
Pleiotropic Roles of the Orthologue of the Drosophila melanogaster Intersex Gene in the Brown Planthopper



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Rearing
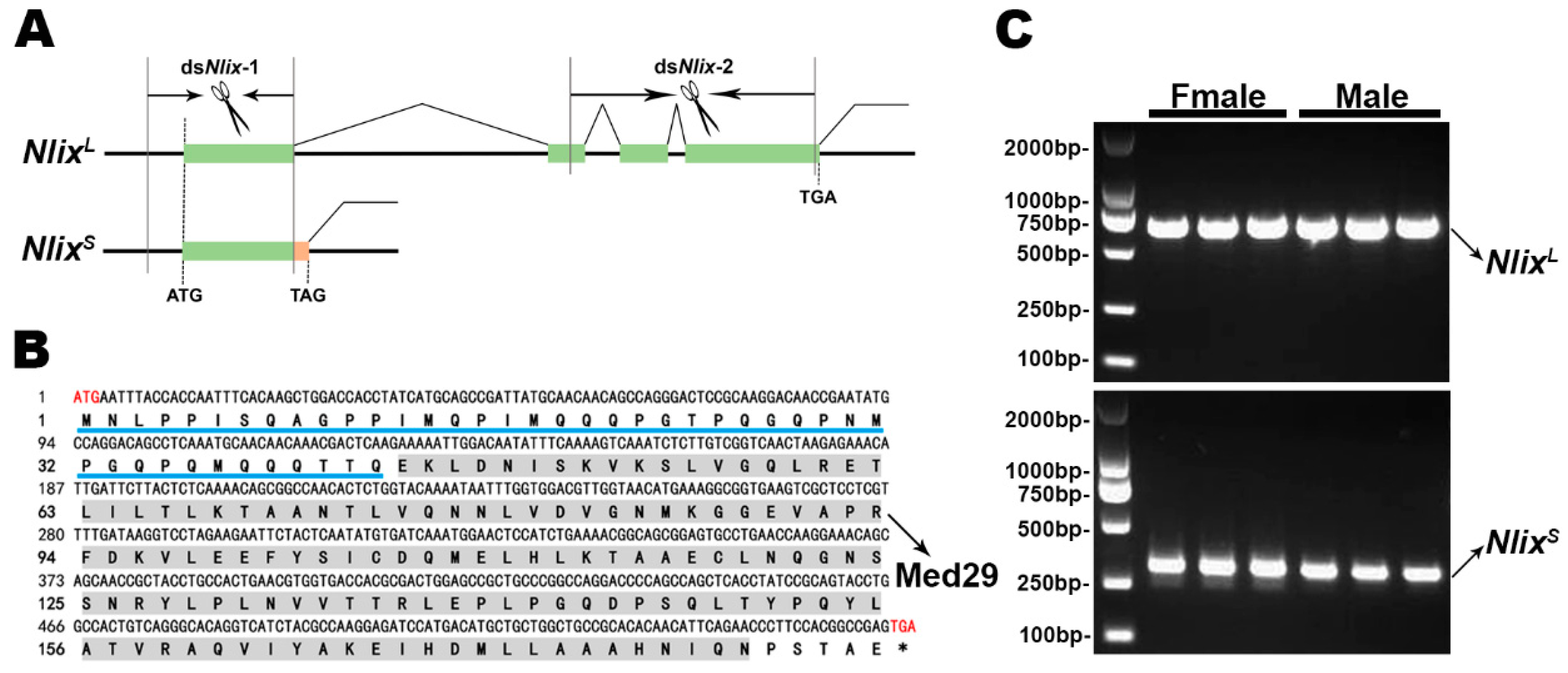
2.2. Searching for Ix Homologs in BPHs and Sequence Analysis
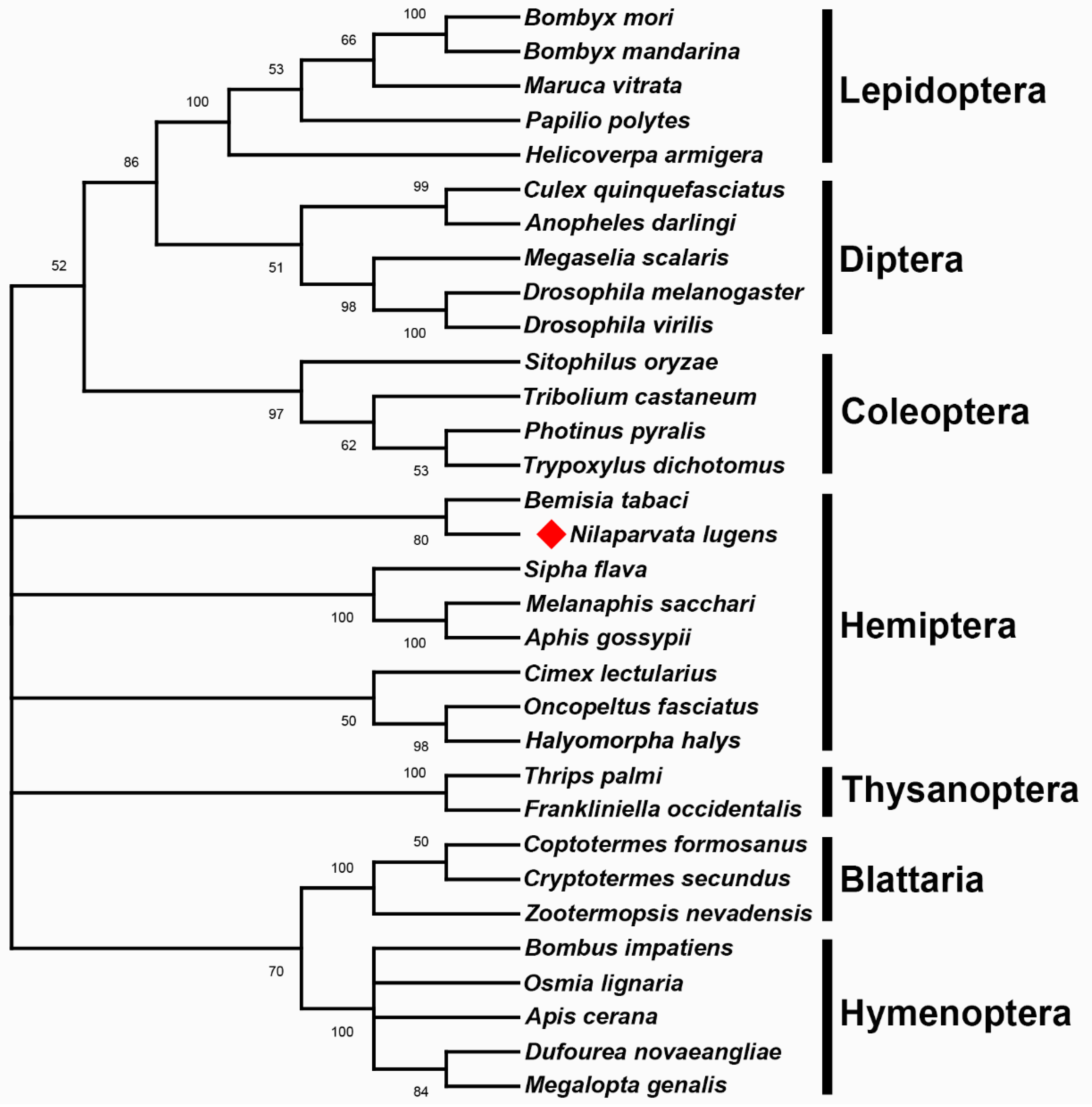
2.3. Sequence Comparison and Phylogenetic Relationships
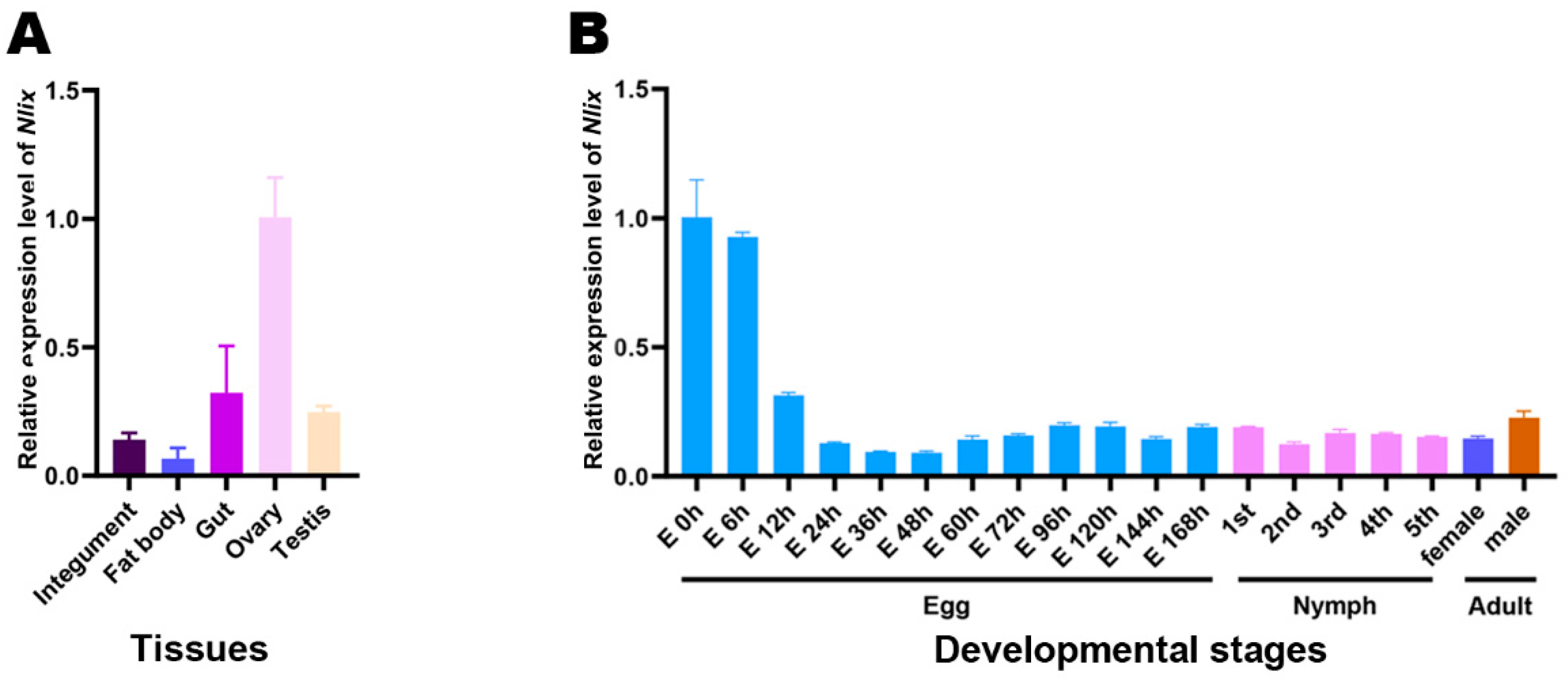
2.4. Expression Pattern Analysis of Nlix
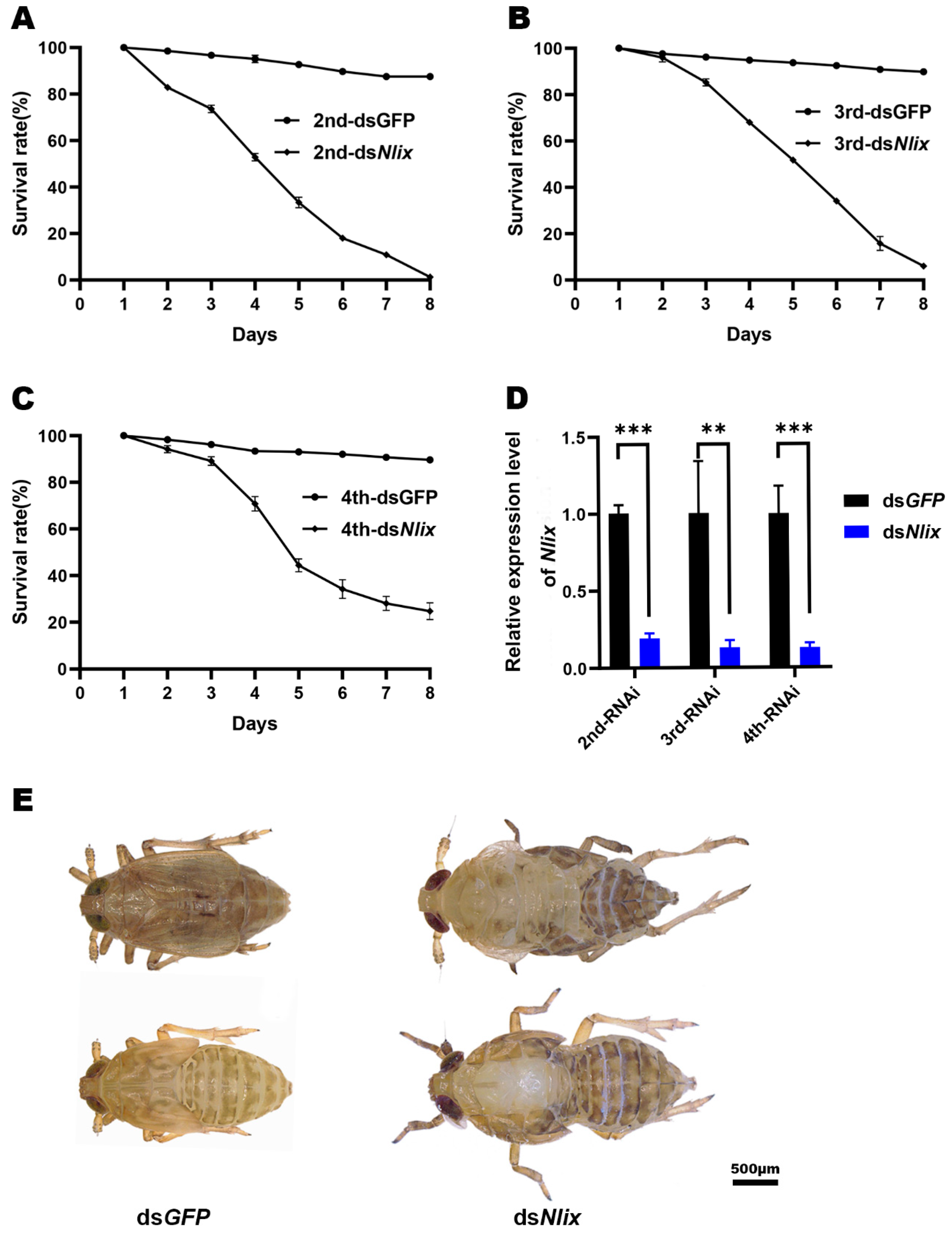
2.5. RNAi Interference
2.6. Fertility Analysis
2.7. Statistical Analysis
3. Results
3.1. Identification and Characterization of the Intersex Homologues in BPH
3.2. Phylogenetic Analysis
3.3. Temporal and Spatial Expression Patterns of Nlix
3.4. Phenotypes and RNAi Effects
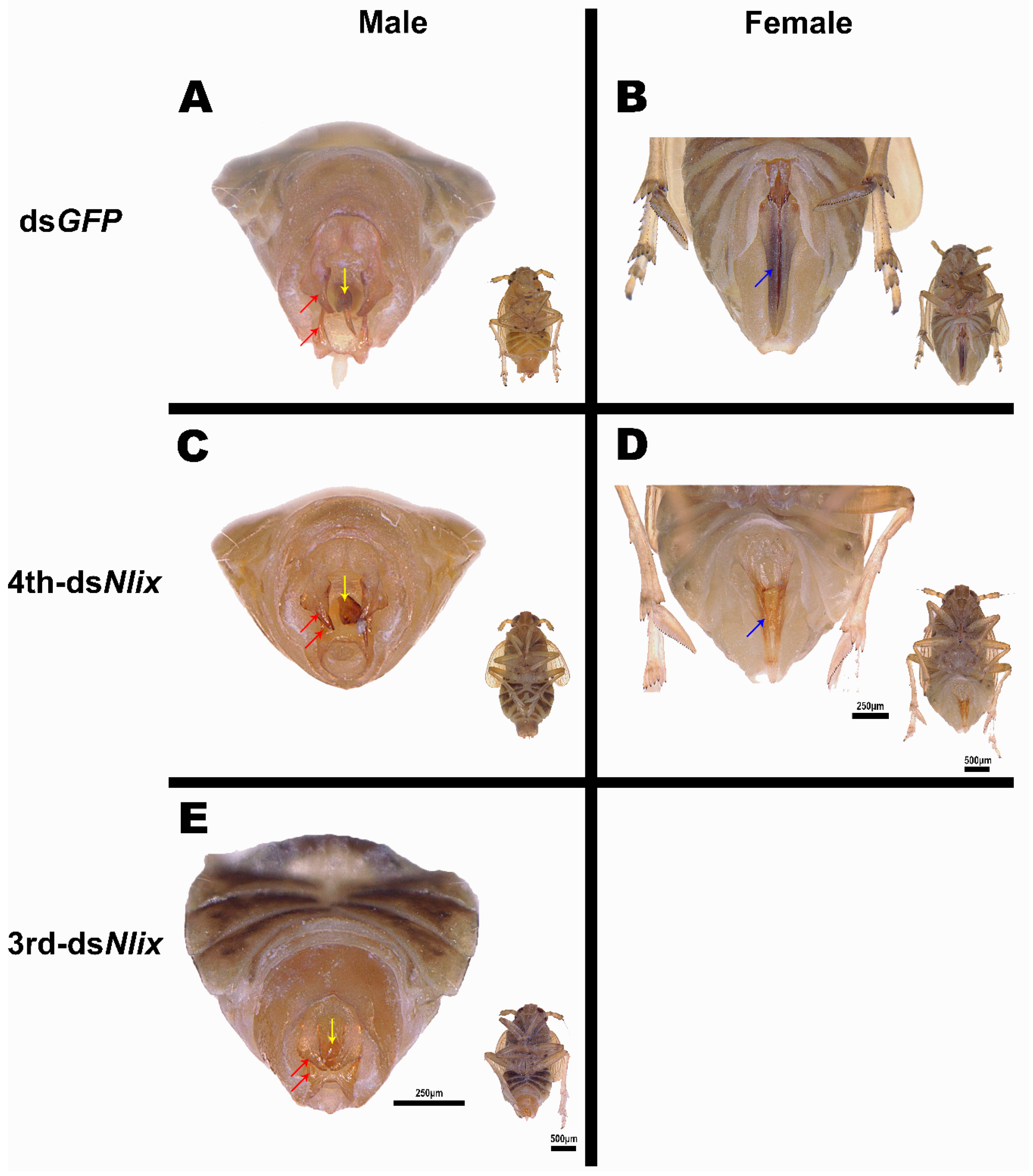
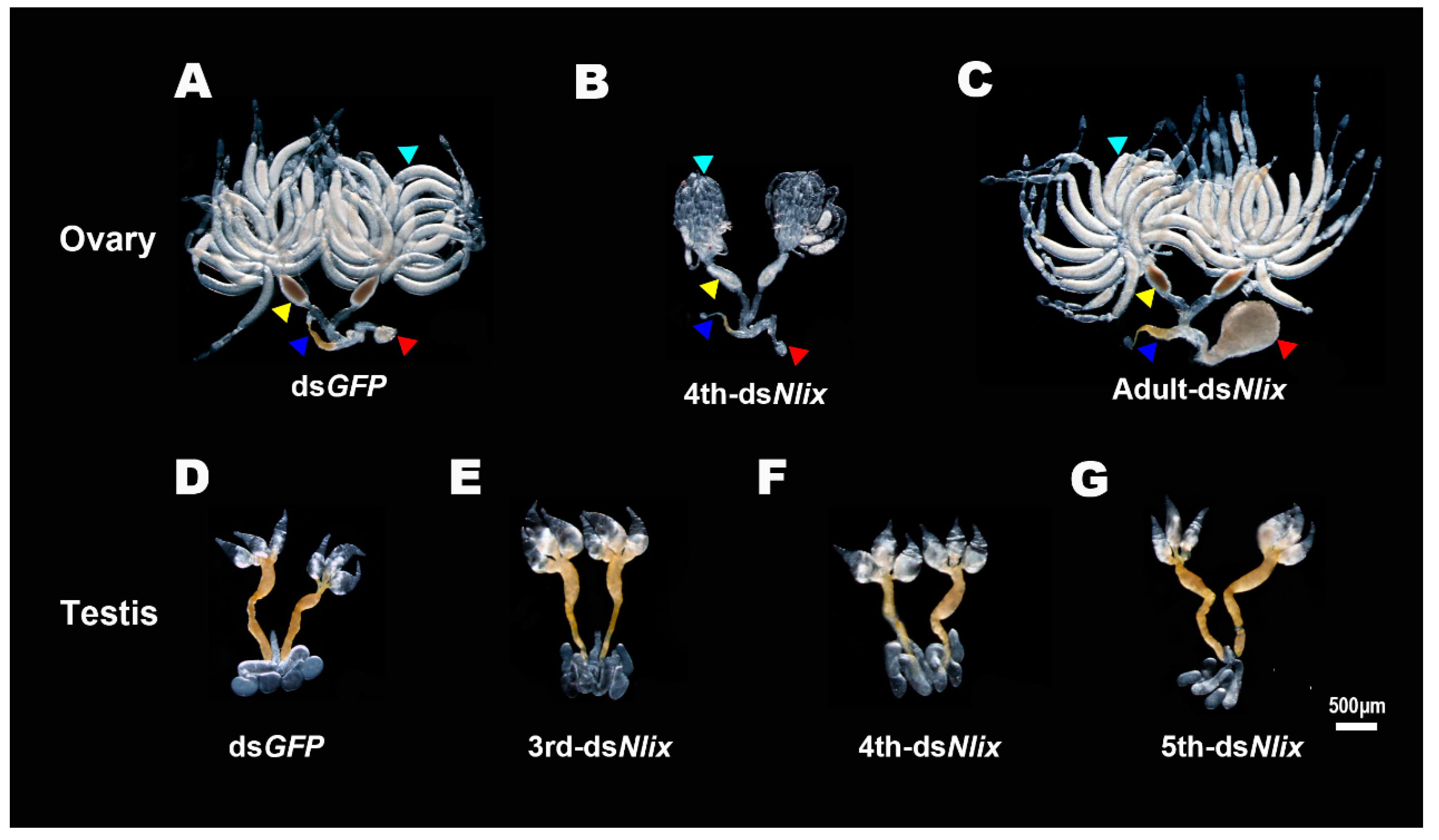
