
Down-Regulation of SlGRAS10 in Tomato Confers Abiotic Stress Tolerance


1
Key Laboratory of Plant Hormones and Development Regulation of Chongqing, School of Life Sciences, Chongqing University, Chongqing 400044, China
2
College of Life Sciences, Henan Normal University, Xinxiang 453007, China
*
Author to whom correspondence should be addressed.
Genes 2021, 12(5), 623; https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050623
Submission received: 9 February 2021
/
Revised: 15 April 2021
/
Accepted: 20 April 2021
/
Published: 22 April 2021
(This article belongs to the Special Issue Genetics and Physiology of Multiple-Stress Tolerance in Crops)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Adverse environmental factors like salt stress, drought, and extreme temperatures, cause damage to plant growth, development, and crop yield. GRAS transcription factors (TFs) have numerous functions in biological processes. Some studies have reported that the GRAS protein family plays significant functions in plant growth and development under abiotic stresses. In this study, we demonstrated the functional characterization of a tomato SlGRAS10 gene under abiotic stresses such as salt stress and drought. Down-regulation of SlGRAS10 by RNA interference (RNAi) produced dwarf plants with smaller leaves, internode lengths, and enhanced flavonoid accumulation. We studied the effects of abiotic stresses on RNAi and wild-type (WT) plants. Moreover, SlGRAS10-RNAi plants were more tolerant to abiotic stresses (salt, drought, and Abscisic acid) than the WT plants. Down-regulation of SlGRAS10 significantly enhanced the expressions of catalase (CAT), peroxidase (POD), and superoxide dismutase (SOD) to reduce the effects of reactive oxygen species (ROS) such as O2− and H2O2. Malondialdehyde (MDA) and proline contents were remarkably high in SlGRAS10-RNAi plants. Furthermore, the expression levels of chlorophyll biosynthesis, flavonoid biosynthesis, and stress-related genes were also enhanced under abiotic stress conditions. Collectively, our conclusions emphasized the significant function of SlGRAS10 as a stress tolerate transcription factor in a certain variety of abiotic stress tolerance by enhancing osmotic potential, flavonoid biosynthesis, and ROS scavenging system in the tomato plant.
1. Introduction
Transcription factors are involved in plant growth. Among them, transcription factors from the GRAS family were recognized based on DNA binding, transcriptional activation features, and nuclear localization [1]. The GRAS plant proteins are important for controlling several features of development, growth, and retorts to the abiotic and biotic conditions. The GRAS protein family name derived from its major three known participants, i.e., gibberellic acid insensitive (GAI), the repressor of GA1 (RGA), and a scarecrow (SCR) [2,3]. Moreover, SCL14, a GRAS protein is known to be important for the stimulation of stress-inducible promoters in Arabidopsis thaliana [4].
The GRAS protein family is different from other proteins that usually have five conserved motifs in the C-terminus [3]. The functions of all GRAS proteins have not been characterized but usually, GRAS proteins function as transcription factors. A few years ago, several transcription factors have been recognized as fundamental members in controlling plant retorts to stresses [5]. For example, CBF and DREB proteins from the AP2 transcription factors attach with the dehydration response element and regulate the expression of stress-responsive genes [6]. High salinity and drought are the main stress elements persuading adverse effects, including plant development, inhibition of seed germination, and reduced fruit production. Several stress-related genes are persuaded in plants mainly controlled by a series of transcription factors under salt and drought stress [7]. GRAS proteins play a significant function under biotic and abiotic stresses in grape ripening [8]. The GRAS family genes play several functions in plant development and growth including gibberellins signal transduction [9,10], phytochrome A signal transduction [11], root radial organization [12], shoot meristem maintenance [13], male gametogenesis [1], and axillary meristem initiation [14,15]. GRAS family has also been related to abiotic stress response and plant disease resistance [16,17,18]. Suppression of SlGRAS6 tomato plants indicated enhances disease resistance [16]. In rice, over-expression of OsGRAS23 enhanced oxidative and drought stress tolerance [19]. In Arabidopsis, Populus gene PeSCL7 ectopic expression increased salt and drought tolerance [17]. In Arabidopsis, a GRAS transcription factor PAT1 (Vitis amurensis), overexpressing plants conferred more drought, cold, and salinity tolerance [20]. In crops like tomato, analyzing the functions of more GRAS genes will support to explicate the mechanisms regulating stress tolerance and probably help the breeding of tolerant accessions.
Tomato (Solanum lycopersicum) is responsive to a series of environmental stresses. Salinity and drought are key causes of stress that result in deleterious effects, such as decreased plant growth, inhibition of seed germination, and reduced fruit production. A significant study has been shown to understand the physiological characteristics of plants in response to salinity and provide sources to grow salinity-tolerant accessions for plant breeders [21]. In tomato, genome-wide analysis of GRAS proteins has been accomplished, and 53 members of GRAS proteins were recognized [22]. However, the role of several GRAS proteins is still unknown in tomatoes. Previously, our study discussed that SlGRAS7 overexpressing plants showed pleiotropic phenotypes, including dwarf plant height, delay flower time, reduced stem diameter, and many others [23]. Furthermore, SlGRAS7 overexpressing plants could enhance tomato tolerance under drought and salt environments [23]. Several functions of GRAS proteins have been described in other plants; it is significant to examine the roles of the remaining GRAS protein family in the tomato plant. In this research, we described the functional characterization of SlGRAS10 (accession number: Solyc03g025170.1.1), which is from the subfamily of PAT1 protein. To learn about the evolutionary relationship of the SlGRAS10 and other organisms, we had constructed a phylogenetic tree of SlGRAS10 and other plant proteins based on the peptide sequences, indicating that the AtSCL8 is the closest orthologous to SlGRAS10. In our study, expression of SlGRAS10 showed higher levels in leaves and red fruit in WT. Moreover, the transcript accumulation of SlGRAS10 sharply enhanced under salt stress, dehydration, wounding and might be caused by exogenous use of GA3, Abscisic acid (ABA), and ACC. To further study the role of SlGRAS10, we constructed an RNAi vector to produce SlGRAS10 down-regulated transgenic lines which have been resulted in pleiotropic phenotypes. To examine whether down-regulation of SlGRAS10 plants confers abiotic stress tolerance, the SlGRAS10-RNAi transgenic plants treated with salt and drought stress. In present work, expression of SlGRAS10 was remarkably higher under abiotic stress has been determined. Generally, the drought stress and ABA signaling share the same components, so we further observed the SlGRAS10-RNAi transgenic plants treated with ABA stress. Our study examined that the down-regulation of SlGRAS10 plants increased tolerance under abiotic stress by modulating the stress-inducible genes expression that are intricate in physiological modifications, such as flavonoid biosynthesis, reactive oxygen species (ROS) scavenging system, chlorophyll biosynthesis pathway and senescence associated genes. We consider that this study might deliver a different vision of the functional characterization of the GRAS gene family to abiotic stress resistance in tomato and different plant species.
2. Materials and Methods
2.1. Phylogenetic Analysis
2.2. Plant Growth
The seeds of the Wild-type (WT) tomato (cv. Micro-tom) were used in this work. WT and SlGRAS10-RNAi tomato plants were grown in a greenhouse. The plants were grown under the same standard environments as we described previously in our study [23], i.e., 6-h dark (18 °C), 18-h light (25 °C), 60% humidity. Tissues including root, stem, leaves from 4 weeks old WT plants, flowers at anthesis stage, fruits at the immature green, mature green, breaker, breaker plus four days, orange and red stages were collected to perform the gene expression analysis. For each tissue, samples were taken from seven healthy plants. All samples were immediately frozen in liquid nitrogen. Before use, samples were kept at −80 °C temperature.
2.3. Vector Construction and Plant Transformation
The 169 bp SlGRAS10 specific sequences were inserted in the pCAMIBA2301 vector. The pCAMIBA2301 vector was used to construct the SlGRAS10-RNAi plasmid. All the primers are shown in Supplementary Table S1. SlGRAS10-RNAi plants produced via Agrobacterium tumefaciens-mediated transformation. T0 seeds were screened in 1/2 Murashige and Skoog (MS) medium with kanamycin (100 mg L−1). To confirm the existence of T-DNA insertions in several lines of SlGRAS10-RNAi, genomic PCR was performed. After confirmation, three successful RNAi lines (RNAi-10, RNAi-11, and RNAi-15) in the T2 generation were selected for all analyses. To identify the repression ratio of SlGRAS10 in RNAi plants, RT-qPCR has been performed.
2.4. Measurement of Phenotypic Characterization
To analyze the alterations in plant architecture between WT and SlGRAS10-RNAi lines, many parameters including leaf width and length, internode length, and plant height have been measured. Length/width of leaves and internode length were measured from fifteen plants of WT and SlGRAS10-RNAi lines. The plant height was calculated 20 days after germination and after that every 7 days. To calculate the flowering time between WT and SlGRAS10-RNAi lines, the days to first visible flower and leaf number before the first inflorescence were noted. Fifteen plants from three selected lines of SlGRAS10-RNAi and WT were used to measure all the above parameters.
2.5. Plant Stress Treatments
For salt stress treatment assay, the roots of the WT tomato seedlings were immersed in 250 mM NaCl solution for 0, 1, 2, 4, 8, 12, 24, and 36 h. The leaves from the stress-treated seedlings were collected. For the dehydration treatment assay, the WT tomato seedlings were softly pulled out from the soil and washed with water, and leaving on dry filter paper at room temperature. The leaves were collected at 0, 1, 2, 4, 8, 12, 24, and 36 h. For the wounding assay, the leaves of WT seedlings were cut with a blade into small pieces and left those leaves on a piece of wetted filter paper in sealed pots for 0, 1, 2, 4, 8, 12, 24, and 36 h at room temperature. For hormonal stress treatment assay, the whole WT tomato seedlings were sprayed, respectively, with water (control), 100 µM abscisic acid (ABA), 50 µM gibberellic acid (GA3), or 100 µM 1-aminocyclopropane-1-carboxylic acid (ACC) solution [26], then were sealed off in plastic immediately and left for 0, 1, 2, 4, 8, 12, 24 and 36 h at room temperature. Each sample was frozen in liquid nitrogen immediately and stored at −80 °C until RNA extraction. Each stress experiment was done with three replications.
Homozygous T2 transgenic lines of SlGRAS10-RNAi and WT seeds were surface-sterilized and sown on Murashige and Skoog (MS) culture medium. To study the tolerance of seedling development to abiotic stress, transgenic seeds of SlGRAS10-RNAi and WT were transmitted to MS medium containing 0 mM (control), 100 mM sodium chloride (NaCl), and 150 mM D-mannitol for 15 days. The seedlings were incubated in a growth chamber under the same conditions described above and measured the root, shoots growth, and seedling weight.
To study the tolerance of seedling development to ABA stress, transgenic seeds of SlGRAS10-RNAi and WT were transmitted to MS medium containing 0 mM (control), and 100 mM (ABA) for 15 days. The seedlings were incubated in a growth chamber under the same conditions described above and measured the seed germination rate, root lengths, and shoots lengths of SlGRAS10-RNAi and WT.
