
Analysis of MCM Proteins’ Role as a Potential Target of Statins in Patients with Acute Type A Aortic Dissection through Bioinformatics

Abstract
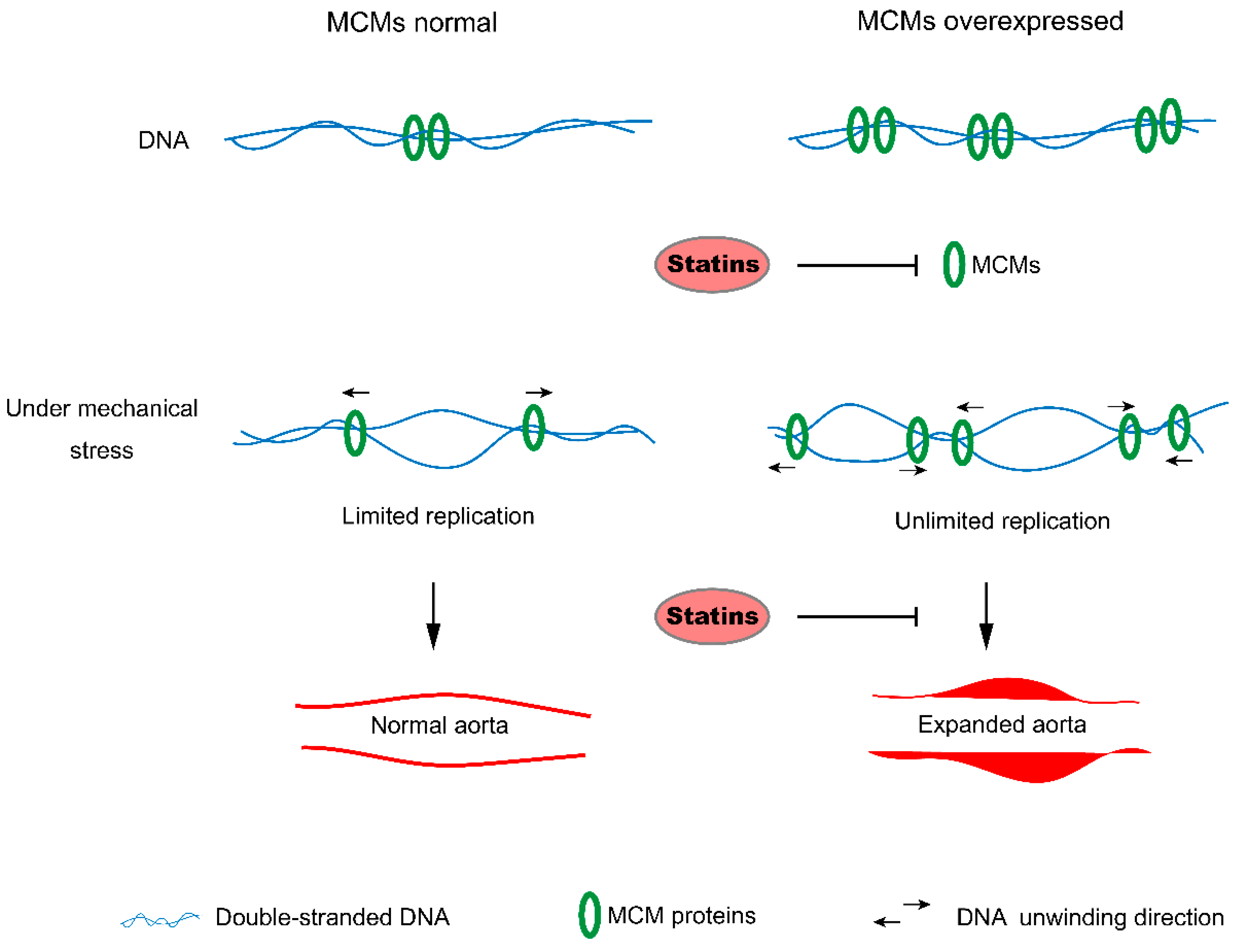
:1. Introduction
2. Materials and Methods
2.1. Data Processing
2.2. Identification of Differentially Expressed Genes (DEGs)
2.3. Weight Gene Correlation Network Analysis (WGCNA)
2.4. Analysis of Regional Variations of Genomic Features
2.5. Immune-Cell Enrichment Analysis
3. Results
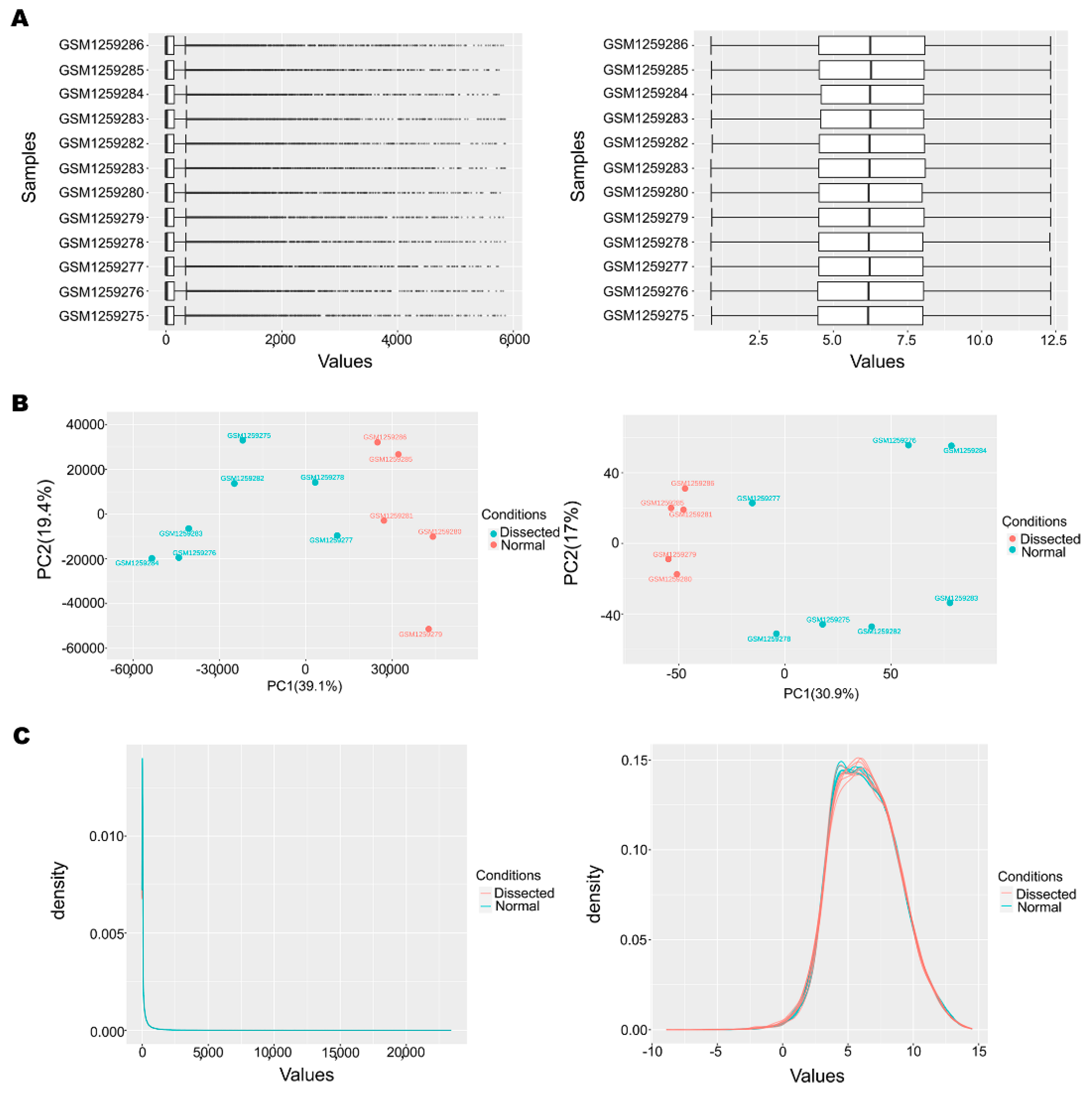
3.1. Data Preprocessing and Normalization
3.2. Identification of DEGs and the Expression of MCM Proteins
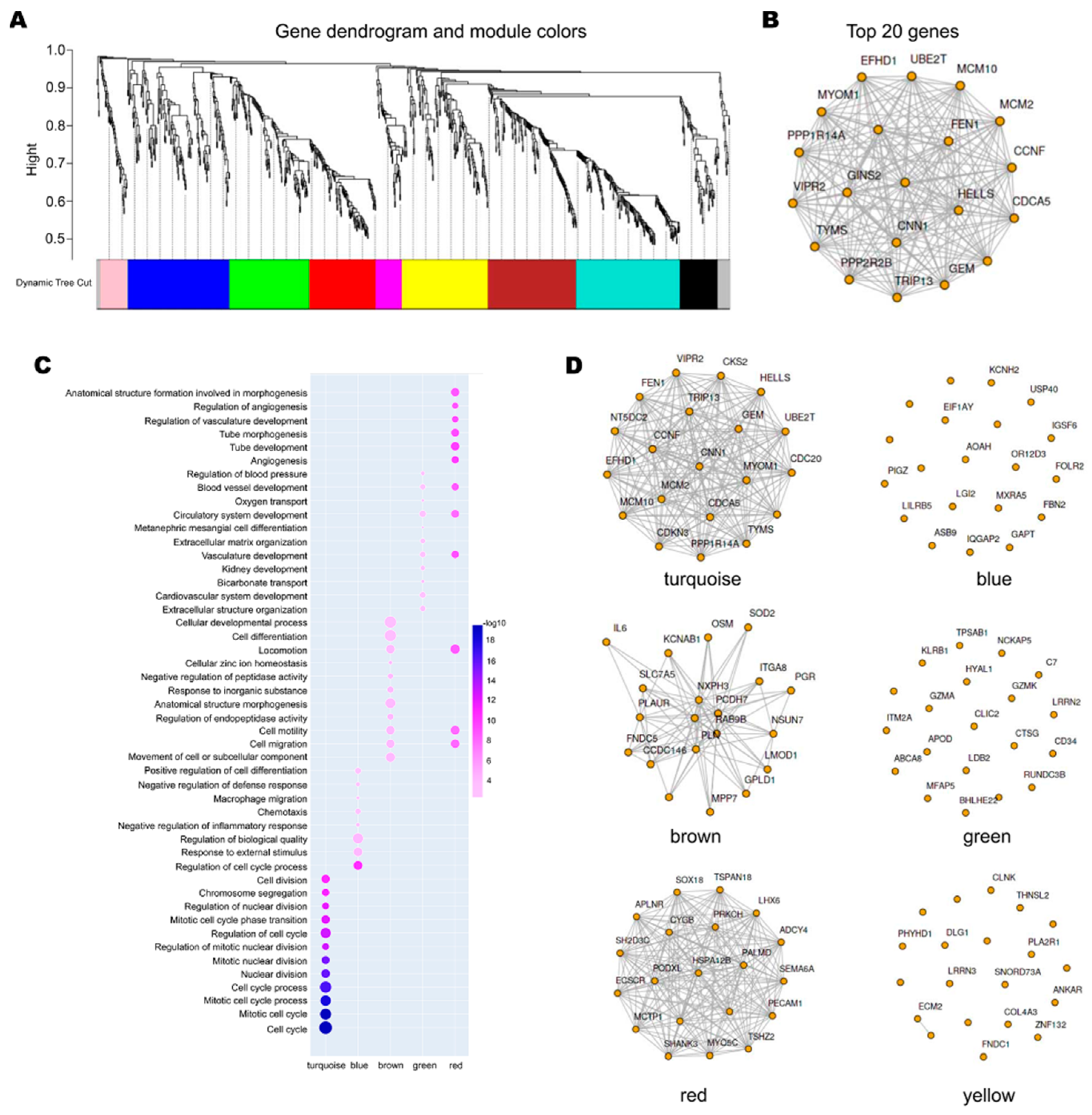
3.3. The Network of DEGs and the Correlation of the Genes with MCM Proteins
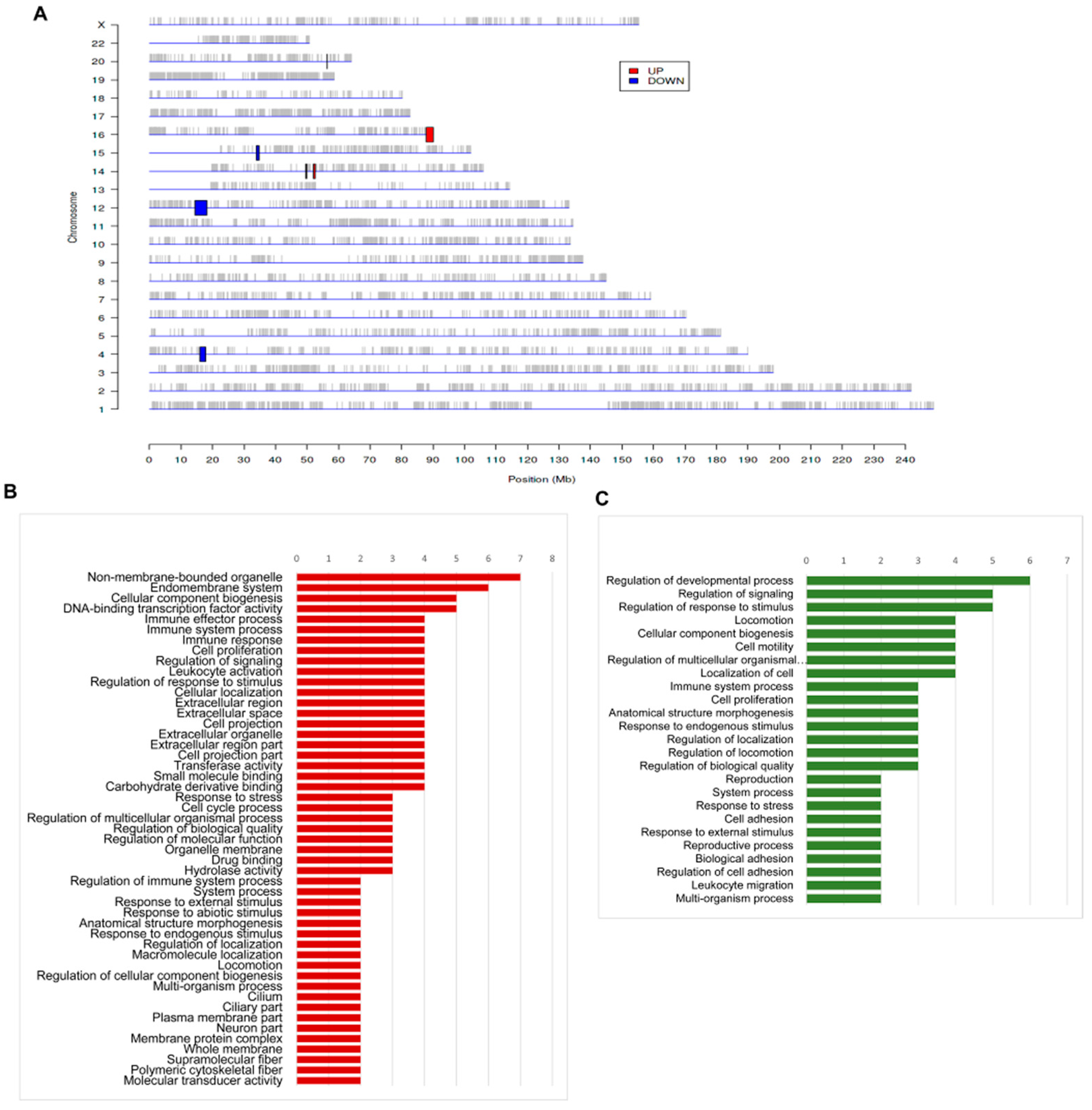
3.4. The Regional Variations of Genomic Features of Aortic Dissection
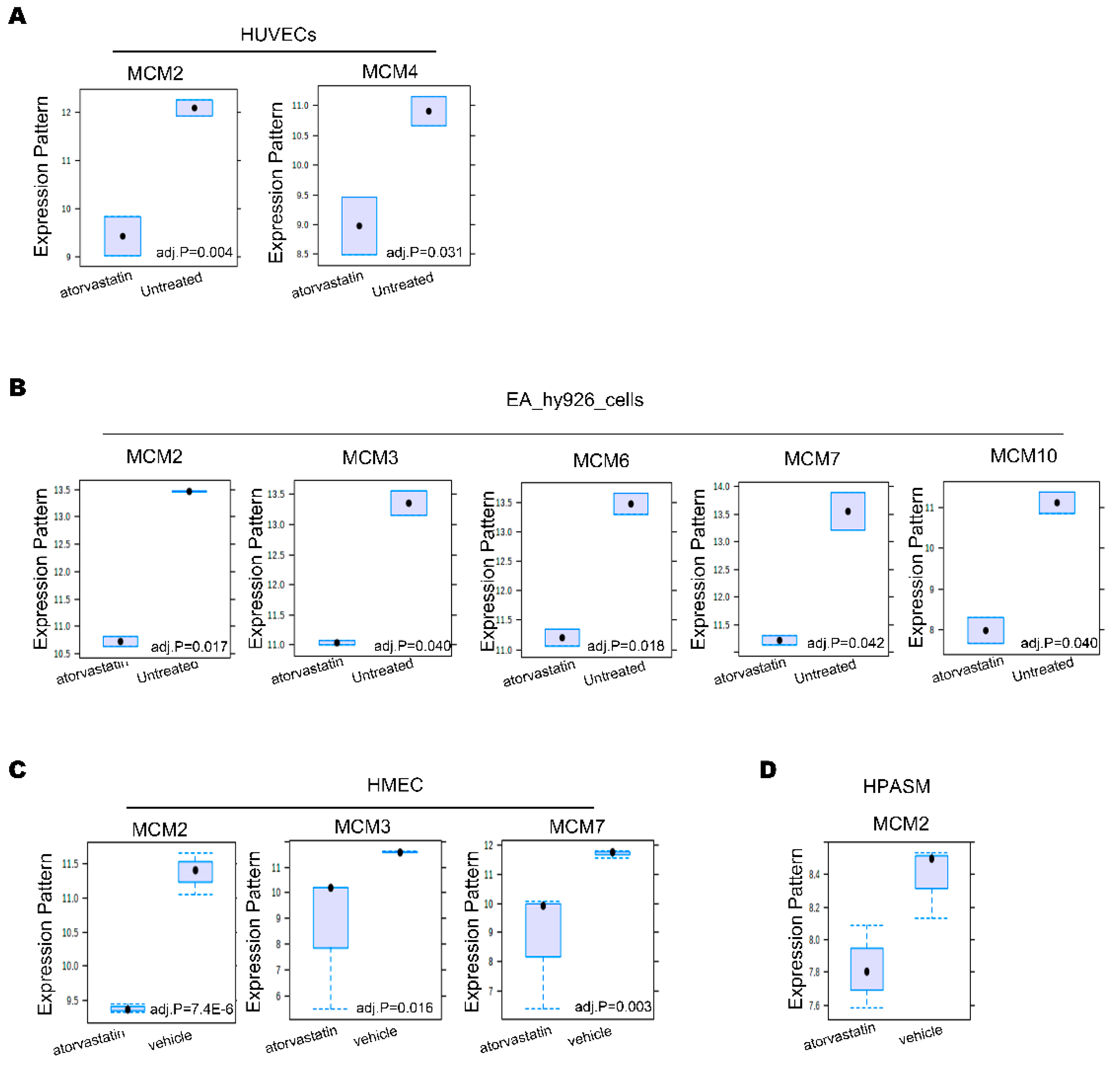
3.5. Statins Inhibit MCM Proteins Expression in Vascular Cell
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vilacosta, I.; Aragoncillo, P.; Canadas, V.; Roman, J.A.S.; Ferreiros, J.; Rodriguez, E. Acute aortic syndrome: A new look at an old conundrum. Postgrad. Med. J. 2010, 86, 52–61. [Google Scholar] [CrossRef]
- White, A.; Broder, J.; Mando-Vandrick, J.; Wendell, J.; Crowe, J. Acute Aortic Emergencies—Part 2 Aortic Dissections. Adv. Emerg. Nurs. J. 2013, 35, 28–52. [Google Scholar] [CrossRef] [PubMed]
