
Newcastle Disease Virus Induced Pathologies Severely Affect the Exocrine and Endocrine Functions of the Pancreas in Chickens

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus
2.2. Chickens, Ethics Statement, and Experimental Design
2.3. Sampling
2.4. Plasma Hormones
2.5. Digestive Enzyme Activities in the Pancreas
2.6. Quantitative Real-Time PCR
2.7. Viral Loads in the Pancreas
2.8. Histopathology
2.9. TUNEL Assay
2.10. Statistical Analysis
3. Results
3.1. Viral Load in the Pancreas
3.2. Histopathology
3.3. Apoptosis in the Pancreas
3.4. Plasma Concentrations of Hormones
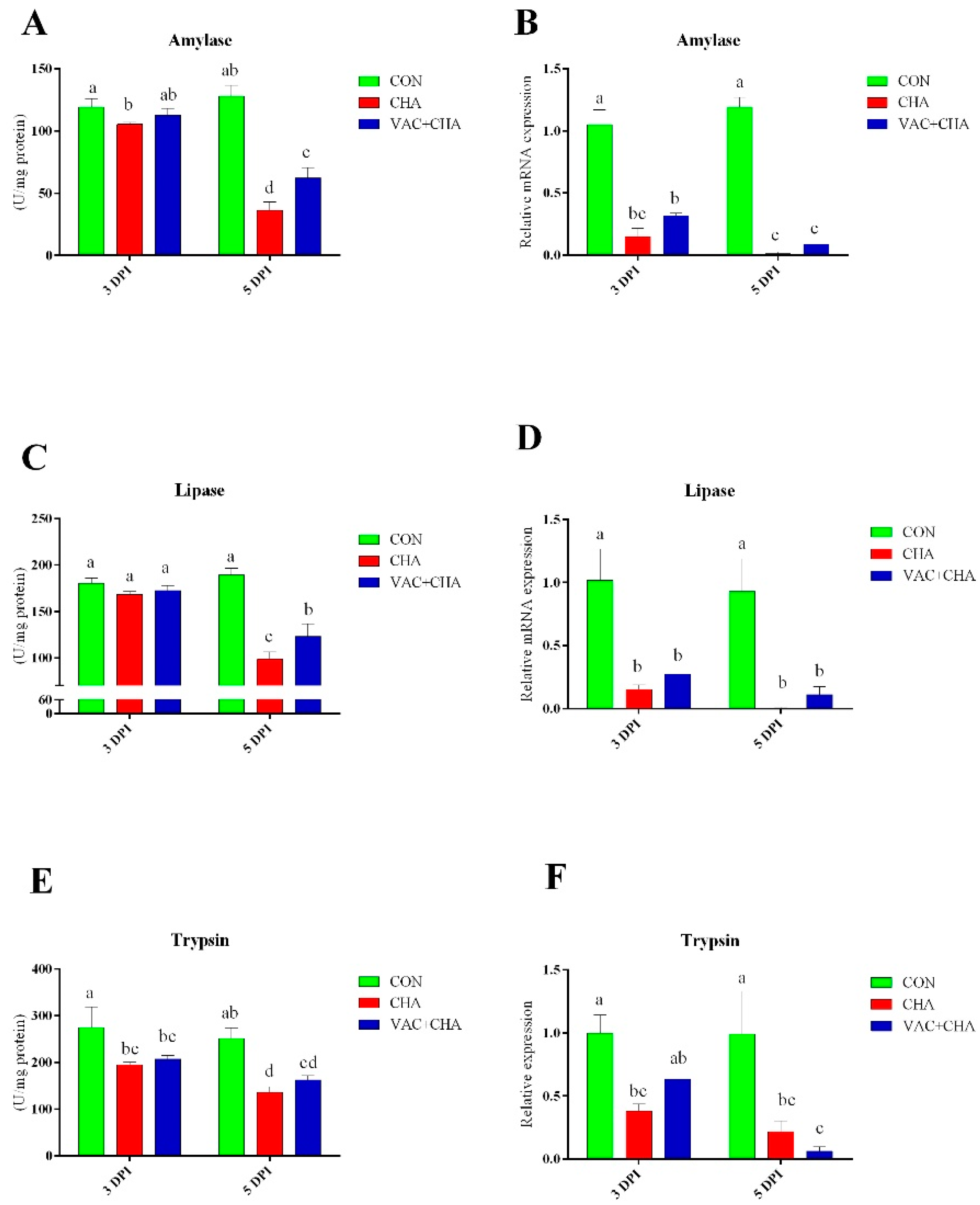
3.5. Activity and Expression of Pancreatic Enzymes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alexander, D.J. Newcastle disease. In Disease of Poultry; Saif, Y.M., Barnes, H.J., Glisson, J.R., Fadly, A.M., McDougald, L.R., Swayne, D.E., Eds.; Iowa State Press: Ames, IA, USA, 2003; pp. 64–87. [Google Scholar]
- Rehman, Z.U.; Meng, C.; Sun, Y.; Mahrose, K.M.; Umar, S.; Ding, C.; Munir, M. Pathobiology of Avian avulavirus 1: Special focus on waterfowl. Vet. Res. 2018, 49, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimitrov, K.M.; Abolnik, C.; Afonso, C.L.; Albina, E.; Bahl, J.; Berg, M.; Briand, F.X.; Brown, I.H.; Choi, K.S.; Chvala, I.; et al. Updated unified phylogenetic classification system and revised nomenclature for Newcastle disease virus. Infect. Genet. Evol. 2019, 74, 103917. [Google Scholar] [CrossRef]
- Chambers, P.; Millar, N.S.; Bingham, R.W.; Emmerson, P.T. Molecular Cloning of Complementary DNA to Newcastle Disease Virus, and Nucleotide Sequence Analysis of the Junction between the Genes Encoding the Haemagglutinin-Neuraminidase and the Large Protein. J. Gen. Virol. 1986, 67, 475–486. [Google Scholar] [CrossRef]
- Collins, P.L.; Wertz, G.W.; Ball, L.A.; Hightower, L.E. Coding assignments of the five smaller mRNAs of Newcastle disease virus. J. Virol. 1982, 43, 1024–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steward, M.; Vipond, I.B.; Millar, N.S.; Emmerson, P.T. RNA editing in Newcastle disease virus. J. Gen. Virol. 1993, 74, 2539–2547. [Google Scholar] [CrossRef]
- Cattoli, G.; Susta, L.; Terregino, C.; Brown, C. Newcastle disease: A review of field recognition and current methods of laboratory detection. J. Vet. Diagn. Investig. 2011, 23, 637–656. [Google Scholar] [CrossRef] [Green Version]
- Meng, C.; Rehman, Z.U.; Liu, K.; Qiu, X.; Tan, L.; Sun, Y.; Liao, Y.; Song, C.; Yu, S.; Ding, Z.; et al. Potential of genotype VII Newcastle disease viruses to cause differential infections in chickens and ducks. Transbound. Emerg. Dis. 2018, 65, 1851–1862. [Google Scholar] [CrossRef] [Green Version]
