
Neuromesodermal Lineage Contribution to CNS Development in Invertebrate and Vertebrate Chordates


Laboratoire de Biologie du Développement de Villefranche-sur-mer (LBDV), Institut de la Mer de Villefranche, Sorbonne Université, CNRS, 06230 Villefranche-sur-mer, France
*
Author to whom correspondence should be addressed.
Genes 2021, 12(4), 592; https://0-doi-org.brum.beds.ac.uk/10.3390/genes12040592
Submission received: 20 February 2021
/
Revised: 12 April 2021
/
Accepted: 13 April 2021
/
Published: 17 April 2021
(This article belongs to the Special Issue The Evolution of Invertebrate Animals)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Ascidians are invertebrate chordates and the closest living relative to vertebrates. In ascidian embryos a large part of the central nervous system arises from cells associated with mesoderm rather than ectoderm lineages. This seems at odds with the traditional view of vertebrate nervous system development which was thought to be induced from ectoderm cells, initially with anterior character and later transformed by posteriorizing signals, to generate the entire anterior-posterior axis of the central nervous system. Recent advances in vertebrate developmental biology, however, show that much of the posterior central nervous system, or spinal cord, in fact arises from cells that share a common origin with mesoderm. This indicates a conserved role for bi-potential neuromesoderm precursors in chordate CNS formation. However, the boundary between neural tissue arising from these distinct neural lineages does not appear to be fixed, which leads to the notion that anterior-posterior patterning and neural fate formation can evolve independently.
1. Introduction
Ascidians are marine invertebrate chordates that are the closest living relatives to the vertebrates [1]. This review is concerned with the tadpole larvae of solitary species, predominantly Halocynthia roretzi and Ciona sp. (Ciona intestinalis, Ciona robusta and Ciona savigni, which will be discussed as “Ciona”). Halocynthia and Ciona belong to different orders of ascidians, but they develop in an almost identical manner in terms of cell lineages and embryo anatomy and morphology even if not necessarily via conserved developmental molecular mechanisms [2,3,4,5,6,7,8,9,10]. At larval stages of development, ascidians possess a notochord and a dorsal neural tube, both synapomorphies of chordates [11]. Ascidian embryos develop with a fixed cell lineage [5]. Most cells are fate-restricted at the onset of gastrulation, when the embryo consists of 112 cells. The larva is composed of in the region of 3000 cells [12]. No stem cells are involved in generating the larval body and the tail elongates by a process of convergence and extension [13,14].
A long-held traditional view of vertebrate CNS formation is that neural tissue is induced from the ectoderm germ layer in a process called “neural induction”, cells choosing between neural and epidermis. This neural induction step induces neural fate with anterior character, part of which is then “transformed” into posterior fates by posteriorizing signals (reviewed in [15,16]). This seemed at odds with the observation in ascidian larvae that the anterior and posterior CNS arise from distinct hemispheres of the early embryo, with animal hemisphere-derived anterior neural tissue arising from binary fate choices with epidermis and the vegetal hemisphere-derived posterior CNS arising from binary fate choices with mesoderm. However, in vertebrates, while some posterior neural tissue does form via a process known as “posteriorization”, it has become clear that the posterior-most CNS originates from a distinct ontology to the anterior CNS and is associated with mesoderm fates [15,17,18,19]. These recent advances may bring the ontologies of the ascidian and vertebrate CNSs closer together than was once believed. Throughout this review we will adopt the following definitions of bi-fated and bi-potential [20]. Bi-fated precursors are those that generate two tissue types (neural and mesoderm, for example), whereas mono-fated implies lineage restriction of a cell to a specific tissue type. This definition does not necessarily imply any type of commitment of cells to a particular fate [21]. We refer to bi-potential precursors as those that can adopt one of two alternative fates based on the signals they receive; i.e., they are competent to form the two cell types, depending upon their environment. This does not rule out the possibility that the local embryonic environment may impose mono-fated development from precursors that are nonetheless bi-potential, with bi-potentiality only being revealed by experimental manipulation. In this review, we provide a detailed description of the embryonic origin of the ascidian larval neuromesoderm-derived CNS and discuss to what extent this is similar to the situation in vertebrates, with particular focus on mice, chick and zebrafish as vertebrate examples.
2. The Ascidian Larval CNS
The ascidian larval neural tube forms through the rolling up of the neural plate, similarly to primary neurulation in vertebrates [14,22,23]. The ascidian larval CNS has been extensively studied in terms of its lineage, molecular mechanisms of development, gene expression patterns and cell types [23,24,25,26,27]. More recently, a larval nervous system connectome has been revealed [28] and collections of single cell transcriptomics generated [29,30,31,32,33]. Emerging studies are now aiming to annotate each neuron identified in the connectome with the gene expression patterns, including neurotransmitter types, described from in situ hybridization/immunostaining analysis or single cell transcriptomics datasets, as well as to the cell lineage history and ultimately to neuronal function and larval behavior [29,34,35,36,37,38,39].
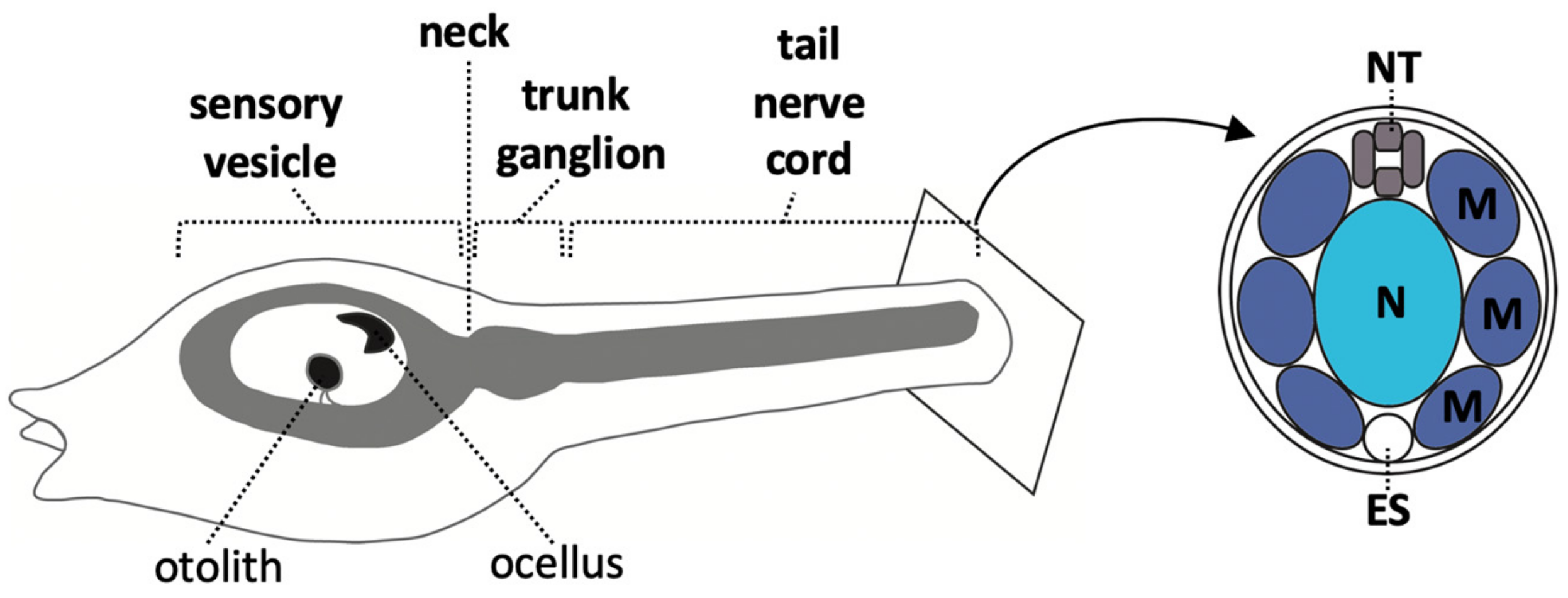
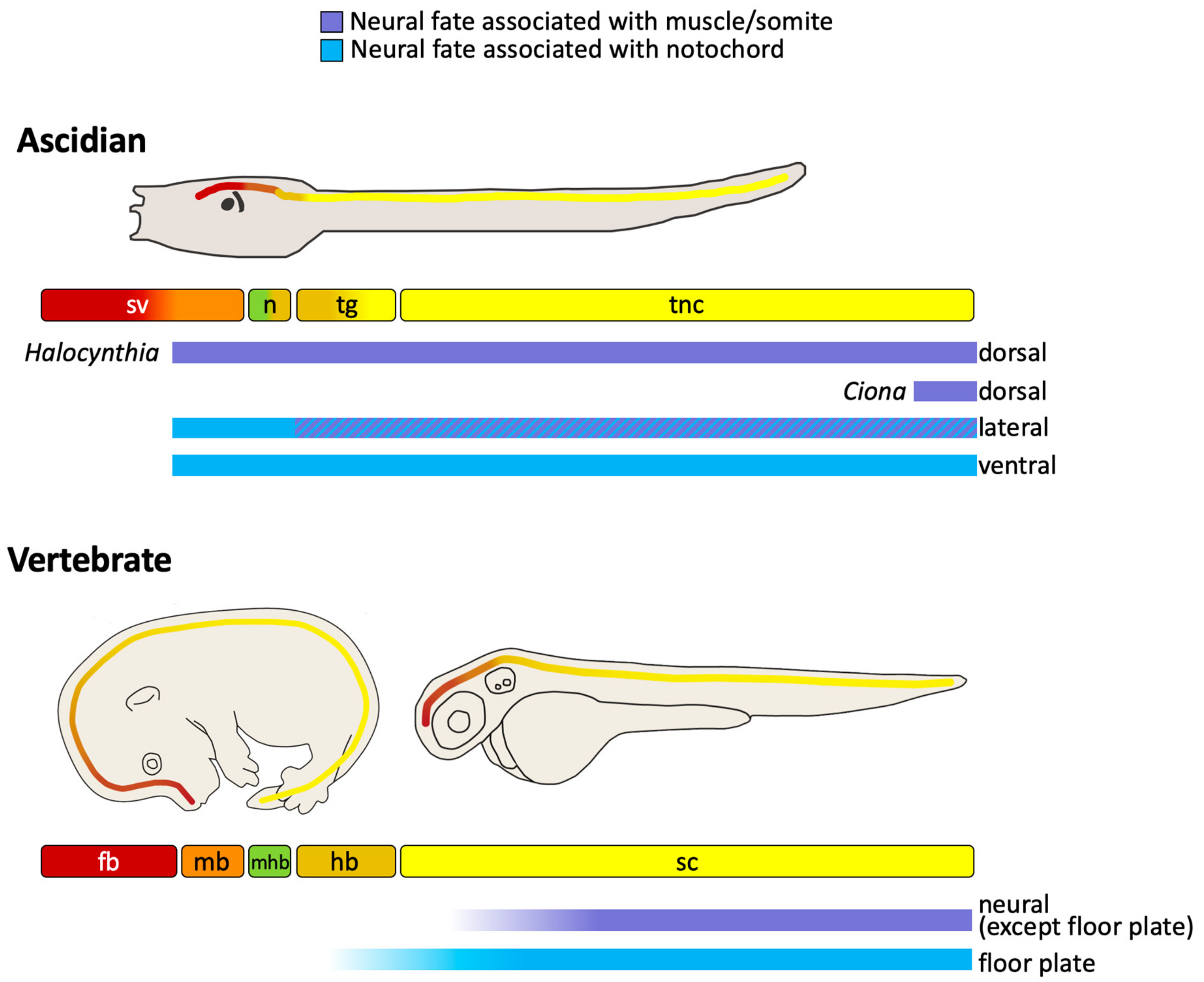
At larvae stages, the ascidian CNS has distinct morphological units which are, from anterior to posterior, the sensory vesicle, a narrow ‘neck’ region, a trunk ganglion, and a tail ‘nerve cord’ (Figure 1) [14]. In the literature, the posterior part of the sensory vesicle is sometimes referred to as the posterior brain [23,24,40] or brain stem [8]. The sensory vesicle contains two pigmented sense organs, an otolith, anchored in the ventral wall and used for gravity perception, and an ocellus, required for photo-reception [24,25,41,42,43]. The trunk ganglion [44] is also referred to as the visceral ganglion [24,45] or motor ganglion, since it contains the motor neurons that innervate the tail muscle [23,28,46,47,48]. Here we use the morphology-based term, ‘trunk ganglion’. The nerve cord is the posterior-most part of the CNS; a tube consisting of only 4 cells in cross section, one dorsal, one ventral and two lateral, though nonetheless, it has recently been shown to harbor neurons [28,47,49,50]. The ascidian larvae CNS is made up of approximately 330 cells of which at least 177 are neurons, located in the sensory vesicle (143), trunk ganglion (25) and tail nerve cord (up to approximately 20) [27,28,47,49,51]. Transcriptomic analysis has identified 41 distinct neural subtypes in larva [29]. For a description of some of these neural cell types, see recent reviews [24,25,27].
Aside from the neurons, the non-neuronal ependymal cells of the CNS (apart from those residing in the tail nerve cord) form an important stem cell-like source that will later give rise to the post-metamorphosis adult nervous system [40,44]. Some larval neurons also persist after metamorphosis, although many others are lost during the process [40].