For the stress experiment, both seedlings of SlGRAS10-RNAi and WT were grown in a growth chamber under normal conditions for two weeks. After that seedlings of SlGRAS10-RNAi and WT were shifted to a greenhouse for 16 h lights (25 °C) and 8 h darks (18 °C). The one-month-old plants of SlGRAS10-RNAi and WT were watered containing 0 (Control), and 200 mM NaCl every 48 h for two weeks to assess the salt tolerance. For drought stress, the one-month-old plants of SlGRAS10-RNAi and WT were treated without water for two weeks. For stress-related gene expression analysis, control and stress-treated leaves of SlGRAS10-RNAi and WT were harvested. To examine the chlorophyll biosynthesis, gene transcription analysis leaves from one-week-old plants after salt and drought stress experiment were collected. The chlorophyll contents and relative water content (RWC) of SlGRAS10-RNAi and WT plants were examined after salt and drought stress treatment. Each stress treatment was completed three times.
2.6. Measurement of Chlorophyll and Relative Water Content
To measure the chlorophyll contents, leaf samples were taken from control, salt, and drought stress treated plants and weighed. All leaf samples were randomly chosen from five individual plants. Samples were crushed with liquid nitrogen and extracted with 80% aqueous acetone (10 mL) (v/v). With the help of a spectrophotometer, the extract was centrifuged at 4000× g for 5 min and the absorbance of the supernatant was noted at 646 and 663 nm. Total Chl (µg mL−1) = 20.29A646 + 8.02A663. According to the method of [27], the measurement of relative water content (RWC) was performed.
2.7. Determination of Antioxidant Enzyme Activities, Proline, MDA, and Soluble Sugar Content
Leaves from control, salt, and drought stress treated plants at the same developmental stage were collected to perform the antioxidant enzyme activities, and Proline, Malondialdehyde (MDA), and soluble sugar contents determination. According to the method [28,29], catalase (CAT), peroxidase (POD), and superoxide dismutase (SOD) were determined. According to the manufacturer’s kit protocol (Jiancheng Bioengineering Company Nanjing, China), the superoxide anion radical (O2−) and H2O2 content were assayed. Proline contents were determined according to the method [30], total MDA contents were measured according to the method [31], and soluble sugar contents according to the method [32].
2.8. Determination of Flavonoid Contents
The flavonoid contents of each sample were determined as described by the Dowd method [33]. Leaves of SlGRAS10-RNAi and WT (2 g/sample) were ground in liquid nitrogen and dissolved in 70% ethanol solution and incubated at room temperature for 24 h. 1 mL of extract ethanol solution were mixed with 0.2 mL of 10% (w/v) AlCl3 solution in methanol, 0.2 mL (1 M) potassium acetate and 5.6 mL distilled water. The mixture was incubated for 30 min at room temperature followed by the measurement of absorbance at 415 nm against the blank. The results were expressed as mg/g of quercetin equivalents in milligrams per gram (mg QE/g) of dry extract.
2.9. RNA Isolation and Real-Time Quantitative PCR Analysis
Total RNAs were isolated from all collected tissues according to the plant RNA kit (OMEGA BIO-TEK). First-strand cDNA synthesis was determined using a PrimeScriptTM RT reagent kit with gDNA Eraser (TAKARA, Kusatsu, Japan). A quantitative Real-Time-PCR (qRT-PCR) was carried out in a Bio-Rad CFX system (Bio-Rad, Hercules, CA, USA) according to the methodology described in [23] using specific primers. Three replicates were used for all samples. SlUBI gene was used for normalization. Relative expression levels were calculated based on the 2−ΔΔCT method. All of the qRT-PCR primers are shown in Supplementary Table S1.
2.10. Statistical Analysis
Each experiment was conducted with three independent biological replicates. The Student’s t-test was used to compare group differences. p-values less than 0.05 were considered to be significant.
3. Results
3.1. SlGRAS10 Is a Member of the PAT1 Subfamily in Tomato
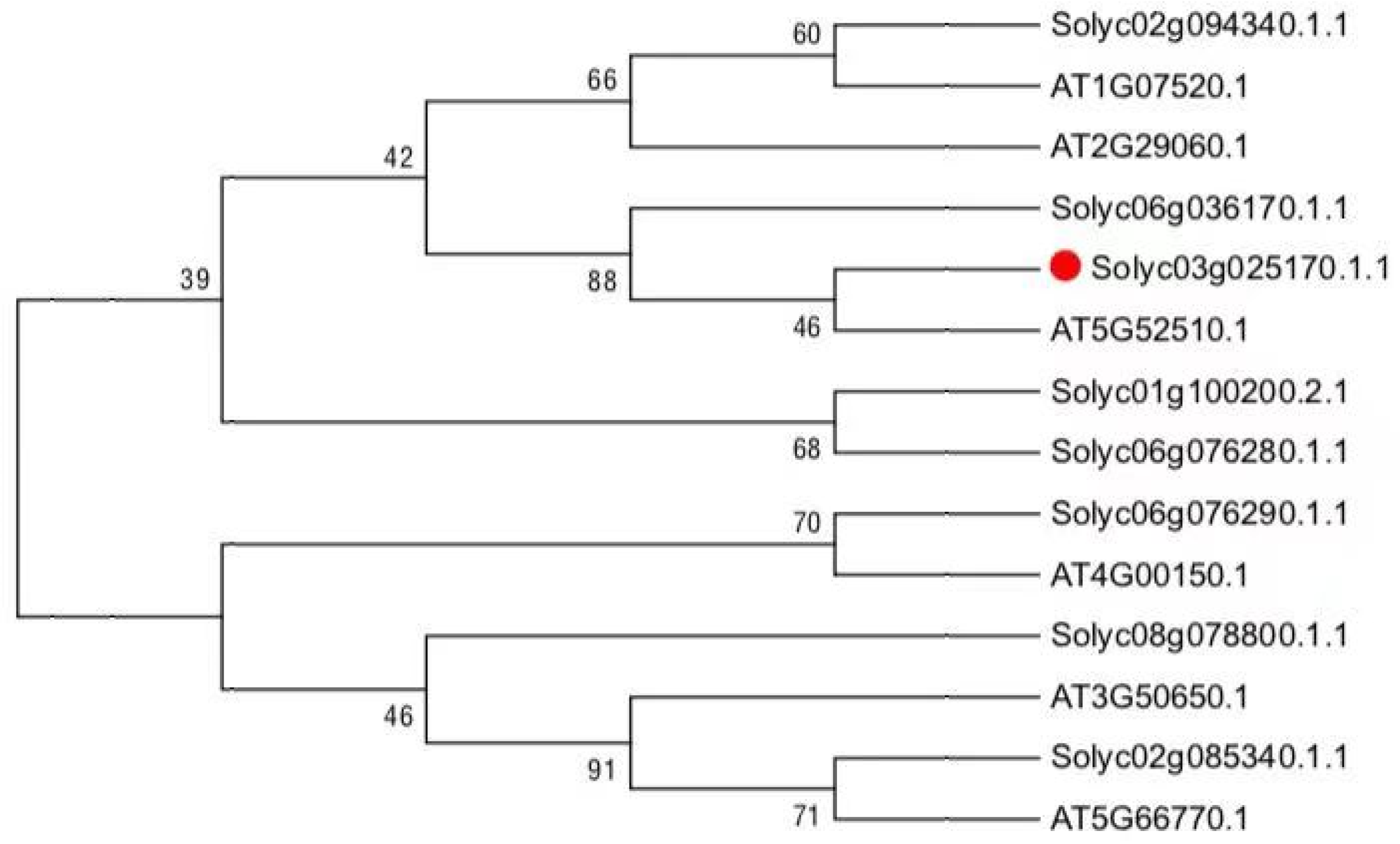
To learn phylogenetic analysis between GRAS family between tomato and Arabidopsis, a phylogenetic tree based on the amino acid sequences was constructed (Figure 1), indicating that the SlGRAS10 belongs to the PAT1 subfamily and is more closely related to AtSCL8 in Arabidopsis thaliana and SlGRAS9 in tomato than to any other proteins among the tree.
3.2. Expression Profiles of SlGRAS10 and under Numerous Stress Treatments of Wild-Type Tomato
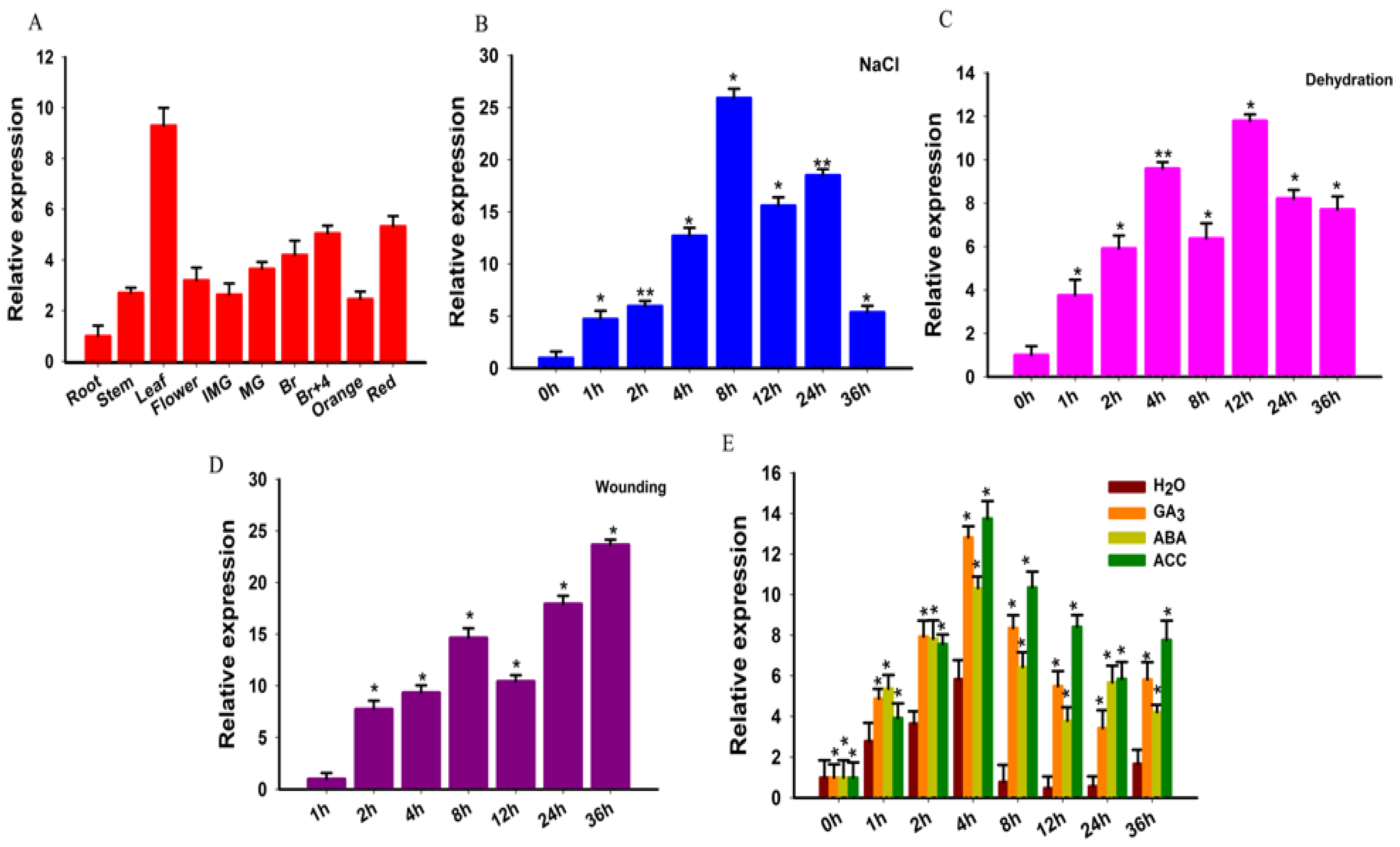
To study the expression profile of SlGRAS10, different samples from wild-type tomato plant were collected such as leaves (L), stems (S), roots (R), flowers (F), immature green fruit (IMG), mature green (MG), breaker (Br), breaker + 4 (Br + 4), and red fruit (R). All tissues for expression pattern analysis frozen in liquid nitrogen and kept at −80 °C (Figure 2A). The primers list is given in Supplementary Table S1. The data indicated that the SlGRAS10 gene expressed in the leaf, breaker + 4, and red fruit, and comparatively lower expression profiles were noticed in other tissues of the tomato plant, indicating its tissue-distinct expression in the tomato plant. The data suggested that SlGRAS10 may be involved in fruit development. To evaluate the dynamic function of phytohormones in the regulation of various developmental and environmental processes, we studied the expression levels of SlGRAS10 under different hormonal applications by quantitative PCR.