- Rehman, Z.U.; Qiu, X.; Sun, Y.; Liao, Y.; Tan, L.; Song, C.; Yu, S.; Ding, Z.; Munir, M.; Nair, V.; et al. Vitamin E Supplementation Ameliorates Newcastle Disease Virus-Induced Oxidative Stress and Alleviates Tissue Damage in the Brains of Chickens. Viruses 2018, 10, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butt, S.L.; Moura, V.; Susta, L.; Miller, P.J.; Hutcheson, J.M.; Cardenas-Garcia, S.; Brown, C.C.; West, F.D.; Afonso, C.L.; Stanton, J.B. Tropism of Newcastle disease virus strains for chicken neurons, astrocytes, oligodendrocytes, and microglia. BMC Vet. Res. 2019, 15, 317. [Google Scholar] [CrossRef] [Green Version]
- Ecco, R.; Susta, L.; Afonso, C.L.; Miller, P.J.; Brown, C. Neurological lesions in chickens experimentally infected with virulent Newcastle disease virus isolates. Avian Pathol. 2011, 40, 145–152. [Google Scholar] [CrossRef]
- Rehman, Z.U.; Ren, S.; Yang, B.; Yang, X.; Butt, S.L.; Afzal, A.; Malik, M.I.; Sun, Y.; Yu, S.; Meng, C.; et al. Newcastle disease virus induces testicular damage and disrupts steroidogenesis in specific pathogen free roosters. Vet. Res. 2020, 51, 84. [Google Scholar] [CrossRef]
- Hussein, E.A.; Hair-Bejo, M.; Omar, A.R.; Arshad, S.S.; Hani, H.; Balakrishnan, K.N.; Yakubu, Y.; Saeed, M.I.; Aini, I. Velogenic newcastle disease virus tissue tropism and pathogenesis of infection in chickens by application of in situ PCR, immunoperoxase staining and HE staining. Microb. Pathog. 2019, 129, 213–223. [Google Scholar] [CrossRef]
- Cui, N.; Huang, X.; Kong, Z.; Huang, Y.; Huang, Q.; Yang, S.; Zhang, L.; Xu, C.; Zhang, X.; Cui, Y. Newcastle Disease Virus Infection Interferes With the Formation of Intestinal Microflora in Newly Hatched Specific-Pathogen-Free Chicks. Front. Microbiol. 2018, 9, 900. [Google Scholar] [CrossRef]
- Nakamura, K.; Ohtsu, N.; Nakamura, T.; Yamamoto, Y.; Yamada, M.; Mase, M.; Imai, K. Pathologic and immunohistochemical studies of Newcastle disease (ND) in broiler chickens vaccinated with ND: Severe nonpurulent encephalitis and necrotizing pancreatitis. Vet. Pathol. 2008, 45, 928–933. [Google Scholar] [CrossRef]
- Susta, L.; Miller, P.J.; Afonso, C.L.; Brown, C.C. Clinicopathological characterization in poultry of three strains of Newcastle disease virus isolated from recent outbreaks. Vet. Pathol. 2011, 48, 349–360. [Google Scholar] [CrossRef]
- El-Bahrawy, A.; Zaid, A.; Sunden, Y.; Sakurai, M.; Ito, H.; Ito, T.; Morita, T. Pathogenesis of Pancreatitis in Chickens after Experimental Infection with 9a5b Newcastle Disease Virus Mutant Isolate. J. Comp. Pathol. 2015, 153, 315–323. [Google Scholar] [CrossRef]
- Meulemans, G.; Roels, S.; van den Berg, T.P.; Godfroid, J.; Decaesstecker, M. Acute pancreatitis in chickens due to non-virulent Newcastle disease virus. Vet. Rec. 1998, 143, 300–303. [Google Scholar] [CrossRef]
- Denbow, D.M. Chapter 14—Gastrointestinal Anatomy and Physiology. In Sturkie’s Avian Physiology, 6th ed.; Scanes, C.G., Ed.; Academic Press: San Diego, CA, USA, 2015; pp. 337–366. [Google Scholar] [CrossRef]
