
Analysis of the Differential Gene and Protein Expression Profiles of Corneal Epithelial Cells Stimulated with Alternating Current Electric Fields

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Alternating Current (AC) Electric Field (EF) Stimulation
2.3. Microarray Hybridization
2.4. Microarray Data Analysis
2.5. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.6. Antibody MicroarrayAanalysis
3. Results
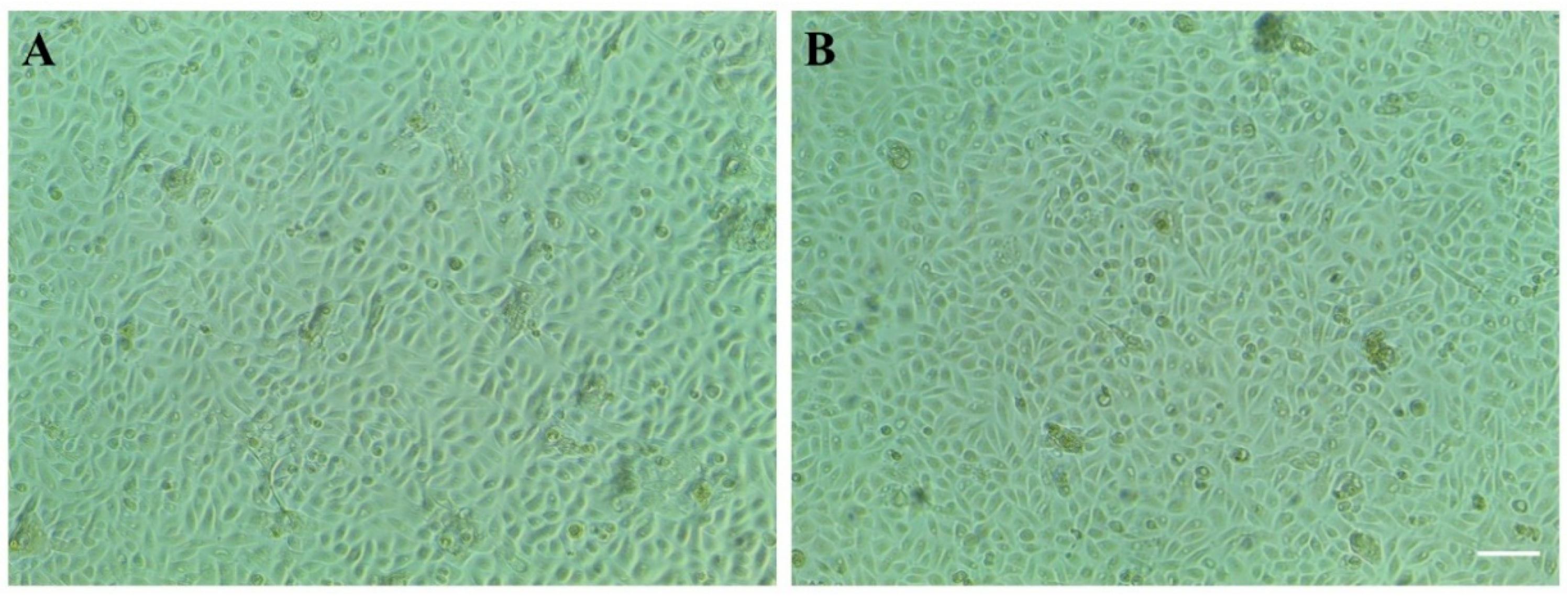
3.1. Stimulation of Telomerase-Immortalized Human Corneal Epithelial (hTCEpi) Cells with Alternate Current (AC) Electric Fields (EFs)
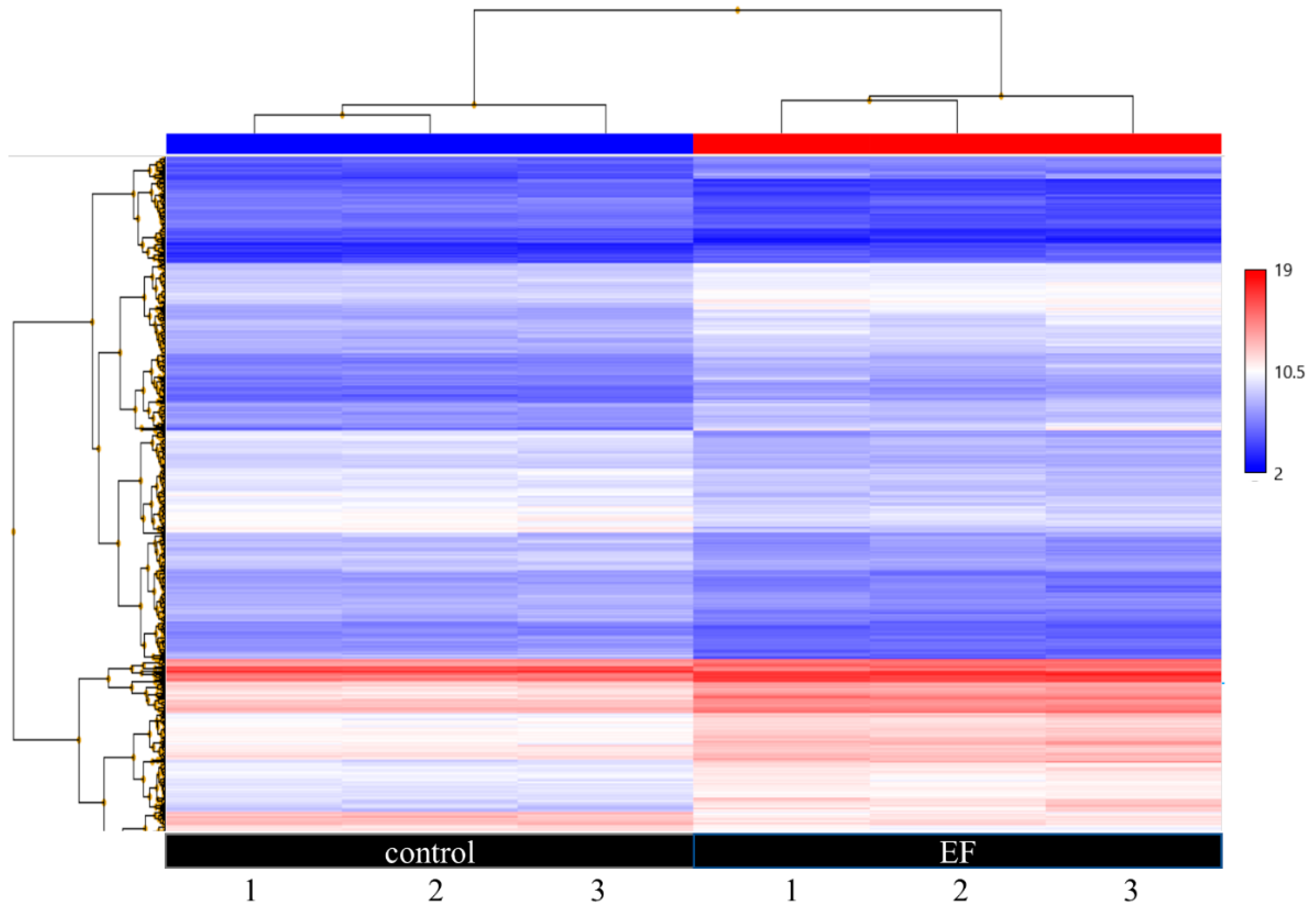
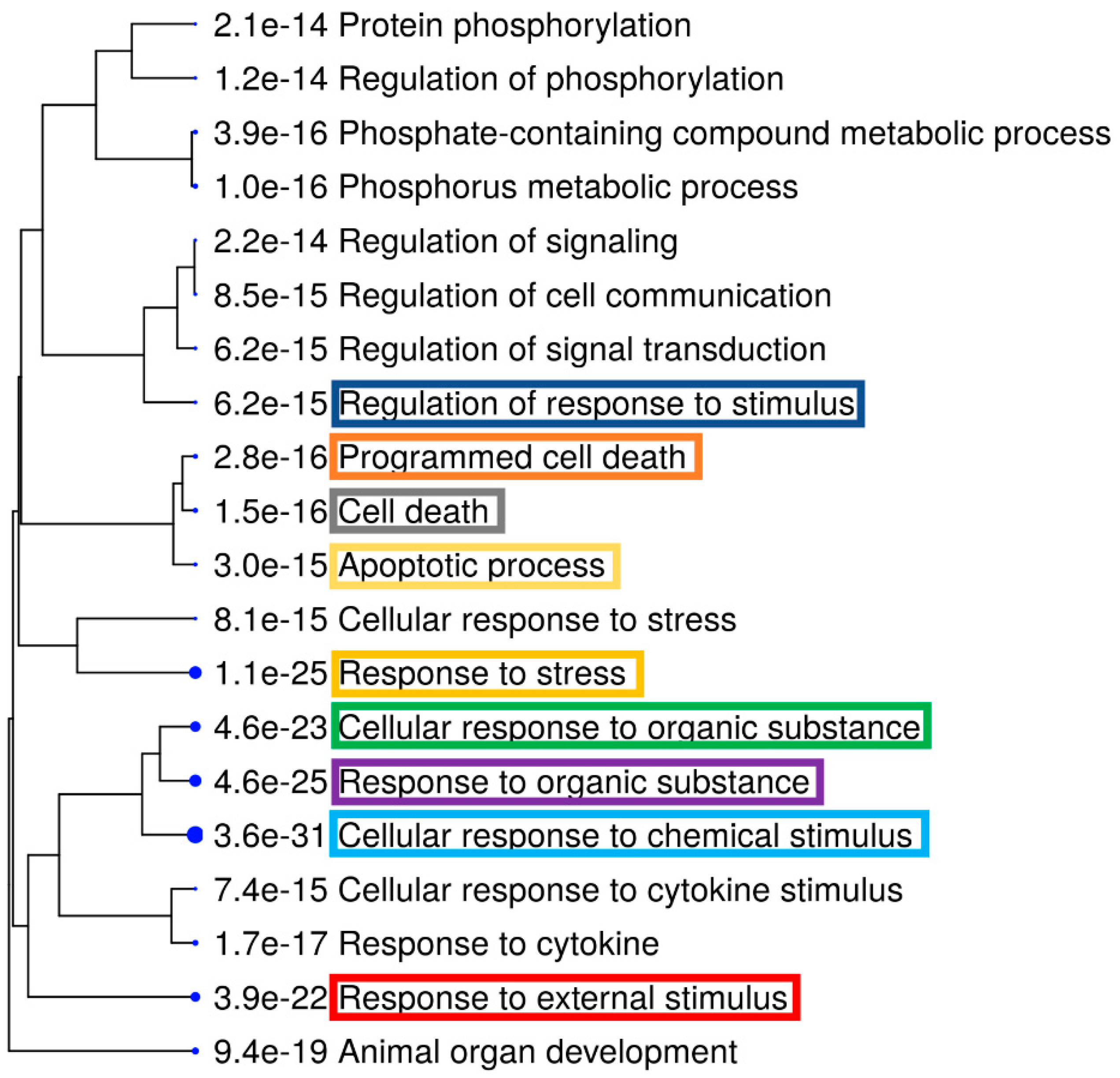
3.2. Gene Expession Profiling (GEP) of AC EF-Stimulated hTCEpi Cells
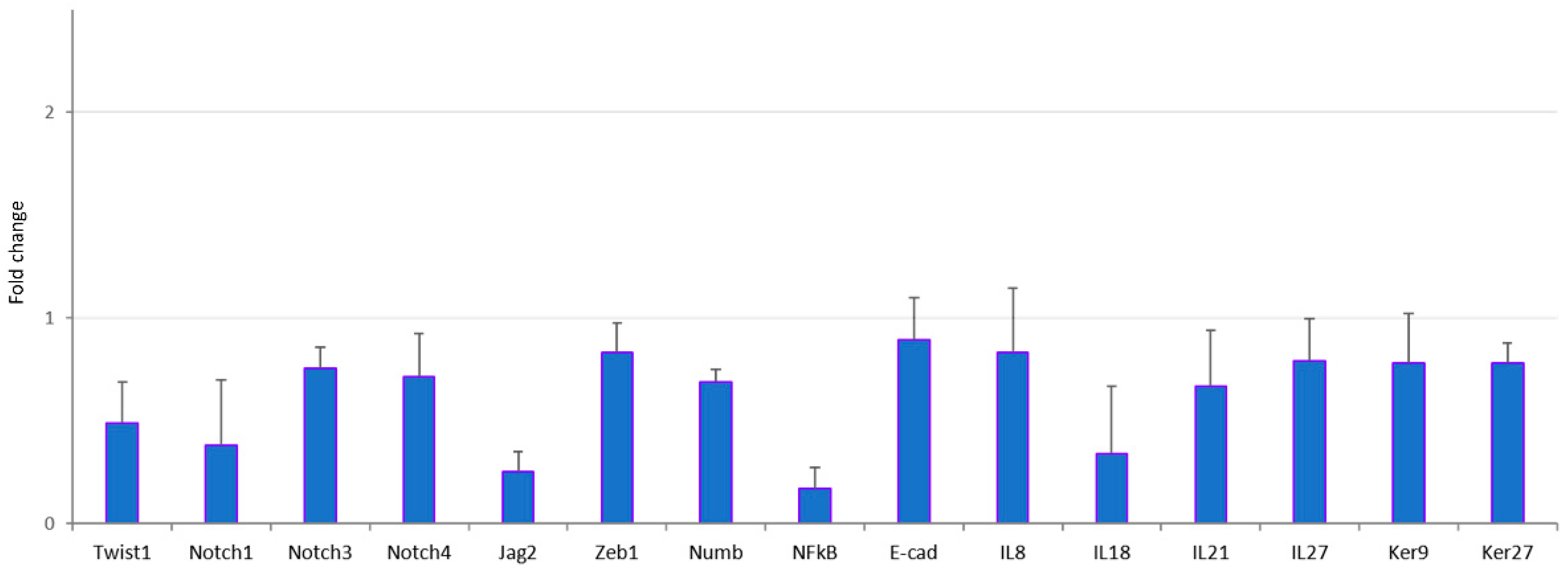
3.3. Validation of Trancriptome Analysis Data by qRT-PCR
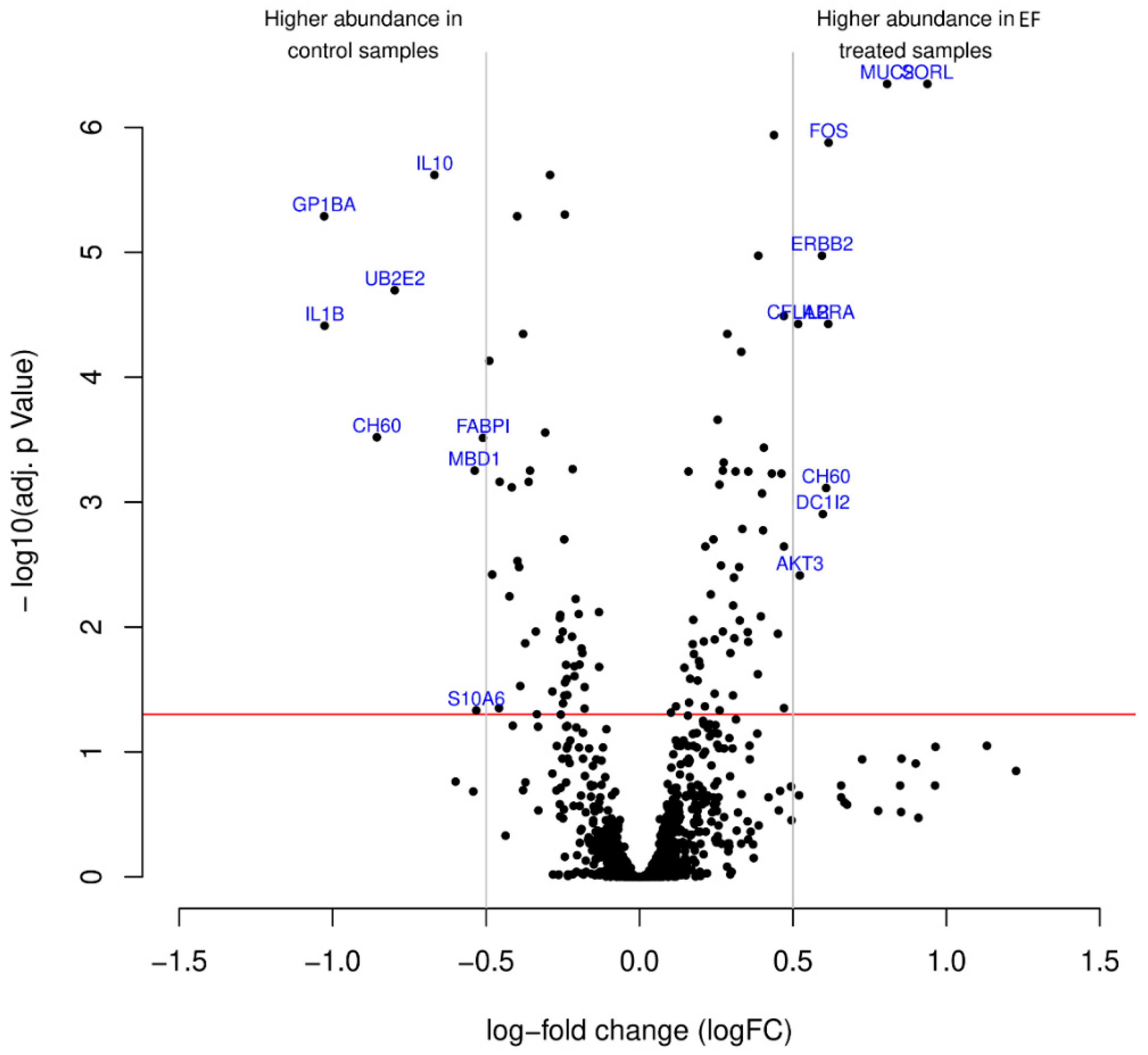
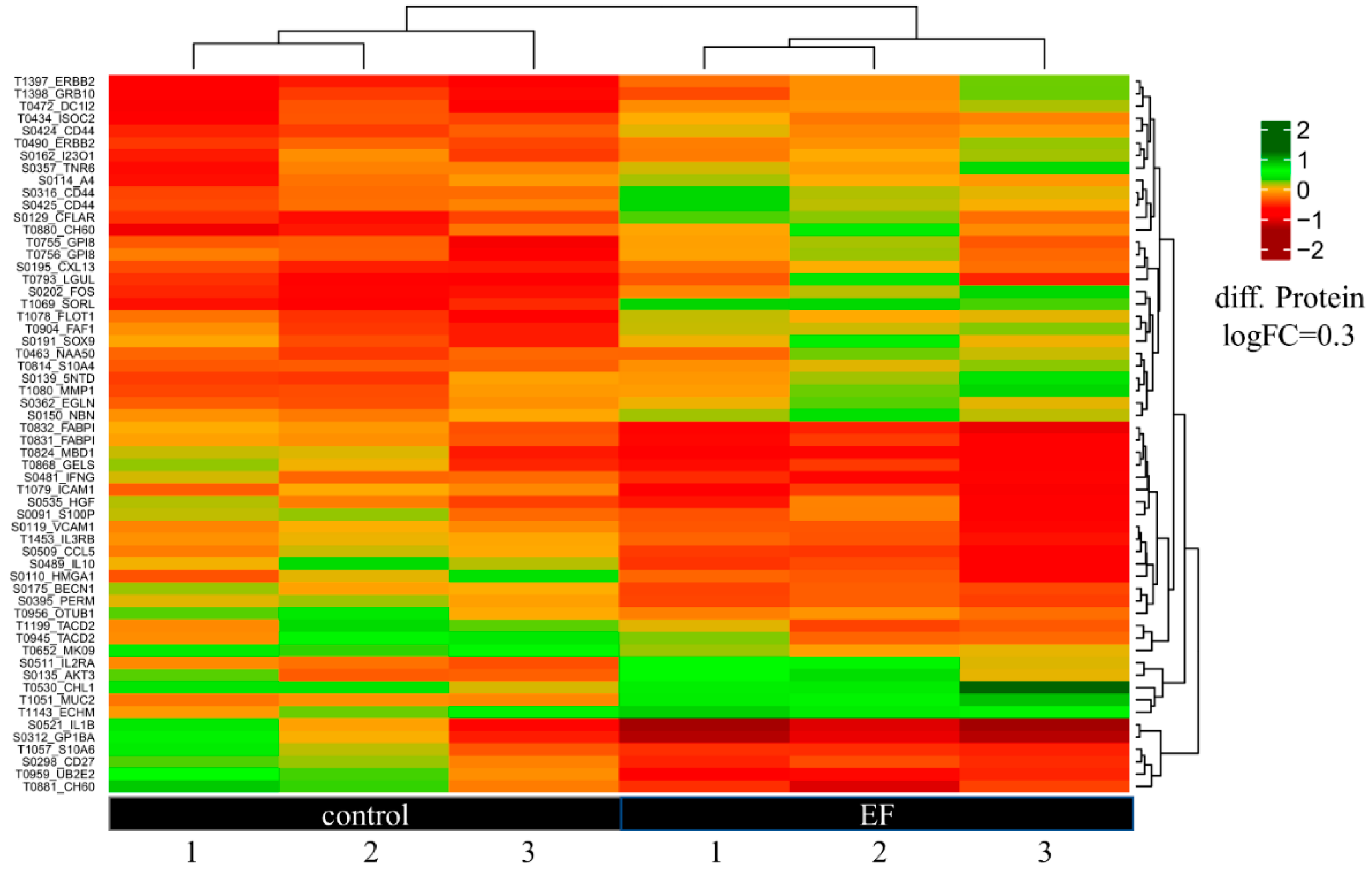
3.4. Antibody Microarray Analysis of Differentially Expressed Proteins in hTCEpi Cells
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reid, B.J.; Zhao, M. The Electrical Response to Injury: Molecular Mechanisms and Wound Healing. Adv. Wound Care 2014, 3, 184–201. [Google Scholar] [CrossRef] [Green Version]
- Vieira, A.C.; Reid, B.; Cao, L.; Mannis, M.J.; Schwab, I.R.; Zhao, M. Ionic Components of Electric Current at Rat Corneal Wounds. PLoS ONE 2011, 6, e17411. [Google Scholar] [CrossRef] [Green Version]
- Nuccitelli, R. Physiological Electric Fields can Influence Cell Motility, Growth, and Polarity. Adv. Mol. Cell Biol. 1988, 2, 213–233. [Google Scholar] [CrossRef]
- Ferrier, J.; Ross, S.M.; Kanehisa, J.; Aubin, J.E. Osteoclasts and osteoblasts migrate in opposite directions in response to a constant electrical field. J. Cell. Physiol. 1986, 129, 283–288. [Google Scholar] [CrossRef]
- Reid, B.; Graue-Hernandez, E.O.; Mannis, M.J.; Zhao, M. Modulating Endogenous Electric Currents in Human Corneal Wounds—A Novel Approach of Bioelectric Stimulation Without Electrodes. Cornea 2011, 30, 338–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Chalmers, L.; Cao, L.; Vieira, A.C.; Mannis, M.; Reid, B. Electrical signaling in control of ocular cell behaviors. Prog. Retin. Eye Res. 2012, 31, 65–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Mehta, A.S.; Zhao, M. Biomedical applications of electrical stimulation. Cell. Mol. Life Sci. 2020, 77, 2681–2699. [Google Scholar] [CrossRef]
- Soong, H.K.; Parkinson, W.C.; Bafna, S.; Sulik, G.L.; Huang, S.C. Movements of cultured corneal epithelial cells and stromal fibroblasts in electric fields. Investig. Ophthalmol. Vis. Sci. 1990, 31, 2278–2282. [Google Scholar]
- Zhao, M.; Forrester, J.V.; McCaig, C.D. A small, physiological electric field orients cell division. Proc. Natl. Acad. Sci. USA 1999, 96, 4942–4946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Dick, A.; Forrester, J.V.; McCaig, C.D. Electric Field–directed Cell Motility Involves Up-regulated Expression and Asymmetric Redistribution of the Epidermal Growth Factor Receptors and Is Enhanced by Fibronectin and Laminin. Mol. Biol. Cell 1999, 10, 1259–1276. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Agius-Fernandez, A.; Forrester, J.V.; McCaig, C.D. Orientation and directed migration of cultured corneal epithelial cells in small electric fields are serum dependent. J. Cell Sci. 1996, 109, 1405–1414. [Google Scholar]
