PPARGC1A rs8192678 and NRF1 rs6949152 Polymorphisms Are Associated with Muscle Fiber Composition in Women

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Subjects and Methods
2.1. Subjects
2.2. Genotyping
2.3. Muscle Biopsy
2.4. Statistical Analysis
3. Results
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Brooke, M.H.; Kaiser, K.K. Muscle fiber types: How many and what kind? Arch. Neurol. 1970, 23, 369–379. [Google Scholar] [CrossRef]
- Kumagai, H.; Tobina, T.; Ichinoseki-Sekine, N.; Kakigi, R.; Tsuzuki, T.; Zempo, H.; Shiose, K.; Yoshimura, E.; Kumahara, H.; Ayabe, M.; et al. Role of selected polymorphisms in determining muscle fiber composition in Japanese men and women. J. Appl. Physiol. 2018, 124, 1377–1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, J.M.; Loenneke, J.P.; Jo, E.; Wilson, G.J.; Zourdos, M.C.; Kim, J.-S. The effects of endurance, strength, and power training on muscle fiber type shifting. J. Strength Cond. Res. 2012, 26, 1724–1729. [Google Scholar] [CrossRef] [PubMed]
- Kriketos, A.D.; Pan, D.A.; Lillioja, S.; Cooney, G.J.; Baur, L.A.; Milner, M.R.; Sutton, J.R.; Jenkins, A.B.; Bogardus, C.; Storlien, L.H. Interrelationships between muscle morphology, insulin action, and adiposity. Am. J. Physiol. 1996, 270 Pt 2, R1332–R1339. [Google Scholar] [CrossRef]
- Toft, I.; Bønaa, K.H.; Lindal, S.; Jenssen, T. Insulin kinetics, insulin action, and muscle morphology in lean or slightly overweight persons with impaired glucose tolerance. Metabolism 1998, 47, 848–854. [Google Scholar] [CrossRef]
- Essén-Gustavsson, B.; Henriksson, J. Enzyme levels in pools of microdissected human muscle fibres of identified type: Adaptive response to exercise. Acta Physiol. Scand. 1984, 120, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Bergh, U.; Thorstensson, A.; Sjödin, B.; Hulten, B.; Piehl, K.; Karlsson, J. Maximal oxygen uptake and muscle fiber types in trained and untrained humans. Med. Sci. Sports 1978, 10, 151–154. [Google Scholar]
- Foster, C.; Costill, D.L.; Daniels, J.T.; Fink, W.J. Skeletal muscle enzyme activity, fiber composition and VO2 max in relation to distance running performance. Eur. J. Appl. Physiol. Occup. Physiol. 1978, 39, 73–80. [Google Scholar] [CrossRef]
- Fry, A.C.; Schilling, B.K.; Staron, R.S.; Hagerman, F.C.; Hikida, R.S.; Thrush, J.T. Muscle fiber characteristics and performance correlates of male Olympic-style weightlifters. J. Strength Cond. Res. 2003, 17, 746–754. [Google Scholar]
- Widrick, J.J.; Stelzer, J.E.; Shoepe, T.C.; Garner, D.P. Functional properties of human muscle fibers after short-term resistance exercise training. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 283, R408–R416. [Google Scholar] [CrossRef] [Green Version]