- Laws, B.M.; Moore, J.H. The lipase and esterase activities of the pancreas and small intestine of the chick. Biochem. J. 1963, 87, 632–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahdavi, R.; Osmanyan, A.K.; Fisinin, V.I.; Ghazi Harsini, S.; Arkhipova, A.L.; Shevyakov, A.N.; Kovalchuk, S.N.; Kosovsky, G.Y. Impact of mash and crumble diets on intestinal amino acids transporters, intestinal morphology and pancreatic enzyme activity of broilers. J. Anim. Physiol. Anim. Nutr. (Berl.) 2018, 102, 1266–1273. [Google Scholar] [CrossRef]
- Yuan, L.; Wang, M.; Zhang, X.; Wang, Z. Effects of protease and non-starch polysaccharide enzyme on performance, digestive function, activity and gene expression of endogenous enzyme of broilers. PLoS ONE 2017, 12, e0173941. [Google Scholar] [CrossRef] [Green Version]
- Dupont, J.; Rideau, N.; Simon, J. Chapter 27—Endocrine Pancreas. In Sturkie’s Avian Physiology, 6th ed.; Scanes, C.G., Ed.; Academic Press: San Diego, CA, USA, 2015; pp. 613–631. [Google Scholar] [CrossRef]
- Kommers, G.D.; King, D.J.; Seal, B.S.; Brown, C.C. Pathogenesis of chicken-passaged Newcastle disease viruses isolated from chickens and wild and exotic birds. Avian Dis. 2003, 47, 319–329. [Google Scholar] [CrossRef]
- Susta, L.; Cornax, I.; Diel, D.G.; Garcia, S.C.; Miller, P.J.; Liu, X.; Hu, S.; Brown, C.C.; Afonso, C.L. Expression of interferon γ by a highly virulent strain of Newcastle disease virus decreases its pathogenicity in chickens. Microb. Pathog. 2013, 61–62, 73–83. [Google Scholar] [CrossRef]
- Dai, Y.; Cheng, X.; Liu, M.; Shen, X.; Li, J.; Yu, S.; Zou, J.; Ding, C. Experimental infection of duck origin virulent Newcastle disease virus strain in ducks. BMC Vet. Res. 2014, 10, 164. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.; Li, Y.; Yuan, R.; Li, X.; Sun, M.; Wang, Z.; Feng, M.; Jiao, P.; Ren, T. Phylogenetic relationships and pathogenicity variation of two Newcastle disease viruses isolated from domestic ducks in Southern China. Virol. J. 2014, 11, 147. [Google Scholar] [CrossRef] [Green Version]
- Piacenti, A.M.; King, D.J.; Seal, B.S.; Zhang, J.; Brown, C.C. Pathogenesis of Newcastle disease in commercial and specific pathogen-free turkeys experimentally infected with isolates of different virulence. Vet. Pathol. 2006, 43, 168–178. [Google Scholar] [CrossRef]
- Cornax, I.; Miller, P.J.; Afonso, C.L. Characterization of live LaSota vaccine strain-induced protection in chickens upon early challenge with a virulent Newcastle disease virus of heterologous genotype. Avian Dis. 2012, 56, 464–470. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Munro, B. Manual of histologic staining methods of the Armed Forces Institute of Pathology. Pathology 1971, 3, 249. [Google Scholar] [CrossRef]
- Qu, Y.; Zhan, Y.; Yang, S.; Ren, S.; Qiu, X.; Rehamn, Z.U.; Tan, L.; Sun, Y.; Meng, C.; Song, C.; et al. Newcastle disease virus infection triggers HMGB1 release to promote the inflammatory response. Virology 2018, 525, 19–31. [Google Scholar] [CrossRef]
- Assaad, H.I.; Hou, Y.; Zhou, L.; Carroll, R.J.; Wu, G. Rapid publication-ready MS-Word tables for two-way ANOVA. Springerplus 2015, 4, 33. [Google Scholar] [CrossRef] [Green Version]