3. Anterior-Posterior Regionalization of the Chordate Neural Tube
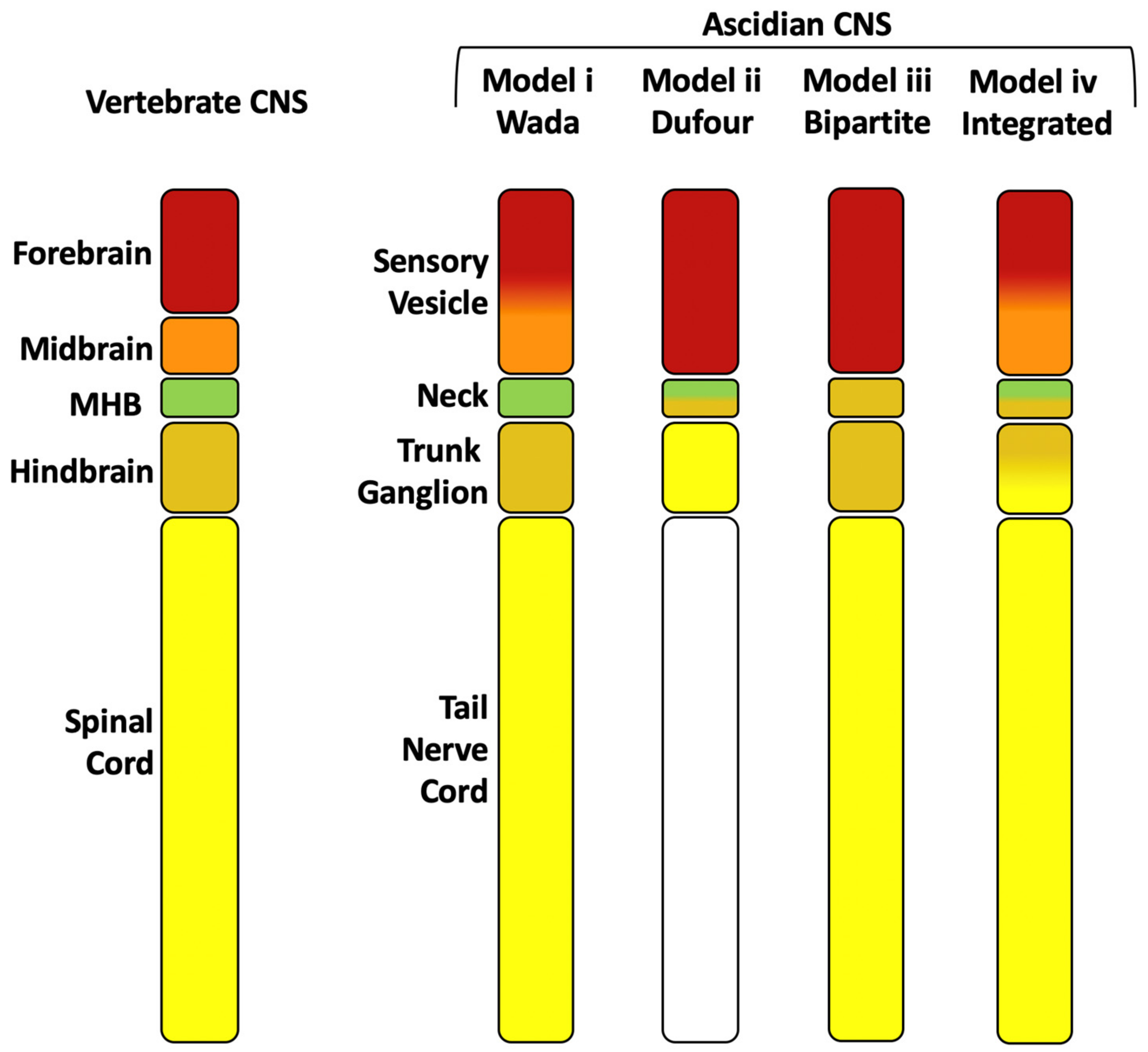
Regional comparisons between ascidian larval and vertebrate CNSs are complicated by a significant simplification of the ascidian CNS that most likely took place in the tunicate lineage [23,52,53,54,55], whereas the vertebrate CNS has evolved more complex structures [55,56]. Nonetheless, comparisons of gene expression profiles, neural function and connectivity along the anterior-posterior axis of the CNS between ascidians and vertebrates can go some way towards providing equivalence between the different domains and have led to several hypothesis (Figure 2) [25,26,44,54,57,58,59,60,61,62,63,64]. At the anterior, the Otx positive domain, or sensory vesicle, likely corresponds to the vertebrate forebrain. Within the Otx positive domain, the presence or absence of a midbrain has been the subject of much debate (Figure 2, models ii and iii have no midbrain equivalent) [26,54,57,62]. However, recent evidence on the function of the posterior sensory vesicle as a sensory processing and integration center, as well as gene expression patterns and neuronal circuitry has suggested that part of the posterior sensory vesicle could be homologous to the vertebrate midbrain [34] (Figure 2, model iv). Between the sensory vesicle and trunk ganglion, the small intervening neck region (not morphologically recognizable in Halocynthia but identifiable by gene expression) of only 6 cells [51], is difficult to assign to a specific vertebrate counterpart, which is further confounded by dynamic gene expression in this region [58]. At late tailbud stages, however, the neck region is neither Otx nor HOX positive [58,64]. This small region may therefore correspond to the mid-hindbrain boundary (MHB; Figure 2 models i, ii) although the presence of a MHB in tunicates is also debated (Figure 2, model iii) and if present, it does not appear to have an organizer function as it does in vertebrates [26,54,57,58,59,63,64]. There is also evidence that the neck region may, in part, correspond to the hindbrain, since some cells labelled by Phox, located in this region, give rise to the visceral motoneurons of the adult, a neuronal cell type that arises in the hindbrain of vertebrates (Figure 2 model ii) [44].
The trunk ganglion contains the motor neurons that control swimming behavior [24,47,65,66] and expresses HOX genes, and has been compared to either the spinal cord (Figure 2 model ii) [44,50,65] or hindbrain (Figure 2, model i, iii) [57,62,63]. A specific gene expression pattern of transcription factors along the anterior-posterior axis of 5 pairs of cholinergic neurons (descending decussating neuron (ddN), interneurons and motoneurons) of the trunk ganglion, which is conserved among different ascidian species, has been compared to a similar order of gene expression along the dorsal-ventral axis of the vertebrate spinal cord [7,67]. Together with the ascending commissural inhibitory interneurons (ACINs) in the anterior nerve cord, the cholinergic motoneurons of the trunk ganglion are comparable to the vertebrate spinal cord central pattern generators that generate rhythmic swimming behavior [65]. However, the most anterior of the trunk ganglion cholinergic neurons, a pair of descending decussating neurons (ddNs), have been likened to the Mauthner cells of tailed vertebrates, neurons which arise in the hindbrain and are involved in the startle response network [68]. At midgastrula stages, FGF8/17/18 is expressed in lateral trunk ganglion precursors just posterior to the neck precursors and it is required for the specification of the neck region itself, which otherwise adopts a posterior sensory vesicle-like fate [26]. This result could be interpreted as a posterior shift in MHB FGF-signaling in the ascidian larval CNS relative to the vertebrate CNS [26,58]. Alternatively, the FGF8/17/18-positive region could be similar to the hindbrain FGF-signaling activity located at rhombomere 4 of vertebrate embryos [57,69,70,71]. We conclude that the trunk ganglion may have homology to both the hindbrain, at least at its anterior end (Mauthner-like cells), and the spinal cord (somatic motoneurons of the central pattern generator) (Figure 2, model iv).
The tail nerve cord is likely to be homologous to the vertebrate spinal cord [57,64] (Figure 2 model i, iii, iv). While it was originally believed that the tail nerve cord was devoid of neurons and was thus removed from any comparative analysis in model ii [44], we now know that neurons are present in the tail nerve cord [28,36,47,65]. The anterior part of the tail nerve cord contains motoneurons (4 cells) and interneurons (2 cells) [27] and the ACINs (usually 3–4 cells) [28,36,47,65,68], that are involved swimming behaviors [47,65,68]. Other cholinergic mid-tail motoneurons or ‘planate neurons’ are located along the tail nerve cord [27,47,68] of which 12-15 cells have been observed [47].
We propose a model (Figure 2, model iv) whereby the sensory vesicle represents the vertebrate Otx positive ‘forebrain’ and ‘midbrain’, with the midbrain corresponding to part of the posterior sensory vesicle [34], the intervening neck region, which is Otx- and HOX-negative at some stages, representing the ‘mid-hindbrain’ and ‘hindbrain’, the trunk ganglion representing a ‘hindbrain-spinal cord’ region and the remaining tail nerve cord, with its neurons, corresponding to the rest of the ‘spinal cord’. This model (iv) fits a tripartite organization, with an Otx positive anterior domain, a posterior HOX positive domain with an Otx/HOX negative MHB domain in between [25,44,58,61,63,64]. However, it has also been suggested that the ascidian CNS could instead be considered bipartite, with only a forebrain and a hindbrain/spinal cord, and lacking a potential MHB domain (Figure 2 model iii) [54,57,58]. It is likely that the debate will continue and boundaries will be refined as more data on neural function and gene expression is acquired.
4. A Large Part of the Ascidian CNS Is Derived from Bi-Fated Neuromesodermal Precursors
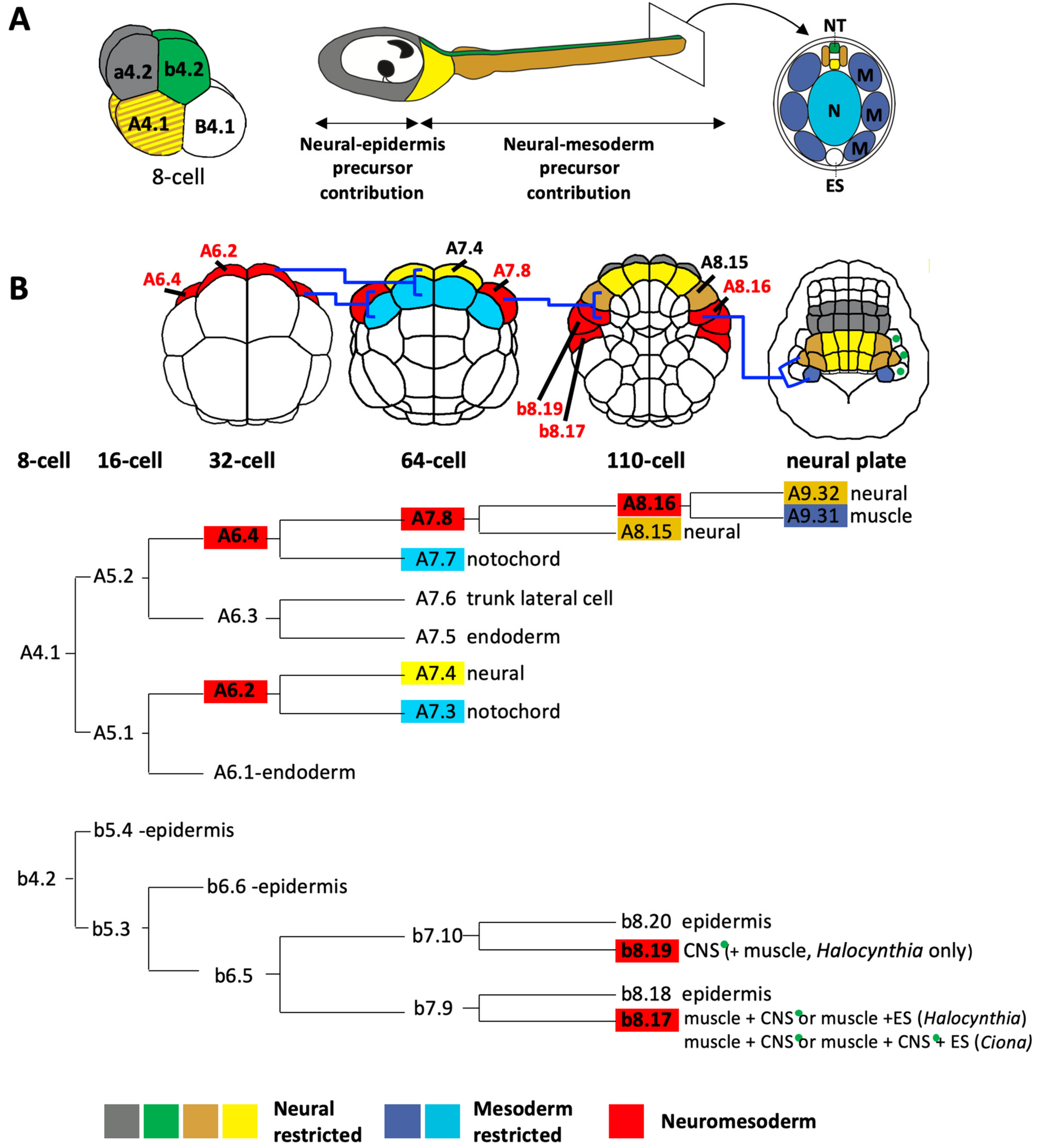
Ascidian embryos develop with a fixed cell cleavage pattern and lineage, allowing the exact lineage relationships between different cell types to be known as long as the cells can be traced and identified [3,8,13,26,37,38,50,58]. The embryos are bilaterally symmetrical so each cell name refers to a pair of cells [72]. At the 8-cell stage of development, the embryo divides along the animal-vegetal axis to generate the four founder lineages. The four animal cells, the a4.2 and b4.2 cell pairs, generate the a- and b-lineages and the four vegetal cells, the A4.1 and B4.1 cell pairs, generate the A- and B-lineages (Figure 3A). a- and b-lineages give rise to predominantly ectoderm: all of the epidermis and the part of the CNS [8,14,45,73]. The vegetal lineages (A- and B-) will generate most of the endoderm and mesoderm but, importantly, a large part of the posterior CNS is also derived from the A-lineages (Figure 3A) [8,14,45,73].
The following description of the neural lineages is based on a number of published studies [8,13,45,74,75]. The a-lineages will generate the anterior part of the sensory vesicle (Figure 3A, grey). The process of neural induction in a- (and b-) line cells begins at the 32-cell stage with induction of the Otx gene in two pairs of neural lineage precursors, following a cell fate choice between neural and epidermal tissue [76,77,78]. At the early gastrula stage the a-lineage neural precursors segregate further into anterior sensory vesicle precursors and, more anteriorly, a specialized region of anterior ectoderm that has been likened to a proto-placodal territory that generates the palps (a sensory adhesive organ) as well as certain epidermal sensory neurons [30,74,79,80]. Manipulating the inducing signal or transcription factor expression during these cell fate decisions suggests that they operate in a binary mode: epidermal vs neural, CNS vs sensory placode [78,81,82,83]. Interestingly, promotion of neural fates in a-lineages involves similar factors (the transcription factors Foxa.a and Zic.r.b and FGF-signaling) to those promoting notochord fates in vegetally-derived A-lineages, but the order in which these factors become active is not the same in the two lineages. Experimentally-induced overlapping expression of Foxa.a and Zic.r.b in a-lineages, mimicking the situation in the notochord lineage, results in a mixed neural and notochord regulatory state in a-lineage neural precursors at early gastrula stages and notochord gene expression extending into the head at tailbud stages [84]. Thus, while the same factors are required for specification of notochord and anterior neural fate, the different temporal sequence of expression of these factors in the a-lineage cells is sufficient to promote anterior neural (‘brain’) fate and to prevent these cells from adopting axial mesoderm fate. More details on the molecular mechanisms governing these neural fate choices can be found in recent reviews [25,60,85].
The b-lineages contribute to the dorsal most row of cells in the posterior part of the CNS (Figure 3A, green cells). Although b-epidermal lineages have been shown to generate neurons of the peripheral nervous system [39,86], there are as yet no reported CNS neurons arising from these lineages. The b-neural lineage precursors arise in the lateral part of the embryo and also contribute to ectoderm, mesoderm and endoderm tissues, though there are difference between Halocynthia and Ciona (Figure 3 and Figure 4) [8]. Epidermis and neuromesodermal fates segregate at the 110-cell (early gastrula) stage of development to give two laterally-positioned neuromesodermal precursors, b8.19 and b8.17 (Figure 3B and Figure 4). In Halocynthia only, b8.19 cells give rise to dorsal neural tube and two muscle cells at the tip of the tail (Figure 3 and Figure 4). In Ciona, this cell generates only dorsal neural tube (thus following a fate choice between neural and epidermis). The b8.17 cell gives rise, in Halocynthia, to 3 muscle cells of the tail as well as either a dorsal neural tube cell or an endodermal strand cell (ventral row of endoderm cells at the tip of the tail), presumably one each from the left or right b8.17 cell (Figure 3 and Figure 4) [8]. In Ciona, each b8.17 gives rise to 2 muscle cells at the tip of the tail and either two cells of the dorsal neural tube or one cell of the dorsal neural tube and one endodermal strand cell (Figure 3 and Figure 4) [8]. This description was based on lineage-tracing. However, by following cells in embryos fixed at sequential stages of development, other authors did not report a contribution of b8.17 to the neural tube in Ciona [14,45]. Therefore, it remains to be confirmed whether or not b8.17 gives rise to the neural tube of Ciona. It is also not clear when the neural tissue segregates from muscle and/or endodermal strand in b-lineage cells.
