Plant Organellar DNA Polymerases Evolved Multifunctionality through the Acquisition of Novel Amino Acid Insertions




Abstract
:1. Introduction
2. Plant Organellar DNA Polymerase as a Replicative Polymerase
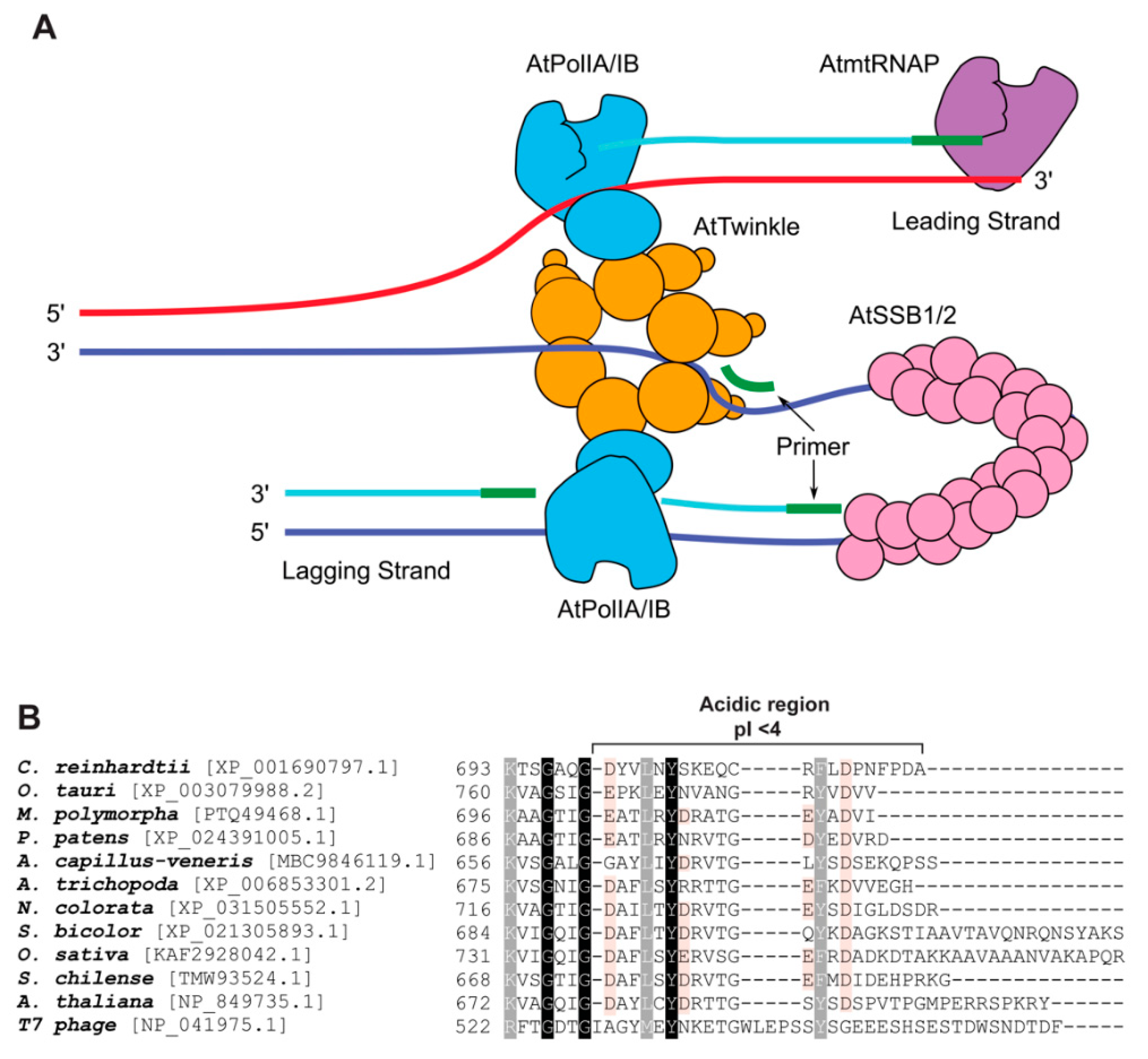
2.1. AtPOLIs Assemble a Replisome In Vitro
2.2. AtPOLIs Present Moderate Fidelity
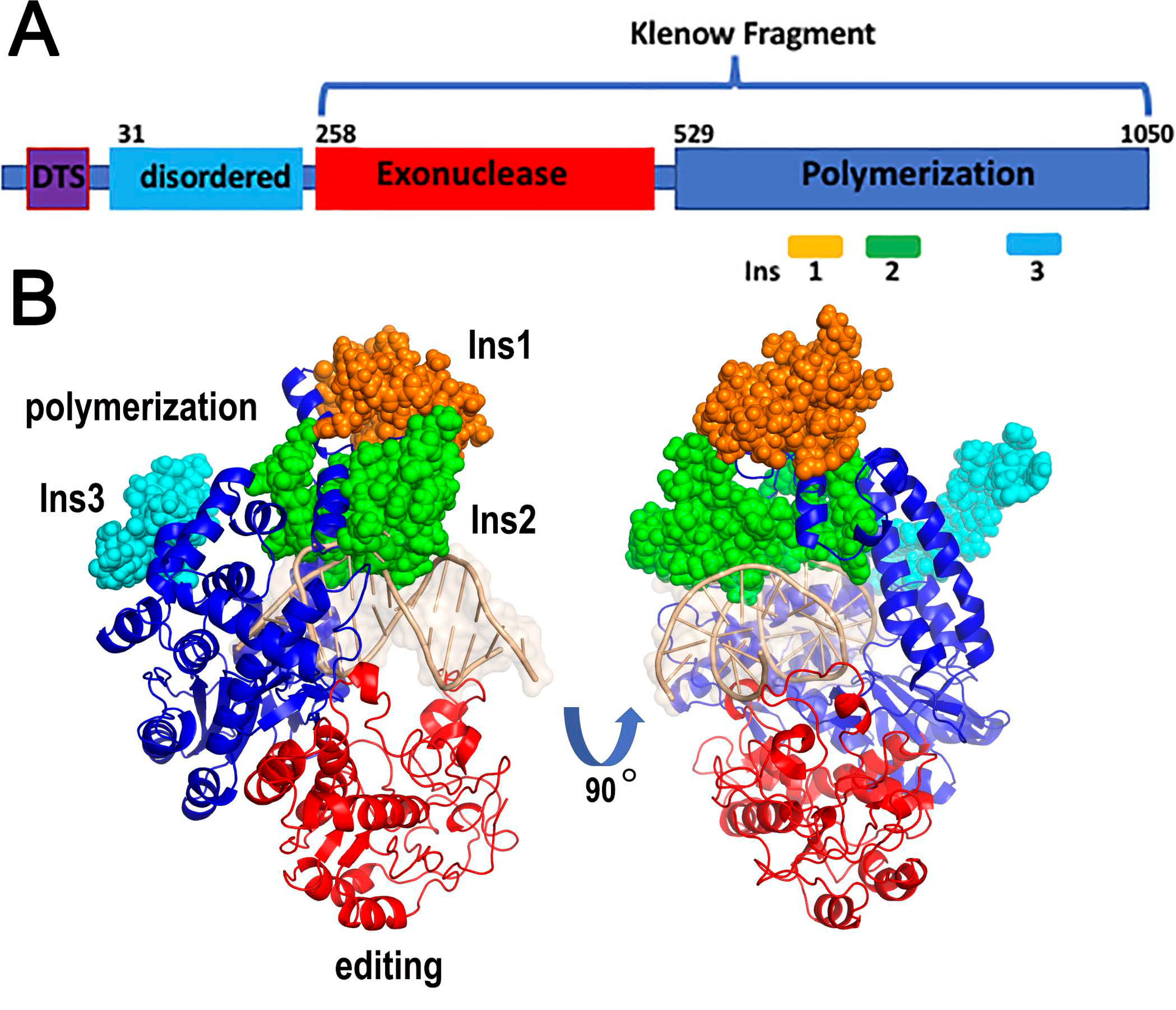
2.3. AtPOLIs Harbor an Insertion Poised to Confer Processivity
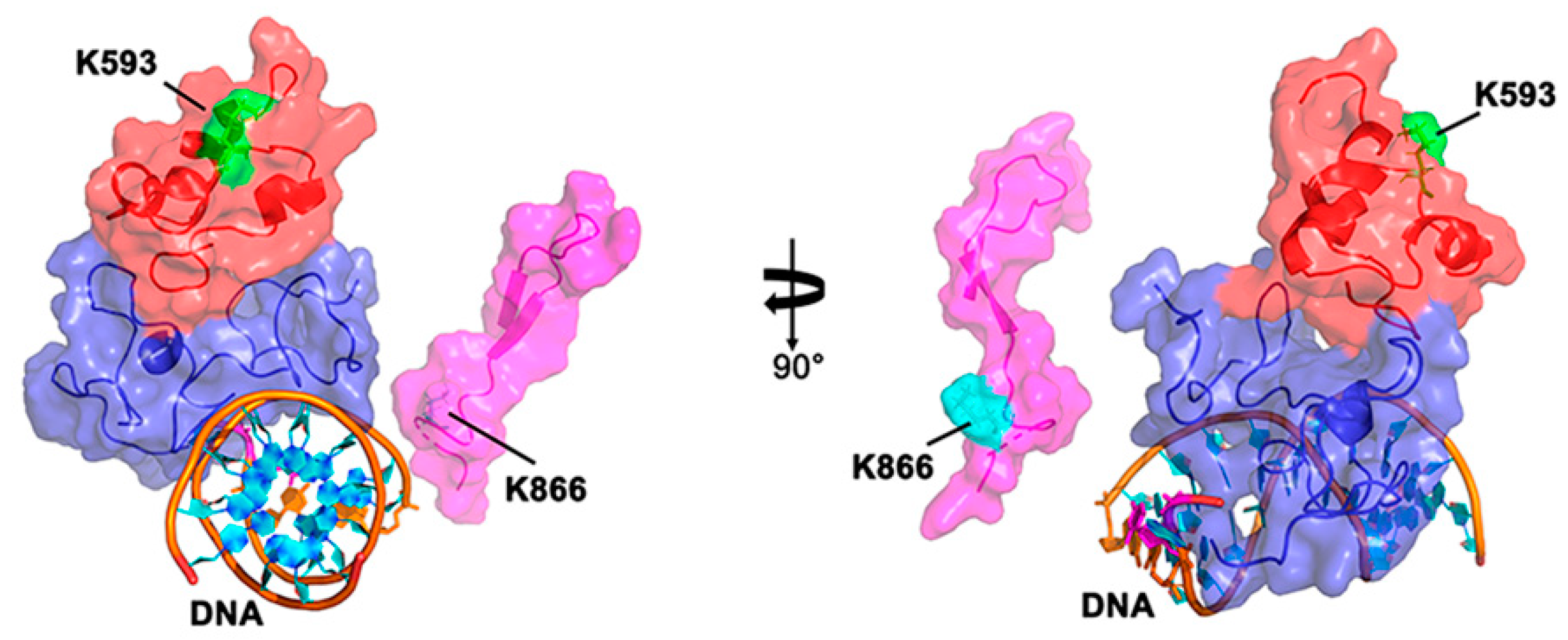
3. Plant Organellar DNA Polymerases Execute Translesion DNA Synthesis
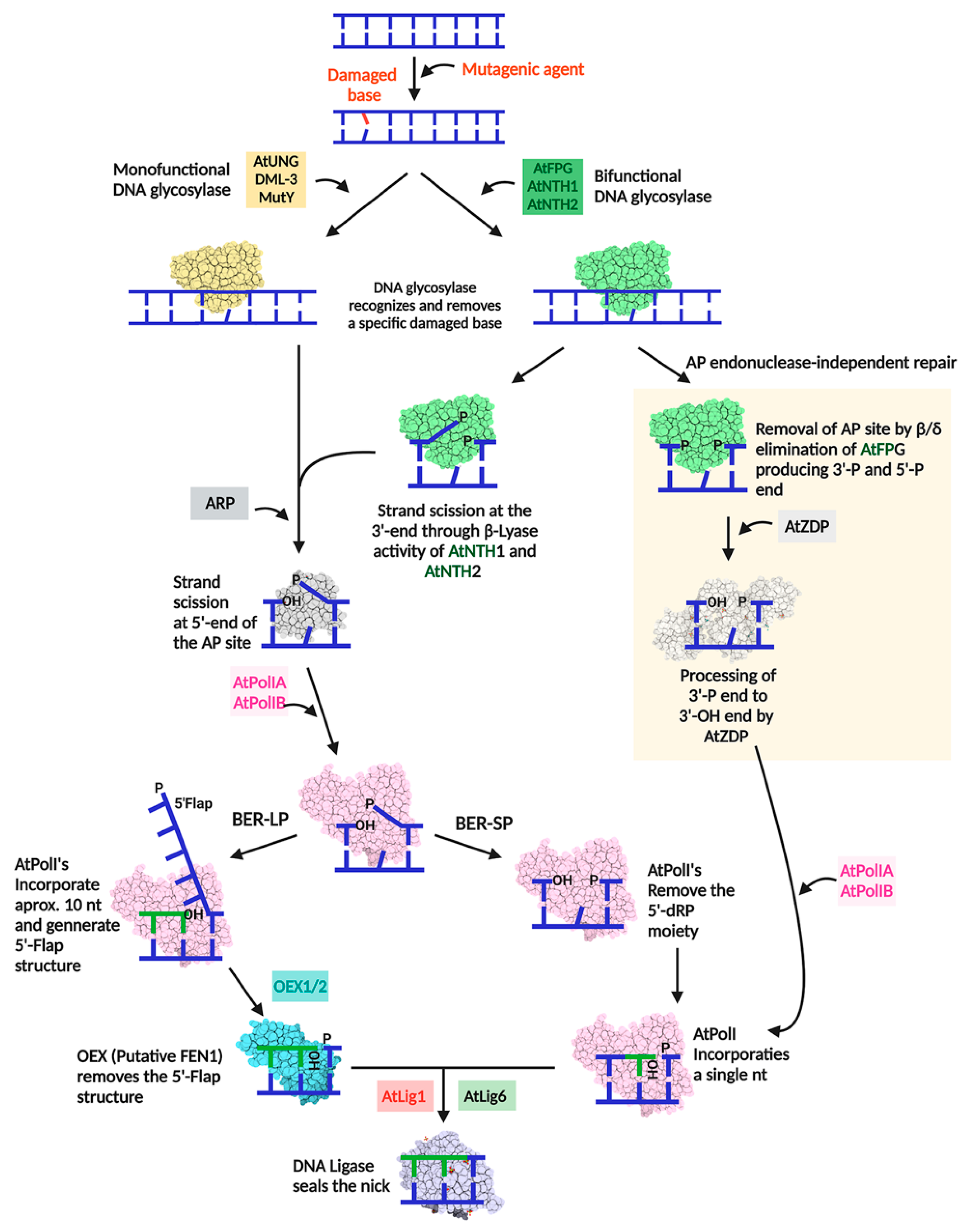
4. Plant Organellar DNA Polymerases Are Involved in Base Excision Repair
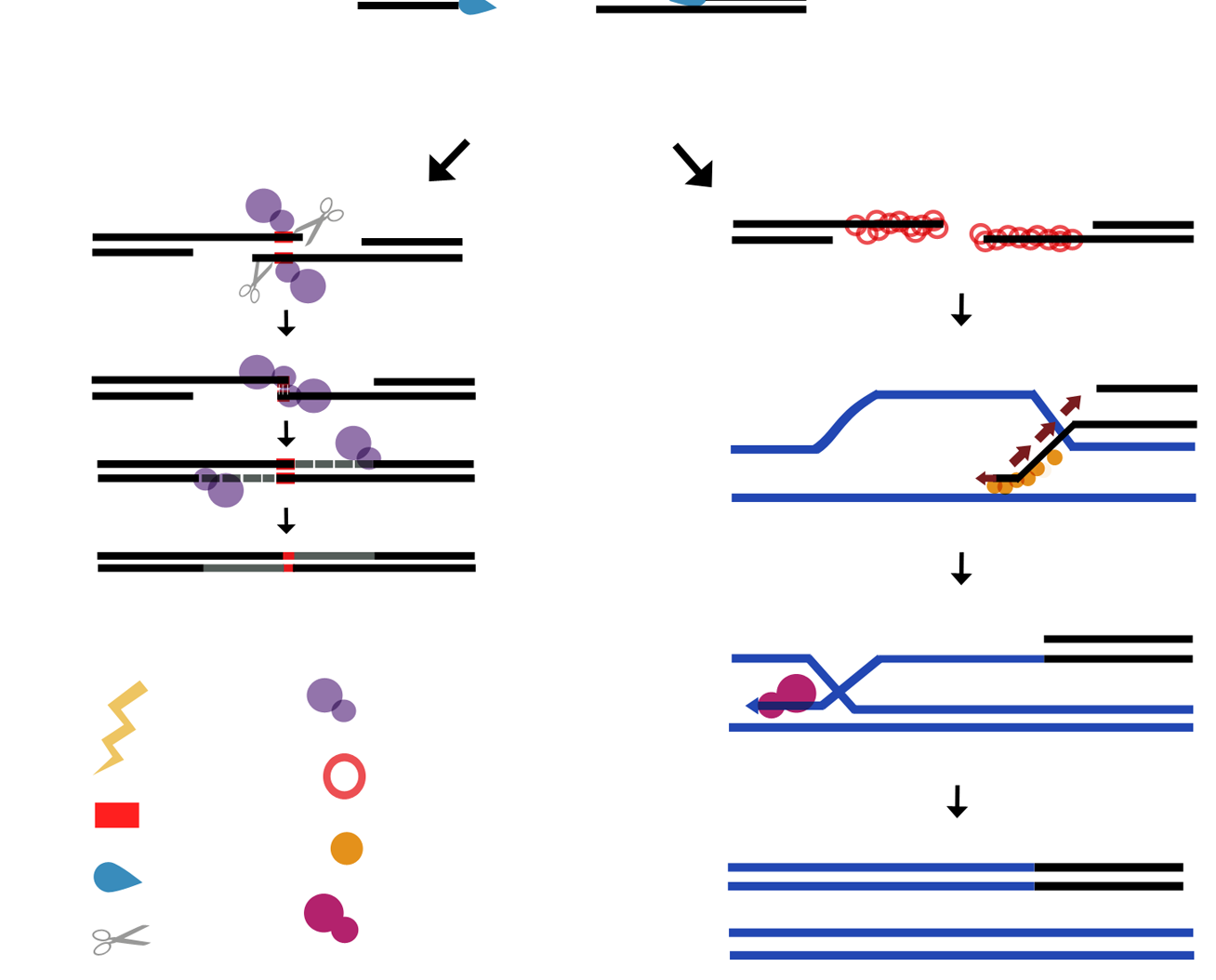
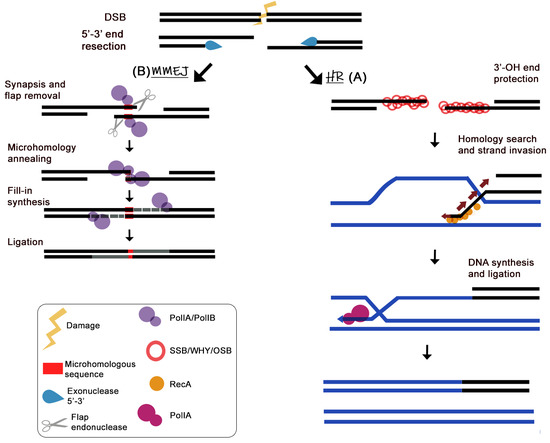
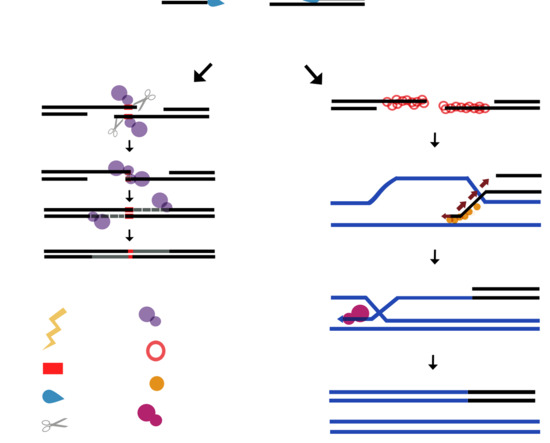
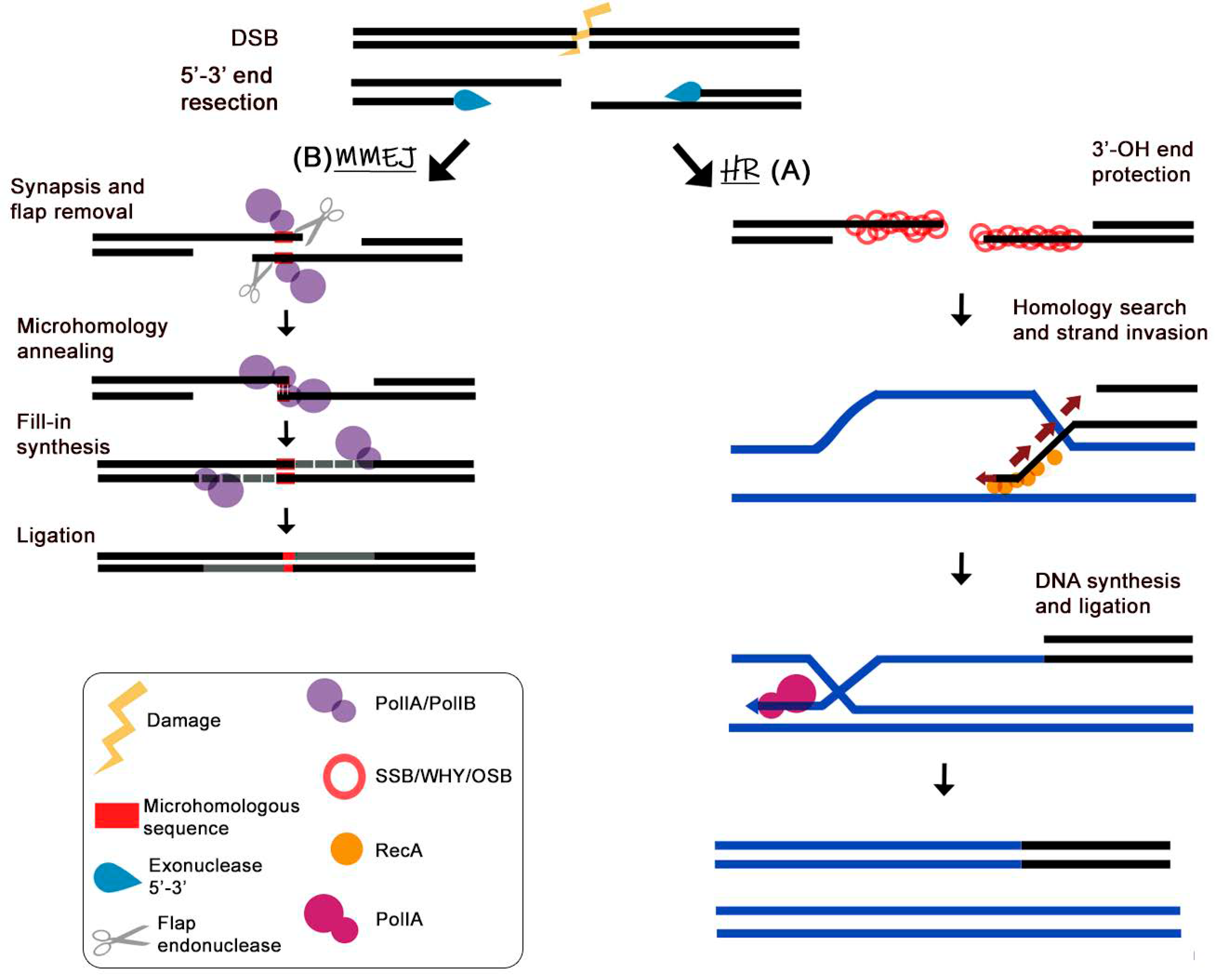
5. Plant Organellar DNA Polymerase Accomplish Micro-Homology-Mediated End-Joining of Double-Stranded Breaks
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gray, M.W. Lynn Margulis and the endosymbiont hypothesis: 50 years later. Mol. Biol. Cell 2017, 28, 1285–1287. [Google Scholar] [CrossRef]
- Oldenburg, D.J.; Bendich, A.J. DNA maintenance in plastids and mitochondria of plants. Front. Plant Sci. 2015, 6, 883. [Google Scholar] [CrossRef] [Green Version]
- Oldenburg, D.J.; Bendich, A.J. Most chloroplast DNA of maize seedlings in linear molecules with defined ends and branched forms. J. Mol. Biol. 2004, 335, 953–970. [Google Scholar] [CrossRef] [PubMed]
- Cermakian, N.; Ikeda, T.M.; Cedergren, R.; Gray, M.W. Sequences homologous to yeast mitochondrial and bacteriophage T3 and T7 RNA polymerases are widespread throughout the eukaryotic lineage. Nucleic Acids Res. 1996, 24, 648–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masters, B.S.; Stohl, L.L.; Clayton, D.A. Yeast mitochondrial RNA polymerase is homologous to those encoded by bacteriophages T3 and T7. Cell 1987, 51, 89–99. [Google Scholar] [CrossRef]
- Shutt, T.E.; Gray, M.W. Twinkle, the mitochondrial replicative DNA helicase, is widespread in the eukaryotic radiation and may also be the mitochondrial DNA primase in most eukaryotes. J. Mol. Evol. 2006, 62, 588–599. [Google Scholar] [CrossRef]
- Longley, M.J.; Nguyen, D.; Kunkel, T.A.; Copeland, W.C. The fidelity of human DNA polymerase γ with and without exonucleolytic proofreading and the p55 accessory subunit. J. Biol. Chem. 2001, 276, 38555–38562. [Google Scholar] [CrossRef] [Green Version]
- Viikov, K.; Valjamae, P.; Sedman, J. Yeast mitochondrial DNA polymerase is a highly processive single-subunit enzyme. Mitochondrion 2011, 11, 119–126. [Google Scholar] [CrossRef]
- Moriyama, T.; Terasawa, K.; Sato, N. Conservation of POPs, the plant organellar DNA polymerases, in eukaryotes. Protist 2011, 162, 177–187. [Google Scholar] [CrossRef]
- Moriyama, T.; Sato, N. Enzymes involved in organellar DNA replication in photosynthetic eukaryotes. Front. Plant Sci. 2014, 5, 480. [Google Scholar] [CrossRef] [Green Version]
- Mori, Y.; Kimura, S.; Saotome, A.; Kasai, N.; Sakaguchi, N.; Uchiyama, Y.; Ishibashi, T.; Yamamoto, T.; Chiku, H.; Sakaguchi, K. Plastid DNA polymerases from higher plants, Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2005, 334, 43–50. [Google Scholar] [CrossRef]
- Takeuchi, R.; Kimura, S.; Saotome, A.; Sakaguchi, K. Biochemical properties of a plastidial DNA polymerase of rice. Plant Mol. Biol. 2007, 64, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Christensen, A.C.; Lyznik, A.; Mohammed, S.; Elowsky, C.G.; Elo, A.; Yule, R.; Mackenzie, S.A. Dual-domain, dual-targeting organellar protein presequences in Arabidopsis can use non-AUG start codons. Plant Cell 2005, 17, 2805–2816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriyama, T.; Terasawa, K.; Fujiwara, M.; Sato, N. Purification and characterization of organellar DNA polymerases in the red alga Cyanidioschyzon merolae. FEBS J. 2008, 275, 2899–2918. [Google Scholar] [CrossRef] [PubMed]