To study the role of SlGRAS10 in the stress treatment, the expression profiles of SlGRAS10 in leaves of tomato seedlings were studied under numerous stress treatments. The SlGRAS10 showed its higher expression at 8 h and then decreased under NaCl conditions (Figure 2B). The expression of SlGRAS10 was induced under dehydration treatments and reached a higher value at 12 h (Figure 2C). Under wounding treatments, the expression of SlGRAS10 showed a higher value at 36 h (Figure 2D). These outcomes propose that SlGRAS10 might play an important role under abiotic stresses. Assuming that phytohormones are associated with intricate signaling pathways and involved in regulating plant reaction to numerous environmental stresses and developmental processes [34], we further investigated the expression patterns of SlGRAS10 under several hormonal applications by quantitative RT-PCR. SlGRAS10 was remarkably induced by ACC application and showed a higher value of 13.7-fold at 4 h, and then decreased but remained at the highest level amongst all hormonal treatments (Figure 2E). The expression of SlGRAS10 was at a maximum peak at 4 h under the ABA application. For GA3, the expression profile of SlGRAS10 increased distinctly at 4 h and showed a higher value of 12.8- fold. The results of expression levels for SlGRAS10 showed that the SlGRAS10 gene may be involved in signal transduction and hormonal stress.
3.3. Phenotypic Characterization of SlGRAS10-RNAi Plants
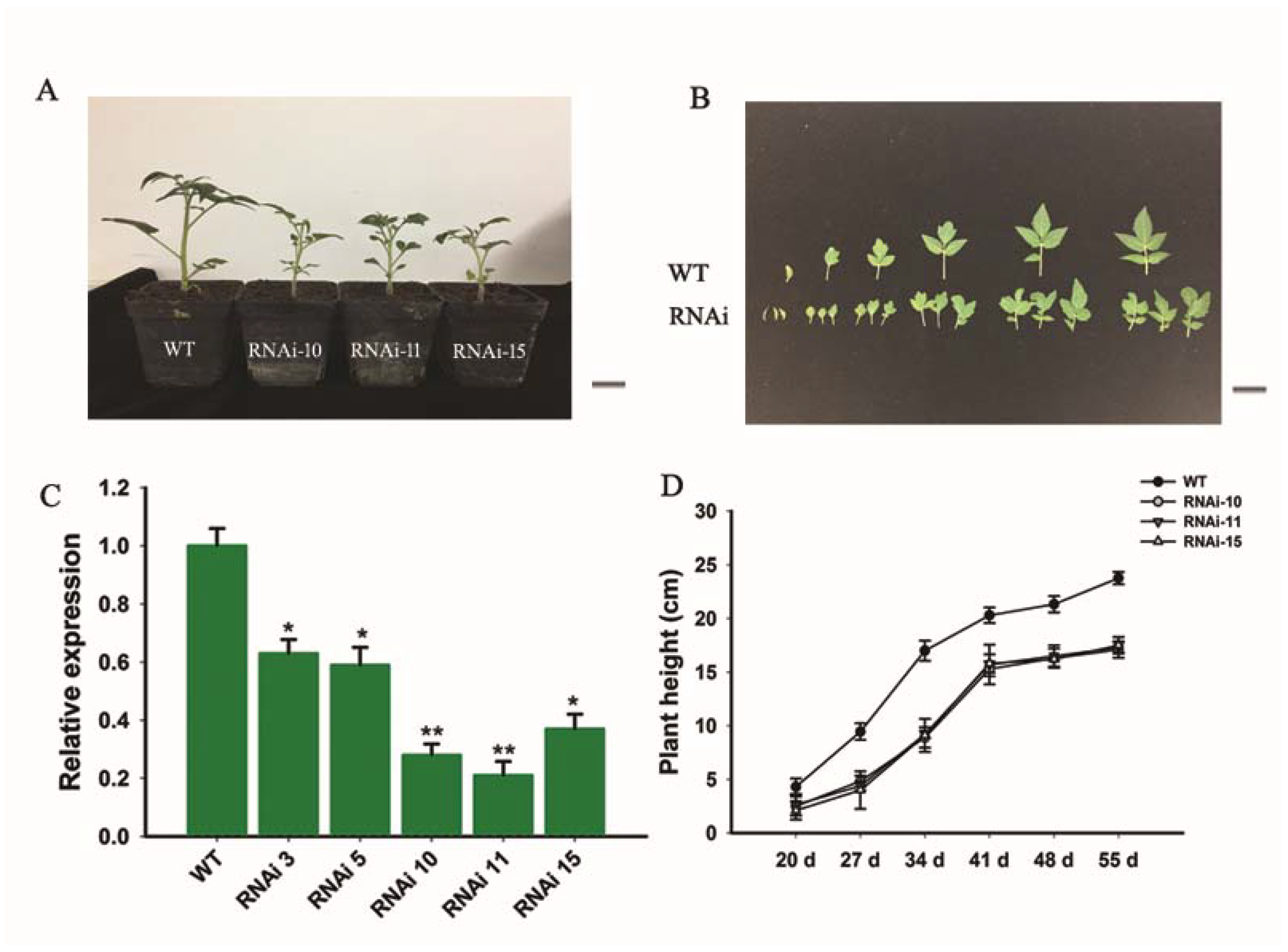
To study the physiological importance of the SlGRAS10 gene, the transgenic tomato plants were produced by RNAi silencing by transformation with Agrobacterium tumefaciens. WT and SlGRAS10-RNAi (L10, L11, and L15) plants are shown in Figure 3A. We found five independent transgenic lines, and the expression pattern of SlGRAS10 was found in all these transgenic lines. The expression patterns of SlGRAS10 in transgenic lines were reduced as compared to WT (Figure 3C). Three out of five transgenic lines of SlGRAS10 were selected due to their lowest expression levels and named RNAi10, RNAi11, and RNAi15. These three transgenic lines of SlGRAS10-RNAi10, SlGRAS10-RNAi11, and SlGRAS10-RNAi15 were examined for more phenotypic and molecular investigations.
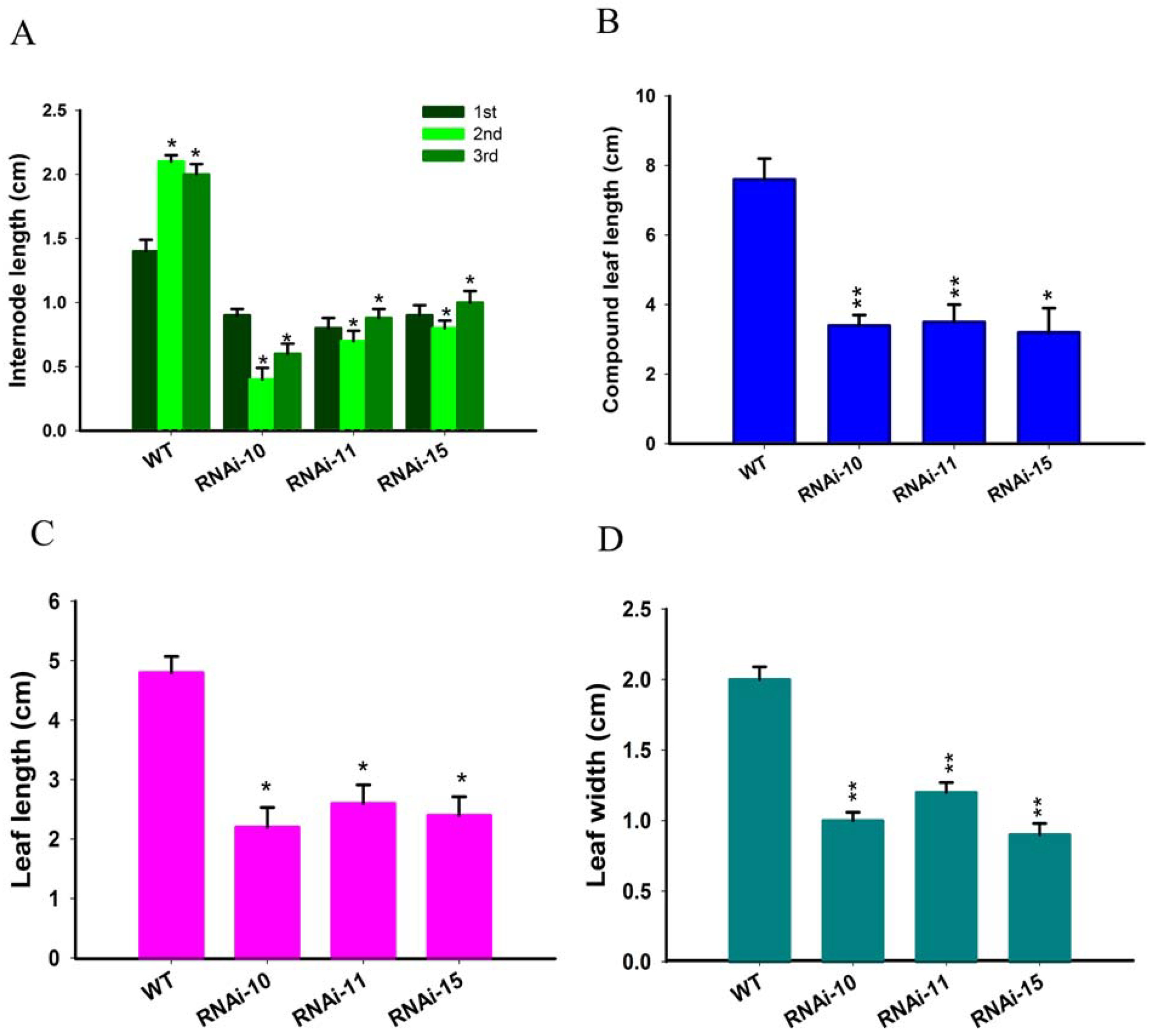
We noted that selected transgenic lines of SlGRAS10-RNAi displayed common phenotypes related to vegetative growth, including short height plants (Figure 3A), and smaller leaves (Figure 3B). We observed and measured the plant height and first three internode lengths of SlGRAS10-RNAi (RNAi10, RNAi11, and RNAi15) and WT. The SlGRAS10-RNAi plants were remarkably smaller than the height of WT plants (Figure 3D). Similarly, the three internode lengths in three SlGRAS10-RNAi lines were smaller than the internode lengths of WT (Figure 4A). The length of compound leaves (Figure 4B), and the length (Figure 4C) and the width (Figure 4D) of mature leaflets were remarkably decreased in three lines of SlGRAS10-RNAi compared to WT.
