Genomic and Phenotypic Diversity of Cultivated and Wild Tomatoes with Varying Levels of Heat Tolerance

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Phenotypic Evaluation
2.3. Genotyping of the Germplasm Collection
2.4. SNP Filtering Summary
2.5. Genetic Population Structure Analysis
2.6. Combined Phenotypic and Genomic Datasets
3. Results
3.1. Variance Components of the Heat Tolerance Traits and Heritability Estimate
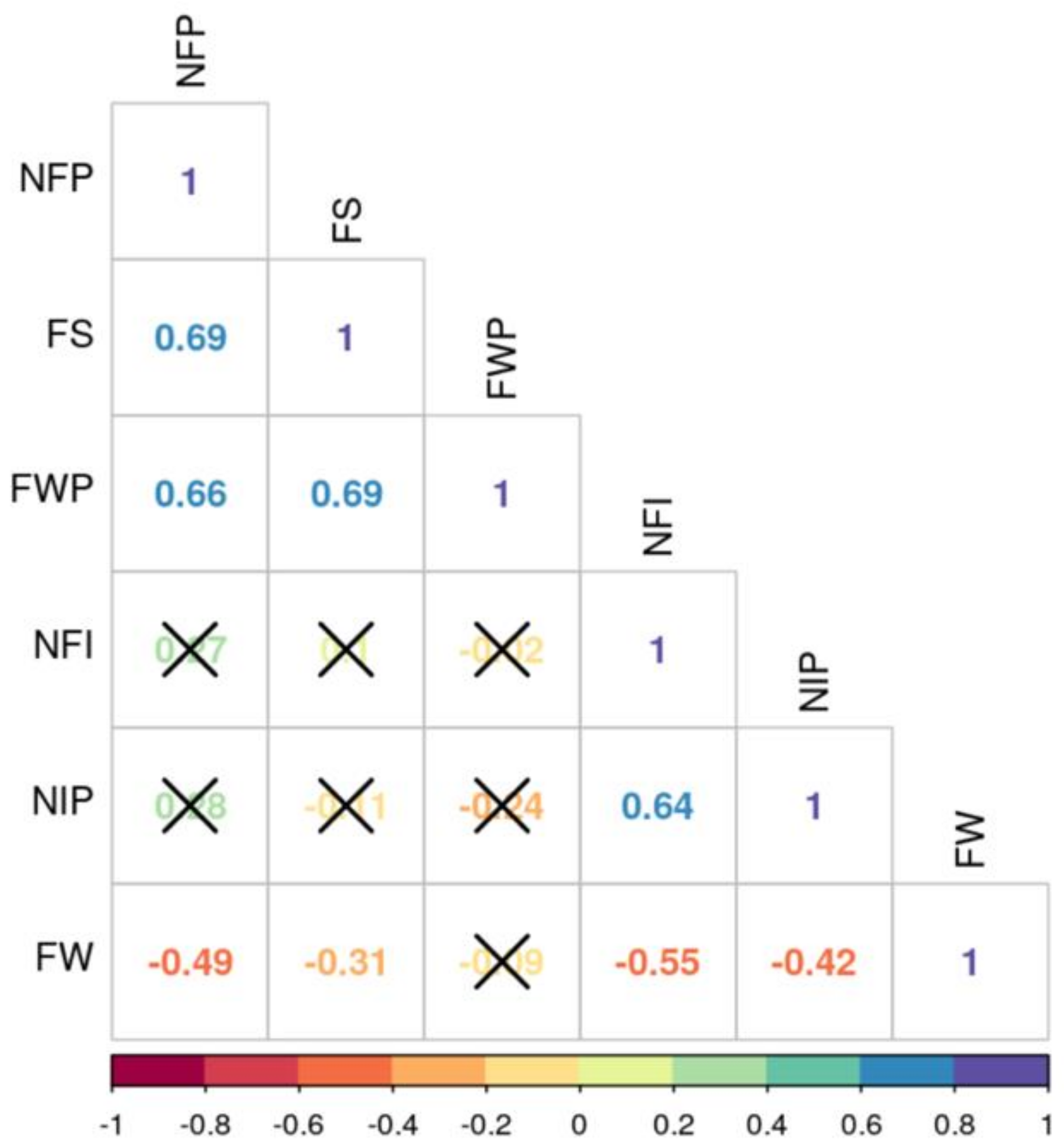
3.2. Correlation Analysis Among the Quantitative Traits
3.3. Phenotypic Based-Clustering Analysis of the Tomato Lines
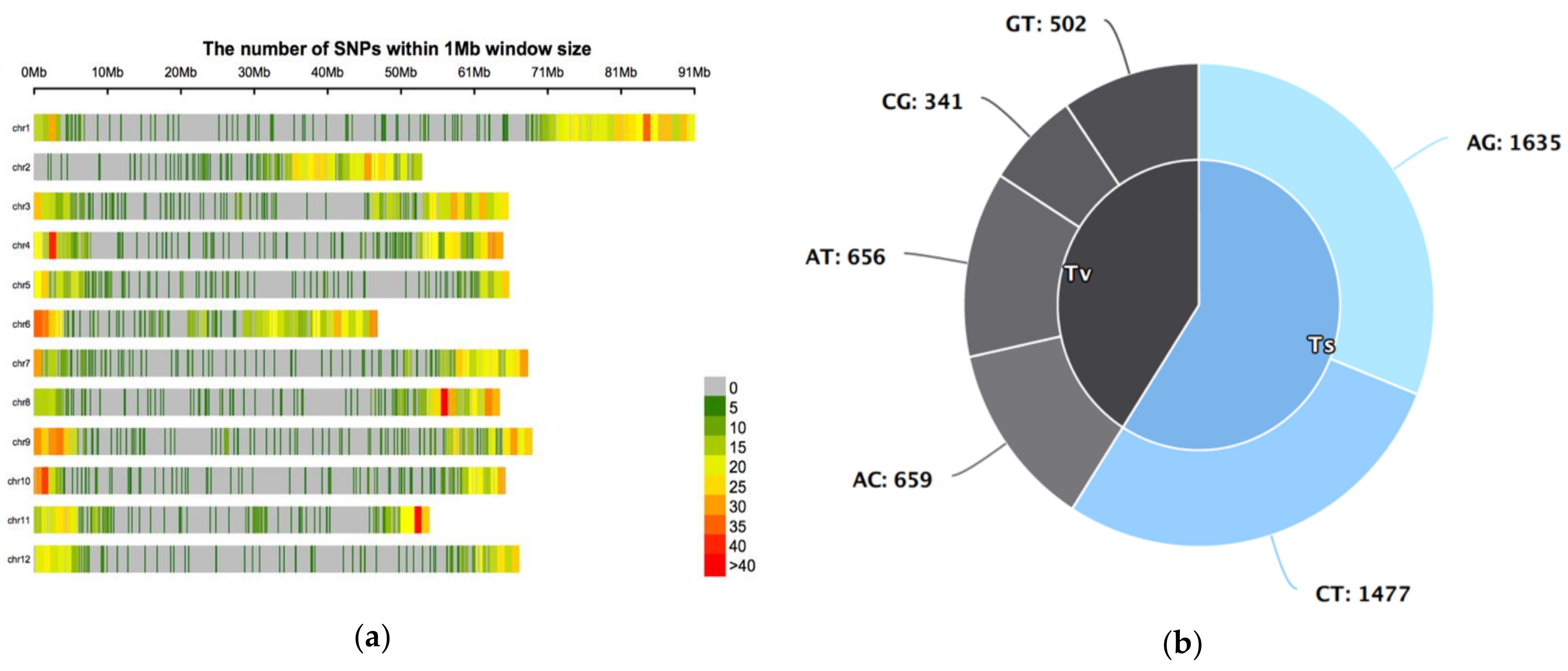
3.4. Characteristics of the 5270 SNPs in the Germplasm Collection
3.5. Population Structure
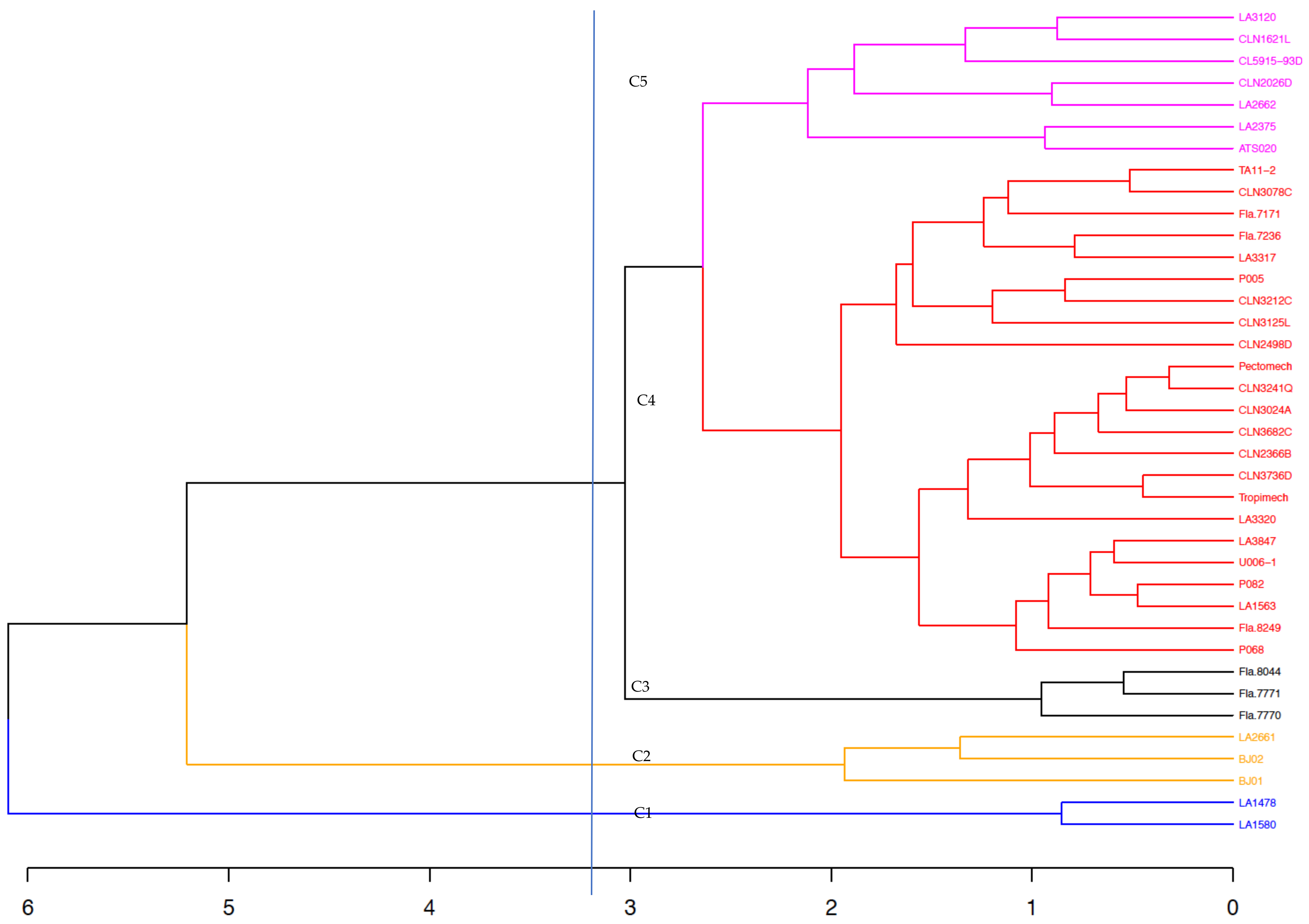
3.6. Genetic Distances Among the Tomato Lines Based on the 5270 SNPs and Hierarchical Clustering