- Zhao, M.; McCaig, C.D.; Agius-Fernandez, A.; Forrester, J.V.; Araki-Sasaki, K. Human corneal epithelial cells reorient and migrate cathodally in a small applied electric field. Curr. Eye Res. 1997, 16, 973–984. [Google Scholar] [CrossRef] [PubMed]
- Iglesia, D.D.S.; Vanable, J.W. Endogenous lateral electric fields around bovine corneal lesions are necessary for and can enhance normal rates of wound healing. Wound Repair Regen. 1998, 6, 531–542. [Google Scholar] [CrossRef]
- Reid, B.; Song, B.; McCaig, C.D.; Zhao, M. Wound healing in rat cornea: The role of electric currents. FASEB J. 2004, 19, 379–386. [Google Scholar] [CrossRef]
- Miyake, K.-I.; Yoshida, M.; Inoue, Y.; Hata, Y. Neuroprotective Effect of Transcorneal Electrical Stimulation on the Acute Phase of Optic Nerve Injury. Investig. Ophthalmol. Vis. Sci. 2007, 48, 2356–2361. [Google Scholar] [CrossRef] [Green Version]
- Song, B.; Zhao, M.; Forrester, J.; McCaig, C. Nerve regeneration and wound healing are stimulated and directed by an endogenous electrical field in vivo. J. Cell Sci. 2004, 117, 4681–4690. [Google Scholar] [CrossRef] [Green Version]
- Shapira, Y.; Sammons, V.; Forden, J.; Guo, G.F.; Kipp, A.; Girgulis, J.; Mishra, T.; de Villers Alant, J.D.; Midha, R. Brief Electrical Stimulation Promotes Nerve Regeneration Following Experimental In-Continuity Nerve Injury. Neurosurgery 2018, 85, 156–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariza, C.A.; Fleury, A.T.; Tormos, C.J.; Petruk, V.; Chawla, S.; Oh, J.; Sakaguchi, D.S.; Mallapragada, S.K. The Influence of Electric Fields on Hippocampal Neural Progenitor Cells. Stem Cell Rev. Rep. 2010, 6, 585–600. [Google Scholar] [CrossRef] [PubMed]
- Biasiotta, A.; D’Arcangelo, D.; Passarelli, F.; Nicodemi, E.M.; Facchiano, A. Ion channels expression and function are strongly modified in solid tumors and vascular malformations. J. Transl. Med. 2016, 14, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.-H.; McCullen, S.D.; Piedrahita, J.A.; Loboa, E.G.; Olby, N.J. Alternating Current Electric Fields of Varying Frequencies: Effects on Proliferation and Differentiation of Porcine Neural Progenitor Cells. Cell. Reprogramming 2013, 15, 405–412. [Google Scholar] [CrossRef] [Green Version]
- McCullen, S.D.; McQuilling, J.P.; Grossfeld, R.M.; Lubischer, J.L.; Clarke, L.I.; Loboa, E.G. Application of Low-Frequency Alternating Current Electric Fields Via Interdigitated Electrodes: Effects on Cellular Viability, Cytoplasmic Calcium, and Osteogenic Differentiation of Human Adipose-Derived Stem Cells. Tissue Eng. Part C Methods 2010, 16, 1377–1386. [Google Scholar] [CrossRef] [Green Version]
- Sahm, F.; Ziebart, J.; Jonitz-Heincke, A.; Hansmann, D.; Dauben, T.; Bader, R. Alternating Electric Fields Modify the Function of Human Osteoblasts Growing on and in the Surroundings of Titanium Electrodes. Int. J. Mol. Sci. 2020, 21, 6944. [Google Scholar] [CrossRef] [PubMed]