3.4. SlGRAS10-RNAi Enhances Tolerance of Seedling Growth to Salt and Drought Stress
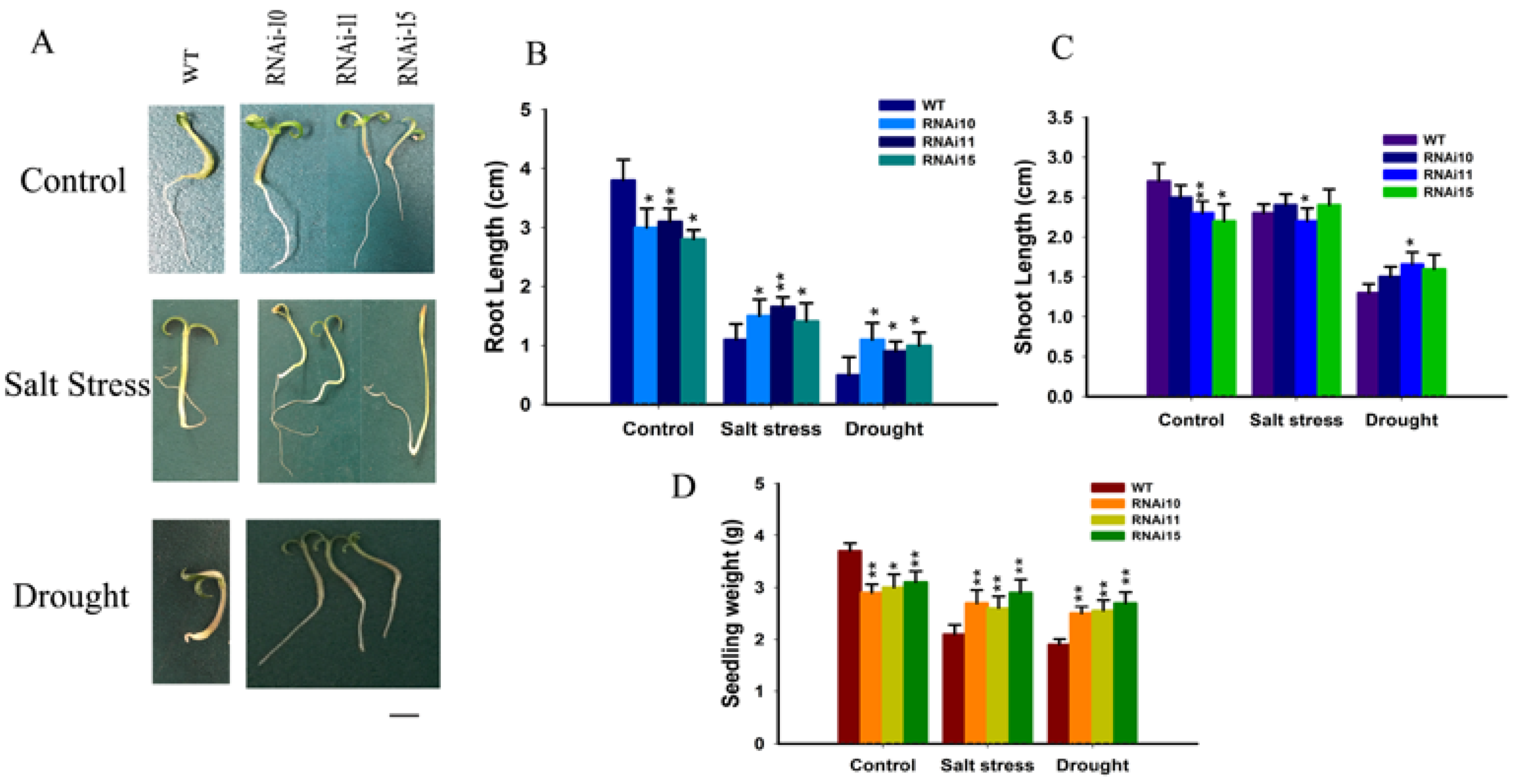
The substantial stimulation of SlGRAS10 expression by salt stress and dehydration showed that it might be playing a significant function in salt stress and drought tolerance of tomatoes (Figure 2B,C). At the stage of post-germination, we studied the different influences of salt stress on development between WT and SlGRAS10-RNAi tomato seedlings to assess the resistance of WT and SlGRAS10-RNAi tomato towards salt stress and drought. Root length was significantly suppressed in WT as well as SlGRAS10-RNAi under salt and drought stress, but the root length of SlGRAS10-RNAi seedlings was much greater than those of WT (Figure 5B). There is no obvious change in shoot length of WT and SlGRAS10-RNAi seedlings under salt stress but the shoot length of WT seedlings was more inhibited under drought stress than those of SlGRAS10-RNAi (Figure 5C). The seedling weight of SlGRAS10-RNAi was greater than the weight of WT seedlings under salt and drought stresses (Figure 5D). The growth of WT seedlings was more reduced, and the root growth of WT seedlings was significantly inhibited as compared to SlGRAS10-RNAi seedlings, showing that silencing of SlGRAS10 enhances seedling growth more tolerant to salt and drought stress. Furthermore, the seedlings of SlGRAS10-RNAi showed more tolerance in fresh weight than WT seedlings when treated with salt and drought stress (Figure 5D).
3.5. SlGRAS10-RNAi Enhances Tolerance of Seedling Growth to ABA Stress
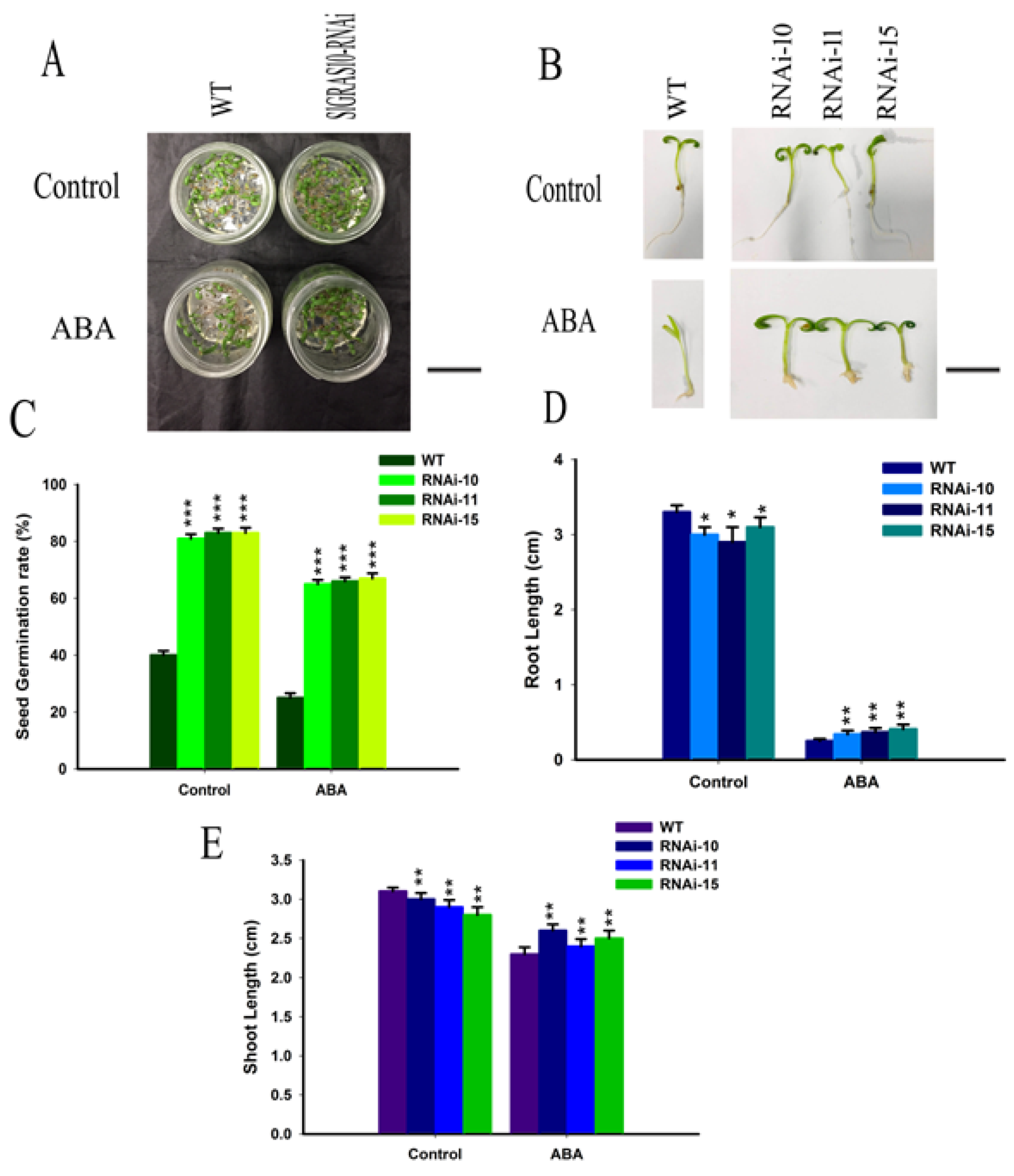
The expression levels of SlGRAS10 are induced by ABA (Figure 2E), it might be playing a significant function in ABA stress. At the stage of post-germination, we studied the ABA stress on the development between WT and SlGRAS10-RNAi tomato seedlings to assess the resistance of WT and SlGRAS10-RNAi tomato towards ABA stress. The seed germination rate was significantly decreased in WT in response to ABA stress, but the seed germination rate of SlGRAS10-RNAi was much greater than that of WT seeds under ABA stress (Figure 6C). Root elongation of both WT and SlGRAS10-RNAi was affected in response to ABA stress but the root length of SlGRAS10-RNAi was slightly higher than that of the WT under ABA stress (Figure 6D). The shoot lengths of the SlGRAS10-RNAi were greater than those of the WT under ABA stress (Figure 6E). Together, these results showed that the seeds and seedlings of SlGRAS10-RNAi tolerate ABA stress.