The A-lineages will generate the lateral and ventral cells of the neural tube from the posterior part of the sensory vesicle to the tail nerve cord (Figure 3, yellow and tan cells). The A-neural lineages are closely associated with mesoderm fates, having segregated from ectoderm lineages at the 8-cell stage of development (Figure 3A). At the 32-cell stage of development, cell division segregates endoderm from the bi-fated neuromesoderm precursors, A6.2 and A6.4, positioned at the anterior margin (Figure 3B, red cells of the 32-cell stage embryo; Figure 4). These neuromesoderm precursors will generate mesoderm (mostly notochord, but also some posterior tail muscle arises from A6.4) and neural tissue (Figure 3 and Figure 4). At the 64-cell stage, A6.2 and A6.4 divide along the animal-vegetal axis segregating notochord (Figure 3B, light blue cells at the 64-cell stage) from neural/muscle fates (yellow and red cells in Figure 3B, 64-cell stage). This ‘notochord-neural’ fate segregation operates as a binary fate choice and thus these precursors can be considered as bi-fated and bi-potential [87,88]. The medial A6.2 cell generates, at the 64-cell stage, a notochord-restricted precursor and a neural-restricted precursor (A7.4, yellow) at the margin (Figure 3B). A7.4 generates the lateral and ventral posterior sensory vesicle and neck as well as the ventral row of cells in the trunk ganglion and tail nerve cord. The lateral A6.4 cells each generate, at the 64-cell stage, a notochord-restricted cell, A7.7 (Figure 3B, blue cell on 64-cell stage embryo) and a neural-muscle precursor, A7.8 (Figure 3B, red cell on 64-cell stage embryo). A7.8 subsequently divides along the medial-lateral axis to give rise to A8.15 and A8.16 (Figure 3B). A8.15 is a mono-fated neural precursor and will generate lateral cells of the trunk ganglion and tail nerve cord. A8.16 is a bi-fated neural-muscle precursor (Figure 3 and Figure 4). At neural plate stages, A8.16 divides along the anterior-posterior axis to generate one neural and one muscle precursor (Figure 3B, tan and dark blue cells at neural plate stage), via a binary fate switch (shown in Ciona [89]). The A-lineages are identical between the two ascidians studied, Halocynthia and Ciona, despite the observation that the molecular mechanisms specifying the muscle cells from the A-lineages has diverged [4,8,75,90,91]. The molecular mechanisms of neuro-mesoderm specification in the A-lineages will be described later (Section 6). In conclusion, a large part of the ascidian neural tube has a neuromesoderm origin.
Appendicularians (or larvaceans) belong to another order of tunicates and exhibit a tadpole body form throughout their adult lives. The cell lineages have been described and fate restriction takes place even earlier than in ascidians. Similar to ascidians, both nervous system and muscle derive from animal and vegetal hemispheres, with a large part of the CNS associated with mesendodermal (including notochord) vegetal lineages, although in appendicularians neural tissue segregates earlier from mesendoderm and there is no neuromesoderm intermediate [92,93].
5. Evidence of Neural-Mesoderm Lineages in Vertebrates
In vertebrates, it was believed for many years that the nervous system was induced with anterior character, following a fate choice between neural and epidermal fates. This anterior character was then ‘transformed’ by posteriorizing signals to generate the entire posterior nervous system (reviewed in [15,16]). The prevailing modern view is that only the anterior nervous system: forebrain, midbrain, hindbrain and anterior part of the spinal cord is induced and transformed from a cellular origin associated with epidermis [19,20,94,95,96]. The posterior part of the nervous system is now thought to arise predominantly from neural-mesoderm precursors (NMps) that generate the paraxial mesoderm and spinal cord [15,17,18,20,95,97,98,99,100,101]. Another neuromesodermal contribution to the vertebrate CNS is from notochord-floor plate precursors. Each of these contributions to the vertebrate CNS will be discussed in turn.
5.1. Neural-Somite Fate Choices
Much of the vertebrate spinal cord is believed to arise from bi-potential precursors, termed neural-mesoderm precursors (NMps), which choose between spinal cord and paraxial mesoderm (plus vasculature) fates [15,17,18,98,100,101]. Early indications for the existence of bi-potential neuromesoderm precursors came from fate mapping and mutant analysis. Fate mapping at early somite stages in mice and chick, revealed that some paraxial mesoderm and neural tissue arose from a similar location, at the node/streak border and caudal lateral epiblast and later in the caudal neural hinge of the tailbud [102,103,104,105,106]. In zebrafish, single-cell fate mapping identified bi-fated neural-muscle precursors at the late blastula stage, which segregate into mono-fated precursors by the early gastrula stage [20,107].
Analysis of mutant cells in mice also indicated a close relationship and potential bi-potentiality between paraxial mesoderm and spinal cord. Examples include FgfR1-mutant cells failing to pass through the primitive streak to form mesoderm and instead forming secondary neural tubes [108]; Wnt-3a mutant cells that ingress through the primitive streak, but fail to migrate laterally to form somites, instead giving rise to secondary neural tubes [109]; and Tbx6 mutants in which posterior somites are converted into supernumerary neural tubes [110]. The interactions between these and other factors and how they drive paraxial mesoderm versus spinal cord fate are now well understood and appear to be conserved among vertebrates [15,17,18,97,98,111,112,113,114,115].
Clonal analysis in mice proved unequivocally that bi-fated neuromesoderm precursors contribute to the neural tube [96]. In mice, while segregation of neural from ectoderm takes place early in development, self-renewing neuromesoderm precursors (neural-somite) continuously contribute to the posterior nervous system (spinal cord) during body axis elongation [96]. Retrospective clonal analysis in zebrafish also revealed that mid- and posterior spinal cord has a closer clonal relationship to muscles derived from paraxial mesoderm than it does to anterior nervous system, strongly arguing for a neural-mesoderm fate choice at the base of the generation of a large part of the spinal cord [20]. Unlike in mice in which neuromesoderm precursors exhibit self-renewing properties during the generation of the spinal cord, in zebrafish, much of the segregation of neural from paraxial mesoderm fates appears to take place earlier, during early gastrulation [20,107]. Resultant mono-fated pools of spinal cord or somite precursors nonetheless retain the capacity to change developmental status between neural and mesoderm, depending on Wnt signaling in the tailbud [100]. Thus, at least in the tailbud, these cells can be considered to remain bi-potential. A small population of NMps retained in the tailbud are largely quiescent during somitogenesis stages and are thought to contribute to only last region of the body axis [20,100,116].
Bi-potential NMps have been successfully created from mouse embryonic stem cells by recreating embryonic conditions (Wnt plus FGF signaling), demonstrating that precursors of posterior spinal cord and paraxial mesoderm pass through a bi-potential state [97]. Anterior neural tissue up to hindbrain levels follows a distinct transcriptional trajectory compared to NMps that generate spinal cord, further confirming the different origins of anterior and posterior nervous system [97]. Furthermore, evidence suggests that anterior and posterior regional identity is imposed on cells prior to their acquisition of neural identity [95]. Specific anterior and posterior enhancers of the pan-neural Sox2 gene in chick and mice provided additional evidence for distinct mechanisms controlling anterior and posterior CNS development [18].
Thus, in vertebrates, at least the posterior spinal cord is generated with a significant contribution from cells that choose between neural and mesoderm (somite) fates [15,18,20,95,101]. The precise extent of the NMp contribution along the anterior-posterior axis of the neural tube is not yet clear, as discussed by Henrique et al., [15].
5.2. Neural-Notochord Fate Choices
There is also evidence for another type of neuromesoderm precursor, which generates the ventral-most part of the neural tube, in particular, the medial floor plate, and notochord in vertebrates. The vertebrate floor plate consists of medial and lateral cell types, which share the expression of a subset of genes [117,118,119,120]. It is the medial floor plate that is associated with notochord fates.
Fate mapping in mice [121,122], chick [105,117] and zebrafish [123,124,125,126,127,128,129] show that floor plate precursors reside in the organizer (early gastrula organizer, node, shield), in a similar region to notochord precursors and in a distinct location compared to those of the rest of the spinal cord.
In chick, bi-fated notochord-floor plate precursors have been identified in the node by single cell labelling [130]. In zebrafish, bi-fated notochord-ventral neural precursors were also found at blastula stages although it is unlikely that these cells are committed in any way [21,128,131]. By the early gastrula stage in zebrafish, notochord and floor plate precursors have segregated into distinct regions of the shield [107,127,128,131,132]. Specification of floor plate identity begins early, during gastrulation, with floor plate character actively maintained in the tailbud at later stages [123,133,134,135]. In mice, fate segregated (i.e., mono-fated) notochord and floor plate precursor pools occupy different regions of the node [122,136,137]. This is consistent with the observation that retrospective lineage tracing in mice identified only one clone contributing to notochord and neural cells (as well as posterior paraxial mesoderm) [96] and suggest that notochord-neural precursors are either very rare or segregate early during development and do not exist as a bi-fated precursor pool.
Evidence for bi-potential notochord-neural precursors can be found under certain conditions in mice, chick and zebrafish. In both chick and zebrafish, mutations or manipulation of signaling pathways result in fate switches between notochord and floor plate fates revealing the presence of bi-potential floor plate-notochord precursors under these conditions [107,132,133,138,139,140]. For example, several mutations effecting notochord formation in zebrafish result in larger medial floor plates in the hindbrain and spinal cord [118,120,123,132,139,140,141,142]. Strikingly, while wild type blastula cells transplanted into the dorsal margin cells of a wild type host generate predominantly notochord, transplantation of ntl (brachyury, a key notochord determinant) mutant cells under the same conditions results in a switch from a notochord to a floor plate contribution [132]. Similarly, in chick, manipulation of the Notch signaling pathway results in notochord-floor plate fate transformations at the level of the spinal cord and posterior hindbrain [138]. In mouse early somite stage embryos (E8.5), transplantation of the rostral node, which gives rise exclusively to notochord, into the border of the primitive streak (that usually produces ventral neural tube and paraxial mesoderm) converts the rostral node notochord precursors into a ventral neural tube fate (not restricted to floor plate) [104]. Transplantation of rostral node cells into the anterior primitive streak, which normally generates paraxial mesoderm and tail bud mesoderm, also converted the rostral node cells from notochord to neural fates. Thus, the rostral node cells can switch from notochord to neural fate even if they do not generate neural fates under normal developmental conditions [104].
Along the anterior-posterior axis, the floor plate cells appear to arise from distinct origins and by distinct mechanisms [119]. In mice, the ventral midline of the fore and midbrain is associated with organizer tissue that arises prior to node formation (early and mid-gastrula organizers, which also generates head mesendoderm) [121] and Shh (Sonic hedgehog) expression in anterior and posterior floor plate are controlled by distinct enhancers [143]. In chick embryos, anterior floor plate (from hindbrain levels) is induced by prechordal mesoderm in a region of epiblast just anterior to the node called ‘area a’; a distinct population of cells compared to more posterior floor plate precursors that arise together with notochord within the node itself [144]. The allocation of cells to anterior and posterior floor plate does not appear to be absolute and is not yet fully understood [119].
In zebrafish, anterior and posterior floor plate expression of Shh is also driven by distinct enhancers [120] and mutations differentially affect either the entire floor plate or specifically the posterior floor plate, suggesting mechanistic differences along the anterior-posterior axis [120,124,132,139]. For example, mutations of the notochord-promoting T-box factor, ntl (Brachyury), are thought to result in conversion of notochord precursors to a floor plate fate [132,139]. Ntl mutations can rescue posterior medial floor plate (MFP) up to the hindbrain, in a Nodal (cyc) mutant background, which would otherwise have no MFP along the entire anterior-posterior axis [139]. Thus, expression of ntl can govern the bi-potentiality of notochord precursors to choose between notochord and floor plate fate, at least in the posterior neural tube of zebrafish. We can conclude that bi-fated and bi-potential notochord-neural (floor plate) cells exist in vertebrate embryos. Exactly when these different fates become committed and the exact extent of their anterior-posterior contribution is not entirely resolved [119,120]. Nonetheless, from the available evidence we can conclude that vertebrate notochord-floor plate precursors contribute to at least the posterior part of the neural tube.
5.3. A Note on the Historical Controversy Surrounding Floor Plate Induction in Chick
There has been some controversy surrounding the formation of the floor plate in chick. Whether floor plate cells are generated as a specific precursor and laid down in the ventral neural tube or whether floor plate is induced in the ventral neural tube has been the subject of much debate [145,146]. Nonetheless it seems now to be accepted that, at least the developmental origin of medial floor plate cells is distinct from those of the rest of the neural tube, and is closely associated with axial mesoderm [120,145,146]. The main contention appears to be whether the cells that share a common origin with notochord are laid down and then induced to form floorplate or whether they are laid down already with floor plate character [120,145,146]. Part of this confusion is likely to have stemmed from the plasticity within the neural tube to form floor plate (e.g., [117,138]) and the fact that there are two types of floor plate, medial and lateral, of which only the medial cells share a common embryonic origin with axial mesoderm [117,118,120].
6. Molecular Mechanisms of Neuromesoderm Fate Segregation in Ascidians
The molecular mechanisms of neural fate segregation are well described in ascidians, particularly in a- and A-lineage [25,60,85]. Despite the similar final body plans shared between ascidian and vertebrate larvae, they often do not appear to develop using the same molecular strategies [90]. Even among ascidian species, the molecular mechanisms required to determine equivalent cell types in equivalent positions can be remarkably divergent [4,5,7,10,91,147,148,149]. In the following discussion, where differences are found between Ciona and Halocynthia, the Ciona data is described. The C. robusta unique gene model IDs for genes discussed in this section are provided at the end of the review (Section 9).
