 4203–4213. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Marsh, E.L.; Ainsworth, E.A.; Leakey, A.D.B.; Amy, M.; Schachtman, D.P. Shifts in microbial communities in soil, rhizosphere and roots of two major crop systems under elevated CO2 and O3. Sci. Rep. 2017, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Castro-Sowinski, S.; Herschkovitz, Y.; Okon, Y.; Jurkevitch, E. Effects of inoculation with plant growth-promoting rhizobacteria on resident rhizosphere microorganisms. FEMS Microbiol. Lett. 2007, 276, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Neumann, G.; Römheld, V. The release of root exudates as affected by the plant physiological status. In The Rhizosphere: Biochemistry and Organic Substances at the Soil-Plant Interface, 2nd ed.; Pinton, R., Varanini, Z., Nannipieri, Z., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 23–72. [Google Scholar]
- Lydell, C.; Dowell, L.; Sikaroodi, M.; Gillevet, P.; Emerson, D. A Population Survey of Members of the Phylum Bacteroidetes Isolated from Salt Marsh Sediments along the East Coast of the United States. Microb. Ecol. 2004, 48, 263–273. [Google Scholar] [CrossRef]
- Lucas, J.A.; Villaraco, A.G.; Ramos, B.; Garcia-Cristobal, J.; Algar, E.; Gutierrez-ma, J. Structural and functional study in the rhizosphere of Oryza sativa L. plants growing under biotic and abiotic stress. J. Appl. Microbiol. 2013, 115, 218–235. [Google Scholar] [CrossRef] [PubMed]
- Riley, D.; Barber, S.A. Salt accumulation at the soybean (Glycine max. (L.) merr.) root-soil interface. Soil Sci. Soc. Am. J. 1970, 34, 154–155. [Google Scholar] [CrossRef]
- Huang, B.; North, G.B.; Nobel, P.S. Soil Sheaths, Photosynthate Distribution to Roots, and Rhizosphere Water Relations for Opuntia ficus-indica. Int. J. Plant Sci. 1993, 154, 425–431. [Google Scholar] [CrossRef]
- Kaushal, M.; Wani, S.P. Plant-growth-promoting rhizobacteria: Drought stress alleviators to ameliorate crop production in drylands. Ann. Microbiol. 2015, 66, 35–42. [Google Scholar] [CrossRef]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; Skz, A. Rhizosphere Multifunctional Pseudomonas putida strain FBKV2 from arid rhizosphere soil and its growth promotional effects on maize under drought stress. Rhizosphere 2016, 1, 4–13. [Google Scholar] [CrossRef]
- Kasim, W.A.; Gaafar, R.M.; Abou-ali, R.M.; Omar, M.N.; Hewait, H.M. Effect of biofilm forming plant growth promoting rhizobacteria on salinity tolerance in barley. Ann. Agric. Sci. 2016, 61, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Bloemberg, G.V.; Lugtenberg, B.J.J. Molecular basis of plant growth promotion and biocontrol by rhizobacteria. Curr. Opin. Plant Biol. 2001, 4, 343–350. [Google Scholar] [CrossRef]
- Mayak, S.; Tirosh, T.; Glick, B.R. Plant growth-promoting bacteria confer resistance in tomato plants to salt stress. Plant Physiol. Biochem. 2004, 42, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Soltani, A.-A. Plant Growth Promoting Characteristics in Some Flavobacterium spp. Isolated from Soils of Iran. J. Agric. Sci. 2010, 2, 106–115. [Google Scholar] [CrossRef]
- Gontia-Mishra, I.; Sapre, S.; Sharma, A.; Tiwari, S. Amelioration of drought tolerance in wheat by the interaction of plant growth-promoting rhizobacteria. Plant Biol. 2016, 18, 992–1000. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M.; Pathology, P. Trichoderma Species—Opportunistic, Avirulent Plant Symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P Dose | Biostimulant | Shoot | Root | Root | Shoot P | Shoot P |
|---|---|---|---|---|---|---|
| Biomass | Length | Concentration | Content | |||
| kg ha−1 | g plant−1 | g plant−1 | m plant−1 | mg g−1 | mg plant−1 | |
| 0 | Control | 300e | 50.5 | 54ab | 0.51g | 23e |