- Gawinecka, J.; Schönrath, F.; Von Eckardstein, A. Acute aortic dissection: Pathogenesis, risk factors and diagnosis. Swiss Med. Wkly. 2017, 147, w14489. [Google Scholar] [CrossRef]
- Suzuki, T.; Isselbacher, E.M.; Nienaber, C.A.; Pyeritz, R.E.; Eagle, K.A.; Tsai, T.T.; Cooper, J.V.; Januzzi, J.L.; Braverman, A.C.; Montgomery, D.G.; et al. Type-Selective Benefits of Medications in Treatment of Acute Aortic Dissection (from the International Registry of Acute Aortic Dissection [IRAD]). Am. J. Cardiol. 2012, 109, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Durham, C.A.; Cambria, R.P.; Wang, L.J.; Ergul, E.A.; Aranson, N.J.; Patel, V.I.; Conrad, M.F. The natural history of medically managed acute type B aortic dissection. J. Vasc. Surg. 2015, 61, 1192–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akutsu, K.; Nejima, J.; Kiuchi, K.; Sasaki, K.; Ochi, M.; Tanaka, K.; Takano, T. Effects of the patent false lumen on the long-term outcome of type B acute aortic dissection. Eur. J. Cardio-Thoracic Surg. 2004, 26, 359–366. [Google Scholar] [CrossRef]
- Kalyanasundaram, A.; Elmore, J.R.; Manazer, J.R.; Golden, A.; Franklin, D.P.; Galt, S.W.; Zakhary, E.M.; Carey, D.J. Simvastatin suppresses experimental aortic aneurysm expansion. J. Vasc. Surg. 2006, 43, 117.e39. [Google Scholar] [CrossRef] [Green Version]