- Fish, R.M.; Geddes, L.A. Conduction of Electrical Current to and Through the Human Body: A Review. Eplasty 2009, 9, 44. [Google Scholar]
- Yan, Y.; Neville, A.; Dowson, D. Biotribocorrosion—an appraisal of the time dependence of wear and corrosion interactions: I. The role of corrosion. J. Phys. D Appl. Phys. 2006, 39, 3200–3205. [Google Scholar] [CrossRef]
- Al-Majed, A.A.; Tam, S.L.; Gordon, T. Electrical Stimulation Accelerates and Enhances Expression of Regeneration-Associated Genes in Regenerating Rat Femoral Motoneurons. Cell. Mol. Neurobiol. 2004, 24, 379–402. [Google Scholar] [CrossRef]
- Hiemer, B.; Krogull, M.; Bender, T.; Ziebart, J.; Krueger, S.; Bader, R.; Jonitz-Heincke, A. Effect of electric stimulation on human chondrocytes and mesenchymal stem cells under normoxia and hypoxia. Mol. Med. Rep. 2018, 18, 2133–2141. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Bai, X.; Ding, Y.; Lee, I.-S. Electrical stimulation as a novel tool for regulating cell behavior in tissue engineering. Biomater. Res. 2019, 23, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashrafi, M.; Alonso-Rasgado, T.; Baguneid, M.; Bayat, A. The efficacy of electrical stimulation in lower extremity cutaneous wound healing: A systematic review. Exp. Dermatol. 2017, 26, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Souffrant, R.; Kluess, D.; Ellenrieder, M.; Mittelmeier, W.; Van Rienen, U.; Bader, R. Evaluation of electric field distribution in electromagnetic stimulation of human femoral head. Bioelectromagnetics 2014, 35, 547–558. [Google Scholar] [CrossRef]
- Robertson, D.M.; Li, L.; Fisher, S.; Pearce, V.P.; Shay, J.W.; Wright, W.E.; Cavanagh, H.D.; Jester, J.V. Characterization of Growth and Differentiation in a Telomerase-Immortalized Human Corneal Epithelial Cell Line. Investig. Ophthalmol. Vis. Sci. 2005, 46, 470–478. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, B.S.; Irizarry, R.A. A framework for oligonucleotide microarray preprocessing. Bioinformatics 2010, 26, 2363–2367. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R Package for Comparing Biological Themes Among Gene Clusters. OMICS J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Reid, B.R.; Nuccitelli, R.; Zhao, M. Non-invasive measurement of bioelectric currents with a vibrating probe. Nat. Protoc. 2007, 2, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Tai, G.; Tai, M.; Zhao, M. Electrically stimulated cell migration and its contribution to wound healing. Burn. Trauma 2018, 6, 20. [Google Scholar] [CrossRef] [Green Version]
- McCaig, C.D. Dynamic aspects of amphibian neurite growth and the effects of an applied electric field. J. Physiol. 1986, 375, 55–69. [Google Scholar] [CrossRef] [Green Version]
- Yao, L.; Shanley, L.; McCaig, C.; Zhao, M. Small applied electric fields guide migration of hippocampal neurons. J. Cell. Physiol. 2008, 216, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Matos, M.A.; Cicerone, M.T. Alternating current electric field effects on neural stem cell viability and differentiation. Biotechnol. Prog. 2010, 26, 664–670. [Google Scholar] [CrossRef]