- Andersen, J.L.; Klitgaard, H.; Saltin, B. Myosin heavy chain isoforms in single fibres from m. vastus lateralis of sprinters: Influence of training. Acta Physiol. Scand. 1994, 151, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Wade, A.J.; Marbut, M.M.; Round, J.M. Muscle fibre type and aetiology of obesity. Lancet 1990, 335, 805–808. [Google Scholar] [CrossRef]
- Gerrits, M.F.; Ghosh, S.; Kavaslar, N.; Hill, B.; Tour, A.; Seifert, E.L.; Beauchamp, B.; Gorman, S.; Stuart, J.; Dent, R.; et al. Distinct skeletal muscle fiber characteristics and gene expression in diet-sensitive versus diet-resistant obesity. J. Lipid Res. 2010, 51, 2394–2404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanner, C.J.; Barakat, H.A.; Dohm, G.L.; Pories, W.J.; MacDonald, K.G.; Cunningham, P.R.; Swanson, M.S.; Houmard, J.A. Muscle fiber type is associated with obesity and weight loss. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E1191–E1196. [Google Scholar] [CrossRef] [Green Version]
- Frisk-Holmberg, M.; Essen, B.; Fredrikson, M.; Strom, G.; Wibell, L. Muscle fibre composition in relation to blood pressure response to isometric exercise in normotensive and hypertensive subjects. Acta Med. Scand. 1983, 213, 21–26. [Google Scholar] [CrossRef]
- Simoneau, J.A.; Bouchard, C. Human variation in skeletal muscle fiber-type proportion and enzyme activities. Am. J. Physiol. 1989, 257 Pt 1, E567–E572. [Google Scholar] [CrossRef]
- Komi, P.V.; Viitasalo, J.H.; Havu, M.; Thorstensson, A.; Sjodin, B.; Karlsson, J. Skeletal muscle fibres and muscle enzyme activities in monozygous and dizygous twins of both sexes. Acta Physiol. Scand. 1977, 100, 385–392. [Google Scholar]
- Simoneau, J.A.; Bouchard, C. Genetic determinism of fiber type proportion in human skeletal muscle. FASEB J. Publ. Fed. Am. Soc. Exp. Biol. 1995, 9, 1091–1095. [Google Scholar] [CrossRef]
- Ahmetov, I.I.; Vinogradova, O.L.; Williams, A.G. Gene polymorphisms and fiber-type composition of human skeletal muscle. Int. J. Sport Nutr. Exerc. Metab. 2012, 22, 292–303. [Google Scholar] [CrossRef]
- Franks, P.W.; Christophi, C.A.; Jablonski, K.A.; Billings, L.K.; Delahanty, L.M.; Horton, E.S.; Knowler, W.C.; Florez, J.C.; Diabetes Prevention Program Research Group. Common variation at PPARGC1A/B and change in body composition and metabolic traits following preventive interventions: The Diabetes Prevention Program. Diabetologia 2014, 57, 485–490. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Puigserver, P.; Andersson, U.; Zhang, C.; Adelmant, G.; Mootha, V.; Troy, A.; Cinti, S.; Lowell, B.; Scarpulla, R.C.; et al. Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell 1999, 98, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Virbasius, J.V.; Scarpulla, R.C. Activation of the human mitochondrial transcription factor A gene by nuclear respiratory factors: A potential regulatory link between nuclear and mitochondrial gene expression in organelle biogenesis. Proc. Natl. Acad. Sci. USA 1994, 91, 1309–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virbasius, J.V.; Virbasius, C.A.; Scarpulla, R.C. Identity of GABP with NRF-2, a multisubunit activator of cytochrome oxidase expression, reveals a cellular role for an ETS domain activator of viral promoters. Genes Dev. 1993, 7, 380–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmetov, I.I.; Williams, A.G.; Popov, D.V.; Lyubaeva, E.V.; Hakimullina, A.M.; Fedotovskaya, O.N.; Mozhayskaya, I.A.; Vinogradova, O.L.; Astratenkova, I.V.; Montgomery, H.E.; et al. The combined impact of metabolic gene polymorphisms on elite endurance athlete status and related phenotypes. Hum. Genet. 2009, 126, 751–761. [Google Scholar] [CrossRef]
- Lin, J.; Wu, H.; Tarr, P.T.; Zhang, C.Y.; Wu, Z.; Boss, O.; Michael, L.F.; Puigserver, P.; Isotani, E.; Olson, E.N.; et al. Transcriptional co-activator PGC-1 alpha drives the formation of slow-twitch muscle fibres. Nature 2002, 418, 797–801. [Google Scholar] [CrossRef]
- Ramachandran, B.; Yu, G.; Gulick, T. Nuclear respiratory factor 1 controls myocyte enhancer factor 2A transcription to provide a mechanism for coordinate expression of respiratory chain subunits. J. Biol. Chem. 2008, 283, 11935–11946. [Google Scholar] [CrossRef] [Green Version]
- Joseph, J.S.; Ayeleso, A.O.; Mukwevho, E. Exercise increases hyper-acetylation of histones on the Cis-element of NRF-1 binding to the Mef2a promoter: Implications on type 2 diabetes. Biochem. Biophys. Res. Commun. 2017, 486, 83–87. [Google Scholar] [CrossRef]
- Handschin, C.; Chin, S.; Li, P.; Liu, F.; Maratos-Flier, E.; LeBrasseur, N.K.; Yan, Z.; Spiegelman, B.M. Skeletal muscle fiber-type switching, exercise intolerance, and myopathy in PGC-1 alpha muscle-specific knock-out animals. J. Biol. Chem. 2007, 282, 30014–30021. [Google Scholar] [CrossRef] [Green Version]
- Eynon, N.; Meckel, Y.; Sagiv, M.; Yamin, C.; Amir, R.; Sagiv, M.; Goldhammer, E.; Duarte, J.A.; Oliveira, J. Do PPARGC1A and PPARα polymorphisms influence sprint or endurance phenotypes? Scand. J. Med. Sci. Sport 2010, 20, e145–e150. [Google Scholar] [CrossRef]
- Lucia, A.; Gómez-Gallego, F.; Barroso, I.; Rabadán, M.; Bandrés, F.; San Juan, A.F.; Chicharro, J.L.; Ekelund, U.; Brage, S.; Earnest, C.P.; et al. PPARGC1A genotype (Gly482Ser) predicts exceptional endurance capacity in European men. J. Appl. Physiol. 2005, 99, 344–348. [Google Scholar] [CrossRef] [Green Version]