- Steinmetz, E.F.; Buckley, C.; Shames, M.L.; Ennis, T.L.; Vanvickle-Chavez, S.J.; Mao, D.; Goeddel, L.A.; Hawkins, C.J.; Thompson, R.W. Treatment with Simvastatin Suppresses the Development of Experimental Abdominal Aortic Aneurysms in Normal and Hypercholesterolemic Mice. Ann. Surg. 2005, 241, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Takagi, H.; Matsui, M.; Umemoto, T. A meta-analysis of clinical studies of statins for prevention of abdominal aortic aneurysm expansion. J. Vasc. Surg. 2010, 52, 1675–1681. [Google Scholar] [CrossRef] [Green Version]
- Raux, M.; Cochennec, F.; Becquemin, J.-P. Statin therapy is associated with aneurysm sac regression after endovascular aortic repair. J. Vasc. Surg. 2012, 55, 1587–1592. [Google Scholar] [CrossRef] [Green Version]
- Sukhija, R.; Aronow, W.S.; Sandhu, R.; Kakar, P.; Babu, S. Mortality and Size of Abdominal Aortic Aneurysm at Long-Term Follow-Up of Patients Not Treated Surgically and Treated with and Without Statins. Am. J. Cardiol. 2006, 97, 279–280. [Google Scholar] [CrossRef]
- Liang, Z.; Li, W.; Liu, J.; Li, J.; He, F.; Jiang, Y.; Yang, L.; Li, P.; Wang, B.; Wang, Y.; et al. Simvastatin suppresses the DNA replication licensing factor MCM7 and inhibits the growth of tamoxifen-resistant breast cancer cells. Sci. Rep. 2017, 7, 41776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Liu, J.; Liang, Z.; He, F.; Yang, L.; Li, P.; Jiang, Y.; Wang, B.; Zhou, C.; Wang, Y.; et al. Simvastatin and Atorvastatin inhibit DNA replication licensing factor MCM7 and effectively suppress RB-deficient tumors growth. Cell Death Dis. 2017, 8, e2673. [Google Scholar] [CrossRef] [Green Version]