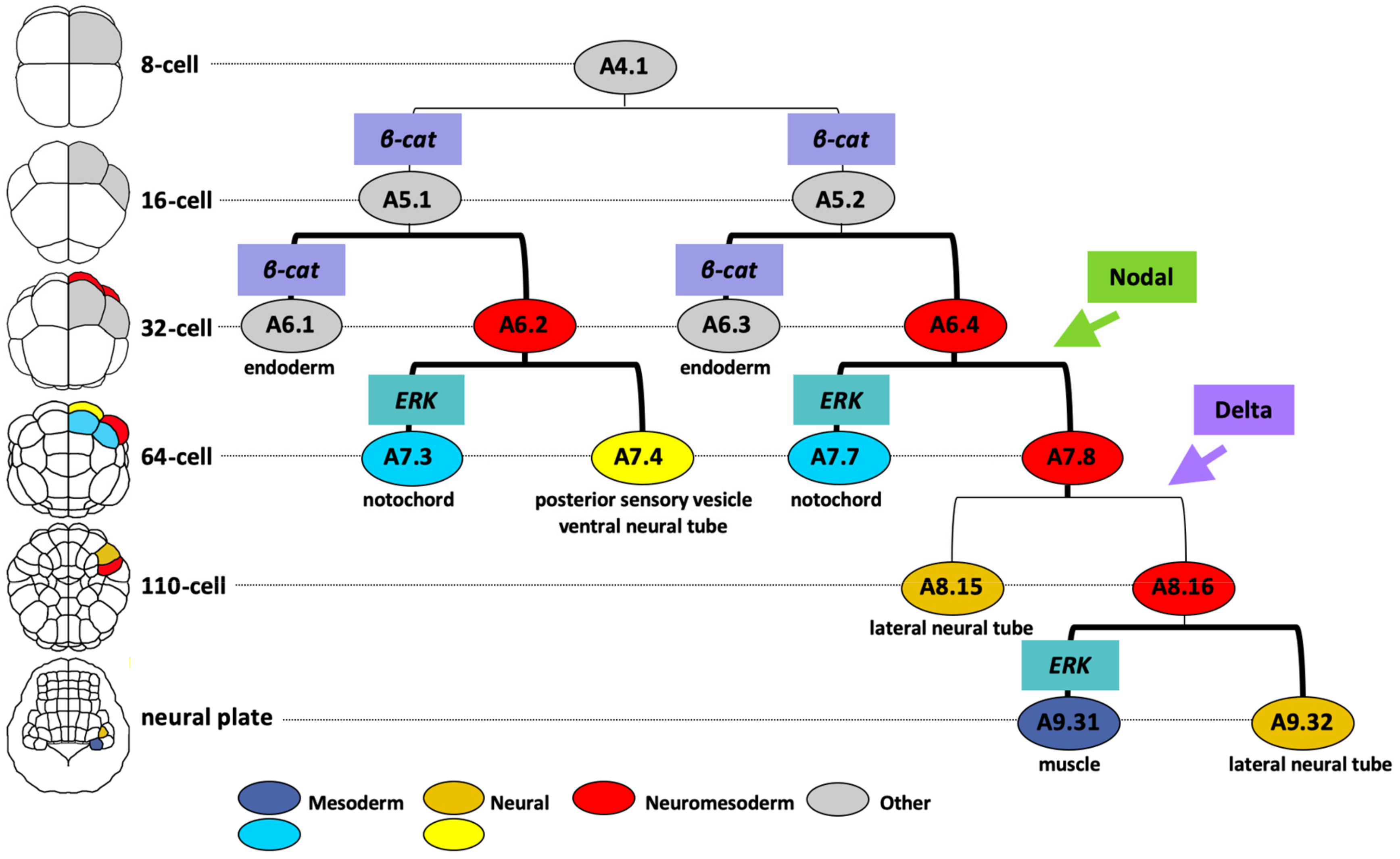
The main neuromesodermal lineage is the A-lineage, arising from the A4.1 blastomere pair at the 8-cell stage of development. A brief overview of the molecular mechanisms leading to neural segregation from the A-lineage is shown in Figure 5. Between the 8- and 16- cell stage β-catenin/TCF transcriptional activities are detected in the vegetal cells, including A5.1 and A5.2 of the A-lineage [150]. β-catenin is required in A5.1 and A5.2 for the specification of endoderm and margin (mesoderm/neural) cell identity and for repression of ectoderm fates [147,150,151,152]. The upstream event leading to nuclear translocation of β-catenin is unknown. At the 32-cell stage, differential activation of β-catenin promotes endoderm over notochord-neural (red cells in Figure 5) fates during this binary fate choice [147,152]. The subsequent binary fate choice sees the notochord lineage (blue cells in Figure 5) segregating from the neural lineage (plus some muscle) at the 64-cell stage with ERK activity in the notochord precursors promoting notochord over neural fates [87,88,153]. The differential pattern of ERK activity is driven by FGF9/16/20 (promoting the Ras-ERK cascade) and the ephrin ligand Efna.d (attenuating the cascade) [88,153,154]. Thus, the segregation of A-neural (plus some muscle)-notochord lineages from the 16- to 64-cell stages requires the following sequence of signaling inputs: β-catenin-ON, β-catenin-OFF and then ERK-OFF (for neural, plus some muscle) or ERK-ON (for notochord). Interestingly, in the vegetal hemisphere of the embryo, the arrangement of notochord and neural precursors in the A-lineage cells is mirrored by a similar arrangement of mesenchyme and muscle precursors in the B-lineage cells of the vegetal hemisphere and these fate segregations are both driven by FGF/ERK signaling with FGF required for notochord and mesenchyme fates that segregate from notochord-neural and muscle-mesenchyme precursors respectively [155]. The attribution of notochord/neural fates to the A-lineages and muscle/mesenchyme fates to the B-lineages is itself governed by the inheritance of a maternal determinant macho-1 (later renamed Zic.r.a) to the B-lineages [155]. Removal of macho-1 results in muscle lineages adopting neural fates and mesenchyme lineages adopting notochord fates [155].
Across the medial-lateral axis of the embryo, Nodal- and Delta-like- signals arising from bilaterally-positioned neighboring cells are required to specify the lateral-most cells of the neural lineage from the 32-cell stage and 64-cell stage, respectively (Figure 5) [26,89,156,157,158]. In the absence of Nodal the A7.8 neuromesoderm precursor adopts fates consistent with an A7.4 identity, whereas the absence of Delta-like leads to a loss of A8.16 lineage identity [4,26,89,156]. The final lineage segregation between neural and mesoderm takes place in this lateral cell at the neural plate stage (mid-gastrula) when differential ERK activity specifies mesoderm (muscle) over neural fate (Figure 5) [89]. This final neuromesoderm segregation thus follows the sequence, from the 16-cell to neural plate stage, of β-catenin-ON, β-catenin-OFF, Nodal-ON, ERK-OFF, Delta/Notch-ON and finally ERK-OFF (for neural fate) or ERK-ON (for muscle fate).
The accumulation of β-catenin in the vegetal lineages of early ascidian embryos is an evolutionary conserved mechanism among metazoans for establishing the primary body axis, with β-catenin defining the site where mesendoderm forms and gastrulation begins [159,160]. In ascidians, the A-lineage neuromesoderm precursors arise in this vegetal embryonic domain and require a β-catenin ON to OFF sequence of activation for the initiation of their specification [147]. The a- and b-lineage derived CNS arises in the animal, or β-catenin OFF, embryonic hemisphere.
In ascidians, the specification of the ventral neural tube appears very different compared to floor plate specification in vertebrates, which requires Nodal and SHH (sonic hedgehog) signaling [119,120]. It seems unlikely that Hedgehog plays a similar role in ascidians. Although a hedgehog gene (hh-2) is expressed in the cells of the ventral neural tube (but notably not in the notochord), this expression is late, from tailbud stages, and there is no evidence that it plays a patterning role in the neural tube of ascidians as it does in vertebrates [161,162,163,164,165]. In ascidians, Nodal is also not implicated in notochord-neural specification, but rather patterns the neural plate to specify lateral over medial identities and actually represses the ventral neural and posterior sensory vesicle fate, restricting their formation to the medial neural lineages [156,166,167]. This is in stark contrast to the positive role for Nodal in floor plate development in vertebrates [120].
In vertebrates, an interplay between Wnt-, FGF- and Retinoic acid (RA-) signaling pathways act to both maintain neuromesoderm (neural-somite) progenitors (Brachyury (T)+/Sox2+) and promote their transition to neural (RA) or mesoderm (Wnt, FGF) via promotion of Brachyury (T) and Tbx6 in the mesoderm and Sox 2 in the neural precursors [115]. The Ciona Sox 1/2/3 gene is not specifically expressed in neural tissue at early stages in ascidians [168]. A role for Wnt- and RA- signaling in A-lineage neuromesoderm formation has not been described in Ciona. In Halocynthia, the mechanisms of A-lineage muscle specification involve similar cellular interactions to those observed in Ciona, but employ different signaling molecules. In Halocynthia, Wnt-5 is required for formation of muscle cells from both the neuro-muscle A- and b-lineages, although, at least for the A-lineage, this cellular interaction takes place before gastrulation [4,91].
Some parallels can be drawn from comparisons of FGF-signaling between ascidians and vertebrates. FGF-signals are transiently required for onset of activation of the pan notochord-neural gene Zic-r.b in the neuromesoderm precursors at the 32-cell stage [169]. At each neuromesoderm lineage segregation, FGF/ERK promotes mesoderm over neural fates, similar to some situations in vertebrates, and activates Brachyury and Tbx6 in segregated notochord and A-lineage muscle precursors respectively [88,89,108,115,153,170].
In vertebrates, expression of the posterior promoting transcription factor Cdx is downstream of Wnt [17,171]. However, in ascidians, a combination of ERK activity, Nodal and Delta are required for Cdx expression in the A-lineage derived neural plate at the mid-gastrula stage, in the lateral, posterior-most row of cells, including the muscle precursor after it has segregated from neural lineages [26,89,157]. These cells will give rise to part of the ventral trunk ganglion and tail nerve cord, the lateral tail nerve cord and the A-lineage tail muscle [8,13]. At tailbud stages Cdx is required in the anterior part of the tail nerve cord to repress more anterior gene expression, but does not appear to be upstream of HOX gene expression as is the case in the vertebrate spinal cord [17,26,171]. Its role in A-lineage muscle specification has not been addressed.
Interestingly, despite the absence of self-renewing precursors or so-called ‘posterior growth’ in ascidian tailbuds, both Wnt (Wnt5 and Wnttun5 (tunicate Wnt family member 5)) and FGF (FGF8/17/18 and FGF9/16/20) ligands are expressed in the tail-tip precursors of Ciona from mid-gastrula stages and retinoic acid (RA) synthesizing enzyme (Raldh2) is expressed more anteriorly in the anterior tail muscle [26,154,168,171,172]. In Ciona, RA is required for HOX-1 gene activation in the neck, trunk ganglion and anterior tail nerve cord as well as the adjacent epidermis at tailbud stage and Wnt signals are required for HOX-12 expression in the posterior most CNS [26,173,174,175]. However, the HOX genes play only a limited role in ascidian larval CNS development [176]. Wnt-, FGF- and RA- signals are involved in patterning the epidermis and associated peripheral nervous system of the ascidian larvae and FGF8/17/18 and Wnt-5 ligands are required for correct tail tip cell morphology and tail elongation [172,175,176]. Thus, while it seems that the posterior patterning mechanisms that operate in vertebrates are functioning in ascidians, they do not appear to play a major role in posterior CNS development or NMp specification in the A-lineage. It would be extremely interesting to address if these patterning mechanisms are involved in the fate choice between neural and mesoderm in the neuromesoderm precursors of the b-lineage, which is found at the very tip of the tail and about which very little is known.
We conclude that the mechanisms of neuromesoderm specification in ascidians and vertebrates have, in the most part, diverged significantly.
7. A Common Origin for Neuromesoderm Precursors at the Base of Chordates?
In summary, neuromesodermal precursors make extensive contributions to both the ascidian and vertebrate CNS (Figure 6). In ascidians, the anterior part of the CNS, the anterior sensory vesicle, is generated from the a-lineages, following a cell fate choice between neural and epidermis. Much of the remaining CNS, which constitutes a large part, is generated from neuromesodermal precursors originating from the vegetal hemisphere (Figure 3, Figure 4 and Figure 6). These lineages contribute to the lateral and ventral neural tube from the posterior part of the sensory vesicle to the tail nerve cord following a cell fate choice between neural and notochord, with a small part arising from the A8.16 lineage that undergoes an additional lineage segregation between neural and muscle (Figure 3 and Figure 4). Small parts of the dorsal neural tube are generated from the b8.17 lineage, which also gives rise to muscle and endodermal strand cells at the tip of the tail, although its contribution to the neural tube requires confirmation in Ciona. The b8.19 lineage generates most of the dorsal neural tube in both Ciona and Halocynthia. While this lineage also generates the tail-tip muscles in Halocynthia, it is neural-restricted in Ciona (Figure 3 and Figure 4).
Neuromesoderm-derived neural precursors also contribute to the generation of the vertebrate CNS. Cells that choose between notochord and neural fates generate the floor plate at least at posterior levels, whereas cells that choose between somite and neural fates give rise to a large part of the spinal cord (Figure 6). Thus, in both ascidians and vertebrates, neuromesoderm precursors are contributing to posterior parts of the CNS.
In ascidians, distinct molecular mechanisms are involved in specification of neural fates from the different lineages of the CNS. For example, FGF is required for ‘neural induction’ in a- and b-lineages [76], whereas inhibition of FGF-signaling is required for neural specification in A-lineages (Figure 5; 64-cell stage) [88]. This is consistent with the hypothesis proposed in vertebrates for distinct molecular mechanisms driving the formation of the anterior and posterior CNS [15,17,18,19,177].
7.1. Neural-Muscle Precursors
While a large part of the vertebrate spinal cord shares a common origin with somite as discussed above, the contribution of bi-fated neural-muscle precursors to the ascidian neural tube is clearly more limited; just a few cells, mostly at the tip of the tail, originate from the lateral A8.16 and b8.17 cells of the early gastrula stage embryo, although the b8.19 neuromesoderm precursor contributes more extensively to the dorsal neural tube in Halocynthia (Figure 3, Figure 4 and Figure 6). While the neuromesodermal lineages are associated with self-renewal at certain axial levels in mice [96], this is limited in zebrafish [20,100] and not at all the case in ascidians.
It would be very interesting to address the origin of muscles in much larger ascidian larvae in which a process known as “caudalization” takes place. Caudalization describes the addition of muscle cells to the tadpole tail without changing the number of cells in other tissue types (i.e., the notochord remains at 40 cells) [178]. These larvae, which some colonial ascidians produce, can contain more than 1000 muscle cells, compared to the 42 of Halocynthia or 36 in Ciona [9,178,179]. These additional muscle cells arise by proliferation [180]. It would be extremely interesting to determine if, in these colonial species with giant larvae, the additional muscles of the tail originate from the neural-mesoderm lineages. Interestingly, in Halocynthia, which exhibits a minimal “caudalization” with the addition of six muscle cells to the tail, the additional muscle cells arise from b-lineage neural-muscle precursors (Figure 3 and Figure 4) [8,178].
Like in vertebrates, the ascidian neural-muscle precursors are associated with posterior neural tube, and germ layer segregation takes place late during development; the segregation of neural and muscle lineages in A- and b-lineages represents one of the latest events of germ-layer segregation to take place [8] However, unlike in vertebrates, A-lineage neural-muscle precursors do not persist beyond gastrulation with their lineage segregation taking place within the neural plate at the mid-gastrula stage (Figure 3B) [96,100,101]. It has not been reported when muscle and neural fates segregate in the b-lineage. The presence of neural-muscle (or somite) precursors in both vertebrates and ascidians supports their presence in the last common ancestor of Olfactores (here and [9]) but the extent of their contribution to the neural tube is variable.
7.2. Notochord-Neural Precursors
Neural tissue arising from bi-fated and bi-potential notochord-neural precursors generate a large part of the CNS of ascidians (Figure 3, Figure 4 and Figure 6). The A6.2 cell of the 32-cell stage embryo is a bi-fated notochord-neural precursor that contributes cells up to forebrain levels in the CNS. The ‘forebrain’ contribution of A-lineage cells is based on the A-lineage origin of the ocellus-associated ciliary photoreceptor cells, most likely corresponding to forebrain-derived retinal or pineal photoreceptors of vertebrates [36,38,53]. At the level of the trunk ganglion and tail nerve cord, A6.2 also contributes to the ventral row of cells (Figure 3 and Figure 4). This ventral contribution bears a strong resemblance to the notochord-neural precursors that generate the medial floor plate of vertebrates. In both ascidians and vertebrates, FoxA and Hedgehog genes are expressed in these ventral cells, although so far there is no evidence that either the notochord or floor plate plays a role in patterning the neural tube in ascidians as they do in vertebrates [17,67,119,162,165,181,182]. The other ‘notochord-neural’ precursors of the 32-cell stage embryo, the A6.4 cell, give rise to notochord, a few posterior muscle cells and to all the lateral cells of the trunk ganglion and tail nerve cord (‘hindbrain-spinal cord’) (Figure 2, Figure 3 and Figure 4). Together, the ascidian notochord-neural precursors generate a much larger part of the neural tube compared to vertebrates; all ventral and lateral cells from ‘posterior forebrain’ (photoreceptors) to ‘spinal cord’ (Figure 6). Despite these differences in the levels of CNS contribution, a common origin for neural and notochord shared between ascidians and vertebrates is nonetheless striking and argues once more for common neuromesodermal origin of part of the neural tube at the base of chordates. Interestingly, the segregation of notochord from neural takes place very early in development in ascidians, prior to gastrulation, similar to zebrafish and perhaps also mouse, although it has not yet been demonstrated if notochord-floor plate cells arise from an early common precursor in mouse.