- Parent, J.S.; Lepage, E.; Brisson, N. Divergent roles for the two PolI-like organelle DNA polymerases of Arabidopsis. Plant Physiol. 2011, 156, 254–262. [Google Scholar] [CrossRef] [Green Version]
- Cupp, J.D.; Nielsen, B.L. Arabidopsis thaliana organellar DNA polymerase IB mutants exhibit reduced mtDNA levels with a decrease in mitochondrial area density. Physiol. Plant. 2013, 149, 91–103. [Google Scholar] [CrossRef]
- Baruch-Torres, N.; Brieba, L.G. Plant organellar DNA polymerases are replicative and translesion DNA synthesis polymerases. Nucleic Acids Res. 2017, 45, 10751–10763. [Google Scholar] [CrossRef] [Green Version]
- Steitz, T.A. DNA polymerases: Structural diversity and common mechanisms. J. Biol. Chem. 1999, 274, 17395–17398. [Google Scholar] [CrossRef] [Green Version]
- Peralta-Castro, A.; Baruch-Torres, N.; Brieba, L.G. Plant organellar DNA primase-helicase synthesizes RNA primers for organellar DNA polymerases using a unique recognition sequence. Nucleic Acids Res. 2017, 45, 10764–10774. [Google Scholar] [CrossRef] [Green Version]
- Baruch-Torres, N.; Yamamoto, J.; Juarez-Quintero, V.; Iwai, S.; Brieba, L.G. Plant organellar DNA polymerases bypass thymine glycol using two conserved lysine residues. Biochem. J. 2020, 477, 1049–1059. [Google Scholar] [CrossRef]
- Zahn, K.E.; Averill, A.M.; Aller, P.; Wood, R.D.; Doublie, S. Human DNA polymerase theta grasps the primer terminus to mediate DNA repair. Nat. Struct. Mol. Biol. 2015, 22, 304–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogg, M.; Seki, M.; Wood, R.D.; Doublie, S.; Wallace, S.S. Lesion bypass activity of DNA polymerase theta (POLQ) is an intrinsic property of the pol domain and depends on unique sequence inserts. J. Mol. Biol. 2011, 405, 642–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morley, S.A.; Peralta-Castro, A.; Brieba, L.G.; Miller, J.; Ong, K.L.; Ridge, P.G.; Oliphant, A.; Aldous, S.; Nielsen, B.L. Arabidopsis thaliana organelles mimic the T7 phage DNA replisome with specific interactions between Twinkle protein and DNA polymerases Pol1A and Pol1B. BMC Plant Biol. 2019, 19, 241. [Google Scholar] [CrossRef] [PubMed]
- Chevigny, N.; Schatz-Daas, D.; Lotfi, F.; Gualberto, J.M. DNA Repair and the Stability of the Plant Mitochondrial Genome. Int. J. Mol. Sci. 2020, 21, 328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gualberto, J.M.; Newton, K.J. Plant Mitochondrial Genomes: Dynamics and Mechanisms of Mutation. Annu. Rev. Plant Biol. 2017, 68, 225–252. [Google Scholar] [CrossRef] [PubMed]
- Morley, S.A.; Nielsen, B.L. Plant mitochondrial DNA. Front. Biosci. 2017, 22, 1023–1032. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.S.; Kennedy, W.D.; Yin, Y.W. Structural insight into processive human mitochondrial DNA synthesis and disease-related polymerase mutations. Cell 2009, 139, 312–324. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Cui, Y.; Fox, T.; Lin, S.; Wang, H.; de Val, N.; Zhou, Z.H.; Yang, W. Structures and operating principles of the replisome. Science 2019, 363, eaav7003. [Google Scholar] [CrossRef]
- Kulczyk, A.W.; Akabayov, B.; Lee, S.J.; Bostina, M.; Berkowitz, S.A.; Richardson, C.C. An interaction between DNA polymerase and helicase is essential for the high processivity of the bacteriophage T7 replisome. J. Biol. Chem. 2012, 287, 39050–39060. [Google Scholar] [CrossRef] [Green Version]
- Hamdan, S.M.; Richardson, C.C. Motors, switches, and contacts in the replisome. Annu. Rev. Biochem. 2009, 78, 205–243. [Google Scholar] [CrossRef] [Green Version]