3.6. SlGRAS10-RNAi Enhances Tomato Plant Tolerance to Salt and Drought
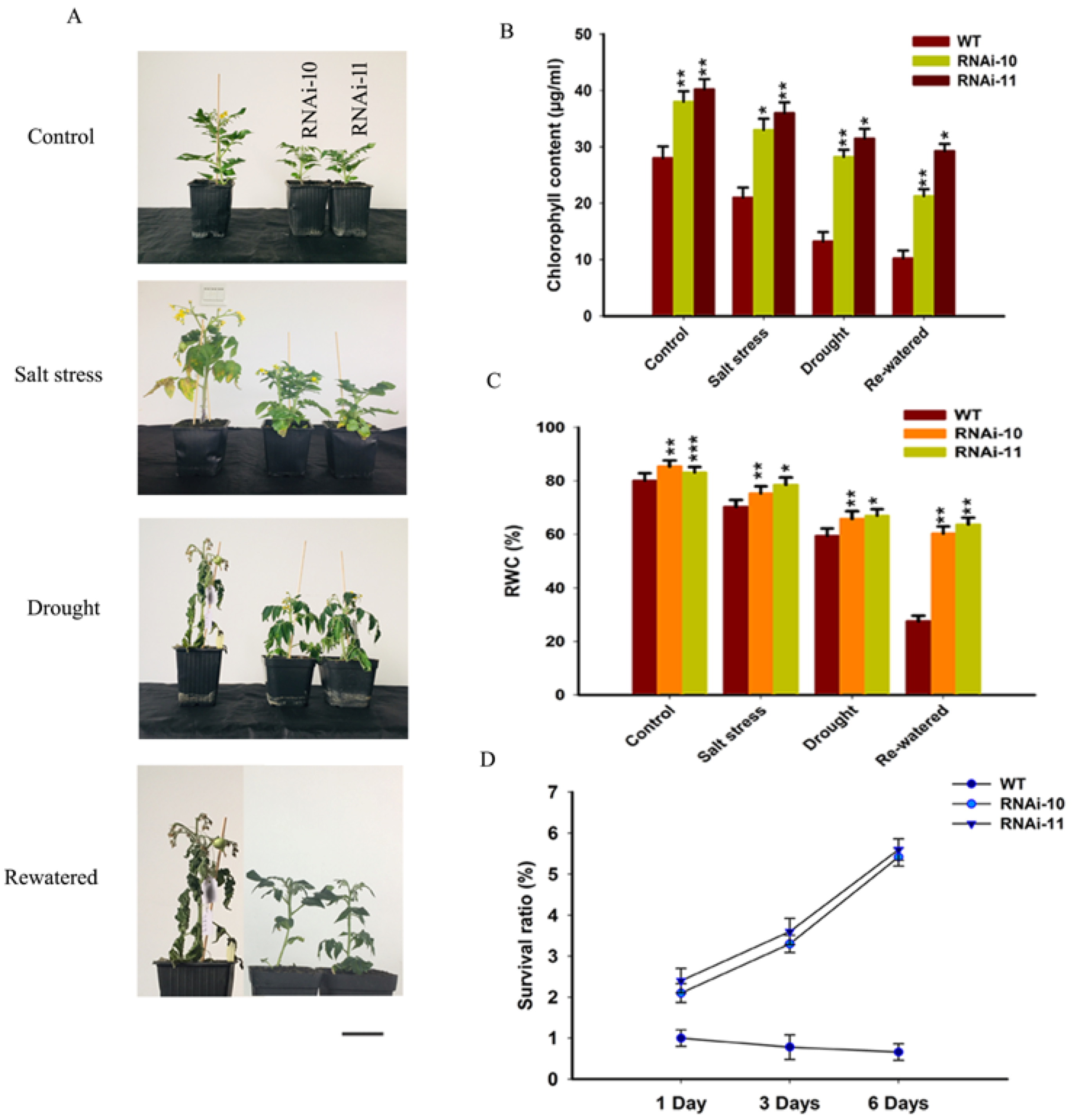
To assess the function of SlGRAS10-RNAi lines in response to salt and drought stresses in soil, one-month-old plants of WT and SlGRAS10-RNAi (L10, L11) were irrigated with water containing 0 (control), 200 mM NaCl every 48 h for 2 weeks to observe salt stress tolerance. To study drought stress tolerance, one-month-old plants of WT and SlGRAS10-RNAi (L10, L11) were deprived of water for up to 2 weeks. The SlGRAS10-RNAi (L10, L11) and WT plants showed significant morphological alterations under stress conditions (Figure 7A). All SlGRAS10-RNAi plants of L10 and L11 were healthier than WT plants in response to salt and drought stress (Figure 7A). However, SlGRAS10-RNAi plants showed slight alterations in their phenotype compared with WT plants under both stresses (Figure 7A). Under salt stress treatment, SlGRAS10-RNAi plants showed less necrosis and chlorosis than the WT plants. WT plants showed typical severe desiccation symptoms under drought stress. All leaves of WT plants were wilted, but there are only lower leaves were wilted in SlGRAS10-RNAi plants under drought stress (Figure 7A). The results indicated that both stress damage in WT plants was more severe than in SlGRAS10-RNAi.
Furthermore, we investigated the total chlorophyll content and relative water content (RWC) in SlGRAS10-RNAi and WT plants under control and stresses conditions. The levels of chlorophyll content and relative water content were much higher in SlGRAS10-RNAi plants than in WT plants under both stress conditions (Figure 7B,C). Together, these results help to provide a suggestion that the SlGRAS10-RNAi confers tolerance under salt and drought stress treatment in tomatoes.
3.7. Enhanced Antioxidant Activity in SlGRAS10-RNAi Plants under Stress Conditions
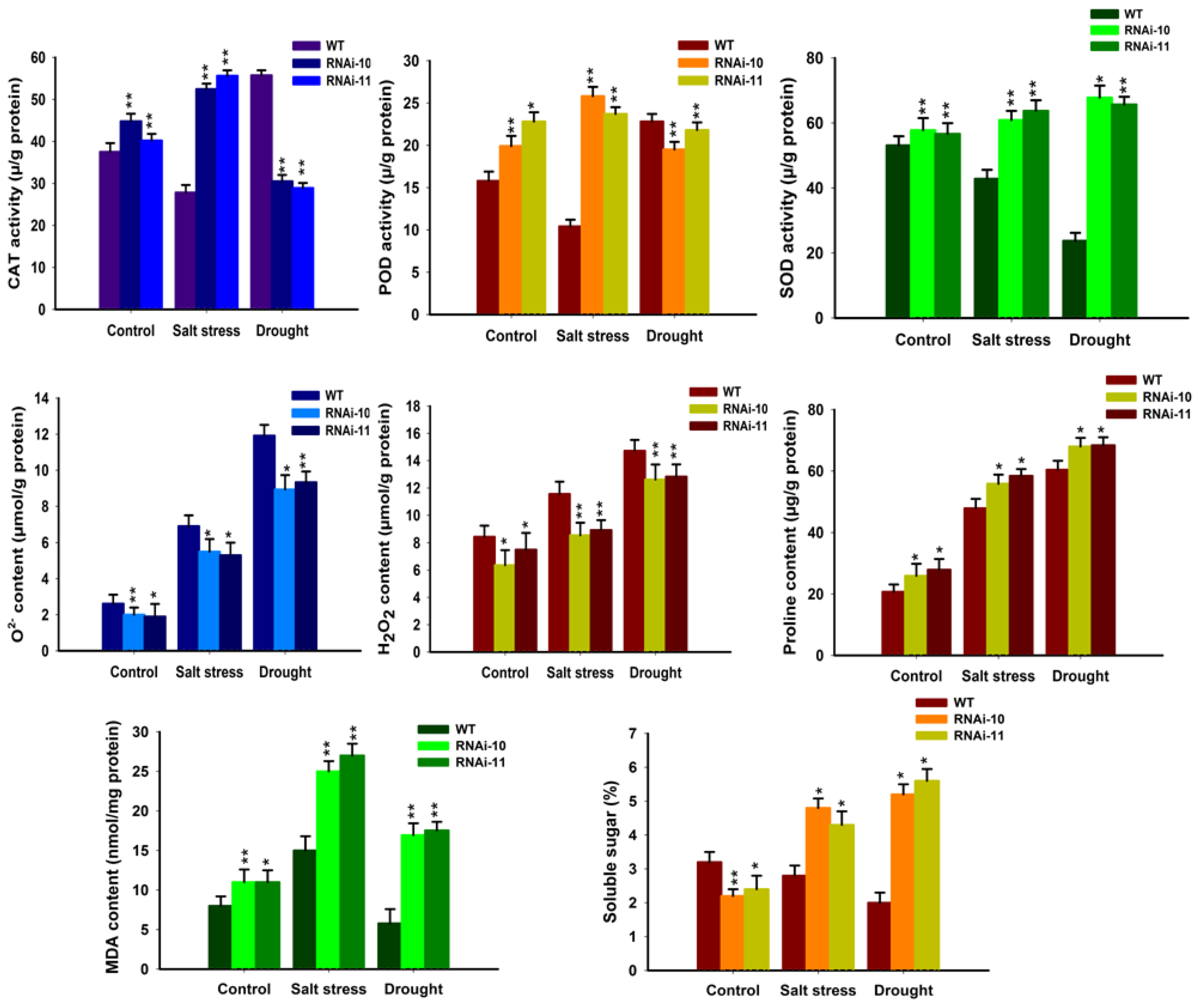
Salt and drought stresses lead to abundant reactive oxygen species (ROS), which harms membrane structure [35]. It was examined that the alterations in the accretion of O2− and H2O2 in WT and SlGRAS10-RNAi plants under salt and drought stresses. It was observed that less O2− and H2O2 gathered in SlGRAS10-RNAi plants than in WT under control and stress conditions (Figure 8). To study the likely physiological mechanism responsible for the enhanced salt and drought tolerance, we examined the alterations in MDA concentration, soluble sugar contents, and total proline contents from SlGRAS10-RNAi and WT plants grown under control and abiotic stress. Total MDA contents were remarkably high in SlGRAS10-RNAi plants than WT plants under control and both stress conditions (Figure 8), suggesting that the SlGRAS10-RNAi induces potential antioxidative processes preventing significant membrane damage. Under salt and drought stresses, we observed that total soluble sugars were collected more in SlGRAS10-RNAi plants than in the WT (Figure 8). Under control and stress conditions, the expression level of total proline contents was higher in SlGRAS10-RNAi plants than in WT (Figure 8). We observed that the RNAi plants accrued more proline contents under both stress conditions than control (Figure 8). Moreover, we studied the antioxidants enzyme activities such as SOD, POD, and CAT from SlGRAS10-RNAi and WT plants under control, salt, and drought stresses. The expressions of CAT and SOD activities in SlGRAS10-RNAi and WT plants were approximately similar under control conditions. For salt stress conditions, the expression pattern of SOD, POD, and CAT were higher in SlGRAS10-RNAi than in the WT plants (Figure 8). Under drought stress conditions, the expression pattern of POD and CAT activities in SlGRAS10-RNAi plants were down-regulated as compared to WT plants but SOD activity was remarkably higher in SlGRAS10-RNAi plants (Figure 8). Overall, these results recommend that SlGRAS10 may help to enhance the tolerance of transgenic plants to salt and drought stresses.