7.3. Ascidian b-Lineage Cells: Neural, Muscle, Endoderm
The contribution of b8.17 cells to dorsal neural tube, muscle and endodermal strand in the tail tip bears some resemblance to axial precursors in the tailbud of zebrafish and chick which generate floor plate, notochord and hypochord (probable endoderm underlying the notochord of zebrafish in a similar position to the endodermal strand of ascidians) [100,105,126,130,133,182,183]. However, the ascidian b-lineage cells contribute to dorsal rather than ventral neural tube and muscle rather than notochord. In addition, these precursors arise at the lateral border of the future neural plate of ascidian embryos, a region proposed to be an evolutionary precursor of neural crest/placode [30,39,56,79]. A proto-neural crest character may explain why b-lineage cells contribute to multiple germ layers (epidermis, neural tube, muscle, endoderm) and may account for the relatively late tissue fate restriction of cells that originate in these lineages [8]. On the other hand, the b8.17 lineage may also represent tailbud-like neural-muscle-endoderm precursors, with the b8.19 lineage of Halocynthia representing a tailbud-like muscle-neural precursor (NMp). Understanding more about the lineage segregation of b-lineage cells and their specification will help shed light on whether they are more neural-border-like or tailbud precursor-like (axial or NMp).
8. Conclusions
We conclude that, in both vertebrates and ascidians, part of the posterior neural tissue shares a common origin with mesoderm, although the mechanisms of specification differ. We therefore propose that a shared developmental origin of mesoderm and neural tissue in ascidian embryos is not a peculiarity of ascidians but descended from a common ancestor in which some neural tissue originates from a neural-epidermis fate choice and some from a neural-mesoderm fate choice and that the extent of these contributions has come to vary in different evolutionary lineages.
While a different embryonic origin of the anterior and posterior CNS in ascidians is entirely consistent with the proposed independent origin of anterior and posterior CNS in vertebrates [15,17,18], the extent of these differential contributions is not the same between ascidians and vertebrates. To explain how the boundary between neuro-epidermis and neuro-mesoderm contributions to the CNS does not appear to be fixed in different chordate embryos, we suggest that the mechanisms that pattern the CNS along the anterior-posterior axis and those that establish neural identity may act, and therefore evolve, independently (or can at least be uncoupled), as suggested by recent work in mice [95].
It has been experimentally demonstrated that cells following distinct transcriptional trajectories can converge upon a similar cell type [184] (see also [92,185]). This suggests that cells from distinct embryonic lineage origins, undergoing distinct transcriptional trajectories should be able to seamlessly unite to form a single functional unit, in this case the CNS. However, this is not unique to the CNS; for example, in ascidians, in addition to CNS, both notochord and muscle arise in distinct lineages via distinct molecular mechanisms [4,9,85]. Ascidians could provide an excellent model system to address the process whereby distinct embryonic and transcriptional trajectories ultimately converge upon a similar cell type.
9. C. robusta Unique Gene IDs
From the KH2012 C. robusta genome assembly. See Ghost (http://ghost.zool.kyoto-u.ac.jp/otherfr_kh.html, accessed 7 April 2021) ([168,186] and ANISEED (http://www.aniseed.cnrs.fr/, accessed 7 April 2021) [187] for further information. Wnt 5 (KH.L152.45), Wnttun5 (tunicate Wnt family member 5, KH.C9.257), FGF9/16/20 (KH.C2.125), FGF8/17/18 (KH.C5.5), ERK (KH.L153.20), Nodal (KH.L106.16), Delta-2/Delta-like (KH.L50.6), β-catenin (KH.C9.53), Hedgehog-2 (KH.C5.544), Raldh2 (KH.C4.697), Sox2/14/21 (KH.S164.12; KH.C1.99), Cdx (KH.C14.408), Brachyury/T (KH.S1404.1), Snail (KH.C3.751), Tbx6 (a multi-copy gene [188], KH.S654.3- tbx6b); HOX-1 (KH.L171.16), HOX-12 (KH.C7.472), Zic.r.b (a multi-copy gene [189], KH.S816.1), macho/Zic-r.a (KH.C1.727), Gata.a (KH.L20.1).
Author Contributions
Conceptualization C.H., H.Y.; writing—original draft preparation, C.H.; writing—review and editing, C.H., H.Y.; funding acquisition, H.Y. All authors have read and agreed to the published version of the manuscript.
Funding
Research in our lab is supported by the Centre National de la Recherche Scientifique (CNRS), Sorbonne University and the Agence Nationale de la Recherche (ANR-17-CE13-0003-01).
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not Applicable.
Data Availability Statement
No new data is reported in this study. Links to web sites accessed are found in Section 9.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Delsuc, F.; Brinkmann, H.; Chourrout, D.; Philippe, H. Tunicates and Not Cephalochordates Are the Closest Living Relatives of Vertebrates. Nature 2006, 439, 965–968. [Google Scholar] [CrossRef] [PubMed]
- Dumollard, R.; Minc, N.; Salez, G.; Aicha, S.B.; Bekkouche, F.; Hebras, C.; Besnardeau, L.; McDougall, A. The Invariant Cleavage Pattern Displayed by Ascidian Embryos Depends on Spindle Positioning along the Cell’s Longest Axis in the Apical Plane and Relies on Asynchronous Cell Divisions. eLife 2017, 6, e19290. [Google Scholar] [CrossRef] [Green Version]
- Guignard, L.; Fiúza, U.-M.; Leggio, B.; Laussu, J.; Faure, E.; Michelin, G.; Biasuz, K.; Hufnagel, L.; Malandain, G.; Godin, C.; et al. Contact Area–Dependent Cell Communication and the Morphological Invariance of Ascidian Embryogenesis. Science 2020, 369, eaar5663. [Google Scholar] [CrossRef] [PubMed]
- Hudson, C.; Yasuo, H. Similarity and Diversity in Mechanisms of Muscle Fate Induction between Ascidian Species. Biol. Cell 2008, 100, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, P. Unfolding a Chordate Developmental Program, One Cell at a Time: Invariant Cell Lineages, Short-Range Inductions and Evolutionary Plasticity in Ascidians. Dev. Biol. 2009, 332, 48–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemaire, P. Evolutionary Crossroads in Developmental Biology: The Tunicates. Development 2011, 138, 2143–2152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowe, E.K.; Stolfi, A. Developmental System Drift in Motor Ganglion Patterning between Distantly Related Tunicates. Evodevo 2018, 9, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishida, H. Cell Lineage Analysis in Ascidian Embryos by Intracellular Injection of a Tracer Enzyme. III. Up to the Tissue Restricted Stage. Dev. Biol. 1987, 121, 526–541. [Google Scholar] [CrossRef]
- Razy-Krajka, F.; Stolfi, A. Regulation and Evolution of Muscle Development in Tunicates. EvoDevo 2019, 10, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stolfi, A.; Lowe, E.K.; Racioppi, C.; Ristoratore, F.; Brown, C.T.; Swalla, B.J.; Christiaen, L. Divergent Mechanisms Regulate Conserved Cardiopharyngeal Development and Gene Expression in Distantly Related Ascidians. Elife 2014, 3, e03728. [Google Scholar] [CrossRef] [PubMed]
- Satoh, N.; Rokhsar, D.; Nishikawa, T. Chordate Evolution and the Three-Phylum System. Proc. Biol. Sci. 2014, 281, 20141729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, A.; Nishida, H. Distinct Parameters Are Involved in Controlling the Number of Rounds of Cell Division in Each Tissue during Ascidian Embryogenesis. J. Exp. Zool. 1999, 284, 379–391. [Google Scholar] [CrossRef]
- Navarrete, I.A.; Levine, M. Nodal and FGF Coordinate Ascidian Neural Tube Morphogenesis. Development 2016, 143, 4665–4675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicol, D.; Meinertzhagen, I.A. Development of the Central Nervous System of the Larva of the Ascidian, Ciona Intestinalis L. II. Neural Plate Morphogenesis and Cell Lineages during Neurulation. Dev. Biol. 1988, 130, 737–766. [Google Scholar] [CrossRef]
- Henrique, D.; Abranches, E.; Verrier, L.; Storey, K.G. Neuromesodermal Progenitors and the Making of the Spinal Cord. Development 2015, 142, 2864–2875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern, C.D.; Charité, J.; Deschamps, J.; Duboule, D.; Durston, A.J.; Kmita, M.; Nicolas, J.-F.; Palmeirim, I.; Smith, J.C.; Wolpert, L. Head-Tail Patterning of the Vertebrate Embryo: One, Two or Many Unresolved Problems? Int. J. Dev. Biol. 2006, 50, 3–15. [Google Scholar] [CrossRef]
- Gouti, M.; Metzis, V.; Briscoe, J. The Route to Spinal Cord Cell Types: A Tale of Signals and Switches. Trends Genet. 2015, 31, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Kondoh, H.; Takada, S.; Takemoto, T. Axial Level-Dependent Molecular and Cellular Mechanisms Underlying the Genesis of the Embryonic Neural Plate. Dev. Growth Differ. 2016, 58, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Al Anber, A.; Martin, B.L. Transformation of a Neural Activation and Patterning Model. EMBO Rep. 2019, 20, e48060. [Google Scholar] [CrossRef] [PubMed]
- Attardi, A.; Fulton, T.; Florescu, M.; Shah, G.; Muresan, L.; Lenz, M.O.; Lancaster, C.; Huisken, J.; van Oudenaarden, A.; Steventon, B. Neuromesodermal Progenitors Are a Conserved Source of Spinal Cord with Divergent Growth Dynamics. Development 2018, 145, dev166728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, R.K. Cell Movements and Cell Fate during Zebrafish Gastrulation. Development 1992, 116, 65–73. [Google Scholar]
- Hashimoto, H.; Robin, F.B.; Sherrard, K.M.; Munro, E.M. Sequential Contraction and Exchange of Apical Junctions Drives Zippering and Neural Tube Closure in a Simple Chordate. Dev. Cell 2015, 32, 241–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasakura, Y.; Mita, K.; Ogura, Y.; Horie, T. Ascidians as Excellent Chordate Models for Studying the Development of the Nervous System during Embryogenesis and Metamorphosis. Dev. Growth Differ. 2012, 54, 420–437. [Google Scholar] [CrossRef] [PubMed]
- Horie, T.; Nakagawa, M.; Sasakura, Y.; Kusakabe, T.G. Cell Type and Function of Neurons in the Ascidian Nervous System. Dev. Growth Differ. 2009, 51, 207–220. [Google Scholar] [CrossRef]
- Hudson, C. The Central Nervous System of Ascidian Larvae. Wiley Interdiscip. Rev. Dev. Biol. 2016, 5, 538–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, K.S.; Stolfi, A.; Levine, M.; Satou, Y. Gene Regulatory Networks Underlying the Compartmentalization of the Ciona Central Nervous System. Development 2009, 136, 285–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, K.; Meinertzhagen, I.A. Neuronal Identity: The Neuron Types of a Simple Chordate Sibling, the Tadpole Larva of Ciona Intestinalis. Curr. Opin. Neurobiol. 2019, 56, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Ryan, K.; Lu, Z.; Meinertzhagen, I.A. The CNS Connectome of a Tadpole Larva of Ciona Intestinalis (L.) Highlights Sidedness in the Brain of a Chordate Sibling. Elife 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Lemaire, L.A.; Wang, W.; Yoon, P.H.; Choi, Y.A.; Parsons, L.R.; Matese, J.C.; Wang, W.; Levine, M.; Chen, K. Comprehensive Single-Cell Transcriptome Lineages of a Proto-Vertebrate. Nature 2019, 571, 349–354. [Google Scholar] [CrossRef]
- Horie, R.; Hazbun, A.; Chen, K.; Cao, C.; Levine, M.; Horie, T. Shared Evolutionary Origin of Vertebrate Neural Crest and Cranial Placodes. Nature 2018, 560, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Horie, T.; Horie, R.; Chen, K.; Cao, C.; Nakagawa, M.; Kusakabe, T.G.; Satoh, N.; Sasakura, Y.; Levine, M. Regulatory Cocktail for Dopaminergic Neurons in a Protovertebrate Identified by Whole-Embryo Single-Cell Transcriptomics. Genes Dev. 2018, 32, 1297–1302. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Wang, W.; Stolfi, A. Single-Cell Transcriptome Profiling of the Ciona Larval Brain. Dev. Biol. 2019, 448, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Sladitschek, H.L.; Fiuza, U.-M.; Pavlinic, D.; Benes, V.; Hufnagel, L.; Neveu, P.A. MorphoSeq: Full Single-Cell Transcriptome Dynamics Up to Gastrulation in a Chordate. Cell 2020, 181, 922–935.e21. [Google Scholar] [CrossRef]
- Borba, C.; Kourakis, M.J.; Schwennicke, S.; Brasnic, L.; Smith, W.C. Visual Processing and Fold-Change Detection by the Larva of the Simple Chordate Ciona. bioRxiv 2020. [Google Scholar] [CrossRef]
- Bostwick, M.; Smith, E.L.; Borba, C.; Newman-Smith, E.; Guleria, I.; Kourakis, M.J.; Smith, W.C. Antagonistic Inhibitory Circuits Integrate Visual and Gravitactic Behaviors. Curr. Biol. 2020, 30, 600–609.e2. [Google Scholar] [CrossRef] [Green Version]
- Kourakis, M.J.; Borba, C.; Zhang, A.; Newman-Smith, E.; Salas, P.; Manjunath, B.; Smith, W.C. Parallel Visual Circuitry in a Basal Chordate. Elife 2019, 8. [Google Scholar] [CrossRef]
- Oonuma, K.; Kusakabe, T.G. Spatio-Temporal Regulation of Rx and Mitotic Patterns Shape the Eye-Cup of the Photoreceptor Cells in Ciona. Dev. Biol. 2019, 445, 245–255. [Google Scholar] [CrossRef]
- Oonuma, K.; Tanaka, M.; Nishitsuji, K.; Kato, Y.; Shimai, K.; Kusakabe, T.G. Revised Lineage of Larval Photoreceptor Cells in Ciona Reveals Archetypal Collaboration between Neural Tube and Neural Crest in Sensory Organ Formation. Dev. Biol. 2016, 420, 178–185. [Google Scholar] [CrossRef]
- Stolfi, A.; Ryan, K.; Meinertzhagen, I.A.; Christiaen, L. Migratory Neuronal Progenitors Arise from the Neural Plate Borders in Tunicates. Nature 2015, 527, 371–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horie, T.; Shinki, R.; Ogura, Y.; Kusakabe, T.G.; Satoh, N.; Sasakura, Y. Ependymal Cells of Chordate Larvae Are Stem-like Cells That Form the Adult Nervous System. Nature 2011, 469, 525–528. [Google Scholar] [CrossRef] [PubMed]
- Esposito, R.; Racioppi, C.; Pezzotti, M.R.; Branno, M.; Locascio, A.; Ristoratore, F.; Spagnuolo, A. The Ascidian Pigmented Sensory Organs: Structures and Developmental Programs. Genesis 2015, 53, 15–33. [Google Scholar] [CrossRef]