- Maciejewska, A.; Sawczuk, M.; Cieszczyk, P.; Mozhayskaya, I.A.; Ahmetov, I.I. The PPARGC1A gene Gly482Ser in Polish and Russian athletes. J. Sports Sci. 2012, 30, 101–113. [Google Scholar] [CrossRef]
- Nishida, Y.; Iyadomi, M.; Higaki, Y.; Tanaka, H.; Kondo, Y.; Otsubo, H.; Horita, M.; Hara, M.; Tanaka, K. Association between the PPARGC1A polymorphism and aerobic capacity in Japanese middle-aged men. Intern. Med. 2015, 54, 359–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrulionyte, L.; Peltola, P.; Chiasson, J.-L.; Laakso, M. Single nucleotide polymorphisms of PPARD in combination with the Gly482Ser substitution of PGC-1A and the Pro12Ala substitution of PPARG2 predict the conversion from impaired glucose tolerance to type 2 diabetes: The STOP-NIDDM trial. Diabetes 2006, 55, 2148–2152. [Google Scholar] [CrossRef] [Green Version]
- Hara, K.; Tobe, K.; Okada, T.; Kadowaki, H.; Akanuma, Y.; Ito, C.; Kimura, S.; Kadowaki, T. A genetic variation in the PGC-1 gene could confer insulin resistance and susceptibility to Type II diabetes. Diabetologia 2002, 45, 740–743. [Google Scholar] [CrossRef] [PubMed]
- Vohl, M.-C.; Houde, A.; Lebel, S.; Hould, F.-S.; Marceau, P. Effects of the peroxisome proliferator-activated receptor-gamma co-activator-1 Gly482Ser variant on features of the metabolic syndrome. Mol. Genet. Metab. 2005, 86, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Steinbacher, P.; Feichtinger, R.G.; Kedenko, L.; Kedenko, I.; Reinhardt, S.; Schönauer, A.L.; Leitner, I.; Sänger, A.M.; Stoiber, W.; Kofler, B.; et al. The single nucleotide polymorphism Gly482Ser in the PGC-1alpha gene impairs exercise-induced slow-twitch muscle fibre transformation in humans. PLoS ONE 2015, 10, e0123881. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Hu, Y.; Feng, L.; Bao, D.; Wang, L.; Li, Y.; Wang, J.; Liu, G.; Xi, Y.; Wen, L.; et al. Is there an association between PPARGC1A genotypes and endurance capacity in Chinese men? Scand. J. Med. Sci. Sports 2008, 18, 195–204. [Google Scholar] [CrossRef]
- Rico-Sanz, J.; Rankinen, T.; Rice, T.; Leon, A.S.; Skinner, J.S.; Wilmore, J.H.; Rao, D.C.; Bouchard, C. Quantitative trait loci for maximal exercise capacity phenotypes and their responses to training in the HERITAGE Family Study. Physiol. Genomics 2004, 16, 256–260. [Google Scholar] [CrossRef]
- Natsume, T.; Ozaki, H.; Kakigi, R.; Kobayashi, H.; Naito, H. Effects of training intensity in electromyostimulation on human skeletal muscle. Eur. J. Appl. Physiol. 2018, 118, 1339–1347. [Google Scholar] [CrossRef]
- He, Z.; Hu, Y.; Feng, L.; Li, Y.; Liu, G.; Xi, Y.; Wen, L.; Lucia, A. NRF-1 genotypes and endurance exercise capacity in young Chinese men. Br. J. Sports Med. 2008, 42, 361–366. [Google Scholar] [CrossRef]