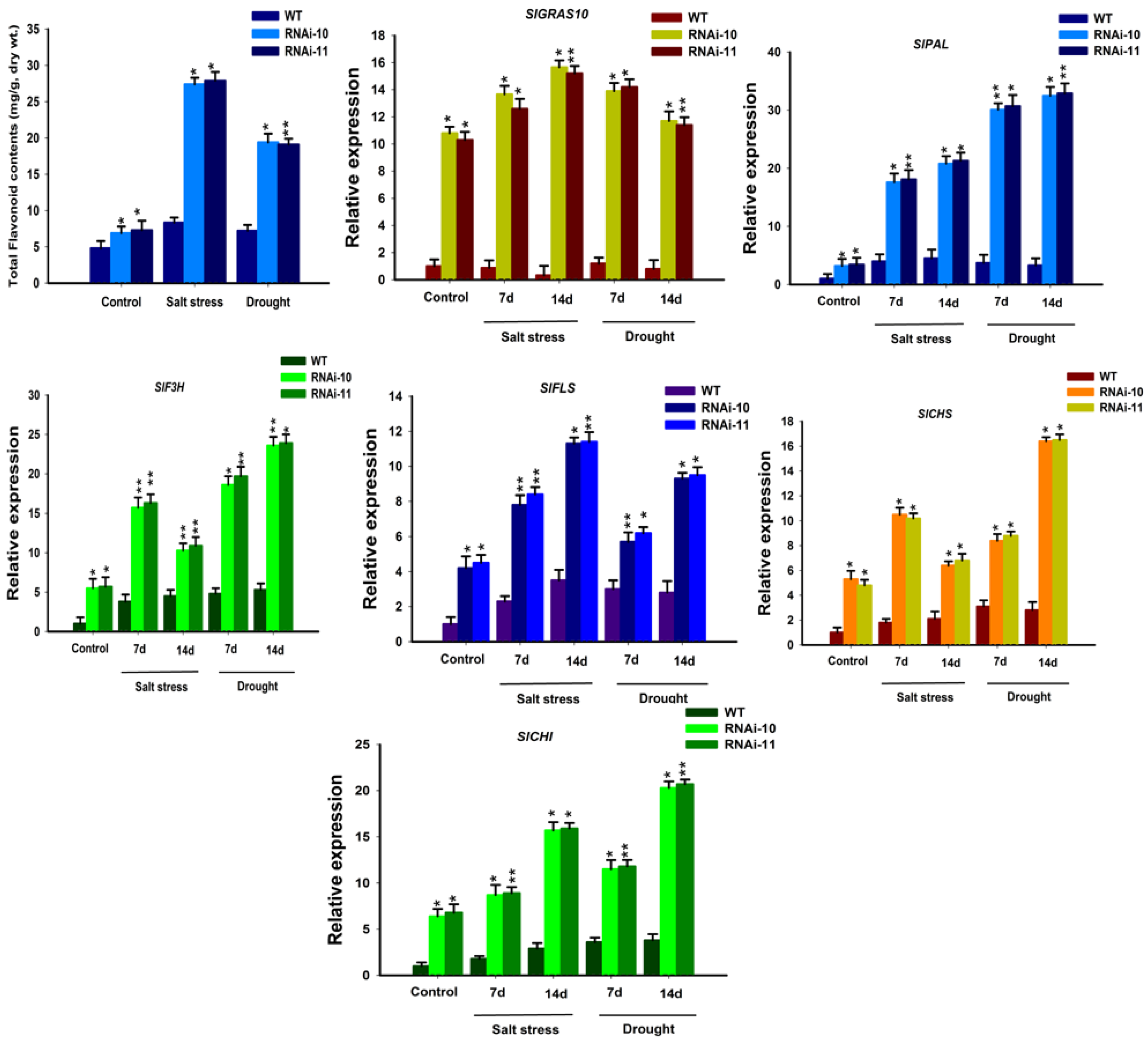
3.8. Up-Regulation of Flavonoid Accumulation in SlGRAS10-RNAi under Stress Conditions
The flavonoid contents were determined in SlGRAS10-RNAi and WT plants under control, salt, and drought stress. It was examined that the expression levels of total flavonoid contents in SlGRAS10-RNAi plants were greater than the WT plants under control and both stresses (Figure 9). Moreover, the flavonoid biosynthesis pathway genes were examined in the SlGRAS10-RNAi and WT plants. The expression patterns of the flavonoid biosynthesis pathway genes (SlPAL, SlF3H, SlFLS, SlCHS, and SlCHI) were up-regulated in SlGRAS10-RNAi compared to WT (Figure 8). The expression of SlGRAS10-RNAi plants was significantly up-regulated than the WT plants in response to salt and drought stress (Figure 9). Our data outcomes showed that SlGRAS10 may induce flavonoid accretion by modulating the flavonoid biosynthesis pathway.
3.9. Enhanced Expression of Chlorophyll Biosynthesis-Related Genes in SlGRAS10-RNAi under Stress Conditions
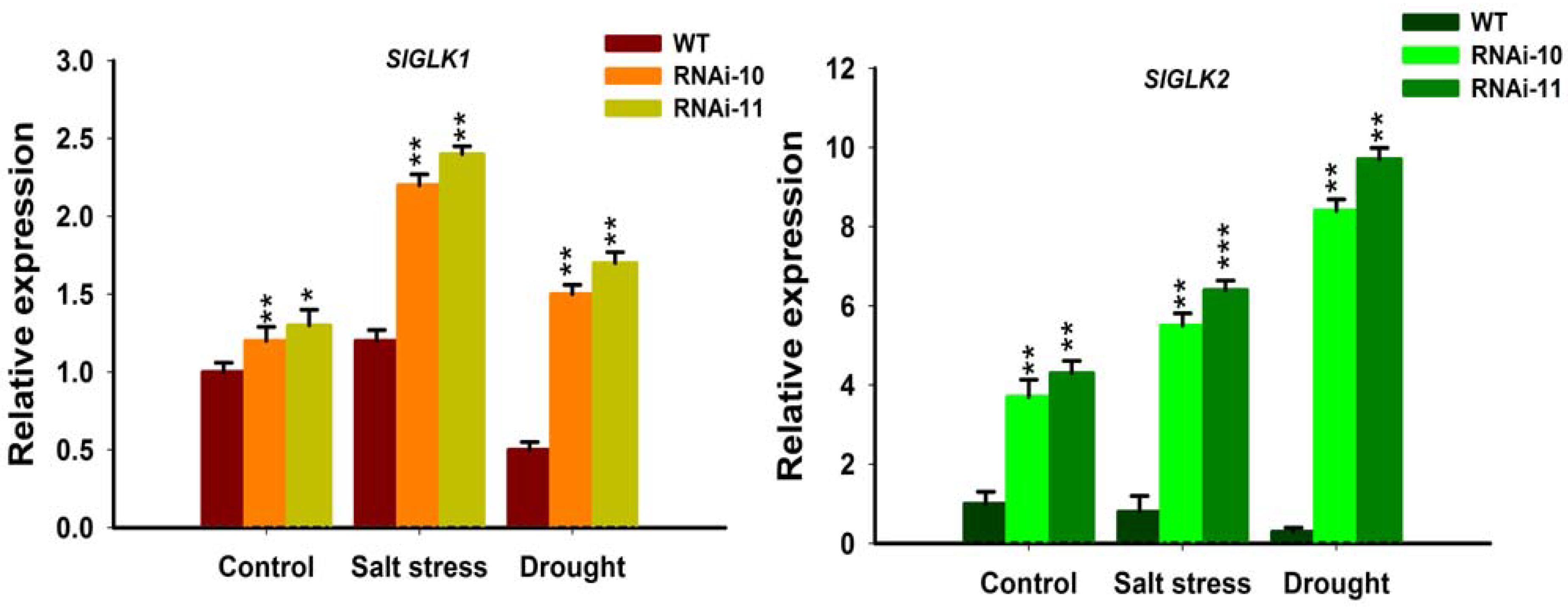
Alteration of chlorophyll contents in the SlGRAS10-RNAi after treatment with salt and drought stress provoked us to examine the influence of the abiotic stresses on chlorophyll biosynthesis at the transcriptional level. Thus, we studied the expressions of both Golden2-like1 (SlGLK1) and Golden2-like2 (SlGLK2) in leaves of SlGRAS10-RNAi and WT plants, suggesting that SlGLK1 and SlGLK2 are up regulators of the chlorophyll biosynthesis mechanism. In our results, the relative expressions of SlGLK1 and SlGLK2 genes showed higher expressions in SlGRAS10-RNAi plants than in the WT under salt and drought stresses (Figure 10). Our results suggest that SlGRAS10 enhanced the expression of chlorophyll biosynthesis genes by modulating the chlorophyll biosynthesis pathway.
3.10. Expression Analysis of Senescence-Associated Genes in SlGRAS10-RNAi under Stress Conditions
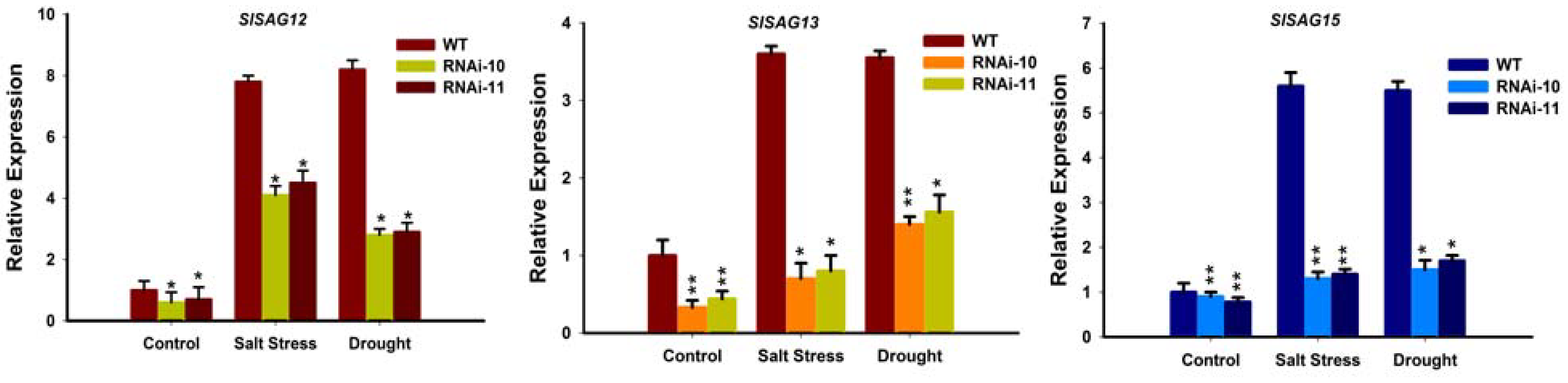
As the Chlorophyll level was altered in SlGRAS10-RNAi as compared to WT plants under stress conditions. Thus, we studied the senescence-associated genes SlSAG12, SlSAG13, and SlSAG15 in the leaves of SlGRAS10-RNAi and WT plants. Leaves from the three-month-old plants of SlGRAS10-RNAi lines and WT plants were used for senescence assay. qRT-PCR results showed that the expression levels of the senescence-associated genes in SlGRAS10-RNAi lines were down-regulated under control conditions as compared with the WT, indicating that the SlGRAS10-RNAi negatively regulates the leaf senescence in tomato (Figure 11). The expression levels of the SlSAG12, SlSAG13, and SlSAG15 were slightly higher under salt stress and drought but significantly lower than the levels of the WT (Figure 11). Together, our results suggest a delay in senescence in SlGRAS10-RNAi plants.