- Jiang, D.; Tresser, J.W.; Horie, T.; Tsuda, M.; Smith, W.C. Pigmentation in the Sensory Organs of the Ascidian Larva Is Essential for Normal Behavior. J. Exp. Biol. 2005, 208, 433–438. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, M.; Sakurai, D.; Goda, M. Direct Evidence for the Role of Pigment Cells in the Brain of Ascidian Larvae by Laser Ablation. J. Exp. Biol. 2003, 206, 1409–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dufour, H.D.; Chettouh, Z.; Deyts, C.; de Rosa, R.; Goridis, C.; Joly, J.-S.; Brunet, J.-F. Precraniate Origin of Cranial Motoneurons. Proc. Natl. Acad. Sci. USA 2006, 103, 8727–8732. [Google Scholar] [CrossRef] [Green Version]
- Cole, A.G.; Meinertzhagen, I.A. The Central Nervous System of the Ascidian Larva: Mitotic History of Cells Forming the Neural Tube in Late Embryonic Ciona Intestinalis. Dev. Biol. 2004, 271, 239–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibboney, S.; Orvis, J.; Kim, K.; Johnson, C.J.; Martinez-Feduchi, P.; Lowe, E.K.; Sharma, S.; Stolfi, A. Effector Gene Expression Underlying Neuron Subtype-Specific Traits in the Motor Ganglion of Ciona. Dev. Biol. 2020, 458, 52–63. [Google Scholar] [CrossRef]
- Horie, T.; Nakagawa, M.; Sasakura, Y.; Kusakabe, T.G.; Tsuda, M. Simple Motor System of the Ascidian Larva: Neuronal Complex Comprising Putative Cholinergic and GABAergic/Glycinergic Neurons. Zool. Sci. 2010, 27, 181–190. [Google Scholar] [CrossRef]
- Stolfi, A.; Levine, M. Neuronal Subtype Specification in the Spinal Cord of a Protovertebrate. Development 2011, 138, 995–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, J.H.; Meinertzhagen, I.A. Neurons of the Ascidian Larval Nervous System in Ciona Intestinalis: I. Central Nervous System. J. Comp. Neurol. 2007, 501, 316–334. [Google Scholar] [CrossRef] [PubMed]
- Nishitsuji, K.; Horie, T.; Ichinose, A.; Sasakura, Y.; Yasuo, H.; Kusakabe, T.G. Cell Lineage and Cis-Regulation for a Unique GABAergic/Glycinergic Neuron Type in the Larval Nerve Cord of the Ascidian Ciona Intestinalis. Dev. Growth Differ. 2012, 54, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Nicol, D.; Meinertzhagen, I.A. Cell Counts and Maps in the Larval Central Nervous System of the Ascidian Ciona Intestinalis (L.). J. Comp. Neurol. 1991, 309, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Holland, L.Z. Tunicates. Curr. Biol. 2016, 26, R146–R152. [Google Scholar] [CrossRef] [Green Version]
- Kusakabe, T.G. Identifying Vertebrate Brain Prototypes in Deuterostomes. In Brain Evolution by Design; Diversity and Commonality in Animals; Shigeno, S., Murakami, Y., Nomura, T., Eds.; Springer Japan: Tokyo, Japan, 2017; pp. 153–186. ISBN 978-4-431-56467-6. [Google Scholar]
- Lacalli, T.C. Prospective Protochordate Homologs of Vertebrate Midbrain and MHB, with Some Thoughts on MHB Origins. Int. J. Biol. Sci. 2006, 2, 104–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, B.; Shimeld, S.M. Evolution of Vertebrate Spinal Cord Patterning. Dev. Dyn. 2019, 248, 1028–1043. [Google Scholar] [CrossRef] [Green Version]
- Shimeld, S.M.; Holland, P.W. Vertebrate Innovations. Proc. Natl. Acad. Sci. USA 2000, 97, 4449–4452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cañestro, C.; Bassham, S.; Postlethwait, J. Development of the Central Nervous System in the Larvacean Oikopleura Dioica and the Evolution of the Chordate Brain. Dev. Biol. 2005, 285, 298–315. [Google Scholar] [CrossRef] [Green Version]
- Ikuta, T.; Saiga, H. Dynamic Change in the Expression of Developmental Genes in the Ascidian Central Nervous System: Revisit to the Tripartite Model and the Origin of the Midbrain-Hindbrain Boundary Region. Dev. Biol. 2007, 312, 631–643. [Google Scholar] [CrossRef] [Green Version]
- Imai, K.S.; Satoh, N.; Satou, Y. Region Specific Gene Expressions in the Central Nervous System of the Ascidian Embryo. Mech. Dev. 2002, 119 (Suppl. 1), S275–S277. [Google Scholar] [CrossRef]
- Liu, B.; Satou, Y. The Genetic Program to Specify Ectodermal Cells in Ascidian Embryos. Dev. Growth Differ. 2020, 62, 301–310. [Google Scholar] [CrossRef]
- Satoh, N. The Ascidian Tadpole Larva: Comparative Molecular Development and Genomics. Nat. Rev. Genet. 2003, 4, 285–295. [Google Scholar] [CrossRef]
- Takahashi, T.; Holland, P.W.H. Amphioxus and Ascidian Dmbx Homeobox Genes Give Clues to the Vertebrate Origins of Midbrain Development. Development 2004, 131, 3285–3294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wada, H.; Satoh, N. Patterning the Protochordate Neural Tube. Curr. Opin. Neurobiol. 2001, 11, 16–21. [Google Scholar] [CrossRef]
- Wada, H.; Saiga, H.; Satoh, N.; Holland, P.W. Tripartite Organization of the Ancestral Chordate Brain and the Antiquity of Placodes: Insights from Ascidian Pax-2/5/8, HOX and Otx Genes. Development 1998, 125, 1113–1122. [Google Scholar]
- Nishino, A.; Okamura, Y.; Piscopo, S.; Brown, E.R. A Glycine Receptor Is Involved in the Organization of Swimming Movements in an Invertebrate Chordate. BMC Neurosci. 2010, 11, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishino, A.; Baba, S.A.; Okamura, Y. A Mechanism for Graded Motor Control Encoded in the Channel Properties of the Muscle ACh Receptor. Proc. Natl. Acad. Sci. USA 2011, 108, 2599–2604. [Google Scholar] [CrossRef] [Green Version]
- Stolfi, A.; Wagner, E.; Taliaferro, J.M.; Chou, S.; Levine, M. Neural Tube Patterning by Ephrin, FGF and Notch Signaling Relays. Development 2011, 138, 5429–5439. [Google Scholar] [CrossRef] [Green Version]
- Ryan, K.; Lu, Z.; Meinertzhagen, I.A. Circuit Homology between Decussating Pathways in the Ciona Larval CNS and the Vertebrate Startle-Response Pathway. Curr. Biol. 2017, 27, 721–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aragon, F.; Pujades, C. FGF Signaling Controls Caudal Hindbrain Specification through Ras-ERK1/2 Pathway. BMC Dev. Biol. 2009, 9, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maves, L.; Jackman, W.; Kimmel, C.B. FGF3 and FGF8 Mediate a Rhombomere 4 Signaling Activity in the Zebrafish Hindbrain. Development 2002, 129, 3825–3837. [Google Scholar]
- Walshe, J.; Maroon, H.; McGonnell, I.M.; Dickson, C.; Mason, I. Establishment of Hindbrain Segmental Identity Requires Signaling by FGF3 and FGF8. Curr. Biol. 2002, 12, 1117–1123. [Google Scholar] [CrossRef] [Green Version]
- Conklin, E.G. The Organisation and Cell Lineage of the Ascidian Egg; The Academy of Natural Sciences: Philadelphia, PA, USA, 1905; pp. 1–119. [Google Scholar]
- Nicol, D.; Meinertzhagen, I.A. Development of the Central Nervous System of the Larva of the Ascidian, Ciona Intestinalis L. I. The Early Lineages of the Neural Plate. Dev. Biol. 1988, 130, 721–736. [Google Scholar] [CrossRef]
- Abitua, P.B.; Gainous, T.B.; Kaczmarczyk, A.N.; Winchell, C.J.; Hudson, C.; Kamata, K.; Nakagawa, M.; Tsuda, M.; Kusakabe, T.G.; Levine, M. The Pre-Vertebrate Origins of Neurogenic Placodes. Nature 2015, 524, 462–465. [Google Scholar] [CrossRef]
- Nishida, H. Determinative Mechanisms in Secondary Muscle Lineages of Ascidian Embryos: Development of Muscle-Specific Features in Isolated Muscle Progenitor Cells. Development 1990, 108, 559–568. [Google Scholar]
- Bertrand, V.; Hudson, C.; Caillol, D.; Popovici, C.; Lemaire, P. Neural Tissue in Ascidian Embryos Is Induced by FGF9/16/20, Acting via a Combination of Maternal GATA and Ets Transcription Factors. Cell 2003, 115, 615–627. [Google Scholar] [CrossRef] [Green Version]
- Hudson, C.; Lemaire, P. Induction of Anterior Neural Fates in the Ascidian Ciona Intestinalis. Mech. Dev. 2001, 100, 189–203. [Google Scholar] [CrossRef]
- Hudson, C.; Darras, S.; Caillol, D.; Yasuo, H.; Lemaire, P. A Conserved Role for the MEK Signalling Pathway in Neural Tissue Specification and Posteriorisation in the Invertebrate Chordate, the Ascidian Ciona Intestinalis. Development 2003, 130, 147–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohtsuka, Y.; Matsumoto, J.; Katsuyama, Y.; Okamura, Y. Nodal Signaling Regulates Specification of Ascidian Peripheral Neurons through Control of the BMP Signal. Development 2014, 141, 3889–3899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, E.; Stolfi, A.; Gi Choi, Y.; Levine, M. Islet Is a Key Determinant of Ascidian Palp Morphogenesis. Development 2014, 141, 3084–3092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, T.; Matsuoka, T.; Satou, Y. A Time Delay Gene Circuit Is Required for Palp Formation in the Ascidian Embryo. Development 2013, 140, 4703–4708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohta, N.; Satou, Y. Multiple Signaling Pathways Coordinate to Induce a Threshold Response in a Chordate Embryo. PLoS Genet. 2013, 9, e1003818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, E.; Levine, M. FGF Signaling Establishes the Anterior Border of the Ciona Neural Tube. Development 2012, 139, 2351–2359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, T.; Satou, Y. Differential Temporal Control of Foxa.a and Zic-r.b Specifies Brain versus Notochord Fate in the Ascidian Embryo. Development 2017, 144, 38–43. [Google Scholar] [CrossRef] [Green Version]
- Satoh, N. Developmental Genomics of Ascidians; Wiley-Blackwell: Hoboken, NJ, USA, 2014; ISBN 978-1-118-65618-1. [Google Scholar]
- Pasini, A.; Amiel, A.; Rothbächer, U.; Roure, A.; Lemaire, P.; Darras, S. Formation of the Ascidian Epidermal Sensory Neurons: Insights into the Origin of the Chordate Peripheral Nervous System. PLoS Biol. 2006, 4, e225. [Google Scholar] [CrossRef] [PubMed]
- Minokawa, T.; Yagi, K.; Makabe, K.W.; Nishida, H. Binary Specification of Nerve Cord and Notochord Cell Fates in Ascidian Embryos. Development 2001, 128, 2007–2017. [Google Scholar] [PubMed]
- Picco, V.; Hudson, C.; Yasuo, H. Ephrin-Eph Signalling Drives the Asymmetric Division of Notochord/Neural Precursors in Ciona Embryos. Development 2007, 134, 1491–1497. [Google Scholar] [CrossRef] [Green Version]
- Hudson, C.; Lotito, S.; Yasuo, H. Sequential and Combinatorial Inputs from Nodal, Delta2/Notch and FGF/MEK/ERK Signalling Pathways Establish a Grid-like Organisation of Distinct Cell Identities in the Ascidian Neural Plate. Development 2007, 134, 3527–3537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemaire, P.; Smith, W.C.; Nishida, H. Ascidians and the Plasticity of the Chordate Developmental Program. Curr. Biol. 2008, 18, R620–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokuoka, M.; Kumano, G.; Nishida, H. FGF9/16/20 and Wnt-5alpha Signals Are Involved in Specification of Secondary Muscle Fate in Embryos of the Ascidian, Halocynthia Roretzi. Dev. Genes Evol. 2007, 217, 515–527. [Google Scholar] [CrossRef]
- Nishida, H.; Stach, T. Cell Lineages and Fate Maps in Tunicates: Conservation and Modification. Zoolog. Sci. 2014, 31, 645–652. [Google Scholar] [CrossRef] [Green Version]
- Stach, T.; Winter, J.; Bouquet, J.-M.; Chourrout, D.; Schnabel, R. Embryology of a Planktonic Tunicate Reveals Traces of Sessility. Proc. Natl. Acad. Sci. USA 2008, 105, 7229–7234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cajal, M.; Lawson, K.A.; Hill, B.; Moreau, A.; Rao, J.; Ross, A.; Collignon, J.; Camus, A. Clonal and Molecular Analysis of the Prospective Anterior Neural Boundary in the Mouse Embryo. Development 2012, 139, 423–436. [Google Scholar] [CrossRef] [Green Version]
- Metzis, V.; Steinhauser, S.; Pakanavicius, E.; Gouti, M.; Stamataki, D.; Ivanovitch, K.; Watson, T.; Rayon, T.; Mousavy Gharavy, S.N.; Lovell-Badge, R.; et al. Nervous System Regionalization Entails Axial Allocation before Neural Differentiation. Cell 2018, 175, 1105–1118.e17. [Google Scholar] [CrossRef] [Green Version]
- Tzouanacou, E.; Wegener, A.; Wymeersch, F.J.; Wilson, V.; Nicolas, J.-F. Redefining the Progression of Lineage Segregations during Mammalian Embryogenesis by Clonal Analysis. Dev. Cell 2009, 17, 365–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouti, M.; Tsakiridis, A.; Wymeersch, F.J.; Huang, Y.; Kleinjung, J.; Wilson, V.; Briscoe, J. In Vitro Generation of Neuromesodermal Progenitors Reveals Distinct Roles for Wnt Signalling in the Specification of Spinal Cord and Paraxial Mesoderm Identity. PLoS Biol. 2014, 12, e1001937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimelman, D. Tales of Tails (and Trunks). In Current Topics in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2016; Volume 116, pp. 517–536. ISBN 978-0-12-802956-5. [Google Scholar]
- Kimelman, D.; Griffin, K.J. Vertebrate Mesendoderm Induction and Patterning. Curr. Opin. Genet. Dev. 2000, 10, 350–356. [Google Scholar] [CrossRef]
- Martin, B.L.; Kimelman, D. Canonical Wnt Signaling Dynamically Controls Multiple Stem Cell Fate Decisions during Vertebrate Body Formation. Dev. Cell 2012, 22, 223–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steventon, B.; Martinez Arias, A. Evo-Engineering and the Cellular and Molecular Origins of the Vertebrate Spinal Cord. Dev. Biol. 2017, 432, 3–13. [Google Scholar] [CrossRef]
- Brown, J.M.; Storey, K.G. A Region of the Vertebrate Neural Plate in Which Neighbouring Cells Can Adopt Neural or Epidermal Fates. Curr. Biol. 2000, 10, 869–872. [Google Scholar] [CrossRef] [Green Version]
- Cambray, N.; Wilson, V. Axial Progenitors with Extensive Potency Are Localised to the Mouse Chordoneural Hinge. Development 2002, 129, 4855–4866. [Google Scholar]
- Cambray, N.; Wilson, V. Two Distinct Sources for a Population of Maturing Axial Progenitors. Development 2007, 134, 2829–2840. [Google Scholar] [CrossRef] [Green Version]
- Catala, M.; Teillet, M.A.; De Robertis, E.M.; Le Douarin, M.L. A Spinal Cord Fate Map in the Avian Embryo: While Regressing, Hensen’s Node Lays down the Notochord and Floor Plate Thus Joining the Spinal Cord Lateral Walls. Development 1996, 122, 2599–2610. [Google Scholar] [PubMed]
- Tam, P.P.; Beddington, R.S. The Formation of Mesodermal Tissues in the Mouse Embryo during Gastrulation and Early Organogenesis. Development 1987, 99, 109–126. [Google Scholar]
- Kimmel, C.B.; Warga, R.M.; Schilling, T.F. Origin and Organization of the Zebrafish Fate Map. Development 1990, 108, 581–594. [Google Scholar]