- Park, J.S.; Park, K.; Moon, H.T.; Woo, D.G.; Na Yang, H.; Park, K.-H. Electrical Pulsed Stimulation of Surfaces Homogeneously Coated with Gold Nanoparticles to Induce Neurite Outgrowth of PC12 Cells. Langmuir 2009, 25, 451–457. [Google Scholar] [CrossRef]
- Farboudab, B.; Nuccitellia, R.; Schwab, I.R.; Isseroff, R. DC Electric Fields Induce Rapid Directional Migration in Cultured Human Corneal Epithelial Cells. Exp. Eye Res. 2000, 70, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.-T.; Zhao, M. Regulation of tissue repair and regeneration by electric fields. Chin. J. Traumatol. 2010, 13, 55–61. [Google Scholar]
- Mulholland, B.; Tuft, S.J.; Khaw, P.T. Matrix metalloproteinase distribution during early corneal wound healing. Eye 2004, 19, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Raghunathan, V.K.; Reid, B.; Wei, D.; Diaz, R.C.; Russell, P.; Murphy, C.J.; Zhao, M. Biomimetic stochastic to-pography and electric fields synergistically enhance directional migration of corneal epithelial cells in a mmp-3-dependent manner. Acta Biomater. 2015, 12, 102–112. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Hu, C.; Lu, C.; Zhang, K.; Han, R.; Lin, C.; Zhao, S.; A, C.; Cheng, C.; Zhao, M.; et al. Applied electric fields suppress osimertinib-induced cytotoxicity via inhibiting FOXO3a nuclear translocation through AKT activation. Carcinogenesis 2019, 41, 600–610. [Google Scholar] [CrossRef]
- Meng, X.; Arocena, M.; Penninger, J.; Gage, F.H.; Zhao, M.; Song, B. PI3K mediated electrotaxis of embryonic and adult neural progenitor cells in the presence of growth factors. Exp. Neurol. 2011, 227, 210–217. [Google Scholar] [CrossRef] [Green Version]
- Yao, L.; Li, Y.; Knapp, J.; Smith, P. Exploration of molecular pathways mediating electric field-directed schwann cell migration by RNA-seq. J. Cell. Physiol. 2015, 230, 1515–1524. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Guan, L.; Fan, P.; Liu, X.; Liu, R.; Liu, Y.; Bai, H. Direct Current Electric Field Stimulates Nitric Oxide Production and Promotes NO-Dependent Angiogenesis: Involvement of the PI3K/Akt Signaling Pathway. J. Vasc. Res. 2020, 57, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Song, B.; Pu, J.; Wada, T.; Reid, B.; Tai, G.; Wang, F.; Guo, A.; Walczysko, P.; Gu, Y.; et al. Electrical signals control wound healing through phosphatidylinositol-3-OH kinase-γ and PTEN. Nat. Cell Biol. 2006, 442, 457–460. [Google Scholar] [CrossRef]
- Yamamoto, K.; Yamamoto, T.; Honjo, K.; Ichioka, H.; Oseko, F.; Kishida, T.; Mazda, O.; Kanamura, N. Electrical stimulation with periodic alternating intervals stimulates neuronal cells to produce neurotrophins and cytokines through activation of mitogen-activated protein kinase pathways. Eur. J. Oral Sci. 2015, 123, 403–408. [Google Scholar] [CrossRef]
- Kawamura, K.; Kano, Y. Electrical stimulation induces neurite outgrowth in PC12m3 cells via the p38 mitogen-activated protein kinase pathway. Neurosci. Lett. 2019, 698, 81–84. [Google Scholar] [CrossRef]