3.11. Expression Analysis of Stress-Related Genes in SlGRAS10-RNAi and WT Plants under Abiotic Stress Conditions
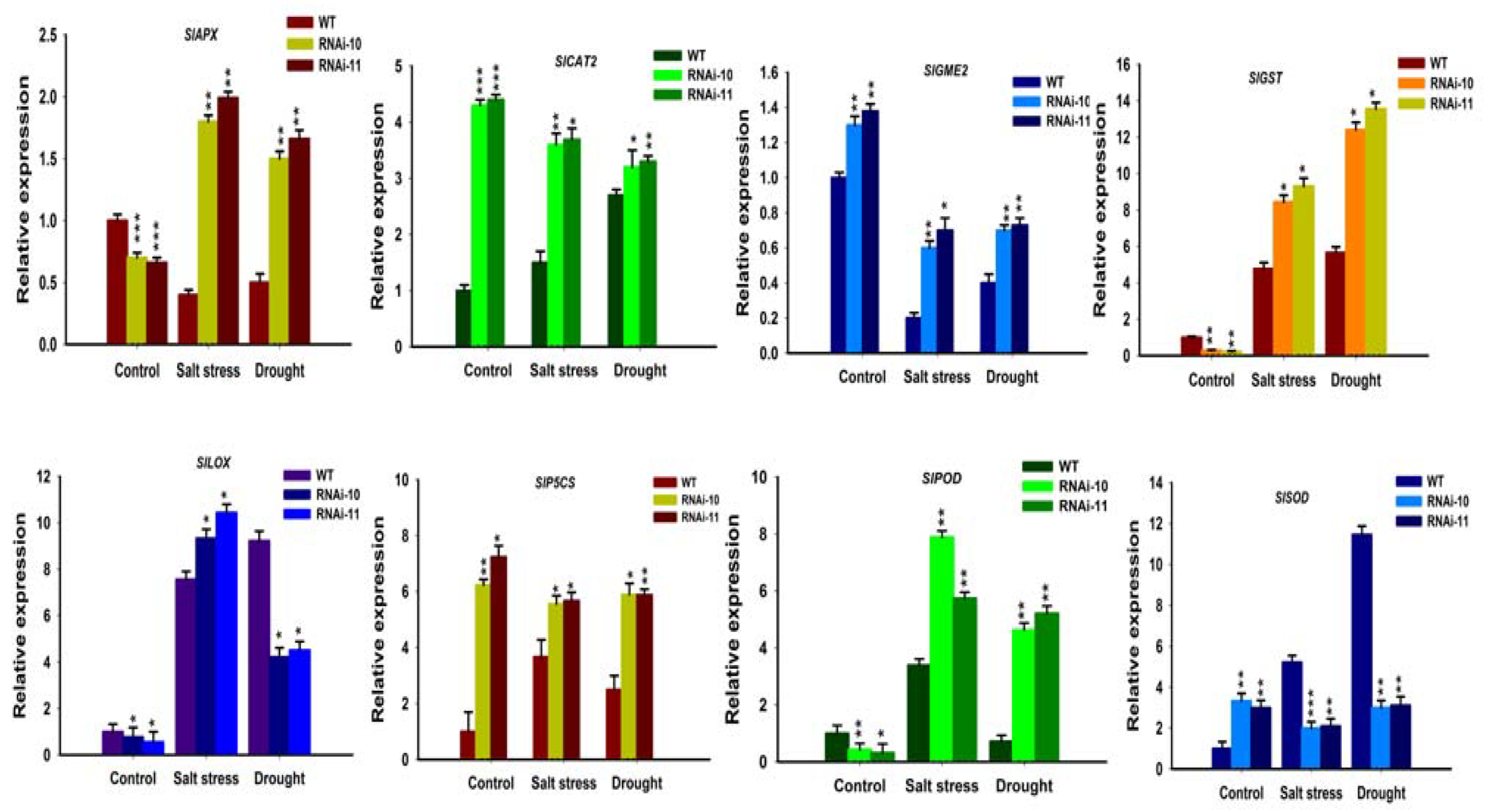
To study the molecular pathways, the expression levels of the stress-related genes were studied by real-time qPCR in SlGRAS10-RNAi lines and WT under salt and drought stress. A key ascorbic acid synthetase (GME2), a key proline synthetase (P5CS), a catalase (CAT2), glutathione S-transferase (GST), superoxide dismutase (SOD), peroxidase (POD), lipoxygenase (LOX), and an ascorbate peroxidase (APX) were studied for SlGRAS10 transcription factor. The relative expressions of SlAPX and SlGST genes showed higher expressions in the SlGRAS10-RNAi plants than the WT under salt and drought stress (Figure 12). The SlPOD gene indicated higher expression levels in SlGRAS10-RNAi plants than in WT after both stress treatments (Figure 12). SlCAT2 and SlP5CS were up-regulated after control conditions in SlGRAS10-RNAi plants, but after salt and drought stress treatment, the expression levels of SlCAT2 and SlP5CS were significantly reduced in WT than SlGRAS10-RNAi plants (Figure 12). The SlLOX gene indicated higher expressions under salt stress conditions in SlGRAS10-RNAi plants compared to WT (Figure 12). Together, our results showed that SlGRAS10 may play role in inducing the gene expression which is involved in stress signaling pathways in tomato.
4. Discussion
Environmental stresses such as salt and drought are primary issues that considerably decrease agricultural yield. The progress in stress tolerates crops will be significantly helpful for current agrarian zones which are likely to abiotic stresses. Plants come across a series of stresses, including drought, salinity, cold, heat, and light so on. Such kinds of environmental stresses badly affect the growth, productivity, and survival ratio of plants [36]. Plants improve their numerous mechanisms and pathways to oppose these stress conditions, which allow them to take on under such environments [37]. Molecular plant biology techniques allow us to determine numerous genes, such as transcription factors which play dynamic functions in temporal and spatial genes expression under stress tolerance which might be caused by one or more biotic and abiotic stresses [38]. From the GRAS protein family in tomato, eleven proteins belong to the PAT1 branch. In this study, the SlGRAS10 gene from the PAT1 branch is functionally described. Our study elucidated the function of a SlGRAS10 in tomato confers abiotic stress tolerance. The tissue expression pattern showed that SlGRAS10 was mainly found in leaves. SlGRAS10 was responsive to salt stress (NaCl), dehydration, wounding, and hormonal treatment (GA3, ABA, and ACC) (Figure 2). Our results suggested that SlGRAS10 may be intricate in leaf growth, abiotic stress-responsive, and mediate hormone signaling.
Down-regulation of SlGRAS10 significantly reduced the plant height (Figure 3A), leaf size as well as internode lengths of tomato (Figure 4A,C,D). In our previous study, overexpressing plants of SlGRAS7 enhanced tolerance to abiotic stress [23]. In the present work, physiological, morphological, and molecular data confirmed that the SlGRAS10 gene confers salt and drought stresses. We produced tomato trans-genetic plants with down-regulation of SlGRAS10 by the RNAi method and showed comprehensive physiological studies. First, we examined the seedling growth of SlGRAS10-RNAi and WT under salt and drought stress and found that SlGRAS10-RNAi enhances seedling growth more tolerate under salt and drought stress than WT (Figure 5A). Our results demonstrated the down-regulation of SlGRAS10 plants more tolerant under salt and drought stress. The immersion of SlGRAS10 tomato plants in response to abiotic stress proved that down-regulation of SlGRAS10 conferred the tomato plants with greater tolerance and responsiveness under salt and drought stress than WT. Therefore, these results appeared to be a positive relationship between the SlGRAS10 and abiotic stress in RNAi lines.
ABA plays a vital role in response to abiotic stress, especially in water-deficient environments, where plant species elevate the cellular level of ABA to combat drought stress. Under drought stress, NCED3 is a dominant contributor to ABA production in Arabidopsis. ABA regulates the dormancy and germination of the seed, inhibiting root growth in response to salinity and drought stresses [39,40]. In our study, we examined the biological parameters in SlGRAS10-RNAi and WT seedlings under control and ABA stress. The SlGRAS10-RNAi seedlings showed a higher germination rate as compared to WT in response to ABA stress (Figure 6). We observed that the root and shoot lengths of WT seedlings are much lower than those of the SlGRAS10-RNAi seedlings. Thus, these results lead to an indication of a positive relationship between the SlGRAS10 and ABA stress in RNAi-lines.
Environmental stresses generally cause several biochemical and physiological modifications in plants. Therefore, we more examined the physiological base responsible for the modifications in tolerance of SlGRAS10-RNAi and WT plants to salt and drought stress. For example, chlorophyll, MDA, and RWC contents are common parameters for assessing abiotic stress resistance and tolerance in plants [41,42,43,44]. In this work, we examined the direct relationship between modifications in SlGRAS10 expression and the biological parameters related to stress tolerance. The SlGRAS10-RNAi plants showed less chlorosis and necrosis than in WT plants after treated with salt, drought stress and after re-watered drought plants (Figure 7A). We examined that the SlGRAS10-RNAi plants collected more chlorophyll contents than WT plants under abiotic stress and re-watered drought plants (Figure 7B). Moreover, water contents in SlGRAS10-RNAi plants were higher than WT plants under salt, drought stresses, and re-watered drought plants (Figure 7C). Moreover, SlGLK1 and SlGLK2 (chlorophyll biosynthetic genes) showed higher expression in SlGRAS10-RNAi lines under control, and both abiotic stresses (Figure 10). Together, these outcomes help to support the argument that the increased tolerance of SlGRAS10 down-regulated plants is associated with more accretion of chlorophyll and RWC contents.