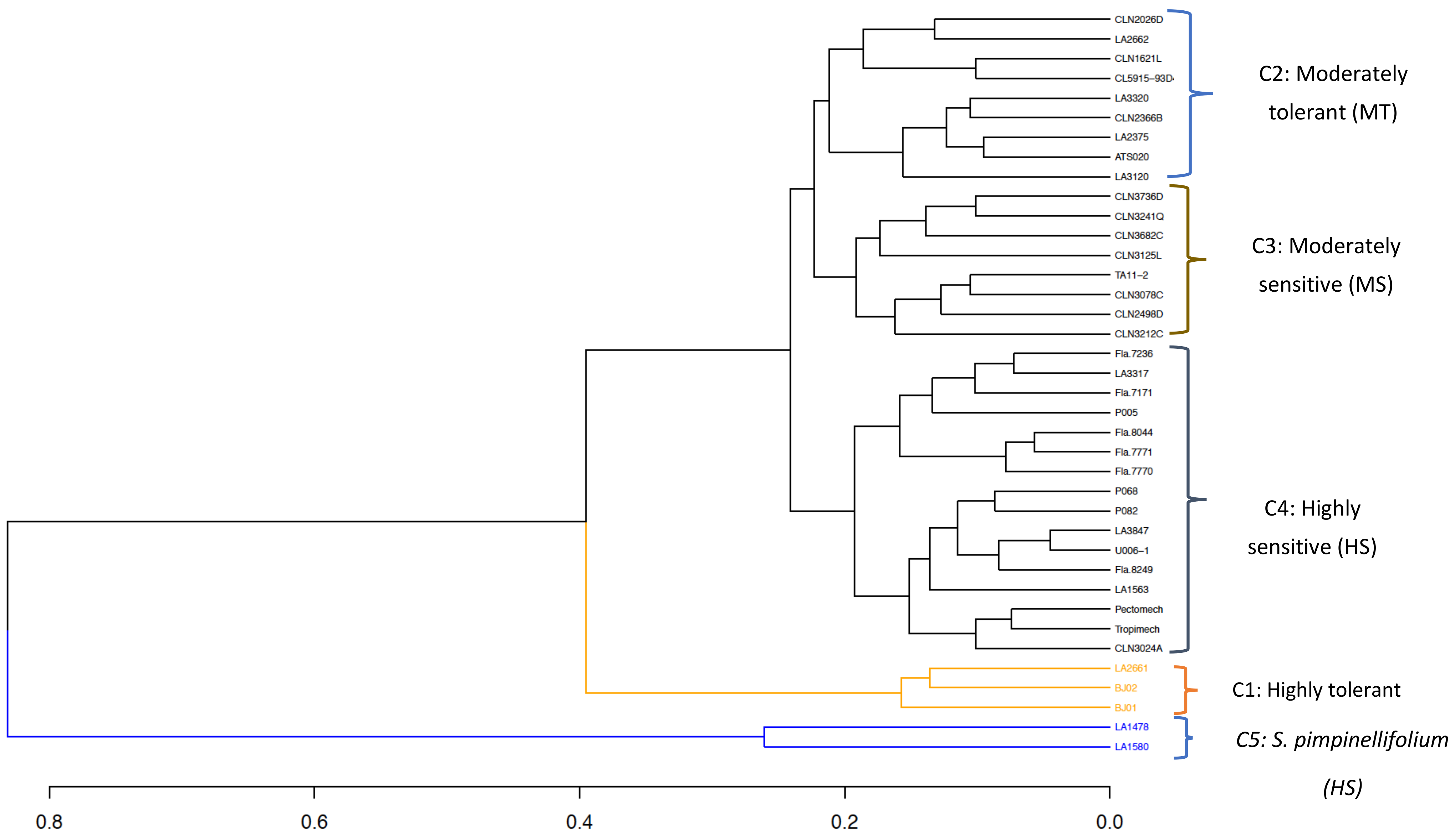
3.7. Combined Phenotypic and Genomic Clustering
3.8. Differentiation Among Populations
4. Discussion
4.1. SNP Quality
4.2. Genetic Diversity in the Germplasm Collection
4.3. Broad Sense Heritability and Trait Associations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. FAO Statistical Division. Available online: http://www.fao.org/faostat/en/#data/TM (accessed on 22 March 2020).