- Sinha, N.K.; Morris, C.F.; Alberts, B.M. Efficient in vitro replication of double-stranded DNA templates by a purified T4 bacteriophage replication system. J. Biol. Chem. 1980, 255, 4290–4293. [Google Scholar] [PubMed]
- Zhang, H.; Lee, S.J.; Zhu, B.; Tran, N.Q.; Tabor, S.; Richardson, C.C. Helicase-DNA polymerase interaction is critical to initiate leading-strand DNA synthesis. Proc. Natl. Acad. Sci. USA 2011, 108, 9372–9377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, A.J.; Richardson, C.C. Gp2.5, the multifunctional bacteriophage T7 single-stranded DNA binding protein. Semin. Cell Dev. Biol. 2019, 86, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Medel, P.L.; Baruch-Torres, N.; Peralta-Castro, A.; Trasvina-Arenas, C.H.; Torres-Larios, A.; Brieba, L.G. Plant organellar DNA polymerases repair double-stranded breaks by microhomology-mediated end-joining. Nucleic Acids Res. 2019, 47, 3028–3044. [Google Scholar] [CrossRef]
- Fuchs, P.; Rugen, N.; Carrie, C.; Elsasser, M.; Finkemeier, I.; Giese, J.; Hildebrandt, T.M.; Kuhn, K.; Maurino, V.G.; Ruberti, C.; et al. Single organelle function and organization as estimated from Arabidopsis mitochondrial proteomics. Plant J. 2020, 101, 420–441. [Google Scholar] [CrossRef] [Green Version]
- Diray-Arce, J.; Liu, B.; Cupp, J.D.; Hunt, T.; Nielsen, B.L. The Arabidopsis At1g30680 gene encodes a homologue to the phage T7 gp4 protein that has both DNA primase and DNA helicase activities. BMC Plant Biol. 2013, 13, 36. [Google Scholar] [CrossRef] [Green Version]
- Edmondson, A.C.; Song, D.; Alvarez, L.A.; Wall, M.K.; Almond, D.; McClellan, D.A.; Maxwell, A.; Nielsen, B.L. Characterization of a mitochondrially targeted single-stranded DNA-binding protein in Arabidopsis thaliana. Mol. Genet. Genomics. 2005, 273, 115–122. [Google Scholar] [CrossRef]
- DeBalsi, K.L.; Hoff, K.E.; Copeland, W.C. Role of the mitochondrial DNA replication machinery in mitochondrial DNA mutagenesis, aging and age-related diseases. Ageing Res. Rev. 2017, 33, 89–104. [Google Scholar] [CrossRef] [Green Version]
- Kunkel, T.A. DNA replication fidelity. J. Biol. Chem. 2004, 279, 16895–16898. [Google Scholar] [CrossRef] [Green Version]
- Ayala-Garcia, V.M.; Baruch-Torres, N.; Garcia-Medel, P.L.; Brieba, L.G. Plant organellar DNA polymerases paralogs exhibit dissimilar nucleotide incorporation fidelity. FEBS J. 2018, 285, 4005–4018. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, K.H.; Li, W.H.; Sharp, P.M. Rates of nucleotide substitution vary greatly among plant mitochondrial, chloroplast, and nuclear DNAs. Proc. Natl. Acad. Sci. USA 1987, 84, 9054–9058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, J.D.; Herbon, L.A. Plant mitochondrial DNA evolves rapidly in structure, but slowly in sequence. J. Mol. Evol. 1988, 28, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Brieba, L.G. Structure-Function Analysis Reveals the Singularity of Plant Mitochondrial DNA Replication Components: A Mosaic and Redundant System. Plants 2019, 8, 533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Waneka, G.; Broz, A.K.; King, C.R.; Sloan, D.B. MSH1 is required for maintenance of the low mutation rates in plant mitochondrial and plastid genomes. Proc. Natl. Acad. Sci. USA 2020, 117, 16448–16455. [Google Scholar] [CrossRef] [PubMed]
- Jackman, S.; Birol, I. Largest Complete Mitochondrial Genome of a. Gymnosperm, Sitka Spruce (Picea sitchensis),. Indicates Complex Physical Structure. bioRxiv 2019. preprint. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Cuthbert, J.M.; Taylor, D.R.; Sloan, D.B. The massive mitochondrial genome of the angiosperm Silene noctiflora is evolving by gain or loss of entire chromosomes. Proc. Natl. Acad. Sci. USA 2015, 112, 10185–10191. [Google Scholar] [CrossRef] [Green Version]