3.5. Nlix Influences Somatic Development of Females and Males
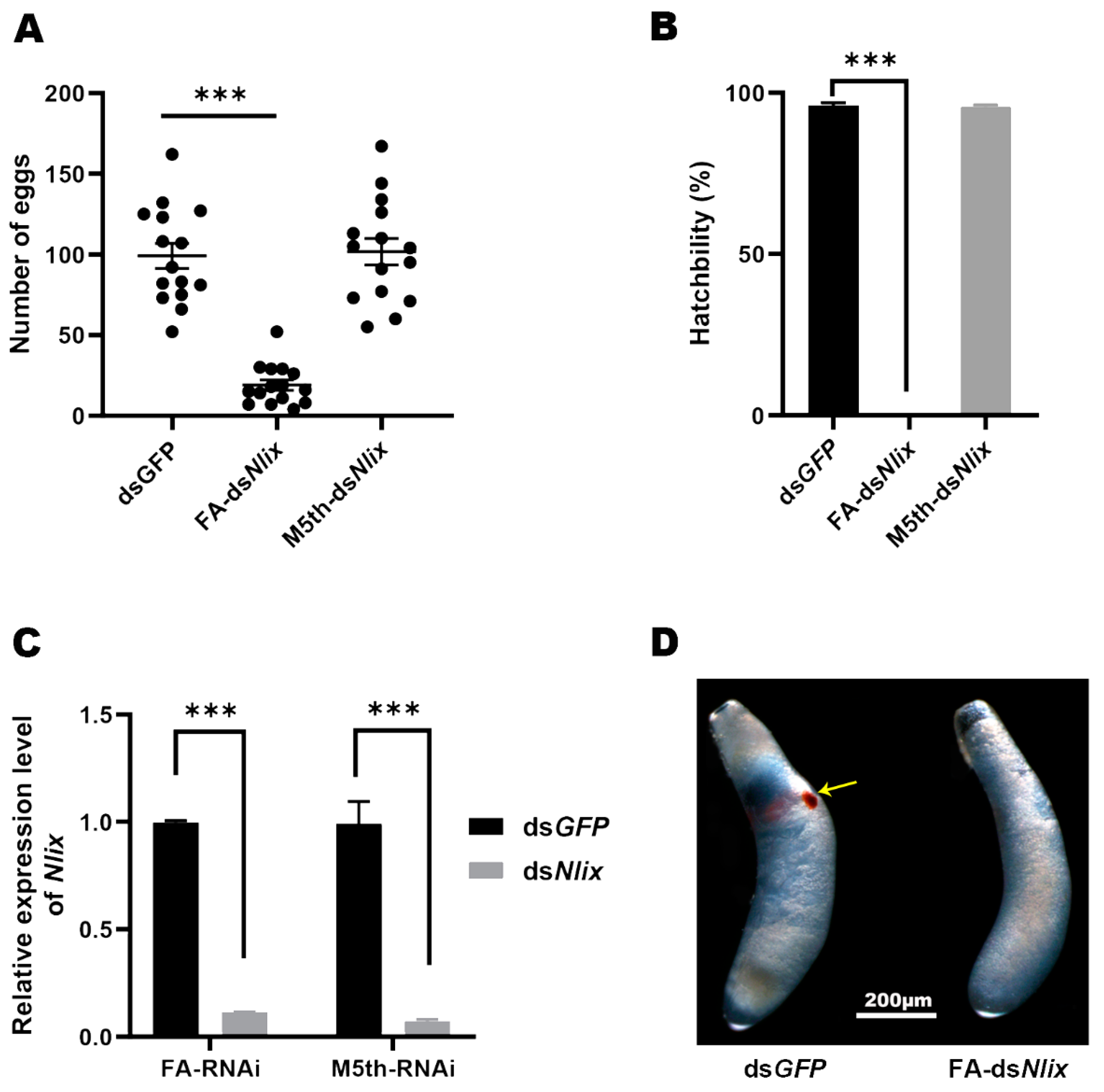
3.6. Effects of Nlix on BPH Fertility and Embryogenesis
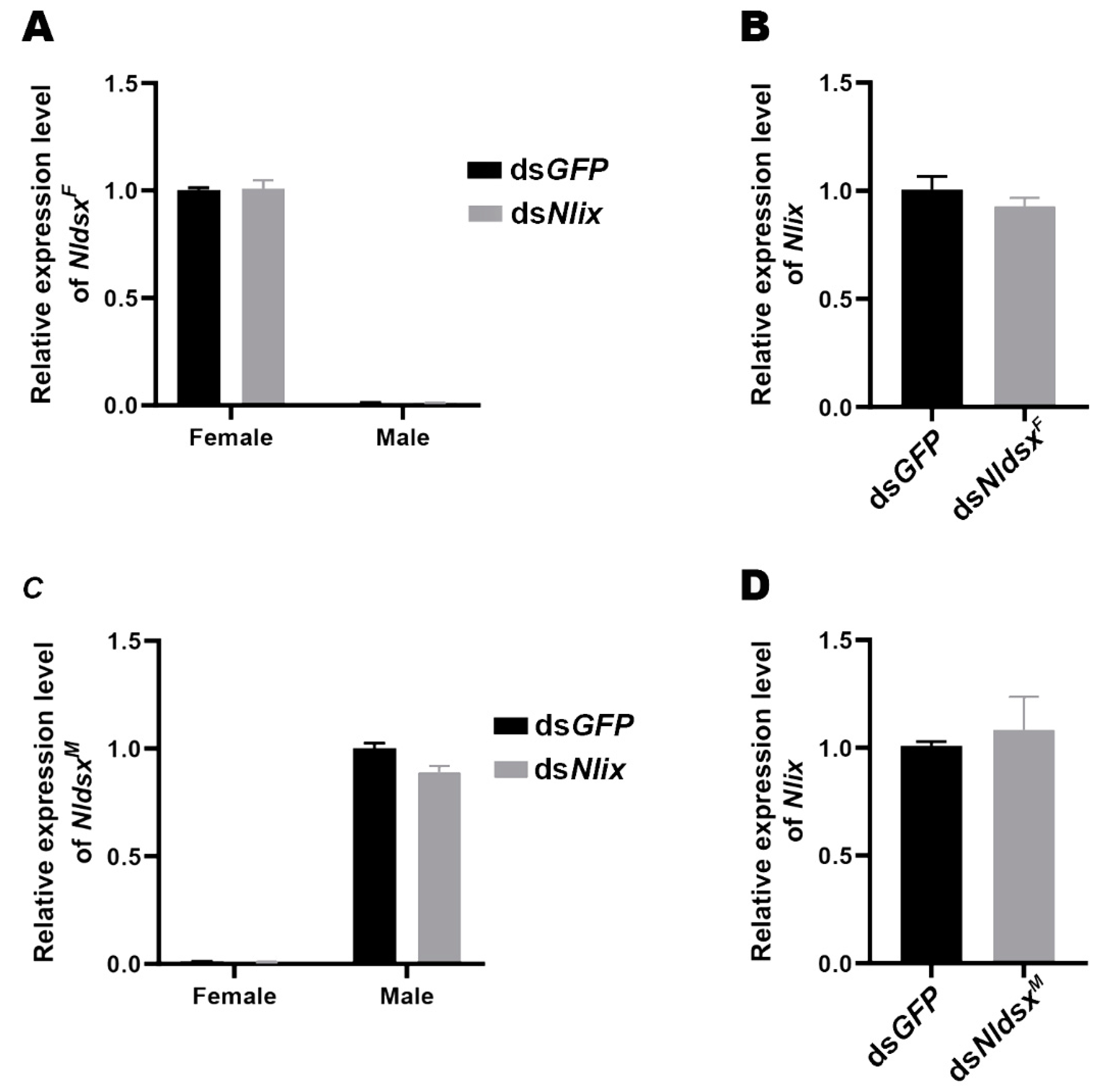
3.7. Nlix Functions Independently of Nldsx
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Garrett-Engele, C.M.; Siegal, M.L.; Manoli, D.S.; Williams, B.C.; Li, H.; Baker, B.S. Intersex, a gene required for female sexual development in Drosophila, is expressed in both sexes and functions together with doublesex to regulate terminal differentiation. Development 2002, 129, 4661–4675. [Google Scholar] [PubMed]
- Xu, J.; Yu, Y.; Chen, K.; Huang, Y. Intersex regulates female external genital and imaginal disc development in the silkworm. Insect. Biochem. Molec. 