- Keatinge, J.D.H.; Lin, L.J.; Ebert, A.W.; Chen, W.Y.; Hughes, J.D.A.; Luther, G.C.; Wang, J.F.; Ravishankar, M. Overcoming biotic and abiotic stresses in the Solanaceae through grafting: Current status and future perspectives. Biol. Agric. Hortic. 2014, 30, 272–287. [Google Scholar] [CrossRef]
- Silva, R.S.; Kumar, L.; Shabani, F.; Picanço, M.C. Assessing the impact of global warming on worldwide open field tomato cultivation through CSIRO-Mk3.0 global climate model. J. Agric. Sci. 2017, 155, 407–420. [Google Scholar] [CrossRef] [Green Version]
- Camejo, D.; Rodríguez, P.; Morales, M.A.; Dell’Amico, J.M.; Torrecillas, A.; Alarcón, J.J. High temperature effects on photosynthetic activity of two tomato cultivars with different heat susceptibility. J. Plant Physiol. 2005, 162, 281–289. [Google Scholar] [CrossRef]
- Zhou, R.; Kjær, K.H.; Rosenqvist, E.; Yu, X.; Wu, Z.; Ottosen, C.O. Physiological Response to Heat Stress During Seedling and Anthesis Stage in Tomato Genotypes Differing in Heat Tolerance. J. Agron. Crop Sci. 2017, 203, 68–80. [Google Scholar] [CrossRef]
- Ayenan, M.A.T.; Danquah, A.; Hanson, P.; Ampomah-dwamena, C.; Sodedji, A.F.K.; Asante, I.K.; Danquah, E.Y. Accelerating Breeding for Heat Tolerance in Tomato (Solanum lycopersicum L.): An Integrated Approach. Agronomy 2019, 9, 720. [Google Scholar] [CrossRef] [Green Version]
- National Aeronautics and Space Administration (NASA). Scientific Consensus: Earth’s Climate is Warming. Available online: https://climate.nasa.gov/scientific-consensus/ (accessed on 20 August 2020).
- Litskas, V.D.; Migeon, A.; Navajas, M.; Tixier, M. Impacts of climate change on tomato, a notorious pest and its natural enemy: Small scale agriculture at higher risk Impacts of climate change on tomato, a notorious pest and its natural enemy: Small scale agriculture at higher risk. Environ. Res. Lett. Lett. 2019, 14, 084041. [Google Scholar] [CrossRef]
- Bita, C.E.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Driedonks, N.; Rieu, I.; Vriezen, W.H. Breeding for plant heat tolerance at vegetative and reproductive stages. Plant Reprod. 2016, 29, 67–79. [Google Scholar] [CrossRef] [Green Version]
- Ruggieri, V.; Calafiore, R.; Schettini, C.; Rigano, M.M.; Olivieri, F.; Frusciante, L.; Barone, A. Exploiting Genetic and Genomic Resources to Enhance Heat-Tolerance in Tomatoes. Agronomy 2019, 9, 22. [Google Scholar] [CrossRef] [Green Version]
- De la Peña, R.; Hughes, J. Improving Vegetable Productivity in a Variable Changing Climate. J. Publshed ICRISAT 2007, 4, 1–22. [Google Scholar]
- Mascher, M.; Schreiber, M.; Scholz, U.; Graner, A.; Reif, J.C.; Stein, N. Genebank genomics bridges the gap between the conservation of crop diversity and plant breeding. Nat. Genet. 2019, 51, 1076–1081. [Google Scholar] [CrossRef]
- Shaheen, M.R.; Ayyub, C.M.; Amjad, M.; Waraich, E.A. Morpho-physiological evaluation of tomato genotypes under high temperature stress conditions. J. Sci. Food Agric. 2016, 96, 2698–2704. [Google Scholar] [CrossRef] [PubMed]
- Paupière, M.J.; van Haperen, P.; Rieu, I.; Visser, R.G.F.; Tikunov, Y.M.; Bovy, A.G. Screening for pollen tolerance to high temperatures in tomato. Euphytica 2017, 213. [Google Scholar] [CrossRef]
- Driedonks, N.; Wolters-Arts, M.; Huber, H.; de Boer, G.J.; Vriezen, W.; Mariani, C.; Rieu, I. Exploring the natural variation for reproductive thermotolerance in wild tomato species. Euphytica 2018, 214. [Google Scholar] [CrossRef] [Green Version]