- Yvert, T.; Miyamoto-Mikami, E.; Murakami, H.; Miyachi, M.; Kawahara, T.; Fuku, N. Lack of replication of associations between multiple genetic polymorphisms and endurance athlete status in Japanese population. Physiol. Rep. 2016, 4, e13003. [Google Scholar] [CrossRef] [PubMed]
- Bamman, M.M.; Hill, V.J.; Adams, G.R.; Haddad, F.; Wetzstein, C.J.; Gower, B.A.; Ahmed, A.; Hunter, G.R. Gender differences in resistance-training-induced myofiber hypertrophy among older adults. J. Gerontol. A Biol. Sci. Med. Sci. 2003, 58, 108–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glenmark, B.; Hedberg, G.; Jansson, E. Changes in muscle fibre type from adolescence to adulthood in women and men. Acta Physiol. Scand. 1992, 146, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Velarde, M.C. Mitochondrial and sex steroid hormone crosstalk during aging. Longev. Health 2014, 3, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.-Q.; Cammarata, P.R.; Baines, C.P.; Yager, J.D. Regulation of mitochondrial respiratory chain biogenesis by estrogens/estrogen receptors and physiological, pathological and pharmacological implications. Biochim. Biophys. Acta. 2009, 1793, 1540–1570. [Google Scholar] [CrossRef] [Green Version]
- Klinge, C.M. Estrogenic control of mitochondrial function and biogenesis. J. Cell. Biochem. 2008, 105, 1342–1351. [Google Scholar] [CrossRef] [Green Version]
- Colom, B.; Alcolea, M.P.; Valle, A.; Oliver, J.; Roca, P.; Garcia-Palmer, F.J. Skeletal muscle of female rats exhibit higher mitochondrial mass and oxidative-phosphorylative capacities compared to males. Cell. Physiol. Biochem. 2007, 19, 205–212. [Google Scholar] [CrossRef]
- Witt, H.; Schubert, C.; Jaekel, J.; Fliegner, D.; Penkalla, A.; Tiemann, K.; Stypmann, J.; Roepcke, S.; Brokat, S.; Mahmoodzadeh, S.; et al. Sex-specific pathways in early cardiac response to pressure overload in mice. J. Mol. Med. 2008, 86, 1013–1024. [Google Scholar] [CrossRef] [Green Version]
- Tcherepanova, I.; Puigserver, P.; Norris, J.D.; Spiegelman, B.M.; McDonnell, D.P. Modulation of estrogen receptor-alpha transcriptional activity by the coactivator PGC-1. J. Biol. Chem. 2000, 275, 16302–16308. [Google Scholar] [CrossRef] [Green Version]
- Mattingly, K.A.; Ivanova, M.M.; Riggs, K.A.; Wickramasinghe, N.S.; Barch, M.J.; Klinge, C.M. Estradiol stimulates transcription of nuclear respiratory factor-1 and increases mitochondrial biogenesis. Mol. Endocrinol. 2008, 22, 609–622. [Google Scholar] [CrossRef] [Green Version]