- Boore, J.L. Animal mitochondrial genomes. Nucleic Acids Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef] [Green Version]
- Backert, S.; Borner, T. Phage T4-like intermediates of DNA replication and recombination in the mitochondria of the higher plant Chenopodium album (L.). Curr. Genet. 2000, 37, 304–314. [Google Scholar] [CrossRef]
- Bendich, A.J. Structural analysis of mitochondrial DNA molecules from fungi and plants using moving pictures and pulsed-field gel electrophoresis. J. Mol. Biol. 1996, 255, 564–588. [Google Scholar] [CrossRef]
- Blanco, L.; Bernad, A.; Lazaro, J.M.; Martin, G.; Garmendia, C.; Salas, M. Highly efficient DNA synthesis by the phage phi 29 DNA polymerase. Symmetrical mode of DNA replication. J. Biol. Chem. 1989, 264, 8935–8940. [Google Scholar]
- Kelman, Z.; Hurwitz, J.; O’Donnell, M. Processivity of DNA polymerases: Two mechanisms, one goal. Structure 1998, 6, 121–125. [Google Scholar] [CrossRef] [Green Version]
- Huber, H.E.; Tabor, S.; Richardson, C.C. Escherichia coli thioredoxin stabilizes complexes of bacteriophage T7 DNA polymerase and primed templates. J. Biol. Chem. 1987, 262, 16224–16232. [Google Scholar] [PubMed]
- Tabor, S.; Huber, H.E.; Richardson, C.C. Escherichia coli thioredoxin confers processivity on the DNA polymerase activity of the gene 5 protein of bacteriophage T7. J. Biol. Chem. 1987, 262, 16212–16223. [Google Scholar] [PubMed]
- Szymanski, M.R.; Kuznetsov, V.B.; Shumate, C.; Meng, Q.; Lee, Y.S.; Patel, G.; Patel, S.; Yin, Y.W. Structural basis for processivity and antiviral drug toxicity in human mitochondrial DNA replicase. EMBO J. 2015, 34, 1959–1970. [Google Scholar] [CrossRef] [Green Version]
- Wuite, G.J.; Smith, S.B.; Young, M.; Keller, D.; Bustamante, C. Single-molecule studies of the effect of template tension on T7 DNA polymerase activity. Nature 2000, 404, 103–106. [Google Scholar] [CrossRef]
- Studwell, P.S.; O’Donnell, M. Processive replication is contingent on the exonuclease subunit of DNA polymerase III holoenzyme. J. Biol. Chem. 1990, 265, 1171–1178. [Google Scholar]
- Tanner, N.A.; Hamdan, S.M.; Jergic, S.; Loscha, K.V.; Schaeffer, P.M.; Dixon, N.E.; van Oijen, A.M. Single-molecule studies of fork dynamics in Escherichia coli DNA replication. Nat. Struct. Mol. Biol. 2008, 15, 170–176. [Google Scholar] [CrossRef] [Green Version]
- Ignatov, A.V.; Bondarenko, K.A.; Makarova, A.V. Non-bulky Lesions in Human DNA: The Ways of Formation, Repair, and Replication. Acta Nat. 2017, 9, 12–26. [Google Scholar] [CrossRef]
- Klungland, A.; Hoss, M.; Gunz, D.; Constantinou, A.; Clarkson, S.G.; Doetsch, P.W.; Bolton, P.H.; Wood, R.D.; Lindahl, T. Base excision repair of oxidative DNA damage activated by XPG protein. Mol. Cell 1999, 3, 33–42. [Google Scholar] [CrossRef]
- Scharer, O.D. Nucleotide excision repair in eukaryotes. Cold Spring Harb. Perspect. Biol. 2013, 5, a012609. [Google Scholar] [CrossRef]
- Goodman, M.F.; Woodgate, R. Translesion DNA polymerases. Cold Spring Harb. Perspect. Biol. 2013, 5, a010363. [Google Scholar] [CrossRef] [PubMed]
- Yang, W. An overview of Y-Family DNA polymerases and a case study of human DNA polymerase eta. Biochemistry 2014, 53, 2793–2803. [Google Scholar] [CrossRef] [PubMed]
- Seki, M.; Masutani, C.; Yang, L.W.; Schuffert, A.; Iwai, S.; Bahar, I.; Wood, R.D. High-efficiency bypass of DNA damage by human DNA polymerase Q. EMBO J. 2004, 23, 4484–4494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kent, T.; Chandramouly, G.; McDevitt, S.M.; Ozdemir, A.Y.; Pomerantz, R.T. Mechanism of microhomology-mediated end-joining promoted by human DNA polymerase theta. Nat. Struct. Mol. Biol. 2015, 22, 230–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Wu, E.Y.; Hellinga, H.W.; Beese, L.S. Structural factors that determine selectivity of a high fidelity DNA polymerase for deoxy-, dideoxy-, and ribonucleotides. J. Biol. Chem. 2012, 287, 28215–28226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krokan, H.E.; Standal, R.; Slupphaug, G. DNA glycosylases in the base excision repair of DNA. Biochem. J. 1997, 325, 1–16. [Google Scholar] [CrossRef]