| 0 | Proradix | 340e | 57.0 | 55ab | 0.61fg | 31de |
| 0 | FZB42 | 350de | 62.8 | 63ab | 0.67efg | 36cde |
| 0 | CFB | 260e | 62.0 | 71ab | 0.70efg | 27de |
| 0 | MCP | 640bc | 36.7 | 46b | 0.72efg | 69abcde |
| 12.5 | Control | 420bcde | 51.2 | 47ab | 0.78defg | 49cde |
| 12.5 | Proradix | 630bcd | 42.2 | 42b | 0.83def | 78abcde |
| 12.5 | FZB42 | 400cde | 46.3 | 58ab | 0.87cdef | 53bcde |
| 12.5 | CFB | 430bcde | 65.7 | 59ab | 0.93cde | 59bcde |
| 12.5 | MCP | 500bcde | 78.1 | 60ab | 0.97bcde | 73abcde |
| 50 | Control | 620bcd | 44.5 | 45b | 1.01bcde | 103abcde |
| 50 | Proradix | 670bc | 62.4 | 62ab | 1.07bcd | 106abcd |
| 50 | FZB42 | 680ab | 58.6 | 70ab | 1.16bc | 119abc |
| 50 | CFB | 770a | 43.3 | 43b | 1.28b | 148a |
| 50 | MCP | 500bcde | 67.0 | 81a | 1.87a | 139ab |
| Analysis of Variance | ||||||
| df | Shoot | Root | Root Length | Shoot P | ||
| Fresh weight | concentration | content | ||||
| Treatment | 14 | ** | ns | * | *** | *** |
| block | 3 | ns | ns | ns | ns | ns |
| P Dose | Bio Stimulant | Red Fruits | Green Fruits | Small Fruits | Total Yield | ||||
|---|---|---|---|---|---|---|---|---|---|
| kg ha−1 | t ha−1 | No | t ha−1 | No | t ha−1 | No | t ha−1 | No | |
| 0 | Control | 14.4e | 13.5d | 1.00 | 1.00 | 1.76 | 4.2 | 17.2b | 187d |
| 0 | Proradix | 21.7cde | 19.8bcd | 0.89 | 0.91 | 0.70 | 1.6 | 23.3ab | 223bcd |
| 0 | FZB42 | 25.9 bcde | 23.9abcd | 0.69 | 0.69 | 0.72 | 1.6 | 27.3ab | 262abcd |
| 0 | CFB | 15.2de | 16.1cd | 0.74 | 0.72 | 1.53 | 3.7 | 17.5b | 205cd |
| 0 | MCP | 33.2bcde | 27.5abcd | 1.15 | 1.13 | 1.44 | 2.9 | 35.8a | 315abcd |
| 12.5 | Control | 36.1bcde | 30.3abcd | 0.30 | 0.25 | 0.34 | 0.7 | 36.7a | 312abcd |
| 12.5 | Proradix | 40.7bcd | 27.8abcd | 0.38 | 0.34 | 0.44 | 1.1 | 41.5a | 293abcd |
| 12.5 | FZB42 | 33.1abcde | 30.8abcd | 0.00 | 0.00 | 0.00 | 0.0 | 33.1a | 308abcd |
| 12.5 | CFB | 31.9abcde | 27.7abcd | 0.29 | 0.31 | 0.57 | 1.6 | 31.7a | 296abcd |
| 12.5 | MCP | 45.5bc | 32.4abc | 0.29 | 0.34 | 0.49 | 1.3 | 46.3a | 341abcd |
| 50 | Control | 56.7a | 40.6a | 0.00 | 0.00 | 0.42 | 0.9 | 57.1a | 415a |
| 50 | Proradix | 57.0a | 40.7a | 0.35 | 0.44 | 0.43 | 1.1 | 57.8a | 422a |
| 50 | FZB42 | 47.3abc | 37.0ab | 0.07 | 0.13 | 0.05 | 0.2 | 47.4a | 374ab |
| 50 | CFB | 52.9a | 34.9ab | 0.12 | 0.09 | 0.02 | 0.1 | 53.0a | 350abc |
| 50 | MCP | 50.8ab | 36.1ab | 0.08 | 0.09 | 0.13 | 0.6 | 51.1a | 368ab |
| Analysis of Variance | |||||||||
| df | Red fruits | Green fruits | Small fruits | Total yield | |||||
| Fresh weight | No | Fresh weight | No | Fresh weight | No | Fresh weight | No | ||
| Treatment | 14 | * | * | ns | ns | * | * | * | * |
| block | 3 | ns | ns | * | * | * | * | ns | ns |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bradáčová, K.; Florea, A.S.; Bar-Tal, A.; Minz, D.; Yermiyahu, U.; Shawahna, R.; Kraut-Cohen, J.; Zolti, A.; Erel, R.; Dietel, K.; et al. Microbial Consortia versus Single-Strain Inoculants: An Advantage in PGPM-Assisted Tomato Production? Agronomy 2019, 9, 105. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9020105
Bradáčová K, Florea AS, Bar-Tal A, Minz D, Yermiyahu U, Shawahna R, Kraut-Cohen J, Zolti A, Erel R, Dietel K, et al. Microbial Consortia versus Single-Strain Inoculants: An Advantage in PGPM-Assisted Tomato Production? Agronomy. 2019; 9(2):105. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9020105
Chicago/Turabian StyleBradáčová, Klára, Andrea S. Florea, Asher Bar-Tal, Dror Minz, Uri Yermiyahu, Raneen Shawahna, Judith Kraut-Cohen, Avihai Zolti, Ran Erel, K. Dietel, and et al. 2019. "Microbial Consortia versus Single-Strain Inoculants: An Advantage in PGPM-Assisted Tomato Production?" Agronomy 9, no. 2: 105. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9020105