- Ruggieri, V.; Francese, G.; Sacco, A.; Alessandro, A.D.; Rigano, M.M.; Parisi, M.; Milone, M.; Cardi, T.; Mennella, G.; Barone, A. An association mapping approach to identify favourable alleles for tomato fruit quality breeding. BMC Plant Biol. 2014, 14, 337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarano, A.; Olivieri, F.; Gerardi, C.; Liso, M.; Chiesa, M.; Chieppa, M.; Frusciante, L.; Barone, A.; Santino, A.; Manuela, M. Selection of tomato landraces with high fruit yield and nutritional quality under elevated temperatures. J. Sci. Food Agric. 2020, 100, 2791–2799. [Google Scholar] [CrossRef]
- Olivieri, F.; Calafiore, R.; Francesca, S.; Schettini, C.; Chiaiese, P.; Rigano, M.M.; Barone, A. High-Throughput Genotyping of Resilient Tomato Landraces to Detect Candidate Genes Involved in the Response to High Temperatures. Genes 2020, 11, 626. [Google Scholar] [CrossRef]
- Alsamir, M.; Ahmad, N.; Arief, V.; Mahmood, T.; Trethowan, R. Phenotypic diversity and marker-trait association studies under heat stress in tomato (Solanum lycopersicum L.). Aust. J. Crop Sci. 2019, 13, 578–587. [Google Scholar] [CrossRef]
- Agre, P.; Asibe, F.; Darkwa, K.; Edemodu, A.; Bauchet, G.; Asiedu, R.; Adebola, P.; Asfaw, A. Phenotypic and molecular assessment of genetic structure and diversity in a panel of winged yam (Dioscorea alata) clones and cultivars. Sci. Rep. 2019, 9, 18221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gediya, L.N.; Patel, D.A.; Kumar, S.; Kumar, D.; Parmar, D.J.; Patel, S.S. Phenotypic variability, path analysis and molecular diversity analysis in chickpea (Cicer arietinum L.). Vegetos 2019, 32, 167–180. [Google Scholar] [CrossRef]
- Ayenan, M.A.T.; Danquah, A.; Hanson, P.; Asante, I.K.; Danquah, E.Y. Identification of new sources of heat tolerance in cultivated and wild tomatoes. Euphytica 2021, 217, 1–16. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix (Version 0.84). 2017. Available online: https://github.com/taiyun/corrplot (accessed on 15 October 2020).
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Falconer, D.S.; Mackay, T.F.C. An Introduction to Quantitative Genetic, 4th ed.; Prentice Hall: London, UK, 1996. [Google Scholar]
- Nyquist, W.E.; Baker, R.J. Estimation of heritability and prediction of selection response in plant populations. CRC. Crit. Rev. Plant Sci. 1991, 10, 235–322. [Google Scholar] [CrossRef]
- Galili, T. dendextend: An R package for visualizing, adjusting, and comparing trees of hierarchical clustering. Bioinformatics 2015, 31, 3718–3720. [Google Scholar] [CrossRef] [Green Version]
- Kilian, A.; Wenzl, P.; Huttner, E.; Carling, J.; Xia, L.; Blois, H.; Caig, V.; Heller-Uszynska, K.; Jaccoud, D.; Hopper, C.; et al. Diversity arrays technology: A generic genome profiling technology on open platforms. Methods Mol. Biol. 2012, 888, 67–89. [Google Scholar] [CrossRef]
- Browning, S.R.; Browning, B.L. Rapid and accurate haplotype phasing and missing-data inference for whole-genome association studies by use of localized haplotype clustering. Am. J. Hum. Genet. 2007, 81, 1084–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Dereeper, A.; Homa, F.; Andres, G.; Sempere, G.; Sarah, G.; Hueber, Y.; Dufayard, J.F.; Ruiz, M. SNiPlay3: A web-based application for exploration and large scale analyses of genomic variations. Nucleic Acids Res. 2015, 43, W295–W300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R Package Version 1.0.6. 2019. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 15 October 2020).
- Jombart, T.; Devillard, S.; Balloux, F.; Falush, D.; Stephens, M.; Pritchard, J.; Pritchard, J.; Stephens, M.; Donnelly, P.; Corander, J.; et al. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 15 October 2020).