- Boesch, P.; Weber-Lotfi, F.; Ibrahim, N.; Tarasenko, V.; Cosset, A.; Paulus, F.; Lightowlers, R.N.; Dietrich, A. DNA repair in organelles: Pathways, organization, regulation, relevance in disease and aging. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2011, 1813, 186–200. [Google Scholar] [CrossRef] [Green Version]
- Gredilla, R. DNA damage and base excision repair in mitochondria and their role in aging. J. Aging Res. 2010, 2011, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Prakash, A.; Doublié, S. Base excision repair in the mitochondria. J. Cell. Biochem. 2015, 116, 1490–1499. [Google Scholar] [CrossRef] [Green Version]
- Daskalova, S.M.; Bai, X.; Hecht, S.M. Study of the Lyase Activity of Human DNA Polymerase β Using Analogues of the Intermediate Schiff Base Complex. Biochemistry 2018, 57, 2711–2722. [Google Scholar] [CrossRef]
- Wong, D.; DeMott, M.S.; Demple, B. Modulation of the 3′→5′-exonuclease activity of human apurinic endonuclease (Ape1) by its 5’-incised Abasic DNA product. J. Biol. Chem. 2003, 278, 36242–36249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beard, W.A.; Horton, J.K.; Prasad, R.; Wilson, S.H. Eukaryotic Base Excision Repair: New Approaches Shine Light on Mechanism. Annu. Rev. Biochem. 2019, 88, 137–162. [Google Scholar] [CrossRef]
- Pedroza-Garcia, J.A.; De Veylder, L.; Raynaud, C. Plant DNA Polymerases. Int. J. Mol. Sci. 2019, 20, 4814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nota, F.; Cambiagno, D.A.; Ribone, P.A.; Alvarez, M.-A.E.; Alvarez, M.E. Expression and function of AtMBD4L, the single gene encoding the nuclear DNA glycosylase MBD4L in Arabidopsis. Plant Sci. 2015, 235, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Rissel, D.; Losch, J.; Peiter, E. The nuclear protein Poly (ADP-ribose) polymerase 3 (At PARP 3) is required for seed storability in Arabidopsis thaliana. Plant Biol. 2014, 16, 1058–1064. [Google Scholar]
- Boesch, P.; Ibrahim, N.; Paulus, F.; Cosset, A.; Tarasenko, V.; Dietrich, A. Plant mitochondria possess a short-patch base excision DNA repair pathway. Nucleic Acids Res. 2009, 37, 5690–5700. [Google Scholar] [CrossRef] [Green Version]
- Gutman, B.L.; Niyogi, K.K. Evidence for base excision repair of oxidative DNA damage in chloroplasts of Arabidopsis thaliana. J. Biol. Chem. 2009, 284, 17006–17012. [Google Scholar] [CrossRef] [Green Version]
- Córdoba-Cañero, D.; Morales-Ruiz, T.; Roldán-Arjona, T.; Ariza, R.R. Single-nucleotide and long-patch base excision repair of DNA damage in plants. Plant J. 2009, 60, 716–728. [Google Scholar] [CrossRef] [Green Version]
- Córdoba-Cañero, D.; Roldán-Arjona, T.; Ariza, R.R. Arabidopsis ARP endonuclease functions in a branched base excision DNA repair pathway completed by LIG1. Plant J. 2011, 68, 693–702. [Google Scholar] [CrossRef]
- Barbado, C.; Córdoba-Cañero, D.; Ariza, R.R.; Roldán-Arjona, T. Nonenzymatic release of N7-methylguanine channels repair of abasic sites into an AP endonuclease-independent pathway in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, E916–E924. [Google Scholar] [CrossRef] [Green Version]
- Ferrando, B.; Furlanetto, A.; Gredilla, R.; Havelund, J.F.; Hebelstrup, K.H.; Moller, I.M.; Stevnsner, T. DNA repair in plant mitochondria—A complete base excision repair pathway in potato tuber mitochondria. Physiol. Plant. 2019, 166, 494–512. [Google Scholar] [CrossRef]
- Sunderland, P.A.; West, C.E.; Waterworth, W.M.; Bray, C.M. An evolutionarily conserved translation initiation mechanism regulates nuclear or mitochondrial targeting of DNA ligase 1 in Arabidopsis thaliana. Plant. J. 2006, 47, 356–367. [Google Scholar] [CrossRef]