- Tye, B.K. MCM Proteins in DNA Replication. Annu. Rev. Biochem. 1999, 68, 649–686. [Google Scholar] [CrossRef] [PubMed]
- Maiorano, D.; Lutzmann, M.; Méchali, M. MCM proteins and DNA replication. Curr. Opin. Cell Biol. 2006, 18, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Enemark, E.J.; Joshua-Tor, L. On helicases and other motor proteins. Curr. Opin. Struct. Biol. 2008, 18, 243–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibarra, A.; Schwob, E.; Méndez, J. Excess MCM proteins protect human cells from replicative stress by licensing backup origins of replication. Proc. Natl. Acad. Sci. USA 2008, 105, 8956–8961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for functional genomics data sets—Update. Nucleic Acids Res. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.; Soufan, O.; Ewald, J.; Hancock, R.E.W.; Basu, N.; Xia, J. NetworkAnalyst 3.0: A visual analytics platform for comprehensive gene expression profiling and meta-analysis. Nucleic Acids Res. 2019, 47, W234–W241. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Benner, M.J.; Hancock, R.E.W. NetworkAnalyst—Integrative approaches for protein–protein interaction network analysis and visual exploration. Nucleic Acids Res. 2014, 42, W167–W174. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; E Gill, E.; Hancock, R.E.W. NetworkAnalyst for statistical, visual and network-based meta-analysis of gene expression data. Nat. Protoc. 