- Ranc, N.; Muños, S.; Xu, J.; Le Paslier, M.C.; Chauveau, A.; Bounon, R.; Rolland, S.; Bouchet, J.P.; Brunel, D.; Causse, M. Genome-Wide association mapping in tomato (Solanum lycopersicum) is possible using genome admixture of solanum lycopersicum var. cerasiforme. G3 Genes Genomes Genet. 2012, 2, 853–864. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, P.; Zhang, Z.; Kroon, D.; Casstevens, T.; Ramdoss, Y.; Buckler, E. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Charrad, M.; Ghazzali, N.; Boiteau, V.; Niknafs, A. NbClust: An R Package for Determining the Relevant Number of Clusters in a Data Set. J. Stat. Softw. 2014, 61, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-6. 2019. Available online: https://CRAN.Rproject.org/package=vegan (accessed on 15 October 2020).
- Simpson, G.L.; Oksanen, J. Analogue: Analogue Matching and Modern Analogue Technique Transfer Function Models. (R Package Version 0.17-4). 2020. Available online: https://cran.r-project.org/package=analogue (accessed on 15 October 2020).
- Weir, B.S.; Cockerham, C.C. Estimating F-Statistics for the Analysis of Population Structure. Evolution 1984, 38, 1358–1370. [Google Scholar] [CrossRef] [PubMed]
- Goudet, J.; Jombart, T. Hierfstat: Estimation and Tests of Hierarchical F-Statistics. R Package Version 0.04-22. 2015. Available online: https://CRAN.R-project.org/package=hierfstat (accessed on 15 October 2020).
- Sim, S.C.; van Deynze, A.; Stoffel, K.; Douches, D.S.; Zarka, D.; Ganal, M.W.; Chetelat, R.T.; Hutton, S.F.; Scott, J.W.; Gardner, R.G.; et al. High-Density SNP Genotyping of Tomato (Solanum lycopersicum L.) Reveals Patterns of Genetic Variation Due to Breeding. PLoS ONE 2012, 7, e45520. [Google Scholar] [CrossRef] [Green Version]
- Tomato Genome Consortium The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641. [CrossRef] [Green Version]
- Serba, D.D.; Muleta, K.T.; St. Amand, P.; Bernardo, A.; Bai, G.; Perumal, R.; Bashir, E. Genetic Diversity, Population Structure, and Linkage Disequilibrium of Pearl Millet. Plant Genome 2019, 12, 180091. [Google Scholar] [CrossRef] [Green Version]
- Pailles, Y.; Ho, S.; Pires, I.S.; Tester, M.; Negrão, S.; Schmöcke, S.M. Genetic diversity and population structure of two tomato species from the galapagos islands. Front. Plant Sci. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuriaga, E.; Blanca, J.M.; Cordero, L.; Sifres, A.; Blas-Cerdán, W.G.; Morales, R.; Nuez, F. Genetic and bioclimatic variation in Solanum pimpinellifolium. Genet. Resour. Crop Evol. 2009, 56, 39–51. [Google Scholar] [CrossRef]
- Bauchet, G.; Causse, M. Genetic diversity in tomato (Solanum lycopersicum) and its wild relatives. In Genetic Diversity in Plants; IN-TECH Education and Publishing: London, UK, 2012; pp. 953–978. [Google Scholar] [CrossRef] [Green Version]
- Tanksley, S.D.; Nelson, J.C. Advanced backcross QTL analysis: A method for the simultaneous discovery and transfer of valuable QTLs from unadapted germplasm into elite breeding lines. Theor. Appl. Genet. 1996, 92, 191–203. [Google Scholar] [CrossRef]
- Grandillo, S.; Tanksley, S.D. Advanced backcross QTL analysis: Results and perspectives. In Proceedings of the International Congress “In the Wake of the Double Helix: From the Green Revolution to the Gene Revolution”; Tuberosa, R., Phillips, R.L., Gale, M., Eds.; Avenue Media: Bologna, Italy, 2005; pp. 115–132. [Google Scholar]
- Nagata, K.; Ando, T.; Nonoue, Y.; Mizubayashi, T.; Kitazawa, N.; Shomura, A.; Matsubara, K.; Ono, N.; Mizobuchi, R.; Shibaya, T.; et al. Advanced backcross QTL analysis reveals complicated genetic control of rice grain shape in a japonica × indica cross. Breed. Sci. 2015, 65, 308–318. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, M.J.; Pastina, M.M.; De Souza, V.F.; Schaffert, R.E.; Carneiro, P.C.S.; Noda, R.W.; de Souza Carneiro, J.E.; Damasceno, C.M.B.; da Costa Parrella, R.A. Phenotypic and molecular characterization of sweet sorghum accessions for bioenergy production. PLoS ONE 2017, 12, e0183504. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.P.; Nodari, R.; Gepts, P. Genetic Diversity in Cultivated Common Bean: I. Allozymes. Crop Sci. 1991, 31, 19–23. [Google Scholar] [CrossRef]