- Geng, K.; Wang, J.; Liu, P.; Tian, X.; Liu, H.; Wang, X.; Hu, C.; Yan, H. Electrical stimulation facilitates the angiogenesis of human umbilical vein endothelial cells through MAPK/ERK signaling pathway by stimulating FGF2 secretion. Am. J. Physiol. Physiol. 2019, 317, C277–C286. [Google Scholar] [CrossRef]
- English, A.W.; Schwartz, G.; Meador, W.; Sabatier, M.J.; Mulligan, A. Electrical stimulation promotes peripheral axon regeneration by enhanced neuronal neurotrophin signaling. Dev. Neurobiol. 2007, 67, 158–172. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Xu, Q.-G.; Franz, C.K.; Zhang, R.; Dalton, C.; Gordon, T.; Verge, V.M.K.; Midha, R.; Zochodne, D.W. Accelerated axon outgrowth, guidance, and target reinnervation across nerve transection gaps following a brief electrical stimulation paradigm. J. Neurosurg. 2012, 116, 498–512. [Google Scholar] [CrossRef]
- Ju, C.; Park, E.; Kim, T.; Kim, T.; Kang, M.; Lee, K.-S.; Park, S.-M. Effectiveness of electrical stimulation on nerve regeneration after crush injury: Comparison between invasive and non-invasive stimulation. PLoS ONE 2020, 15, e0233531. [Google Scholar] [CrossRef]
- Zuo, K.J.; Gordon, T.; Chan, K.M.; Borschel, G.H. Electrical stimulation to enhance peripheral nerve regeneration: Update in molecular investigations and clinical translation. Exp. Neurol. 2020, 332, 113397. [Google Scholar] [CrossRef]
- Gordon, T. Electrical stimulation to enhance axon regeneration after peripheral nerve injuries in animal models and humans. Neurotherapeutics 2016, 13, 295–310. [Google Scholar] [CrossRef] [Green Version]
- Jara, J.S.; Agger, S.; Hollis, E.R.I. Functional Electrical Stimulation and the Modulation of the Axon Regeneration Program. Front. Cell Dev. Biol. 2020, 8, 736. [Google Scholar] [CrossRef]
- Gygi, S.P.; Rochon, Y.; Franza, B.R.; Aebersold, R. Correlation between Protein and mRNA Abundance in Yeast. Mol. Cell. Biol. 1999, 19, 1720–1730. [Google Scholar] [CrossRef] [Green Version]
- Greenbaum, D.; Colangelo, C.; Williams, K.; Gerstein, M. Comparing protein abundance and mRNA expression levels on a genomic scale. Genome Biol. 2003, 4, 117. [Google Scholar] [CrossRef] [Green Version]
- Waters, K.M.; Pounds, J.G.; Thrall, B.D. Data merging for integrated microarray and proteomic analysis. Brief. Funct. Genom. Proteom. 2006, 5, 261–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyler, S.E.B. Nature’s Electric Potential: A Systematic Review of the Role of Bioelectricity in Wound Healing and Regenerative Processes in Animals, Humans, and Plants. Front. Physiol. 2017, 8, 02017. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Primer | Primer Sequence (5′-3′) | GenBank Accession |
|---|---|---|---|
| Twist1 | Fwd | CAGCCTGAGCAACAGCGAGGA | NM_000474 |
| Rev | CGAACGCCTCGTTCAGCGAC | ||
| Notch1 | Fwd | GCAGTGCTTCCAGAGTGCCACC | NM_017617 |
| Rev | AACCGGAACTTCTTGGTCTCCAGG | ||
| Notch3 | Fwd | GGCCAGGCCATGGTCTTCC | NM_000435 |
| Rev | TCAGGGAACCAGAGGGTGCTG | ||
| Notch4 | Fwd | AGGCTGGAGCCAACCCCAAC | NM_004557 |
| Rev | CAGCCCCTTCCAGCAGCGT | ||
| Jag2 | Fwd | CCTGATTGGCGGCTATTACTGTGA | NM_002226 |
| Rev | CCCGCAGCCATCGATCACTC | ||
| Zeb1 | Fwd | GCAGGGCACACCAGAAGCCA | NM_001128128 |
| Rev | GTGAGCTATAGGAGCCAGAATGGGAA | ||
| Numb | Fwd | CAAGCTAATGGCACTGACTCAGCC | NM_001005743 |