- Chansiripornchai, N.; Sasipreeyajan, J. Efficacy of live B1 or Ulster 2C Newcastle disease vaccines simultaneously vaccinated with inactivated oil adjuvant vaccine for protection of Newcastle disease virus in broiler chickens. Acta Vet. Scand. 2006, 48, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owen, J.C.; Nakamura, A.; Coon, C.A.; Martin, L.B. The effect of exogenous corticosterone on West Nile virus infection in Northern Cardinals (Cardinalis cardinalis). Vet. Res. 2012, 43. [Google Scholar] [CrossRef] [Green Version]
- Ives, A.M.; Bertke, A.S. Stress hormones epinephrine and corticosterone selectively modulate herpes simplex virus 1 (HSV-1) and HSV-2 productive infections in adult sympathetic, but not sensory, neurons. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- Gross, W.; Siegel, P.; DuBose, R. Some effects of feeding corticosterone to chickens. Poult. Sci. 1980, 59, 516–522. [Google Scholar] [CrossRef]
- Zhao, J.; Lin, H.; Jiao, H.; Song, Z. Corticosterone suppresses insulin-and NO-stimulated muscle glucose uptake in broiler chickens (Gallus gallus domesticus). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2009, 149, 448–454. [Google Scholar] [CrossRef]
- Hazelwood, R.L. CHAPTER 20—Pancreas. In Sturkie’s Avian Physiology, 5th ed.; Whittow, G.C., Ed.; Academic Press: San Diego, CA, USA, 2000; pp. 539–555. [Google Scholar] [CrossRef]
- Rehman, Z.U.; Che, L.; Ren, S.; Liao, Y.; Qiu, X.; Yu, S.; Sun, Y.; Tan, L.; Song, C.; Liu, W.; et al. Supplementation of Vitamin E Protects Chickens from Newcastle Disease Virus-Mediated Exacerbation of Intestinal Oxidative Stress and Tissue Damage. Cell Physiol. Biochem. 2018, 47, 1655–1666. [Google Scholar] [CrossRef]
- Alison, B. The distribution and ontogeny of gastrin/CCK-, somatostatin-and neurotensin-immunoreactive cells in the gastrointestinal tract of the chicken. Histol. Histopathol. 1989, 4, 55–62. [Google Scholar]
- Ku, S.K.; Lee, J.H.; Lee, H.S. An immunohistochemical study of the insulin-, glucagon- and somatostatin-immunoreactive cells in the developing pancreas of the chicken embryo. Tissue Cell 2000, 32, 58–65. [Google Scholar] [CrossRef]
- Malik, N.; Moaeen-ud-Din, M.; Zhao, R. Ontogeny of mRNA expression of somatostatin and its receptors in chicken embryos in association with methylation status of their promoters. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2013, 165, 260–270. [Google Scholar] [CrossRef]
- Trabucchi, M.; Tostivint, H.; Lihrmann, I.; Blähser, S.; Vallarino, M.; Vaudry, H. Characterization of the cDNA encoding a somatostatin variant in the chicken brain: Comparison of the distribution of the two somatostatin precursor mRNAs. J. Comp. Neurol. 2003, 461, 441–451. [Google Scholar] [CrossRef]
- Low, M.J. Somatostatin. In Encyclopedia of Hormones; Henry, H.L., Norman, A.W., Eds.; Academic Press: New York, NY, USA, 2003; pp. 379–388. [Google Scholar] [CrossRef]