2015, 10, 823–844. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.X.; Son, E.W.; Yao, R. iDEP: An integrated web application for differential expression and pathway analysis of RNA-Seq data. BMC Bioinform. 2018, 19, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrari, F.; Solari, A.; Battaglia, C.; Bicciato, S. PREDA: An R-package to identify regional variations in genomic data. Bioinformatics 2011, 27, 2446–2447. [Google Scholar] [CrossRef] [Green Version]
- Aran, D.; Hu, Z.; Butte, A.J. xCell: Digitally portraying the tissue cellular heterogeneity landscape. Genome Biol. 2017, 18. [Google Scholar] [CrossRef] [Green Version]
- Nienaber, C.A.; Clough, R.E.; Sakalihasan, N.; Suzuki, T.; Gibbs, R.; Mussa, F.; Jenkins, M.P.; Thompson, M.M.; Evangelista, A.; Yeh, J.S.M.; et al. Aortic dissection. Nat. Rev. Dis. Prim. 2016, 2, 1–18. [Google Scholar] [CrossRef]
- Members, T.F.; Erbel, R.; Aboyans, V.; Boileau, C.; Bossone, E.; Di Bartolomeo, R.; Eggebrecht, H.; Evangelista, A.; Falk, V.; Frank, H.; et al. 2014 ESC Guidelines on the diagnosis and treatment of aortic diseases. Eur. Hear. J. 2014, 35, 2873–2926. [Google Scholar] [CrossRef] [Green Version]
- Jiang, T.; Si, L. Identification of the molecular mechanisms associated with acute type A aortic dissection through bioinformatics methods. Braz. J. Med Biol. Res. 2019, 52, e8950. [Google Scholar] [CrossRef]