- Almagro, A.J.J.; Salvatore, M.; Emanuelsson, O.; Winther, O.; von Heijne, G.; Elofsson, A.; Nielsen, H. Detecting sequence signals in targeting peptides using deep learning. Life Sci. Alliance 2019, 2, e201900429. [Google Scholar] [CrossRef] [Green Version]
- Trasviña-Arenas, C.H.; Baruch-Torres, N.; Cordoba-Andrade, F.J.; Ayala-García, V.M.; García-Medel, P.L.; Díaz-Quezada, C.; Peralta-Castro, A.; Ordaz-Ortiz, J.J.; Brieba, L.G. Identification of a unique insertion in plant organellar DNA polymerases responsible for 5′-dRP lyase and strand-displacement activities: Implications for Base Excision Repair. DNA Repair 2018, 65, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Garcıa-Dıaz, M.; Bebenek, K.; Kunkel, T.A.; Blanco, L. Identification of an Intrinsic 5′-Deoxyribose-5-phosphate Lyase Activity in Human DNA Polymerase λ A Possible role in base excision repair. J. Biol. Chem. 2001, 276, 34659–34663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deterding, L.J.; Prasad, R.; Mullen, G.P.; Wilson, S.H.; Tomer, K.B. Mapping of the 5′-2-deoxyribose-5-phosphate lyase active site in DNA polymerase β by mass spectrometry. J. Biol. Chem. 2000, 275, 10463–10471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariotti, L.G.; Pirovano, G.; Savage, K.I.; Ghita, M.; Ottolenghi, A.; Prise, K.M.; Schettino, G. Use of the γ-H2AX assay to investigate DNA repair dynamics following multiple radiation exposures. PLoS ONE 2013, 8, e79541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varas, J.; Sanchez-Moran, E.; Copenhaver, G.P.; Santos, J.L.; Pradillo, M. Analysis of the Relationships between DNA Double-Strand Breaks, Synaptonemal Complex and Crossovers Using the Atfas1–4 Mutant. PLoS Genet. 2015, 11, e1005301. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, D.; Nam, A.; Oldenburg, D.J.; Bendich, A.J. Reactive Oxygen Species, Antioxidant Agents, and DNA Damage in Developing Maize Mitochondria and Plastids. Front. Plant Sci. 2020, 11, 596. [Google Scholar] [CrossRef]
- Anand, R.P.; Lovett, S.T.; Haber, J.E. Break-induced DNA replication. Cold Spring Harb. Perspect. Biol. 2013, 5, a010397. [Google Scholar] [CrossRef] [Green Version]
- Miller-Messmer, M.; Kuhn, K.; Bichara, M.; Le Ret, M.; Imbault, P.; Gualberto, J.M. RecA-dependent DNA repair results in increased heteroplasmy of the Arabidopsis mitochondrial genome. Plant Physiol. 2012, 159, 211–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tremblay-Belzile, S.; Lepage, E.; Zampini, E.; Brisson, N. Short-range inversions: Rethinking organelle genome stability: Template switching events during DNA replication destabilize organelle genomes. Bioessays 2015, 37, 1086–1094. [Google Scholar] [CrossRef] [PubMed]
- Davila, J.I.; Arrieta-Montiel, M.P.; Wamboldt, Y.; Cao, J.; Hagmann, J.; Shedge, V.; Xu, Y.Z.; Weigel, D.; Mackenzie, S.A. Double-strand break repair processes drive evolution of the mitochondrial genome in Arabidopsis. BMC Biol. 2011, 9, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marechal, A.; Parent, J.S.; Veronneau-Lafortune, F.; Joyeux, A.; Lang, B.F.; Brisson, N. Whirly proteins maintain plastid genome stability in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 14693–14698. [Google Scholar] [CrossRef] [Green Version]
- Kwon, T.; Huq, E.; Herrin, D.L. Microhomology-mediated and nonhomologous repair of a double-strand break in the chloroplast genome of Arabidopsis. Proc. Natl. Acad. Sci. USA 2010, 107, 13954–13959. [Google Scholar] [CrossRef] [Green Version]
- Hanscom, T.; McVey, M. Regulation of Error-Prone DNA Double-Strand Break Repair and Its Impact on Genome Evolution. Cells 2020, 9, 1657. [Google Scholar] [CrossRef]