2019, 108, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Cavaliere, D.; Di Cara, F.; Polito, L.C.; Digilio, F.A. Cloning and functional characterization of the intersex homologous gene in the pest lepidopteron Maruca vitrata. Int. J. Dev. Biol. 2009, 53, 1057–1062. [Google Scholar] [CrossRef] [PubMed]
- Gotoh, H.; Zinna, R.A.; Warren, I.; Denieu, M.; Niimi, T.; Dworkin, I.; Emlen, D.J.; Miura, T.; Lavine, L.C. Identification and functional analyses of sex determination genes in the sexually dimorphic stag beetle Cyclommatus metallifer. BMC Genom. 2016, 17, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Yang, J.; Huo, Z.; Wang, S.; Wu, Q.; Zhou, X.; Xie, W.; Zhang, Y. Characteristic and functional study of intersex, a gene related to female fertility in Bemisia tabaci. Front. Physiol. 2020, 11, 55. [Google Scholar] [CrossRef] [PubMed]
- Aspiras, A.C.; Smith, F.W.; Angelini, D.R. Sex-specific gene interactions in the patterning of insect genitalia. Dev. Biol. 2011, 360, 369–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegal, M.L.; Baker, B.S. Functional conservation and divergence of intersex, a gene required for female differentiation in Drosophila melanogaster. Dev. Genes Evol. 2005, 215, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, R.N.; Chatterjee, P.; Kuthe, S.; Acharyya-Ari, M.; Chatterjee, R. Intersex (ix) mutations of Drosophila melanogaster cause nonrandom cell death in genital disc and can induce tumours in genitals in response to decapentaplegic (dpp(disk)) mutations. J. Genet. 2015, 94, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Chase, B.A.; Baker, B.S. A genetic analysis of intersex, a gene regulating sexual differentiation in Drosophila melanogaster females. Genetics 1995, 139, 1649–1661. [Google Scholar] [CrossRef] [PubMed]