| Rev | CTGAGGGTGGCCTGCAGTGC | ||
| NFkB | Fwd | GCACCTAGCTGCCAAAGAAGGAC | NM_001165412 |
| Rev | GCTCCTGCTGCTTTGAGAAGAGC | ||
| E-cadherin | Fwd | GCTTGGATTTTGAGGCCAAGCA | NM_004360 |
| Rev | AATCTCCAGCCAGTTGGCAGTGTC | ||
| IL8 | Fwd | CGTGGCTCTCTTGGCAGCC | NM_000584 |
| Rev | TGTGTTGGCGCAGTGTGGTC | ||
| IL18 | Fwd | CCAAGGAAATCGGCCTCTATTTG | NM_001562 |
| Rev | TCAGGAGGATTCATTTCCTTAAAGGA | ||
| IL21 | Fwd | GCCACATGATTAGAATGCGTCAAC | NM_021803 |
| Rev | TGTATTTGCTGACTTTAGTTGGGCC | ||
| IL27 | Fwd | CGGAGCGTCTCTGCTTCATCTC | NM_145659 |
| Rev | AGGTTGAATCCTGCAGCCAGC | ||
| Ker9 | Fwd | CTCTCCTGGACATTGACAACACTCGC | NM_000226 |
| Rev | AGCACCTGCCGCAGGCCAT | ||
| Ker27 | Fwd | GCGAGCTGAGTACGAAGCCCTCG | NM_181537 |
| Rev | CTCATTCCGGGCTGAGGTGGTG | ||
| GAPDH | Fwd | GGATATTGTTGCCATCAATGACCC | NM_002046 |
| Rev | TCTCGCTCCTGGAAGATGGTGA |
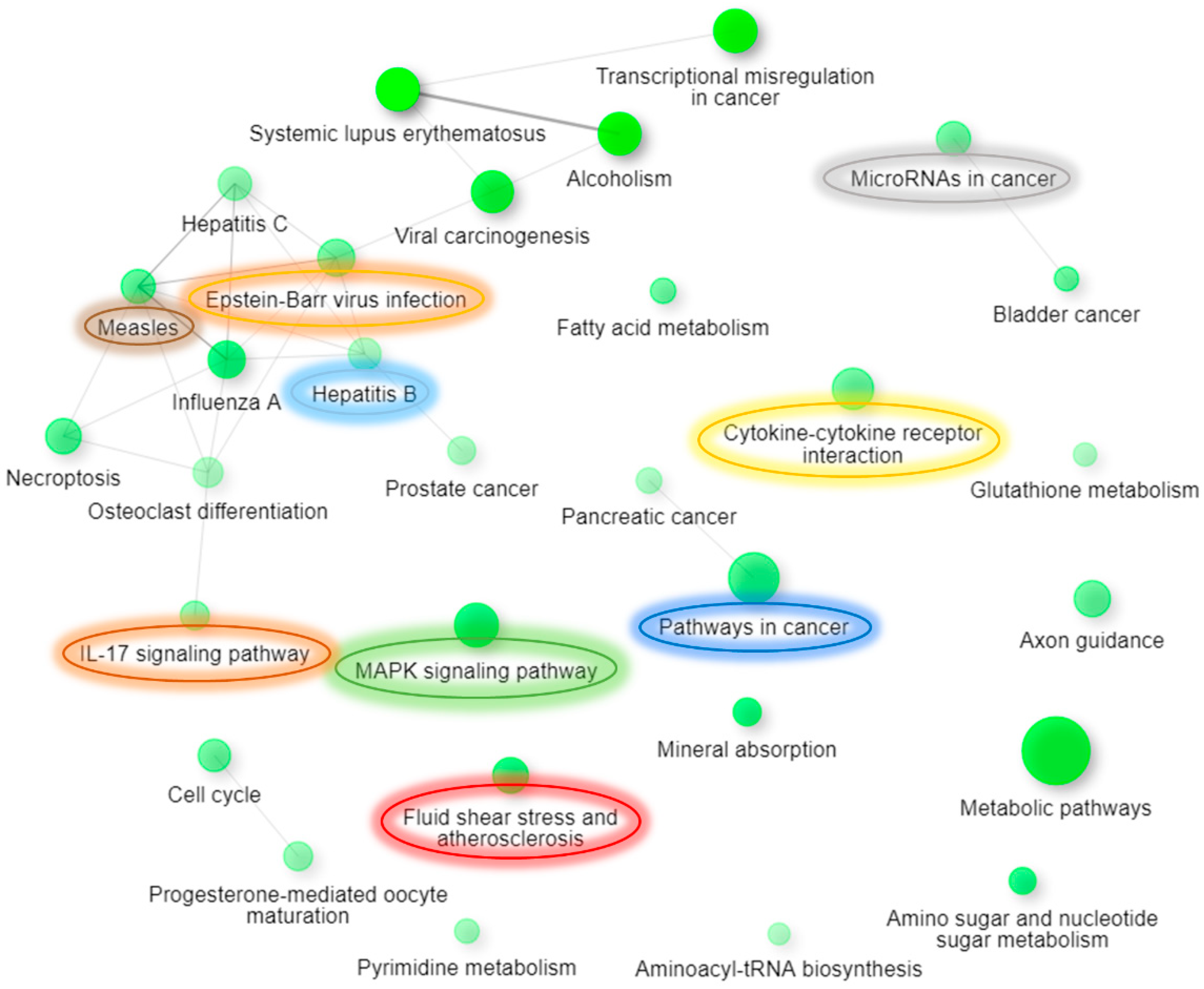
| KEGG Pathway | Pathway Description | Observed Gene Count | False Discovery Rate |
|---|---|---|---|
| hsa00520 | Amino sugar and nucleotide sugar metabolism | 13 of 48 | 0.0036 |
| hsa04010 | MAPK signaling pathway | 35 of 293 | 0.0036 |
| hsa04978 | Mineral absorption | 13 of 51 | 0.0036 |
| hsa05162 | Measles | 21 of 133 | 0.0036 |
| hsa05164 | Influenza A | 25 of 168 | 0.0036 |
| hsa05202 | Transcriptional misregulation in cancer | 25 of 169 | 0.0036 |
| hsa05418 | Fluid shear stress and atherosclerosis | 22 of 133 | 0.0036 |
| hsa01100 | Metabolic pathways | 100 of 1250 | 0.0074 |
| hsa01212 | Fatty acid metabolism | 11 of 48 | 0.0074 |
| hsa05160 | Hepatitis C | 19 of 131 | 0.0074 |
| hsa05200 | Pathways in cancer | 49 of 515 | 0.0074 |
| hsa05203 | Viral carcinogenesis | 24 of 183 | 0.0074 |
| hsa05206 | MicroRNAs in cancer | 21 of 149 | 0.0074 |
| hsa05219 | Bladder cancer | 10 of 41 | 0.0074 |
| hsa04110 | Cell cycle | 18 of 123 | 0.0076 |
| hsa05168 | Herpes simplex infection | 23 of 181 | 0.0076 |
| hsa04360 | Axon guidance | 22 of 173 | 0.0093 |
| hsa04657 | IL-17 signaling pathway | 14 of 92 | 0.0194 |
| hsa04060 | Cytokine-cytokine receptor interaction | 28 of 263 | 0.0206 |
| hsa04914 | Progesterone-mediated oocyte maturation | 14 of 94 | 0.021 |
| hsa00970 | Aminoacyl-tRNA biosynthesis | 9 of 44 | 0.0213 |
| hsa05161 | Hepatitis B | 18 of 142 | 0.0229 |
| hsa01040 | Biosynthesis of unsaturated fatty acids | 6 of 23 | 0.0426 |
| hsa05322 | Systemic lupus erythematosus | 13 of 94 | 0.0454 |
| hsa04217 | Necroptosis | 18 of 155 | 0.0474 |
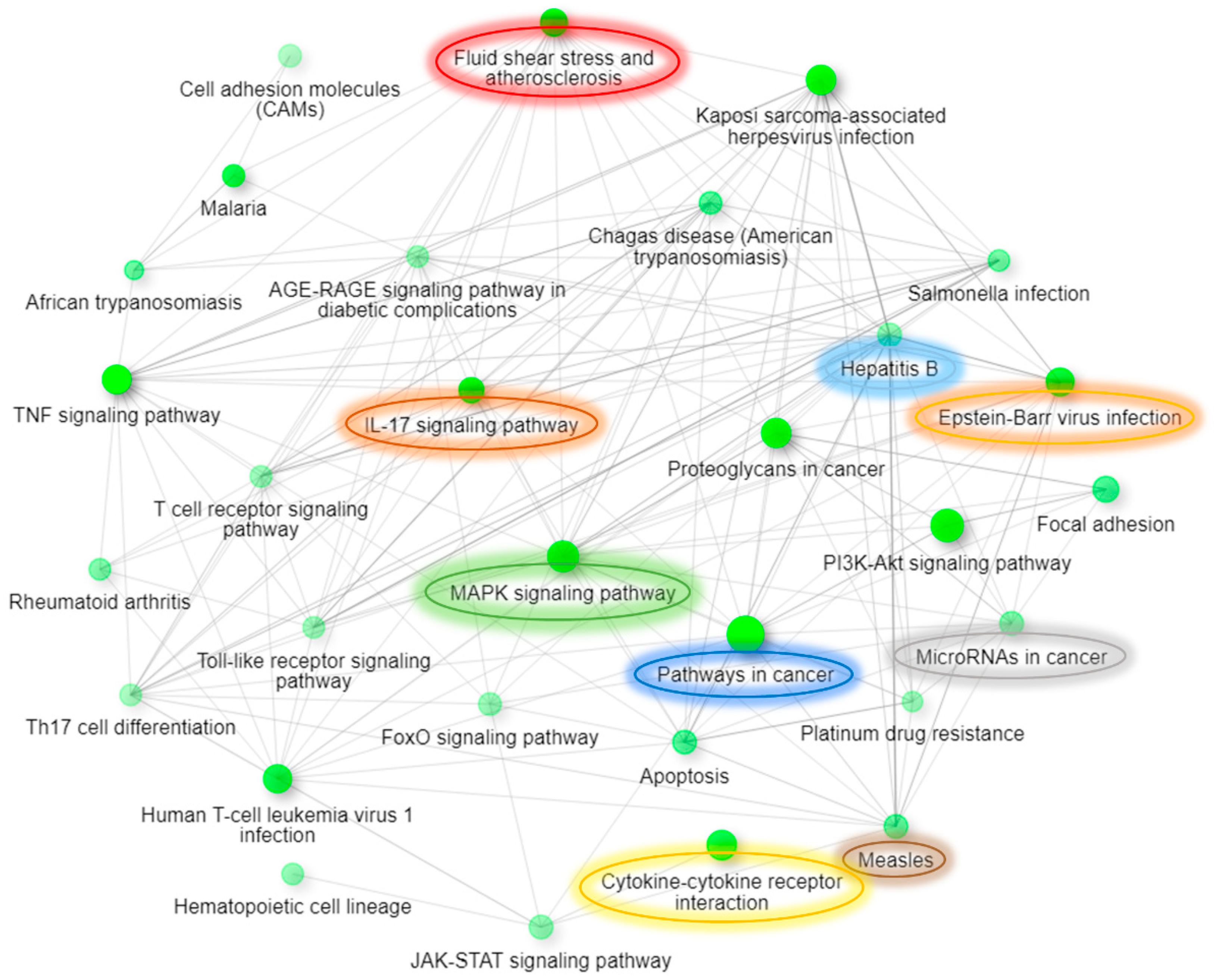
| KEGG Pathway | Pathway Description | Protein Count | False Discovery Rate | Matching Proteins in Network |
|---|---|---|---|---|
| hsa04668 | TNF signaling pathway | 17 | 1.95 × 10−13 | MAPK12, MAPK1, AKT3, ICAM1, VCAM1, IL15, CSF2, FOS, CASP3, CFLAR, SELE, FAS, MAPK9, TNF, MAP2K4, PGAM5, CCL5 |
| hsa05200 | Pathways in cancer | 27 | 2.95 × 10−11 | MAPK1, HGF, IFNG, CCND2, AKT3, IL7, FGF2, STAT3, TP53, ERBB2, IL15, HSP90B1, IL7R, FOS, CASP3, GSTM1, KLK3, MET, MMP1, CASP9, FAS, RAC1, IL2RA, ETS1, JAK3, MAPK9, CUL2 |
| hsa05418 | Fluid shear stress and atherosclerosis | 16 | 2.95 × 10−11 | MAPK12, IFNG, AKT3, KDR, ICAM1, TP53, VCAM1, HSP90B1, FOS, GSTM1, SELE, CAV3, RAC1, MAPK9, TNF, MAP2K4 |