- Seol, J.H.; Shim, E.Y.; Lee, S.E. Microhomology-mediated end joining: Good, bad and ugly. Mutation Res. 2018, 809, 81–87. [Google Scholar] [CrossRef]
- Tadi, S.K.; Sebastian, R.; Dahal, S.; Babu, R.K.; Choudhary, B.; Raghavan, S.C. Microhomology-mediated end joining is the principal mediator of double-strand break repair during mitochondrial DNA lesions. Mol. Biol. Cell 2016, 27, 223–235. [Google Scholar] [CrossRef]
- Crespan, E.; Czabany, T.; Maga, G.; Hubscher, U. Microhomology-mediated DNA strand annealing and elongation by human DNA polymerases lambda and beta on normal and repetitive DNA sequences. Nucleic Acids Res. 2012, 40, 5577–5590. [Google Scholar] [CrossRef] [Green Version]
- Ceccaldi, R.; Liu, J.C.; Amunugama, R.; Hajdu, I.; Primack, B.; Petalcorin, M.I.; O’Connor, K.W.; Konstantinopoulos, P.A.; Elledge, S.J.; Boulton, S.J.; et al. Homologous-recombination-deficient tumours are dependent on Poltheta-mediated repair. Nature 2015, 518, 258–262. [Google Scholar] [CrossRef] [Green Version]
- Yousefzadeh, M.J.; Wyatt, D.W.; Takata, K.; Mu, Y.; Hensley, S.C.; Tomida, J.; Bylund, G.O.; Doublie, S.; Johansson, E.; Ramsden, D.A.; et al. Mechanism of suppression of chromosomal instability by DNA polymerase POLQ. PLoS Genet 2014, 10, e1004654. [Google Scholar] [CrossRef] [Green Version]
- Mateos-Gomez, P.A.; Gong, F.; Nair, N.; Miller, K.M.; Lazzerini-Denchi, E.; Sfeir, A. Mammalian polymerase theta promotes alternative NHEJ and suppresses recombination. Nature 2015, 518, 254–257. [Google Scholar] [CrossRef] [Green Version]
- Brambati, A.; Barry, R.M.; Sfeir, A. DNA polymerase theta (Poltheta)—An error-prone polymerase necessary for genome stability. Curr. Opin. Genet. Dev. 2020, 60, 119–126. [Google Scholar] [CrossRef]
- Wood, R.D.; Doublie, S. DNA polymerase theta (POLQ), double-strand break repair, and cancer. DNA Repair 2016, 44, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Black, S.J.; Ozdemir, A.Y.; Kashkina, E.; Kent, T.; Rusanov, T.; Ristic, D.; Shin, Y.; Suma, A.; Hoang, T.; Chandramouly, G.; et al. Molecular basis of microhomology-mediated end-joining by purified full-length Poltheta. Nat. Commun. 2019, 10, 4423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, J.A.; Cooper, C.D.O.; Aitkenhead, H.; Gileadi, O. Structure of the Helicase Domain of DNA Polymerase Theta Reveals a Possible Role in the Microhomology-Mediated End-Joining Pathway. Structure 2015, 23, 2319–2330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beagan, K.; Armstrong, R.L.; Witsell, A.; Roy, U.; Renedo, N.; Baker, A.E.; Scharer, O.D.; McVey, M. Drosophila DNA polymerase theta utilizes both helicase-like and polymerase domains during microhomology-mediated end joining and interstrand crosslink repair. PLoS Genet. 2017, 13, e1006813. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Yang, W. Template and primer requirements for DNA Pol theta-mediated end joining. Proc. Natl. Acad. Sci. USA 2018, 115, 7747–7752. [Google Scholar] [CrossRef] [Green Version]
- Sfeir, A.; Symington, L.S. Microhomology-Mediated End Joining: A Back-up Survival Mechanism or Dedicated Pathway? Trends Biochem. Sci. 2015, 40, 701–714. [Google Scholar] [CrossRef] [Green Version]
- Cappadocia, L.; Marechal, A.; Parent, J.S.; Lepage, E.; Sygusch, J.; Brisson, N. Crystal structures of DNA-Whirly complexes and their role in Arabidopsis organelle genome repair. Plant Cell 2010, 22, 1849–1867. [Google Scholar] [CrossRef] [Green Version]