- Bottrell, D.G.; Schoenly, K.G. Resurrecting the ghost of green revolutions past: The brown planthopper as a recurring threat to high-yielding rice production in tropical Asia. J. Asia Pac. Entomol. 2012, 15, 122–140. [Google Scholar] [CrossRef]
- Zhuo, J.; Lei, C.; Shi, J.; Xu, N.; Xue, W.; Zhang, M.; Ren, Z.; Zhang, H.; Zhang, C. Tra-2 mediates cross-talk between sex determination and wing polyphenism in female Nilaparvata lugens. Genetics 2017, 207, 1067–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burtis, K.C.; Baker, B.S. Drosophila doublesex gene controls somatic sexual differentiation by producing alternatively spliced mRNAs encoding related sex-specific polypeptides. Cell 1989, 56, 997–1010. [Google Scholar] [CrossRef]
- Zhuo, J.; Hu, Q.; Zhang, H.; Zhang, M.; Jo, S.B.; Zhang, C. Identification and functional analysis of the doublesex gene in the sexual development of a hemimetabolous insect, the brown planthopper. Insect. Biochem. Molec. 2018, 102, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using Real-Time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Xue, J.; Lu, B.; Zhang, X.; Zhuo, J.; He, S.; Ma, X.; Jiang, Y.; Fan, H.; Xu, J.; et al. Two insulin receptors determine alternative wing morphs in planthoppers. Nature 2015, 519, 464–467. [Google Scholar] [CrossRef] [PubMed]
- Pan, P.; Ye, Y.; Lou, Y.; Lu, J.; Cheng, C.; Shen, Y.; Moussian, B.; Zhang, C. A comprehensive omics analysis and functional survey of cuticular proteins in the brown planthopper. Proc. Nat. Acad. Sci. USA 2018, 115, 5175–5180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Zhang, W.; Bayrer, J.R.; Weiss, M.A. Doublesex and the regulation of sexual dimorphism in Drosophila