- Wright, D.C.; Han, D.-H.; Garcia-Roves, P.M.; Geiger, P.C.; Jones, T.E.; Holloszy, J.O. Exercise-induced mitochondrial biogenesis begins before the increase in muscle PGC-1alpha expression. J. Biol. Chem. 2007, 282, 194–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gleyzer, N.; Vercauteren, K.; Scarpulla, R.C. Control of mitochondrial transcription specificity factors (TFB1M and TFB2M) by nuclear respiratory factors (NRF-1 and NRF-2) and PGC-1 family coactivators. Mol. Cell. Biol. 2005, 25, 1354–1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wende, A.R.; Schaeffer, P.J.; Parker, G.J.; Zechner, C.; Han, D.H.; Chen, M.M.; Hancock, C.R.; Lehman, J.J.; Huss, J.M.; McClain, D.A.; et al. A role for the transcriptional coactivator PGC-1alpha in muscle refueling. J. Biol. Chem. 2007, 282, 36642–36651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvo, J.A.; Daniels, T.G.; Wang, X.; Paul ALin, J.; Spiegelman, B.M.; Stevenson, S.C.; Rangwala, S.M. Muscle-specific expression of PPARgamma coactivator-1alpha improves exercise performance and increases peak oxygen uptake. J. Appl. Physiol. 2008, 104, 1304–1312. [Google Scholar] [CrossRef] [PubMed]
- Michael, L.F.; Wu, Z.; Cheatham, R.B.; Puigserver, P.; Adelmant, G.; Lehman, J.J.; Kelly, D.P.; Spiegelman, B.M. Restoration of insulin-sensitive glucose transporter (GLUT4) gene expression in muscle cells by the transcriptional coactivator PGC-1. Proc. Natl. Acad. Sci. USA 2001, 98, 3820–3825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
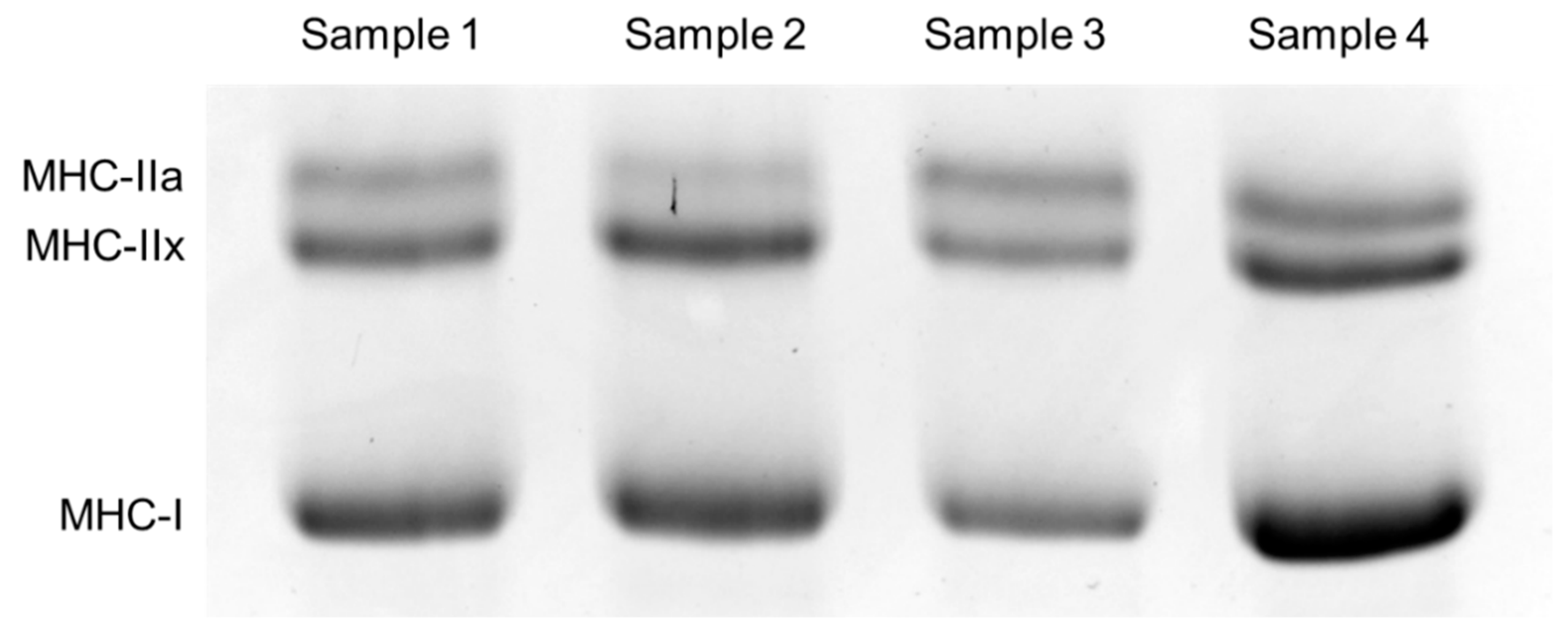
- Pandorf, C.E.; Caiozzo, V.J.; Haddad, F.; Baldwin, K.M. A rationale for SDS-PAGE of MHC isoforms as a gold standard for determining contractile phenotype. J. Appl. Physiol. 2010, 108, 222. [Google Scholar]
- Serrano, N.; Colenso-Semple, L.M.; Lazauskus, K.K.; Siu, J.W.; Bagley, J.R.; Lockie, R.G.; Costa, P.B.; Galpin, A.J. Extraordinary fast-twitch fiber abundance in elite weightlifters. PLoS ONE 2019, 14, e0207975. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Men (n = 107) | Women (n = 107) | |
|---|---|---|
| Age (years) | 47.5 ± 17.9 | 48.3 ± 16.3 |
| Height (cm) | 169.7 ± 5.9 | 156.7 ± 5.9 *** |
| Weight (kg) | 74.7 ± 11.2 | 62.5 ± 9.9 *** |
| BMI (kg/m2) | 25.9 ± 3.8 | 25.5 ± 4.3 |
| MHC-I (%) | 40.7 ± 11.5 | 50.3 ± 11.2 *** |
| MHC-IIa (%) | 35.9 ± 8.2 | 30.9 ± 8.2 *** |
| MHC-IIx (%) | 23.4 ± 9.1 | 18.8 ± 8.3 *** |
| Gene Name (rs Number) | Genotype | p (AIC) | ||||
|---|---|---|---|---|---|---|
| PPARGC1A (rs8192678) | Additive | Dominant | Recessive | |||
| Men | GG (n = 31) | GA (n = 51) | AA (n = 22) | GG vs. GA vs. AA | GG + GA vs. AA | GG vs. GA + AA |
| MHC-I (%) | 40.3 ± 10.7 | 41.6 ± 10.8 | 39.3 ± 14.6 | 0.854 (795.8) | 0.985 (795.9) | 0.764 (795.8) |
| MHC-IIa (%) | 37.5 ± 7.3 | 34.4 ± 7.2 | 36.9 ± 11.0 | 0.366 (724.2) | 0.965 (725.1) | 0.150 (722.9) |
| MHC-IIx (%) | 22.2 ± 8.9 | 24.0 ± 9.0 | 23.8 ± 10.0 | 0.589 (759.0) | 0.981 (759.3) | 0.392 (758.6) |
| Women | (n = 36) | (n = 52) | (n = 18) | |||