- Ciruna, B.G.; Schwartz, L.; Harpal, K.; Yamaguchi, T.P.; Rossant, J. Chimeric Analysis of Fibroblast Growth Factor Receptor-1 (Fgfr1) Function: A Role for FGFR1 in Morphogenetic Movement through the Primitive Streak. Development 1997, 124, 2829–2841. [Google Scholar] [PubMed]
- Yoshikawa, Y.; Fujimori, T.; McMahon, A.P.; Takada, S. Evidence That Absence of Wnt-3a Signaling Promotes Neuralization Instead of Paraxial Mesoderm Development in the Mouse. Dev. Biol. 1997, 183, 234–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, D.L.; Papaioannou, V.E. Three Neural Tubes in Mouse Embryos with Mutations in the T-Box Gene Tbx6. Nature 1998, 391, 695–697. [Google Scholar] [CrossRef]
- Goto, H.; Kimmey, S.C.; Row, R.H.; Matus, D.Q.; Martin, B.L. FGF and Canonical Wnt Signaling Cooperate to Induce Paraxial Mesoderm from Tailbud Neuromesodermal Progenitors through Regulation of a Two-Step Epithelial to Mesenchymal Transition. Development 2017, 144, 1412–1424. [Google Scholar] [CrossRef] [Green Version]
- Gouti, M.; Delile, J.; Stamataki, D.; Wymeersch, F.J.; Huang, Y.; Kleinjung, J.; Wilson, V.; Briscoe, J. A Gene Regulatory Network Balances Neural and Mesoderm Specification during Vertebrate Trunk Development. Dev. Cell 2017, 41, 243–261.e7. [Google Scholar] [CrossRef]
- Wilson, V.; Olivera-Martinez, I.; Storey, K.G. Stem Cells, Signals and Vertebrate Body Axis Extension. Development 2009, 136, 1591–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wymeersch, F.J.; Huang, Y.; Blin, G.; Cambray, N.; Wilkie, R.; Wong, F.C.K.; Wilson, V. Position-Dependent Plasticity of Distinct Progenitor Types in the Primitive Streak. Elife 2016, 5, e10042. [Google Scholar] [CrossRef]
- Sambasivan, R.; Steventon, B. Neuromesodermal Progenitors: A Basis for Robust Axial Patterning in Development and Evolution. Front. Cell Dev. Biol. 2021, 8, 607516. [Google Scholar] [CrossRef] [PubMed]
- Bouldin, C.M.; Snelson, C.D.; Farr, G.H.; Kimelman, D. Restricted Expression of Cdc25a in the Tailbud Is Essential for Formation of the Zebrafish Posterior Body. Genes Dev. 2014, 28, 384–395. [Google Scholar] [CrossRef] [Green Version]
- Charrier, J.-B.; Lapointe, F.; Le Douarin, N.M.; Teillet, M.-A. Dual Origin of the Floor Plate in the Avian Embryo. Development 2002, 129, 4785–4796. [Google Scholar] [PubMed]
- Odenthal, J.; van Eeden, F.J.; Haffter, P.; Ingham, P.W.; Nüsslein-Volhard, C. Two Distinct Cell Populations in the Floor Plate of the Zebrafish Are Induced by Different Pathways. Dev. Biol. 2000, 219, 350–363. [Google Scholar] [CrossRef] [Green Version]
- Placzek, M.; Briscoe, J. The Floor Plate: Multiple Cells, Multiple Signals. Nat. Rev. Neurosci. 2005, 6, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Strähle, U.; Lam, C.S.; Ertzer, R.; Rastegar, S. Vertebrate Floor-Plate Specification: Variations on Common Themes. Trends Genet. 2004, 20, 155–162. [Google Scholar] [CrossRef]
- Kinder, S.J.; Tsang, T.E.; Wakamiya, M.; Sasaki, H.; Behringer, R.R.; Nagy, A.; Tam, P.P. The Organizer of the Mouse Gastrula Is Composed of a Dynamic Population of Progenitor Cells for the Axial Mesoderm. Development 2001, 128, 3623–3634. [Google Scholar] [PubMed]
- Wilson, V.; Beddington, R.S. Cell Fate and Morphogenetic Movement in the Late Mouse Primitive Streak. Mech. Dev. 1996, 55, 79–89. [Google Scholar] [CrossRef]
- Halpern, M.E.; Ho, R.K.; Walker, C.; Kimmel, C.B. Induction of Muscle Pioneers and Floor Plate Is Distinguished by the Zebrafish No Tail Mutation. Cell 1993, 75, 99–111. [Google Scholar] [CrossRef]
- Halpern, M.E.; Thisse, C.; Ho, R.K.; Thisse, B.; Riggleman, B.; Trevarrow, B.; Weinberg, E.S.; Postlethwait, J.H.; Kimmel, C.B. Cell-Autonomous Shift from Axial to Paraxial Mesodermal Development in Zebrafish Floating Head Mutants. Development 1995, 121, 4257–4264. [Google Scholar]
- Kudoh, T.; Concha, M.L.; Houart, C.; Dawid, I.B.; Wilson, S.W. Combinatorial Fgf and Bmp Signalling Patterns the Gastrula Ectoderm into Prospective Neural and Epidermal Domains. Development 2004, 131, 3581–3592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latimer, A.J.; Appel, B. Notch Signaling Regulates Midline Cell Specification and Proliferation in Zebrafish. Dev. Biol. 2006, 298, 392–402. [Google Scholar] [CrossRef] [Green Version]
- Melby, A.E.; Warga, R.M.; Kimmel, C.B. Specification of Cell Fates at the Dorsal Margin of the Zebrafish Gastrula. Development 1996, 122, 2225–2237. [Google Scholar] [PubMed]
- Shih, J.; Fraser, S.E. Distribution of Tissue Progenitors within the Shield Region of the Zebrafish Gastrula. Development 1995, 121, 2755–2765. [Google Scholar] [PubMed]
- Shih, J.; Fraser, S.E. Characterizing the Zebrafish Organizer: Microsurgical Analysis at the Early-Shield Stage. Development 1996, 122, 1313–1322. [Google Scholar] [PubMed]
- Selleck, M.A.; Stern, C.D. Fate Mapping and Cell Lineage Analysis of Hensen’s Node in the Chick Embryo. Development 1991, 112, 615–626. [Google Scholar] [PubMed]
- Kimmel, C.B.; Warga, R.M. Cell Lineage and Developmental Potential of Cells in the Zebrafish Embryo. Trends Genet. 1988, 4, 68–74. [Google Scholar] [CrossRef]
- Amacher, S.L.; Draper, B.W.; Summers, B.R.; Kimmel, C.B. The Zebrafish T-Box Genes No Tail and Spadetail Are Required for Development of Trunk and Tail Mesoderm and Medial Floor Plate. Development 2002, 129, 3311–3323. [Google Scholar] [PubMed]
- Row, R.H.; Tsotras, S.R.; Goto, H.; Martin, B.L. The Zebrafish Tailbud Contains Two Independent Populations of Midline Progenitor Cells That Maintain Long-Term Germ Layer Plasticity and Differentiate in Response to Local Signaling Cues. Development 2016, 143, 244–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampath, K.; Rubinstein, A.L.; Cheng, A.M.; Liang, J.O.; Fekany, K.; Solnica-Krezel, L.; Korzh, V.; Halpern, M.E.; Wright, C.V. Induction of the Zebrafish Ventral Brain and Floorplate Requires Cyclops/Nodal Signalling. Nature 1998, 395, 185–189. [Google Scholar] [CrossRef]
- Tian, J.; Yam, C.; Balasundaram, G.; Wang, H.; Gore, A.; Sampath, K. A Temperature-Sensitive Mutation in the Nodal-Related Gene Cyclops Reveals That the Floor Plate Is Induced during Gastrulation in Zebrafish. Development 2003, 130, 3331–3342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, Y.; Epstein, D.J. Distinct Regulators of Shh Transcription in the Floor Plate and Notochord Indicate Separate Origins for These Tissues in the Mouse Node. Development 2003, 130, 3891–3902. [Google Scholar] [CrossRef] [Green Version]
- Yamanaka, Y.; Tamplin, O.J.; Beckers, A.; Gossler, A.; Rossant, J. Live Imaging and Genetic Analysis of Mouse Notochord Formation Reveals Regional Morphogenetic Mechanisms. Dev. Cell 2007, 13, 884–896. [Google Scholar] [CrossRef] [Green Version]
- Gray, S.D.; Dale, J.K. Notch Signalling Regulates the Contribution of Progenitor Cells from the Chick Hensen’s Node to the Floor Plate and Notochord. Development 2010, 137, 561–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halpern, M.E.; Hatta, K.; Amacher, S.L.; Talbot, W.S.; Yan, Y.L.; Thisse, B.; Thisse, C.; Postlethwait, J.H.; Kimmel, C.B. Genetic Interactions in Zebrafish Midline Development. Dev. Biol. 1997, 187, 154–170. [Google Scholar] [CrossRef] [Green Version]
- Schäfer, M.; Rembold, M.; Wittbrodt, J.; Schartl, M.; Winkler, C. Medial Floor Plate Formation in Zebrafish Consists of Two Phases and Requires Trunk-Derived Midkine-a. Genes Dev. 2005, 19, 897–902. [Google Scholar] [CrossRef] [Green Version]
- Odenthal, J.; Haffter, P.; Vogelsang, E.; Brand, M.; van Eeden, F.J.; Furutani-Seiki, M.; Granato, M.; Hammerschmidt, M.; Heisenberg, C.P.; Jiang, Y.J.; et al. Mutations Affecting the Formation of the Notochord in the Zebrafish, Danio Rerio. Development 1996, 123, 103–115. [Google Scholar]
- Strähle, U.; Blader, P.; Ingham, P.W. Expression of Axial and Sonic Hedgehog in Wildtype and Midline Defective Zebrafish Embryos. Int. J. Dev. Biol. 1996, 40, 929–940. [Google Scholar]
- Epstein, D.J.; McMahon, A.P.; Joyner, A.L. Regionalization of Sonic Hedgehog Transcription along the Anteroposterior Axis of the Mouse Central Nervous System Is Regulated by Hnf3-Dependent and -Independent Mechanisms. Development 1999, 126, 281–292. [Google Scholar] [PubMed]
- Patten, I.; Kulesa, P.; Shen, M.M.; Fraser, S.; Placzek, M. Distinct Modes of Floor Plate Induction in the Chick Embryo. Development 2003, 130, 4809–4821. [Google Scholar] [CrossRef] [Green Version]
- Le Douarin, N.M.; Halpern, M.E. Discussion Point. Origin and Specification of the Neural Tube Floor Plate: Insights from the Chick and Zebrafish. Curr. Opin. Neurobiol. 2000, 10, 23–30. [Google Scholar] [CrossRef]
- Placzek, M.; Dodd, J.; Jessell, T.M. Discussion Point. The Case for Floor Plate Induction by the Notochord. Curr. Opin. Neurobiol. 2000, 10, 15–22. [Google Scholar] [CrossRef]
- Hudson, C.; Kawai, N.; Negishi, T.; Yasuo, H. β-Catenin-Driven Binary Fate Specification Segregates Germ Layers in Ascidian Embryos. Curr. Biol. 2013, 23, 491–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobral, D.; Tassy, O.; Lemaire, P. Highly Divergent Gene Expression Programs Can Lead to Similar Chordate Larval Body Plans. Curr. Biol. 2009, 19, 2014–2019. [Google Scholar] [CrossRef]
- Takatori, N.; Kumano, G.; Saiga, H.; Nishida, H. Segregation of Germ Layer Fates by Nuclear Migration-Dependent Localization of Not MRNA. Dev. Cell 2010, 19, 589–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothbacher, U.; Bertrand, V.; Lamy, C.; Lemaire, P. A Combinatorial Code of Maternal GATA, Ets and -Catenin-TCF Transcription Factors Specifies and Patterns the Early Ascidian Ectoderm. Development 2007, 134, 4023–4032. [Google Scholar] [CrossRef] [Green Version]
- Imai, K.; Takada, N.; Satoh, N.; Satou, Y. (β)-Catenin Mediates the Specification of Endoderm Cells in Ascidian Embryos. Development 2000, 127, 3009–3020. [Google Scholar] [PubMed]
- Imai, K.S.; Hudson, C.; Oda-Ishii, I.; Yasuo, H.; Satou, Y. Antagonism between β-Catenin and Gata.a Sequentially Segregates the Germ Layers of Ascidian Embryos. Development 2016, 143, 4167–4172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, G.J.; Kumano, G.; Nishida, H. Cell Fate Polarization in Ascidian Mesenchyme/Muscle Precursors by Directed FGF Signaling and Role for an Additional Ectodermal FGF Antagonizing Signal in Notochord/Nerve Cord Precursors. Development 2007, 134, 1509–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, K.S.; Satoh, N.; Satou, Y. Early Embryonic Expression of FGF4/6/9 Gene and Its Role in the Induction of Mesenchyme and Notochord in Ciona Savignyi Embryos. Development 2002, 129, 1729–1738. [Google Scholar] [PubMed]
- Kobayashi, K.; Sawada, K.; Yamamoto, H.; Wada, S.; Saiga, H.; Nishida, H. Maternal Macho-1 Is an Intrinsic Factor That Makes Cell Response to the Same FGF Signal Differ between Mesenchyme and Notochord Induction in Ascidian Embryos. Development 2003, 130, 5179–5190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, C.; Yasuo, H. Patterning across the Ascidian Neural Plate by Lateral Nodal Signalling Sources. Development 2005, 132, 1199–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mita, K.; Fujiwara, S. Nodal Regulates Neural Tube Formation in the Ciona Intestinalis Embryo. Dev. Genes Evol. 2007, 217, 593–601. [Google Scholar] [CrossRef] [Green Version]
- Imai, K.S.; Levine, M.; Satoh, N.; Satou, Y. Regulatory Blueprint for a Chordate Embryo. Science 2006, 312, 1183–1187. [Google Scholar] [CrossRef] [PubMed]
- Loh, K.M.; van Amerongen, R.; Nusse, R. Generating Cellular Diversity and Spatial Form: Wnt Signaling and the Evolution of Multicellular Animals. Dev. Cell 2016, 38, 643–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, S.Q.; Bowerman, B. Animal Development: An Ancient β-Catenin Switch? Curr. Biol. 2013, 23, R313–315. [Google Scholar] [CrossRef] [Green Version]
- Briscoe, J.; Ericson, J. The Specification of Neuronal Identity by Graded Sonic Hedgehog Signalling. Semin. Cell Dev. Biol. 1999, 10, 353–362. [Google Scholar] [CrossRef] [Green Version]
- Hudson, C.; Ba, M.; Rouvière, C.; Yasuo, H. Divergent Mechanisms Specify Chordate Motoneurons: Evidence from Ascidians. Development 2011, 138, 1643–1652. [Google Scholar] [CrossRef] [Green Version]
- Islam, A.F.M.T.; Moly, P.K.; Miyamoto, Y.; Kusakabe, T.G. Distinctive Expression Patterns of Hedgehog Pathway Genes in the Ciona Intestinalis Larva: Implications for a Role of Hedgehog Signaling in Postembryonic Development and Chordate Evolution. Zoolog. Sci. 2010, 27, 84–90. [Google Scholar] [CrossRef]
- Katsuyama, Y.; Okada, T.; Matsumoto, J.; Ohtsuka, Y.; Terashima, T.; Okamura, Y. Early Specification of Ascidian Larval Motor Neurons. Dev. Biol. 2005, 278, 310–322. [Google Scholar] [CrossRef] [Green Version]
- Takatori, N.; Satou, Y.; Satoh, N. Expression of Hedgehog Genes in Ciona Intestinalis Embryos. Mech. Dev. 2002, 116, 235–238. [Google Scholar] [CrossRef]
- Hudson, C.; Sirour, C.; Yasuo, H. Snail Mediates Medial-Lateral Patterning of the Ascidian Neural Plate. Dev. Biol. 2015, 403, 172–179. [Google Scholar] [CrossRef]
- Hudson, C.; Yasuo, H. A Signalling Relay Involving Nodal and Delta Ligands Acts during Secondary Notochord Induction in Ciona Embryos. Development 2006, 133, 2855–2864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, K.S.; Hino, K.; Yagi, K.; Satoh, N.; Satou, Y. Gene Expression Profiles of Transcription Factors and Signaling Molecules in the Ascidian Embryo: Towards a Comprehensive Understanding of Gene Networks. Development 2004, 131, 4047–4058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, C.; Sirour, C.; Yasuo, H. Co-Expression of Foxa.a, Foxd and Fgf9/16/20 Defines a Transient Mesendoderm Regulatory State in Ascidian Embryos. Elife 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.-L.; Akiyama, R.; Bessho, Y.; Matsui, T. ERK Activity Dynamics during Zebrafish Embryonic Development. Int. J. Mol. Sci. 2018, 20, 109. [Google Scholar] [CrossRef] [Green Version]