| hsa04010 | MAPK signaling pathway | 21 | 3.06 × 10−11 | MAPK12, MAPK1, HGF, HSPB1, AKT3, KDR, FGF2, TP53, ERBB2, CD14, FOS, CASP3, MET, FAS, RAC1, HSPA1A, MAPK9, TNF, MAP2K4, DDIT3, NTF4 |
| hsa04060 | Cytokine-cytokine receptor interaction | 20 | 3.06 × 10−11 | HGF, IFNG, TNFSF10, CCR7, IL7, KDR, CXCL13, IL15, CSF2, CCL11, IL7R, MET, FAS, IL2RA, CCL8, TNF, IL10, TNFSF14, CCL5, TNFRSF9 |
| hsa04657 | IL-17 signaling pathway | 13 | 2.29 × 10−10 | MAPK12, MAPK1, IFNG, CSF2, HSP90B1, CCL11, FOS, CASP3, MMP1, MAPK4, MAPK9, TNF, MUC5B |
| hsa04151 | PI3K-Akt signaling pathway | 21 | 3.07 × 10−10 | MAPK1, HGF, VTN, CCND2, AKT3, IL7, KDR, FGF2, TP53, ERBB2, ITGA5, HSP90B1, IL7R, MET, CASP9, RAC1, THBS3, IL2RA, JAK3, PPP2R5D, NTF4 |
| hsa04933 | AGE-RAGE signaling pathway in diabetic complications | 11 | 4.79 × 10−8 | MAPK12, MAPK1, AKT3, STAT3, ICAM1, VCAM1, CASP3, SELE, RAC1, MAPK9, TNF |
| hsa04660 | T cell receptor signaling pathway | 11 | 4.95 × 10−8 | MAPK12, MAPK1, IFNG, AKT3, CSF2, FOS, CD28, MAPK9, TNF, PTPRC, IL10 |
| hsa04620 | Toll-like receptor signaling pathway | 11 | 6.21 × 10−8 | MAPK12, MAPK1, AKT3, CD14, FOS, CD86, RAC1, MAPK9, TNF, MAP2K4, CCL5 |
| hsa04510 | Focal adhesion | 14 | 6.84 × 10−8 | MAPK1, HGF, VTN, CCND2, AKT3, KDR, ERBB2, ITGA5, MET, CAV3, RAC1, THBS3, ACTN1, MAPK9 |
| hsa05164 | Influenza A | 13 | 8.98 × 10−8 | MAPK12, MAPK1, IFNG, TNFSF10, AKT3, ICAM1, CASP9, FAS, HSPA1A, MAPK9, TNF, MAP2K4, CCL5 |
| hsa04630 | Jak-STAT signaling pathway | 12 | 3.49 × 10−7 | IFNG, CCND2, AKT3, IL7, STAT3, IL15, CSF2, IL7R, AOX1, IL2RA, JAK3, IL10 |
| hsa04068 | FoxO signaling pathway | 11 | 3.99 × 10−7 | MAPK12, MAPK1, TNFSF10, CCND2, AKT3, STAT3, ATM, IL7R, BNIP3, MAPK9, IL10 |
| hsa05162 | Measles | 11 | 4.63 × 10−7 | IFNG, TNFSF10, CCND2, AKT3, STAT3, TP53, CD28, FAS, HSPA1A, IL2RA, JAK3 |
| hsa05161 | Hepatitis B | 11 | 8.35 × 10−7 | MAPK1, AKT3, STAT3, TP53, FOS, CASP3, CASP9, FAS, MAPK9, TNF, MAP2K4 |
| hsa04664 | Fc epsilon RI signaling pathway | 8 | 2.19 × 10−6 | MAPK12, MAPK1, AKT3, CSF2, RAC1, MAPK9, TNF, MAP2K4 |
| hsa05168 | Herpes simplex infection | 11 | 6.12 × 10−6 | CDC34, IFNG, TP53, IL15, FOS, CASP3, FAS, MAPK9, TNF, TNFSF1, CCL5 |
| hsa04514 | Cell adhesion molecules (CAMs) | 10 | 4.92 × 10−6 | SELP, ICAM1, VCAM1, CD28, SELE, CD86, VTCN1, CD99, PDCD1LG2, PTPRC |
| hsa04370 | VEGF signaling pathway | 7 | 9.52 × 10−6 | MAPK12, MAPK1, HSPB1, AKT3, KDR, CASP9, RAC1 |
| hsa04217 | Necroptosis | 10 | 1.05 × 10−5 | IFNG, TNFSF10, STAT3, CFLAR, FAS, FAF1, JAK3, MAPK9, TNF, PGAM5 |
| hsa04115 | p53 signaling pathway | 7 | 1.92 × 10−5 | CCND2, TP53, ATM, CASP3, CASP9, FAS, CHEK2 |
| hsa04917 | Prolactin signaling pathway | 7 | 2.02 × 10−5 | MAPK12, MAPK1, CCND2, AKT3, STAT3, FOS, MAPK9 |
| hsa04062 | Chemokine signaling pathway | 10 | 3.16 × 10−5 | MAPK1, CCR7, AKT3, STAT3, CXCL13, CCL11, RAC1, CCL8, JAK3, CCL5 |
| hsa04512 | ECM-receptor interaction | 7 | 5.07 × 10−5 | VTN, ITGA5, GP1BA, CD47, THBS3, HMMR, CD44 |
| hsa04071 | Sphingolipid signaling pathway | 8 | 5.57 × 10−5 | MAPK12, MAPK1, AKT3, TP53, RAC1, MAPK9, TNF, PPP2R5D |