- Elliott, D.E.; Blum, A.M.; Li, J.; Metwali, A.; Weinstock, J.V. Preprosomatostatin messenger RNA is expressed by inflammatory cells and induced by inflammatory mediators and cytokines. J. Immunol. 1998, 160, 3997–4003. [Google Scholar]
- Briard, N.; Guillaume, V.; Frachebois, C.; Rico-Gomez, M.; Sauze, N.; Oliver, C.; Dutour, A. Endotoxin injection increases growth hormone and somatostatin secretion in sheep. Endocrinology 1998, 139, 2662–2669. [Google Scholar] [CrossRef]
- Revhaug, A.; Lygren, I.; Lundgren, T.I.; Jorde, R.; Burhol, P.G.; Giercksky, K.E. Release of gastrointestinal peptides during E. coli endotoxinaemia. Acta Chir. Scand. 1984, 150, 535–539. [Google Scholar]
- Hendricks, E.E.; Ludlage, E.; Bussell, S.; George, K.; Wegner, F.H.; Mansfield, K.G. Wasting syndrome and disruption of the somatotropic axis in simian immunodeficiency virus-infected macaques with Mycobacterium avium complex infection. J. Infect. Dis. 2004, 190, 2187–2194. [Google Scholar] [CrossRef] [Green Version]
- Weihe, E.; Bette, M.; Preuss, M.A.; Faber, M.; Schafer, M.K.; Rehnelt, J.; Schnell, M.J.; Dietzschold, B. Role of virus-induced neuropeptides in the brain in the pathogenesis of rabies. Dev. Biol. 2008, 131, 73–81. [Google Scholar]
- Linscheid, P.; Seboek, D.; Müller, B.; Zulewski, H.; Christ-Crain, M.; Keller, U.; Langer, I. Somatostatin Is Expressed and Secreted by Human Adipose Tissue upon Infection and Inflammation. J. Clin. Endocrinol. Metab. 2004, 89, 4833–4839. [Google Scholar] [CrossRef] [Green Version]
- Takeba, Y.; Suzuki, N.; Takeno, M.; Asai, T.; Tsuboi, S.; Hoshino, T.; Sakane, T. Modulation of synovial cell function by somatostatin in patients with rheumatoid arthritis. Arthritis Rheum. 1997, 40, 2128–2138. [Google Scholar] [CrossRef]
- Sakane, T.; Suzuki, N. The role of somatostatin in the pathophysiology of rheumatoid arthritis. Clin. Exp. Rheumatol. 1998, 16, 745–749. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rehman, Z.U.; Ren, S.; Butt, S.L.; Manzoor, Z.; Iqbal, J.; Anwar, M.N.; Sun, Y.; Qiu, X.; Tan, L.; Liao, Y.; et al. Newcastle Disease Virus Induced Pathologies Severely Affect the Exocrine and Endocrine Functions of the Pancreas in Chickens. Genes 2021, 12, 495. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12040495
Rehman ZU, Ren S, Butt SL, Manzoor Z, Iqbal J, Anwar MN, Sun Y, Qiu X, Tan L, Liao Y, et al. Newcastle Disease Virus Induced Pathologies Severely Affect the Exocrine and Endocrine Functions of the Pancreas in Chickens. Genes. 2021; 12(4):495. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12040495
Chicago/Turabian StyleRehman, Zaib Ur, Shanhui Ren, Salman Latif Butt, Zahid Manzoor, Javid Iqbal, Muhammad Naveed Anwar, Yingjie Sun, Xusheng Qiu, Lei Tan, Ying Liao, and et al. 2021. "Newcastle Disease Virus Induced Pathologies Severely Affect the Exocrine and Endocrine Functions of the Pancreas in Chickens" Genes 12, no. 4: 495. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12040495