- Zaegel, V.; Guermann, B.; Le Ret, M.; Andres, C.; Meyer, D.; Erhardt, M.; Canaday, J.; Gualberto, J.M.; Imbault, P. The plant-specific ssDNA binding protein OSB1 is involved in the stoichiometric transmission of mitochondrial DNA in Arabidopsis. Plant Cell 2006, 18, 3548–3563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, J.; Day, A. Construction of a highly error-prone DNA polymerase for developing organelle mutation systems. Nucleic Acids Res. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Sousa, R. Structural and mechanistic relationships between nucleic acid polymerases. Trends Biochem. Sci. 1996, 21, 186–190. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Gene | Activity | TAIR ID | Localization + | References for Cellular Localization |
|---|---|---|---|---|---|
| Family A DNA polymerase | AtPOLIA | DNA polymerase | AT1G50840 | M, C | [13,35] |
| Family A DNA polymerase | AtPOLIB | DNA polymerase | AT3G20540 | M, C | [13,35] |
| DNA primase-helicase | AtTWINKLE | DNA primase DNA helicase | AT1G30680 | M, C | [35,36] |
| SSB1 | AtSSB1 | Single-stranded binding protein | AT4G11060 | M, C | [35] |
| SSB2 | AtSSB2 | Single-stranded binding protein | AT3G18580 | M | [35,37] |
| Enzyme | Gene | TAIR ID | Mitochondrial (Predicted TargetP [83]) | Chloroplast (Predicted TargetP [83]) | Reference for | Substrate |
|---|---|---|---|---|---|---|
| Uracil DNA glycosylase | AtUNG | AT3G18630 | + (Experimental) | + (Predicted) | [76] | U, 5-FU |
| Endonuclease III 1 | AtNTH1 | AT2G31450 | + (Predicted) | + (Experimental) | [77] | Tg, 5-hC, 5-hU, Fapy lesions |
| Endonuclease III 2 | AtNTH2 | AT1G05900 | + (Predicted) | + (Experimental) | [77] | Tg, 5-hC, 5-hU, Fapy lesions |
| Formamidopyrimidine DNA Glycosylase | AtFPG | AT1G52500 | + (Predicted) | - (Predicted) | n.d | Fapy lesions and 8oxoG |
| 5-meC and thymine-DNA glycosylase | DML-3 | AT4G34060 | - (Predicted) | - (Predicted) | n.d | T:G, U:G, 5-MeC, halogenated pyrimidines, 5-FU, Tg:G |
| Adenine DNA Glycosylase | MutY | AT4G12740 | + (Predicted) | + (Predicted) | n.d | A across 8oxoG |
| Polynucleotide 5′-Kinase/3′-Phosphatase | AtZDP | AT3G14890 | + (Predicted) | + (Predicted) | n.d | Dephosphorilates 3′-P ends producing 3′-OH |
| Xth Endonuclease | ARP | AT2G41460 | + (Predicted) | + (Experimental) | [77] | AP sites, DHU, α-dA |
| Family A DNA polymerase | AtPOLIA | AT1G50840 | + (Experimental) | + (Experimental) | [13] | Lyase and strand-displacement |
| Family A DNA polymerase | AtPOLIB | AT3G20540 | + (Experimental) | + (Experimental) | [13] | Lyase and strand-displacement |
| FEN1 | OEX1 | AT3G52050 | + (Predicted) | - (Predicted) | n.d | 5′-Flap structure |
| FEN1 | OEX2 | AT1G34380 | - (Predicted) | + (Predicted) | n.d | 5′-Flap structure |
| ATP-dependent DNA Ligase | AtLig1 | AT1G08130 | + (Experimental) | - (Experimental) | [82] | Nick-containing DNA |
| ATP-dependent DNA Ligase | AtLig6 | AT1G49250 | - (Predicted) | + (Predicted) | n.d | Nick-containing DNA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peralta-Castro, A.; García-Medel, P.L.; Baruch-Torres, N.; Trasviña-Arenas, C.H.; Juarez-Quintero, V.; Morales-Vazquez, C.M.; Brieba, L.G. Plant Organellar DNA Polymerases Evolved Multifunctionality through the Acquisition of Novel Amino Acid Insertions. Genes 2020, 11, 1370. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11111370
Peralta-Castro A, García-Medel PL, Baruch-Torres N, Trasviña-Arenas CH, Juarez-Quintero V, Morales-Vazquez CM, Brieba LG. Plant Organellar DNA Polymerases Evolved Multifunctionality through the Acquisition of Novel Amino Acid Insertions. Genes. 2020; 11(11):1370. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11111370
Chicago/Turabian StylePeralta-Castro, Antolín, Paola L. García-Medel, Noe Baruch-Torres, Carlos H. Trasviña-Arenas, Víctor Juarez-Quintero, Carlos M. Morales-Vazquez, and Luis G. Brieba. 2020. "Plant Organellar DNA Polymerases Evolved Multifunctionality through the Acquisition of Novel Amino Acid Insertions" Genes 11, no. 11: 1370. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11111370