- Melchinger, A.E.; Gumber, R.K. Overview of Heterosis and Heterotic Groups in Agronomic Crops. Concepts Breed. Heterosis Crop Plants 1998, 25, 29–44. [Google Scholar] [CrossRef]
- Rick, C.M.; Fobes, J.F.; Holle, M. Genetic variation in Lycopersicon pimpinellifolium: Evidence of evolutionary change in mating systems. Plant Syst. Evol. 1977, 127, 139–170. [Google Scholar] [CrossRef]
- Xu, J.; Driedonks, N.; Rutten, M.J.M.; Vriezen, W.H.; de Boer, G.-J.; Rieu, I. Mapping quantitative trait loci for heat tolerance of reproductive traits in tomato (Solanum lycopersicum). Mol. Breed. 2017, 37, 58. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.W. Fla. 7771, a Medium-large, Heat- tolerant, Jointless-pedicel Tomato. HortScience 2000, 35, 968–969. [Google Scholar] [CrossRef] [Green Version]
- Scott, J.W.; Olson, S.M.; Bryan, H.H.; Bartz, J.A.; Maynard, D.N.; Stoffella, P.J. “Solar Fire” hybrid tomato: Fla. 7776 tomato breeding line. HortScience 2006, 41, 1504–1505. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Weerakoon, W.M.W.; Maruyama, A.; Ohba, K. Impact of Humidity on Temperature-Induced Grain Sterility in Rice (Oryza sativa L.). J. Agron. Crop Sci. 2008, 194, 135–140. [Google Scholar] [CrossRef]
- Sherzod, R.; Yang, E.Y.; Cho, M.C.; Chae, S.Y.; Chae, W.B. Physiological traits associated with high temperature tolerance differ by fruit types and sizes in tomato (Solanum lycopersicum L.). Hortic. Environ. Biotechnol. 2020, 61, 837–847. [Google Scholar] [CrossRef]
- Panthee, D.R.; Kressin, J.P.; Piotrowski, A. Heritability of flower number and fruit set under heat stress in tomato. HortScience 2018, 53, 1294–1299. [Google Scholar] [CrossRef] [Green Version]
- Zörb, C.; Piepho, H.-P.; Zikeli, S.; Horneburg, B. Heritability and Variability of Quality Parameters of Tomatoes in Outdoor Production. Research 2020, 2020, 6707529. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lines/Accessions | Species | Heat Tolerance Status | Sources of Accessions/Lines |
|---|---|---|---|
| Fla.7770 | S. lycopersicum | Sensitive | University of Florida |
| Fla.7771 | S. lycopersicum | Sensitive | University of Florida |
| Fla.7171 | S. lycopersicum | Sensitive | University of Florida |
| Fla.8044 | S. lycopersicum | Sensitive | University of Florida |
| Fla.7236 | S. lycopersicum | Sensitive | University of Florida |
| Fla.8249 | S. lycopersicum | Sensitive | University of Florida |
| Fla.7776 | S. lycopersicum | Sensitive | University of Florida |
| LA2661 (Nagcarlang) | S. lycopersicum | Tolerant | TGRC, UC Davis |
| LA3317 (Campbell 28) | S. lycopersicum | Tolerant | TGRC, UC Davis |
| LA2662 (Saladette) | S. lycopersicum | Tolerant | TGRC, UC Davis |
| LA3120 (Malintka 101) | S. lycopersicum | Tolerant | TGRC, UC Davis |
| LA1563 | S. lycopersicum | Sensitive | TGRC, UC Davis |
| LA2375 (San Marzano) | S. lycopersicum | Tolerant | TGRC, UC Davis |
| LA3320 (Hotset) | S. lycopersicum | Sensitive | TGRC, UC Davis |
| LA0657 (Beaverlodge) | S. lycopersicum | Tolerant | TGRC, UC Davis |
| LA3847 (NCHS-1) | S. lycopersicum | Sensitive | TGRC, UC Davis |
| LA1580 | S. pimpinellifolium | Sensitive | TGRC, UC Davis |
| LA2854 | S. pimpinellifolium | Sensitive | TGRC, UC Davis |
| LA1478 | S. pimpinellifolium | Sensitive | TGRC, UC Davis |
| U006-1 | S. lycopersicum | Sensitive | Off-type isolated from LA1563 during seed multiplication |
| CLN3212C | S. lycopersicum | Tolerant | WorldVeg |
| CLN1621L | S. lycopersicum | Tolerant | WorldVeg |
| CLN3125L | S. lycopersicum | Tolerant | WorldVeg |
| CLN3241Q | S. lycopersicum | Tolerant | WorldVeg |
| CLN3078C | S. lycopersicum | Tolerant | WorldVeg |
| CLN3682C | S. lycopersicum | Tolerant | WorldVeg |