melanogaster. J. Biol. Chem. 2008, 283, 7280–7292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, S.; Tomomori-Sato, C.; Banks, C.A.S.; Parmely, T.J.; Sorokina, I.; Brower, C.S.; Conaway, R.C.; Conaway, J.W. A mammalian homolog of Drosophila melanogaster transcriptional coactivator Intersex is a subunit of the mammalian Mediator complex. J. Biol. Chem. 2003, 278, 49671–49674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, S.; Tomomori-Sato, C.; Parmely, T.J.; Florens, L.; Zybailov, B.; Swanson, S.K.; Banks, C.A.S.; Jin, J.; Cai, Y.; Washburn, M.P.; et al. A set of consensus mammalian mediator subunits identified by multidimensional protein identification technology. Mol. Cell 2004, 14, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Arunkumar, K.P.; Nagaraju, J. Drosophila intersex orthologue in the silkworm, Bombyx mori and related species. Genetica 2011, 139, 141–147. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.-H.; Xie, Y.-C.; Li, H.-J.; Zhuo, J.-C.; Zhang, C.-X. Pleiotropic Roles of the Orthologue of the Drosophila melanogaster Intersex Gene in the Brown Planthopper. Genes 2021, 12, 379. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030379
Zhang H-H, Xie Y-C, Li H-J, Zhuo J-C, Zhang C-X. Pleiotropic Roles of the Orthologue of the Drosophila melanogaster Intersex Gene in the Brown Planthopper. Genes. 2021; 12(3):379. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030379
Chicago/Turabian StyleZhang, Hou-Hong, Yu-Cheng Xie, Han-Jing Li, Ji-Chong Zhuo, and Chuan-Xi Zhang. 2021. "Pleiotropic Roles of the Orthologue of the Drosophila melanogaster Intersex Gene in the Brown Planthopper" Genes 12, no. 3: 379. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030379