| hsa04064 | NF-kappa B signaling pathway | 7 | 0.00011 | ICAM1, ATM, VCAM1, CD14, CFLAR, TNF, TNFSF14 |
| hsa04066 | HIF-1 signaling pathway | 7 | 0.00013 | MAPK1, IFNG, ENO1, AKT3, STAT3, ERBB2, CUL2 |
| hsa04014 | Ras signaling pathway | 10 | 0.00016 | MAPK1, HGF, AKT3, KDR, FGF2, MET, RAC1, ETS1, MAPK9, NTF4 |
| hsa04722 | Neurotrophin signaling pathway | 7 | 0.00034 | MAPK12, MAPK1, AKT3, TP53, RAC1, MAPK9, NTF4 |
| hsa04926 | Relaxin signaling pathway | 7 | 0.00066 | MAPK12, MAPK1, AKT3, FOS, MMP1, MAPK9, MAP2K4 |
| hsa04015 | Rap1 signaling pathway | 8 | 0.0015 | MAPK12, MAPK1, HGF, AKT3, KDR, FGF2, MET, RAC1 |
| hsa05219 | Bladder cancer | 4 | 0.0017 | MAPK1, TP53, ERBB2, MMP1 |
| hsa04110 | Cell cycle | 6 | 0.0025 | CCND2, TP53, ATM, CDC25C, CHEK2, CDKN1C |
| hsa04012 | ErbB signaling pathway | 5 | 0.0027 | MAPK1, AKT3, ERBB2, MAPK9, MAP2K4 |
| hsa04550 | Signaling pathways regulating pluripotency of stem cells | 6 | 0.0042 | MAPK12, MAPK1, AKT3, FGF2, STAT3, JAK3 |
| hsa04920 | Adipocytokine signaling pathway | 4 | 0.0089 | AKT3, STAT3, MAPK9, TNF |
| hsa04919 | Thyroid hormone signaling pathway | 5 | 0.0094 | MAPK1, AKT3, NCOR1, TP53, CASP9 |
| hsa04662 | B cell receptor signaling pathway | 4 | 0.0096 | MAPK1, AKT3, FOS, RAC1 |
| hsa04915 | Estrogen signaling pathway | 5 | 0.0154 | MAPK1, AKT3, HSP90B1, FOS, HSPA1A |
| hsa04024 | cAMP signaling pathway | 6 | 0.0179 | MAPK1, SOX9, AKT3, FOS, RAC1, MAPK9 |
| hsa04912 | GnRH signaling pathway | 4 | 0.0179 | MAPK12, MAPK1, MAPK9, MAP2K4 |
| hsa04310 | Wnt signaling pathway | 5 | 0.0197 | MMP7, CCND2, TP53, RAC1, MAPK9 |
| hsa04810 | Regulation of actin cytoskeleton | 6 | 0.0213 | MAPK1, FGF2, ITGA5, CD14, RAC1, ACTN1 |
| hsa04141 | Protein processing in endoplasmic reticulum | 5 | 0.0298 | AMFR, HSP90B1, HSPA1A, MAPK9, DDIT3 |
| hsa04621 | NOD-like receptor signaling pathway | 5 | 0.0329 | MAPK12, MAPK1, MAPK9, TNF, CCL5 |
| hsa04622 | RIG-I-like receptor signaling pathway | 3 | 0.0468 | MAPK12, MAPK9, TNF |
| hsa03320 | PPAR signaling pathway | 3 | 0.0497 | FABP2, FABP5, MMP1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kowtharapu, B.S.; Damaraju, J.; Singh, N.K.; Ziebart, J.; Bader, R.; Koczan, D.; Stachs, O. Analysis of the Differential Gene and Protein Expression Profiles of Corneal Epithelial Cells Stimulated with Alternating Current Electric Fields. Genes 2021, 12, 299. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12020299
Kowtharapu BS, Damaraju J, Singh NK, Ziebart J, Bader R, Koczan D, Stachs O. Analysis of the Differential Gene and Protein Expression Profiles of Corneal Epithelial Cells Stimulated with Alternating Current Electric Fields. Genes. 2021; 12(2):299. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12020299
Chicago/Turabian StyleKowtharapu, Bhavani S., Jyoti Damaraju, Nitesh Kumar Singh, Josefin Ziebart, Rainer Bader, Dirk Koczan, and Oliver Stachs. 2021. "Analysis of the Differential Gene and Protein Expression Profiles of Corneal Epithelial Cells Stimulated with Alternating Current Electric Fields" Genes 12, no. 2: 299. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12020299