| CLN3024A | S. lycopersicum | Tolerant | WorldVeg |
| CL5915-93D4-1-0-3 | S. lycopersicum | Tolerant | WorldVeg |
| CLN2366B | S. lycopersicum | Tolerant | WorldVeg |
| CLN2498D | S. lycopersicum | Tolerant | WorldVeg |
| CLN2026D | S. lycopersicum | Tolerant | WorldVeg |
| CLN3736D | S. lycopersicum | Sensitive | WorldVeg |
| TA11-2 | S. lycopersicum | Tolerant | Off-type isolated from CLN2498D during seed multiplication |
| BJ01 | S. lycopersicum | Tolerant | Benin |
| BJ02 | S. lycopersicum | Tolerant | Benin |
| ATS020 | S. lycopersicum | Tolerant | Ghana |
| P082 | S. lycopersicum | Sensitive | Ghana |
| P005 | S. lycopersicum | Sensitive | Ghana |
| P068 | S. lycopersicum | Sensitive | Ghana |
| WACI1 | S. lycopersicum | Tolerant | Ghana |
| Tropimech | S. lycopersicum | Sensitive | Commercial OPV variety |
| Pectomech | S. lycopersicum | Sensitive | Commercial OPV variety |
| Siberia | S. lycopersicum | Unknown* | WorldVeg |
| Tstar-29-4-3-12-6-11-4 | S. lycopersicum | Unknown* | WorldVeg |
| CLN4220F5-186 | S. lycopersicum | Unknown* | WorldVeg |
| Locations | Range of Night Temperatures (°C) | Range of Daytime Temperatures (°C) | Range of Night Relative Humidity (%) | Range of Day Relative Humidity (%) |
|---|---|---|---|---|
| Legon (greenhouse) | 24.83–27.78 | 27.19–33.30 | 92.80–100.00 | 65.95–89.86 |
| Ze (open field) | 25.3–32.00 | 27.60–33.70 | 53.50–95.50 | 42.00–82.00 |
| Table | NIP | NFI | NFP | FW | FWP | FS |
|---|---|---|---|---|---|---|
| 41,176 | 3.39 | 684.48 | 716.10 | 50,747 | 118.23 | |
| 12,890 | 0.37 | 616.71 | 147.30 | 113,896 | 101.73 | |
| 6,607 | 0.81 | 216.67 | 207.10 | 61,459 | 64.20 | |
| Broad sense heritability | 0.84 | 0.90 | 0.65 | 0.85 | 0.41 | 0.64 |
| Traits | Cluster 1 (Highly Tolerant) | Cluster 2 (Moderately Tolerant) | Cluster 3 (Moderately Sensitive) | Cluster 4 (Highly Sensitive) | Cluster 5 (S. pimpinellifolium, Highly Sensitive) | |
|---|---|---|---|---|---|---|
| Number of flowers per inflorescence | 6.00 ± 0.00 | 7.00 ± 1.00 | 5.00 ± 1.00 | 4.00 ± 1.00 | 11.00 ± 0.00 | |
| Fruit set percentage (%) | 47.52 ± 5.21 | 27.60 ± 13.37 | 20.17 ± 5.32 | 11.15 ± 6.39 | 1.78 ± 0.70 | |
| Phenotypic parameters | Individual fruit weight (g) | 11.60 ± 1.82 | 27.13 ± 7.50 | 36.08 ± 12.84 | 61.00 ± 30.56 | 0.69 ± 0.36 |
| Fruit weight per plant (g) | 1347.05 ± 275.92 | 537.11 ± 204.58 | 432.99 ± 224.39 | 300.13 ± 206.13 | 9.21 ± 0.14 | |
| Number of inflorescences per plant | 121.00 ± 47.00 | 39.00 ± 19.00 | 36.00 ± 7.00 | 32.00 ± 8.00 | 921 ± 62 | |
| Number of fruits per plant | 114.00 ± 15.00 | 23.00 ± 15.00 | 14.00 ± 4.00 | 6.00 ± 4.00 | 15.00 ± 7.00 | |
| O.HET | 0.01 | 0.01 | 0.01 | 0.01 | 0.04 | |
| Genotypic parameters | E.HET | 0.04 | 0.06 | 0.08 | 0.06 | 0.14 |
| MAF | 0.03 | 0.04 | 0.06 | 0.04 | 0.13 | |
| PIC | 0.03 | 0.05 | 0.07 | 0.05 | 0.11 |
| Cluster 1 | Cluster 2 | Cluster 3 | Cluster 4 | |
|---|---|---|---|---|
| Cluster 2 | 0.10 | |||
| Cluster 3 | 0.41 | 0.26 | ||
| Cluster 4 | 0.20 | 0.14 | 0.41 | |
| Cluster 5 | 0.74 | 0.79 | 0.75 | 0.83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayenan, M.A.T.; Danquah, A.; Agre, P.A.; Hanson, P.; Asante, I.K.; Danquah, E.Y. Genomic and Phenotypic Diversity of Cultivated and Wild Tomatoes with Varying Levels of Heat Tolerance. Genes 2021, 12, 503. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12040503
Ayenan MAT, Danquah A, Agre PA, Hanson P, Asante IK, Danquah EY. Genomic and Phenotypic Diversity of Cultivated and Wild Tomatoes with Varying Levels of Heat Tolerance. Genes. 2021; 12(4):503. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12040503
Chicago/Turabian StyleAyenan, Mathieu Anatole Tele, Agyemang Danquah, Paterne A. Agre, Peter Hanson, Isaac Kwadwo Asante, and Eric Yirenkyi Danquah. 2021. "Genomic and Phenotypic Diversity of Cultivated and Wild Tomatoes with Varying Levels of Heat Tolerance" Genes 12, no. 4: 503. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12040503