- Martin, B.L.; Kimelman, D. Wnt Signaling and the Evolution of Embryonic Posterior Development. Curr. Biol. 2009, 19, R215–219. [Google Scholar] [CrossRef] [Green Version]
- Pasini, A.; Manenti, R.; Rothbächer, U.; Lemaire, P. Antagonizing Retinoic Acid and FGF/MAPK Pathways Control Posterior Body Patterning in the Invertebrate Chordate Ciona Intestinalis. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Feinberg, S.; Roure, A.; Piron, J.; Darras, S. Antero-Posterior Ectoderm Patterning by Canonical Wnt Signaling during Ascidian Development. PLoS Genet. 2019, 15, e1008054. [Google Scholar] [CrossRef] [Green Version]
- Kanda, M.; Ikeda, T.; Fujiwara, S. Identification of a Retinoic Acid-Responsive Neural Enhancer in the Ciona Intestinalis Hox1 Gene. Dev. Growth Differ. 2013, 55, 260–269. [Google Scholar] [CrossRef]
- Sasakura, Y.; Kanda, M.; Ikeda, T.; Horie, T.; Kawai, N.; Ogura, Y.; Yoshida, R.; Hozumi, A.; Satoh, N.; Fujiwara, S. Retinoic Acid-Driven Hox1 Is Required in the Epidermis for Forming the Otic/Atrial Placodes during Ascidian Metamorphosis. Development 2012, 139, 2156–2160. [Google Scholar] [CrossRef] [Green Version]
- Ikuta, T.; Satoh, N.; Saiga, H. Limited Functions of HOX Genes in the Larval Development of the Ascidian Ciona Intestinalis. Development 2010, 137, 1505–1513. [Google Scholar] [CrossRef] [Green Version]
- Polevoy, H.; Gutkovich, Y.E.; Michaelov, A.; Volovik, Y.; Elkouby, Y.M.; Frank, D. New Roles for Wnt and BMP Signaling in Neural Anteroposterior Patterning. EMBO Rep. 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, W.R.; Swalla, B.J. Evolution of Alternate Modes of Development in Ascidians. Bioessays 1992, 14, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Satoh, N. Developmental Biology of Ascidians; Developmental and Cell Biology Series; Cambridge University Press: Cambridge, UK; New York, NY, USA, 1994; ISBN 978-0-521-35221-5. [Google Scholar]
- Cavey, M.J. Myogenic Events in Compound Ascidian Larvae. Am. Zool. 1982, 22, 807–815. [Google Scholar] [CrossRef] [Green Version]
- Corbo, J.C.; Erives, A.; Di Gregorio, A.; Chang, A.; Levine, M. Dorsoventral Patterning of the Vertebrate Neural Tube Is Conserved in a Protochordate. Development 1997, 124, 2335–2344. [Google Scholar]
- Teillet, M.A.; Lapointe, F.; Le Douarin, N.M. The Relationships between Notochord and Floor Plate in Vertebrate Development Revisited. Proc. Natl. Acad. Sci. USA 1998, 95, 11733–11738. [Google Scholar] [CrossRef] [Green Version]
- Appel, B.; Fritz, A.; Westerfield, M.; Grunwald, D.J.; Eisen, J.S.; Riley, B.B. Delta-Mediated Specification of Midline Cell Fates in Zebrafish Embryos. Curr. Biol. 1999, 9, 247–256. [Google Scholar] [CrossRef] [Green Version]
- Briggs, J.A.; Li, V.C.; Lee, S.; Woolf, C.J.; Klein, A.; Kirschner, M.W. Mouse Embryonic Stem Cells Can Differentiate via Multiple Paths to the Same State. Elife 2017, 6, e26945. [Google Scholar] [CrossRef]
- Arendt, D.; Musser, J.M.; Baker, C.V.H.; Bergman, A.; Cepko, C.; Erwin, D.H.; Pavlicev, M.; Schlosser, G.; Widder, S.; Laubichler, M.D.; et al. The Origin and Evolution of Cell Types. Nat. Rev. Genet. 2016, 17, 744–757. [Google Scholar] [CrossRef] [PubMed]
- Satou, Y.; Kawashima, T.; Shoguchi, E.; Nakayama, A.; Satoh, N. An Integrated Database of the Ascidian, Ciona Intestinalis: Towards Functional Genomics. Zool. Sci. 2005, 22, 837–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brozovic, M.; Dantec, C.; Dardaillon, J.; Dauga, D.; Faure, E.; Gineste, M.; Louis, A.; Naville, M.; Nitta, K.R.; Piette, J.; et al. ANISEED 2017: Extending the Integrated Ascidian Database to the Exploration and Evolutionary Comparison of Genome-Scale Datasets. Nucleic Acids Res. 2018, 46, D718–D725. [Google Scholar] [CrossRef] [PubMed]
- Takatori, N.; Hotta, K.; Mochizuki, Y.; Satoh, G.; Mitani, Y.; Satoh, N.; Satou, Y.; Takahashi, H. T-Box Genes in the Ascidian Ciona Intestinalis: Characterization of CDNAs and Spatial Expression. Dev. Dyn. 2004, 230, 743–753. [Google Scholar] [CrossRef] [PubMed]
- Satou, Y.; Imai, K.S. Ascidian Zic Genes. Adv. Exp. Med. Biol. 2018, 1046, 87–106. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Morphological features of the larval CNS. Schematic drawing of the larval CNS, enlarged within the larval shape outline for illustration purposes. Right is a cross-section though the tail showing the chordate organization of axial notochord (N, light blue), lateral muscles (M, dark blue) and the neural tube with a four-cell configuration (NT, dark grey). ES = endodermal strand.
Figure 1.
Morphological features of the larval CNS. Schematic drawing of the larval CNS, enlarged within the larval shape outline for illustration purposes. Right is a cross-section though the tail showing the chordate organization of axial notochord (N, light blue), lateral muscles (M, dark blue) and the neural tube with a four-cell configuration (NT, dark grey). ES = endodermal strand.

Figure 2.
CNS organization along the anterior-posterior axis in ascidians and vertebrates. Left is a schematic drawing of the vertebrate CNS compartments indicated as boxes, with forebrain colored in red, midbrain in orange, mid-hindbrain (MHB) in green, hindbrain in tan and spinal cord in yellow. Right are four models of the ascidian CNS with the different morphological compartments (sensory vesicle, neck, trunk ganglion and tail nerve cord) indicated as boxes. Colors indicate equivalence to vertebrate CNS compartments in four different models. White indicates no (or uncertain) homology. Models are based on information in [63]-Model i; [44]-Model ii; [57]-Model iii; with Model iv showing an integrated summary of previous and recent data (see text for details).
Figure 2.
CNS organization along the anterior-posterior axis in ascidians and vertebrates. Left is a schematic drawing of the vertebrate CNS compartments indicated as boxes, with forebrain colored in red, midbrain in orange, mid-hindbrain (MHB) in green, hindbrain in tan and spinal cord in yellow. Right are four models of the ascidian CNS with the different morphological compartments (sensory vesicle, neck, trunk ganglion and tail nerve cord) indicated as boxes. Colors indicate equivalence to vertebrate CNS compartments in four different models. White indicates no (or uncertain) homology. Models are based on information in [63]-Model i; [44]-Model ii; [57]-Model iii; with Model iv showing an integrated summary of previous and recent data (see text for details).

Figure 3.
Lineage contributions to the larval CNS. (A) 8-cell stage embryo in lateral view (left) showing the four founder lineages, a4.2 (grey) and b4.2 (green) of the animal hemisphere and A4.1 (tan/yellow) and B4.1 (white) of the vegetal hemisphere. Middle shows a schematic drawing of the larval CNS with the contributions from the a4.2 (grey), b4.2 (green) and A4.1 (tan and yellow) shown and summarizing the main contributions from neural-epidermis precursors and neural-mesoderm precursors. Right is a cross-section though the tail showing the chordate organization of axial notochord (N, light blue), lateral muscles (M, dark blue) and the neural tube (NT, green, yellow and tan). For NT, contributions of founder lineages are indicated with colors whereas notochord derives from A-lineage (mostly) and B-lineage and muscle derives from B-(mostly), A- and b-lineage. ES-endoderm strand. (B) Schematic drawings of embryos in vegetal pole view and cell lineages of one A4.1 and one b4.2 showing the bi-fated neuromesoderm precursors in red. Other colors indicate fate-restricted lineages following the same color code as (A) The green dots indicate b-line CNS precursors, the fate segregation of which is not fully resolved.
Figure 3.
Lineage contributions to the larval CNS. (A) 8-cell stage embryo in lateral view (left) showing the four founder lineages, a4.2 (grey) and b4.2 (green) of the animal hemisphere and A4.1 (tan/yellow) and B4.1 (white) of the vegetal hemisphere. Middle shows a schematic drawing of the larval CNS with the contributions from the a4.2 (grey), b4.2 (green) and A4.1 (tan and yellow) shown and summarizing the main contributions from neural-epidermis precursors and neural-mesoderm precursors. Right is a cross-section though the tail showing the chordate organization of axial notochord (N, light blue), lateral muscles (M, dark blue) and the neural tube (NT, green, yellow and tan). For NT, contributions of founder lineages are indicated with colors whereas notochord derives from A-lineage (mostly) and B-lineage and muscle derives from B-(mostly), A- and b-lineage. ES-endoderm strand. (B) Schematic drawings of embryos in vegetal pole view and cell lineages of one A4.1 and one b4.2 showing the bi-fated neuromesoderm precursors in red. Other colors indicate fate-restricted lineages following the same color code as (A) The green dots indicate b-line CNS precursors, the fate segregation of which is not fully resolved.

Figure 4.
Lineage tracing of the bi-fated neuromesoderm precursors. The neuromesoderm precursor labelled is indicated on the left in green and contribution to neural and mesoderm fates indicated by green on the tailbud drawings [8,75].

Figure 5.
Molecular mechanisms of A-lineage neuromesoderm segregation in Ciona from 8-cell stage to neural segregation. One A4.1 founder lineage is shown (the other side is identical). Cells are colored following the key at the bottom. β-catenin and ERK activity is depicted by boxes on the lineage bars, just above the cell name; signals arising from neighboring cells (Nodal and Delta) are depicted by boxes and arrows. Thick lineage brackets indicate cell division along the animal-vegetal axis (16-to-32-cell stage, 32-to-64-cell stage) or anterior-posterior axis (neural plate stage), whereas thin brackets indicate cell division along the medial-lateral axis. The positions of cells following each cell division is shown on the embryo drawings on the left, using the same color code, with the stages indicated.
Figure 5.
Molecular mechanisms of A-lineage neuromesoderm segregation in Ciona from 8-cell stage to neural segregation. One A4.1 founder lineage is shown (the other side is identical). Cells are colored following the key at the bottom. β-catenin and ERK activity is depicted by boxes on the lineage bars, just above the cell name; signals arising from neighboring cells (Nodal and Delta) are depicted by boxes and arrows. Thick lineage brackets indicate cell division along the animal-vegetal axis (16-to-32-cell stage, 32-to-64-cell stage) or anterior-posterior axis (neural plate stage), whereas thin brackets indicate cell division along the medial-lateral axis. The positions of cells following each cell division is shown on the embryo drawings on the left, using the same color code, with the stages indicated.

Figure 6.
Relative contribution of neuromesoderm precursors to the larval CNS in ascidians and vertebrates along the anterior-posterior axis. The animal drawings show the positions of the equivalent CNS regions along the anterior-posterior axis, with the same color code as in Figure 2. MHB is not colored. Mouse and zebrafish drawings are taken from ref.[101](part of their Figure 3), with permission from Elsevier. Below the drawings is a schematic representation of the anatomical compartments with colors showing equivalent regions, from Figure 2. For ascidians, model iv is taken. Below the models the contributions of neuromesoderm (bi-fated or bi-potential) precursors is indicated, with neural-somite/muscle precursors in purple and neural-notochord in blue. The diagonal striped colored box represents the A7.8 lineage, which segregates from notochord at the 64-cell stage as a neural-muscle precursor. Ascidian neuromesoderm contributions extent into more anterior regions of the CNS compared to vertebrates. Vertebrate summary based on [20,119,120,144]. sv, sensory vesicle; n, neck; tg, trunk ganglion; tnc, tail nerve cord; fb, forebrain; mb, midbrain; mhb, mid-hindbrain boundary; hb, hindbrain; sc, spinal cord.
Figure 6.
Relative contribution of neuromesoderm precursors to the larval CNS in ascidians and vertebrates along the anterior-posterior axis. The animal drawings show the positions of the equivalent CNS regions along the anterior-posterior axis, with the same color code as in Figure 2. MHB is not colored. Mouse and zebrafish drawings are taken from ref.[101](part of their Figure 3), with permission from Elsevier. Below the drawings is a schematic representation of the anatomical compartments with colors showing equivalent regions, from Figure 2. For ascidians, model iv is taken. Below the models the contributions of neuromesoderm (bi-fated or bi-potential) precursors is indicated, with neural-somite/muscle precursors in purple and neural-notochord in blue. The diagonal striped colored box represents the A7.8 lineage, which segregates from notochord at the 64-cell stage as a neural-muscle precursor. Ascidian neuromesoderm contributions extent into more anterior regions of the CNS compared to vertebrates. Vertebrate summary based on [20,119,120,144]. sv, sensory vesicle; n, neck; tg, trunk ganglion; tnc, tail nerve cord; fb, forebrain; mb, midbrain; mhb, mid-hindbrain boundary; hb, hindbrain; sc, spinal cord.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hudson, C.; Yasuo, H. Neuromesodermal Lineage Contribution to CNS Development in Invertebrate and Vertebrate Chordates. Genes 2021, 12, 592. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12040592
AMA Style
Hudson C, Yasuo H. Neuromesodermal Lineage Contribution to CNS Development in Invertebrate and Vertebrate Chordates. Genes. 2021; 12(4):592. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12040592
Chicago/Turabian StyleHudson, Clare, and Hitoyoshi Yasuo. 2021. "Neuromesodermal Lineage Contribution to CNS Development in Invertebrate and Vertebrate Chordates" Genes 12, no. 4: 592. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12040592
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.