- Todorov, I.T.; Attaran, A.; E Kearsey, S. BM28, a human member of the MCM2-3-5 family, is displaced from chromatin during DNA replication. J. Cell Biol. 1995, 129, 1433–1445. [Google Scholar] [CrossRef]
- Young, M.R.; Tye, B.K. Mcm2 and Mcm3 are constitutive nuclear proteins that exhibit distinct isoforms and bind chromatin during specific cell cycle stages of Saccharomyces cerevisiae. Mol. Biol. Cell 1997, 8, 1587–1601. [Google Scholar] [CrossRef] [Green Version]
- Krude, T.; Musahl, C.; A Laskey, R.; Knippers, R. Human replication proteins hCdc21, hCdc46 and P1Mcm3 bind chromatin uniformly before S-phase and are displaced locally during DNA replication. J. Cell Sci. 1996, 109, 309–318. [Google Scholar]
- Aparicio, O.M.; Weinstein, D.M.; Bell, S.P. Components and Dynamics of DNA Replication Complexes in S. cerevisiae: Redistribution of MCM Proteins and Cdc45p during S Phase. Cell 1997, 91, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Torrens, C.; Kelsall, C.J.; Hopkins, L.A.; Anthony, F.W.; Curzen, N.P.; Hanson, M.A. Atorvastatin Restores Endothelial Function in Offspring of Protein-Restricted Rats in a Cholesterol-Independent Manner. Hypertension 2009, 53, 661–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, L.H.; Berger, J.; Tranquilli, M.; Elefteraides, J.A. Effect of Statin Drugs on Thoracic Aortic Aneurysms. Am. J. Cardiol. 2013, 112, 1240–1245. [Google Scholar] [CrossRef] [PubMed]