| MHC-I (%) | 47.7 ± 12.1 | 50.3 ± 11.0 | 54.4 ± 7.7 | 0.042 (809.1) * | 0.084 (810.3) | 0.112 (810.7) |
| MHC-IIa (%) | 32.3 ± 8.3 | 30.1 ± 8.0 | 30.4 ± 9.0 | 0.368 (750.3) | 0.854 (751.1) | 0.241 (749.7) |
| MHC-IIx (%) | 20.1 ± 8.9 | 19.6 ± 7.4 | 15.2 ± 8.0 | 0.072 (749.2) | 0.033 (747.8) * | 0.348 (751.7) |
| NRF1 (rs6949152) | Additive | Dominant | Recessive | |||
| Men | AA (n = 77) | AG (n = 21) | GG (n = 6) | AA vs. AG vs. GG | AA + AG vs. GG | AA vs. AG + GG |
| MHC-I (%) | 39.8 ± 11.4 | 44.3 ± 11.6 | 40.2 ± 13.3 | 0.334 (794.9) | 0.713 (795.7) | 0.142 (793.6) |
| MHC-IIa (%) | 36.0 ± 8.5 | 34.8 ± 7.1 | 38.0 ± 9.2 | 0.853 (725.0) | 0.341 (724.1) | 0.794 (725.0) |
| MHC-IIx (%) | 24.2 ± 9.1 | 20.9 ± 9.3 | 21.7 ± 8.3 | 0.188 (757.6) | 0.713 (759.2) | 0.125 (756.9) |
| Women | (n = 59) | (n = 41) | (n = 6) | |||
| MHC-I (%) | 52.7 ± 10.5 | 46.3 ± 11.5 | 51.0 ± 6.6 | 0.033 (808.7) * | 0.891 (813.3) | 0.008 (806.0) ** |
| MHC-IIa (%) | 29.8 ± 8.7 | 32.4 ± 7.4 | 31.2 ± 8.7 | 0.221 (749.6) | 0.886 (751.1) | 0.155 (749.0) |
| MHC-IIx (%) | 17.5 ± 7.7 | 21.3 ± 8.7 | 17.8 ± 5.1 | 0.108 (749.9) | 0.745 (752.5) | 0.035 (748.0) * |
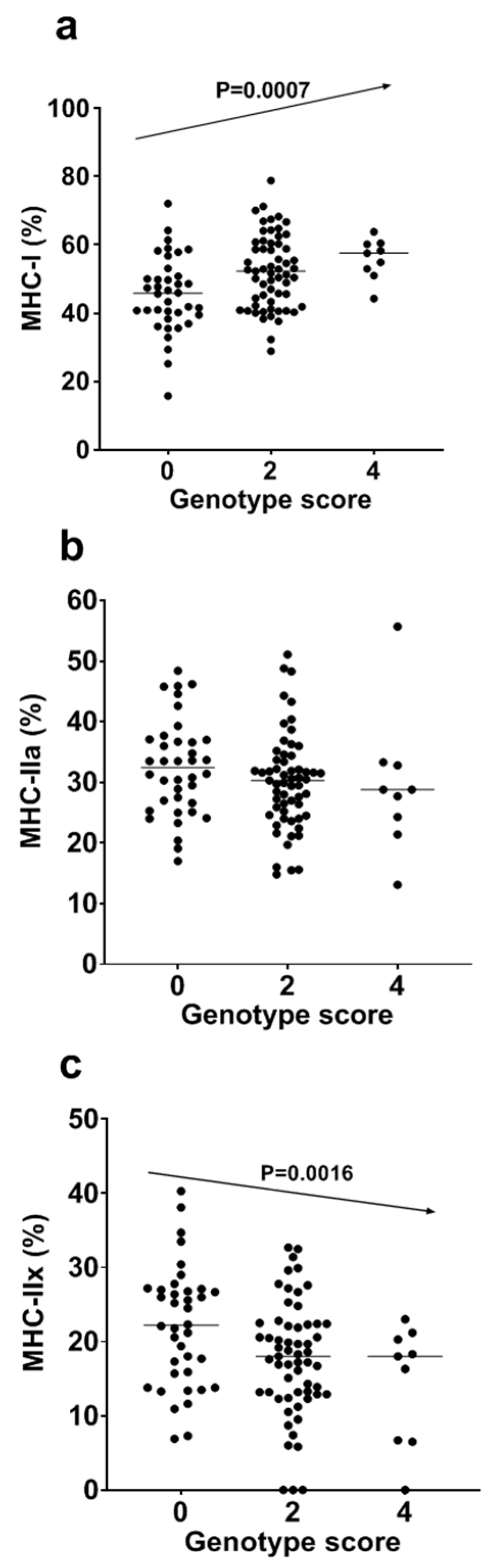
| − | Model | R2 (Adjusted R2) | β for GS (95% CI), p Value |
|---|---|---|---|
| MHC-I | 1 | 0.104 (0.096) | 2.94 (1.26–4.61), p = 0.0007 |
| 2 | 0.140 (0.123) | 2.82 * (1.17–4.47), p = 0.0010 | |
| MHC-IIa | 1 | 0.018 (0.009) | −0.92 (−2.22−0.38), p = 0.1657 |
| 2 | 0.053 (0.035) | −0.83 * (−0.19−0.00), p = 0.2014 | |
| MHC-IIx | 1 | 0.091 (0.083) | −2.02 (−3.26–(−0.78)), p = 0.0016 |
| 2 | 0.096 (0.078) | −1.99 * (−3.24–(−0.75)), p = 0.0020 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yvert, T.; Miyamoto-Mikami, E.; Tobina, T.; Shiose, K.; Kakigi, R.; Tsuzuki, T.; Takaragawa, M.; Ichinoseki-Sekine, N.; Pérez, M.; Kobayashi, H.; et al. PPARGC1A rs8192678 and NRF1 rs6949152 Polymorphisms Are Associated with Muscle Fiber Composition in Women. Genes 2020, 11, 1012. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11091012
Yvert T, Miyamoto-Mikami E, Tobina T, Shiose K, Kakigi R, Tsuzuki T, Takaragawa M, Ichinoseki-Sekine N, Pérez M, Kobayashi H, et al. PPARGC1A rs8192678 and NRF1 rs6949152 Polymorphisms Are Associated with Muscle Fiber Composition in Women. Genes. 2020; 11(9):1012. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11091012
Chicago/Turabian StyleYvert, Thomas, Eri Miyamoto-Mikami, Takuro Tobina, Keisuke Shiose, Ryo Kakigi, Takamasa Tsuzuki, Mizuki Takaragawa, Noriko Ichinoseki-Sekine, Margarita Pérez, Hiroyuki Kobayashi, and et al. 2020. "PPARGC1A rs8192678 and NRF1 rs6949152 Polymorphisms Are Associated with Muscle Fiber Composition in Women" Genes 11, no. 9: 1012. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11091012