- Tazaki, J.; Morimoto, T.; Sakata, R.; Okabayashi, H.; Yamazaki, F.; Nishiwaki, N.; Mitsudo, K.; Kimura, T. Impact of statin therapy on patients with coronary heart disease and aortic aneurysm or dissection. J. Vasc. Surg. 2014, 60, 604–612.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masaki, N.; For the STANP trial investigators; Kumagai, K.; Sasaki, K.; Matsuo, S.; Motoyoshi, N.; Adachi, O.; Akiyama, M.; Kawamoto, S.; Tabayashi, K.; et al. Suppressive effect of pitavastatin on aortic arch dilatation in acute stanford type B aortic dissection: Analysis of STANP trial. Gen. Thorac. Cardiovasc. Surg. 2018, 66, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Kohno, M.; Shinomiya, K.; Abe, S.; Noma, T.; Kondo, I.; Oshita, A.; Takeuchi, H.; Takagi, Y.; Yukiiri, K.; Mizushige, K.; et al. Inhibition of Migration and Proliferation of Rat Vascular Smooth Muscle Cells by a New HMG-CoA Reductase Inhibitor, Pitavastatin. Hypertens. Res. 2002, 25, 279–285. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | logFC | adj-P-Val |
|---|---|---|
| MCM2 | 2.62753 | 0.00817 |
| MCM3 | 0.66044 | 0.08468 |
| MCM4 | 2.37032 | 0.00656 |
| MCM5 | 1.52354 | 0.03502 |
| MCM6 | 0.86740 | 0.01988 |
| MCM7 | 0.78220 | 0.06388 |
| MCM10 | 3.21558 | 0.00684 |
| Regulation | Chr. | Start | End | Size | Band | Genes |
|---|---|---|---|---|---|---|
| Up | 14 | 49622140 | 49895068 | 0.273 | q21 | POLE2, KLHDC1, ARF6 |
| Up | 14 | 52037015 | 52595274 | 0.558 | q22 | PTGER2, TXNDC16, GPR137C |
| Up | 16 | 87849765 | 90074319 | 2.225 | q24 | ZNF469, ZFPM1, CYBA, MVD, RNF166, CTU2, APRT, GALNS, TRAPPC2L, ZNF778, SNORD68; CHMP1A, CDK10, SPATA2L, ZNF276, MC1R, TUBB3, CENPBD1, DBNDD1, GAS8, PRDM7 |
| Down | 4 | 16023727 | 17805706 | 1.782 | p15 | TAPT1, LDB2, QDPR, LAP3, MED28, DCAF16 |
| Down | 12 | 14434283 | 18200488 | 3.766 | p12 | C12orf60, ART4, MGP, ERP27, ARHGDIB, RERG, PTPRO, EPS8, STRAP, MGST1, LMO3, RERGL |
| Down | 15 | 33952715 | 34792889 | 0.84 | q14 | PGBD4, KATNBL1, SLC12A6, NOP10, LPCAT4, GOLGA8A, GOLGA8B, ACTC1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, Z.; Zhang, Y.; Chen, Q.; Hao, J.; Wang, H.; Li, Y.; Yan, Y. Analysis of MCM Proteins’ Role as a Potential Target of Statins in Patients with Acute Type A Aortic Dissection through Bioinformatics. Genes 2021, 12, 387. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030387
Liang Z, Zhang Y, Chen Q, Hao J, Wang H, Li Y, Yan Y. Analysis of MCM Proteins’ Role as a Potential Target of Statins in Patients with Acute Type A Aortic Dissection through Bioinformatics. Genes. 2021; 12(3):387. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030387
Chicago/Turabian StyleLiang, Zheyong, Yongjian Zhang, Qiang Chen, Junjun Hao, Haichen Wang, Yongxin Li, and Yang Yan. 2021. "Analysis of MCM Proteins’ Role as a Potential Target of Statins in Patients with Acute Type A Aortic Dissection through Bioinformatics" Genes 12, no. 3: 387. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030387