Abiotic stresses generally induce accretion of ROS and lipid peroxidation which lead to oxidative stress [45]. MDA is a decomposition product of polyunsaturated fatty acids and has been commonly used as a parameter for lipid peroxidation [46]. In this study, we examined that SlGRAS10-RNAi lines showed more accretion of MDA contents than WT plants when treated to salt and drought stress (Figure 8). This might be because of the protective function of antioxidant enzymes in SlGRAS10-RNAi lines, suggesting that the synergistic effect of various ROS-scavenging enzymes can effectively improve the antioxidant capacity of plants, thereby reducing the degree of oxidative stress in SlGRAS10-RNAi lines. Thus, these results suggest that SlGRAS10 may confer more tolerate towards oxidative stress in plants. We believed that the down-regulation of SlGRAS10 improves the ability of transgenic plants to scavenge reactive oxygen species. Antioxidant enzymes such as CAT, POD, and SOD reduce oxidative and osmotic deterioration to cellular enzymes, plasma membrane proteins, and integrity in plants [35,47]. The down-regulation of SlGRAS10 plant lines showed more accumulation of proline and total soluble sugar contents than WT towards salinity and drought stress (Figure 6). SlGRAS10-RNAi lines showed a higher accumulation of H2O2 and O2− contents than the WT plants after treated to both stresses (Figure 8). Probably, soluble sugar and proline contents secure SlGRAS10-RNAi against ROS, reducing oxidative loss. ROS scavenging enzymes, for example, POD, SOD, CAT, GST, and APX reduce oxidative loss and increase abiotic stress tolerance in plants [48]. Here, the expression levels of CAT and POD were higher in SlGRAS10-RNAi plants than in WT under control and salt stress treatments (Figure 8), and the SOD activity in SlGRAS10-RNAi plants was up-regulated under control and both stresses (Figure 8). These results showed that the ROS scavenging ability in SlGRAS10-RNAi plants was greatly improved and the down-regulation of SlGRAS10 in tomato plants confers abiotic stress tolerance. Plants are often affected by biotic and abiotic environments, and therefore they have developed distinguished physiological and developmental pathways by gene expression regulation under biotic and abiotic stresses [49]. These results showed that the expression of stress-related genes such as SlAPX, SlGST SlPOD, SlCAT2, SlGST SlP5CS, and SlLOX genes in SlGRAS10-RNAi plants was much higher under both stress conditions than in WT (Figure 12). Thus, we examined that the increased tolerance to abiotic stress is primarily attributable to considerably higher expression levels of stress-responsive genes. Together, the expression level alterations of stress-related genes showed a dynamic regulatory function of the SlGRAS10 under abiotic stress environments, and certainly influence salt stress tolerance.
Flavonoids belong to a group of secondary metabolites that have the distinct antioxidant capability in plants [50]. These secondary metabolites can improve plant tolerance under abiotic stresses because of their ability to reduce free radicals, peroxides, superoxide which were accumulated during stress conditions [51,52]. Therefore, we supposed that the accretion of secondary metabolites (flavonoids) can increase tolerance to oxidative and osmotic stress. It was determined that down-regulation of SlGRAS10 plants accumulates more flavonoids, as well as the flavonoid biosynthesis pathway genes, showed higher expressions in SlGRAS10-RNAi lines in response to salt and drought stress conditions than in WT (Figure 9). These findings suggest that up-regulation of flavonoid accumulation leads to enhanced tolerance to oxidative stresses.
Therefore, together our study provides evidence that the down-regulation of SlGRAS10 confers abiotic stress tolerance in tomatoes. The higher expression levels of flavonoid biosynthesis genes, chlorophyll biosynthesis genes, and ROS scavenging genes may cause an increase in the capability of SlGRAS10-RNAi tomato plants to deal with related stresses.
5. Conclusions
In summary, our present work concerned the physiological and morphological aspects of SlGRAS10-RNAi transgenic tomato under abiotic stress environments, including molecular mechanisms for expression levels of stress-related, chlorophyll, senescence-associated, and flavonoid biosynthesis genes. We concluded that the down-regulation of the SlGRAS10 gene improves tolerance to abiotic stresses by increased chlorophyll contents, flavonoid accumulation, and ROS scavenging enzymes. Therefore, these physiological alterations improve and enhance the tomato plant’s ability to sustain under abiotic stress environments. Our work provides evidence of GRAS functions against abiotic stresses as and helps to enhance our knowledge about their functions under abiotic stresses.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/genes12050623/s1, Table S1: List of primers used in cloning and qRT-PCR.
Author Contributions
Conceptualization, S.H.; methodology, S.H.; software, S.H. and Y.Y.L.; validation, Y.Y.L., and N.L.; formal analysis, S.H.; investigation, S.H.; resources, S.H.; data curation, S.H.; writing—original draft preparation, S.H.; writing—review and editing, S.H., Y.Y.L., and N.L.; visualization, S.H.; supervision, S.H.; project administration, S.H.; funding acquisition, S.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Key Research and Development Program (2016YFD0400101) and the National Natural Science Foundation of China (Nos. 31772370 and 31572175).
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not Applicable.
Data Availability Statement
No datasets were generated and analyzed for this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Morohashi, K.; Minami, M.; Takase, H.; Hotta, Y.; Hiratsuka, K. Isolation and characterization of a novel GRAS gene that regulates meiosis-associated gene expression. J. Biol. Chem. 2003, 278, 20865–20873. [Google Scholar] [CrossRef] [Green Version]
- Pysh, L.D.; Wysocka-Diller, J.W.; Camilleri, C.; Bouchez, D.; Benfey, P.N. The GRAS gene family in Arabidopsis: Sequence characterization and basic expression analysis of the SCARECROW-LIKE genes. Plant J. 1999. [Google Scholar] [CrossRef]
- Bolle, C. The role of GRAS proteins in plant signal transduction and development. Planta 2004, 218, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Fode, B.; Siemsen, T.; Thurow, C.; Weigel, R.; Gatz, C. The Arabidopsis GRAS Protein SCL14 Interacts with Class II TGA Transcription Factors and Is Essential for the Activation of Stress-Inducible Promoters. Plant Cell 2008, 20, 3122–3135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, K.B.; Foley, R.C.; Oñate-Sánchez, L. Transcription factors in plant defense and stress responses. Curr. Opin. Plant Biol. 2002, 5, 430–436. [Google Scholar] [CrossRef]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Molecular responses to dehydration and low temperature: Differences and cross-talk between two stress signaling pathways. Curr. Opin. Plant Biol. 2000, 3, 217–223. [Google Scholar] [CrossRef]
- Hirayama, T.; Shinozaki, K. Research on plant abiotic stress responses in the post-genome era: Past, present and future. Plant J. 2010, 61, 1041–1052. [Google Scholar] [CrossRef]
- Grimplet, J.; Agudelo-Romero, P.; Teixeira, R.T.; Martinez-Zapater, J.M.; Fortes, A.M. Structural and Functional Analysis of the GRAS Gene Family in Grapevine Indicates a Role of GRAS Proteins in the Control of Development and Stress Responses. Front. Plant Sci. 2016, 7, 353. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Carol, P.; Richards, D.E.; King, K.E.; Cowling, R.J.; Murphy, G.P.; Harberd, N.P. The Arabidopsis GAI gene defines a signaling pathway that negatively regulates gibberellin responses. Genes Dev. 1997, 11, 3194–3205. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, A.; Ueguchi-Tanaka, M.; Sonoda, Y.; Kitano, H.; Koshioka, M.; Futsuhara, Y.; Matsuoka, M.; Yamaguchi, J. Slender rice, a constitutive gibberellin response mutant, is caused by a null mutation of the SLR1 gene, an ortholog of the height-regulating gene GAI/RGA/RHT/D8. Plant Cell 2001, 13, 999–1010. [Google Scholar] [CrossRef] [Green Version]
- Bolle, C.; Koncz, C.; Chua, N.H. PAT1, a new member of the GRAS family, is involved in phytochrome A signal transduction. Genes Dev. 2000, 14, 1269–1278. [Google Scholar]
- Helariutta, Y.; Fukaki, H.; Wysocka-Diller, J.; Nakajima, K.; Jung, J.; Sena, G.; Hauser, M.-T.; Benfey, P.N. The SHORT-ROOT Gene Controls Radial Patterning of the Arabidopsis Root through Radial Signaling. Cell 2000, 101, 555–567. [Google Scholar] [CrossRef] [Green Version]
- Stuurman, J.; Jäggi, F.; Kuhlemeier, C. Shoot meristem maintenance is controlled by a GRAS-gene mediated signal from differentiating cells. Genes Dev. 2002, 16, 2213–2218. [Google Scholar] [CrossRef] [Green Version]
- Greb, T.; Clarenz, O.; Schäfer, E.; Müller, D.; Herrero, R.; Schmitz, G.; Theres, K. Molecular analysis of the LATERAL SUPPRESSOR gene in Arabidopsis reveals a conserved control mechanism for axillary meristem formation. Genes Dev. 2003, 17, 1175–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Qian, Q.; Fu, Z.; Wang, Y.; Xiong, G.; Zeng, D.; Wang, X.; Liu, X.; Teng, S.; Hiroshi, F.; et al. Control of tillering in rice. Nature 2003, 422, 618–621. [Google Scholar] [CrossRef] [PubMed]
- Mayrose, M.; Ekengren, S.K.; Melech-Bonfil, S.; Martin, G.B.; Sessa, G. A novel link between tomato GRAS genes, plant disease resistance and mechanical stress response. Mol. Plant Pathol. 2006, 7, 593–604. [Google Scholar] [CrossRef]
- Ma, H.S.; Liang, D.; Shuai, P.; Xia, X.L.; Yin, W.L. The salt-and drought-inducible poplar GRAS protein SCL7 confers salt and drought tolerance in Arabidopsis thaliana. J. Exp. Bot. 2010, 61, 4011–4019. [Google Scholar] [CrossRef] [Green Version]
- Cui, H. Killing two birds with one stone: Transcriptional regulators coordinate development and stress responses in plants. Plant Signal. Behav. 2012, 7, 701–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, K.; Chen, S.; Li, T.; Ma, X.; Liang, X.; Ding, X.; Liu, H.; Luo, L. OsGRAS23, a rice GRAS transcription factor gene, is involved in drought stress response through regulating expression of stress-responsive genes. BMC Plant Biol. 2015, 15, 141. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Fang, L.; Karungo, S.K.; Zhang, L.; Gao, Y.; Li, S.; Xin, H. Overexpression of VaPAT1, a GRAS transcription factor from Vitis amurensis, confers abiotic stress tolerance in Arabidopsis. Plant Cell Rep. 2016, 35, 655–666. [Google Scholar] [CrossRef] [PubMed]
- Cuartero, J.; Bolarín, M.C.; Asíns, M.J.; Moreno, V. Increasing salt tolerance in the tomato. J. Exp. Bot. 2006, 57, 1045–1058. [Google Scholar] [CrossRef]
- Huang, W.; Xian, Z.; Kang, X.; Tang, N.; Li, Z. Genome-wide identification, phylogeny and expression analysis of GRAS gene family in tomato. BMC Plant Biol. 2015, 15, 209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habib, S.; Waseem, M.; Li, N.; Yang, L.; Li, Z. Overexpression of SlGRAS7 Affects Multiple Behaviors Leading to Confer Abiotic Stresses Tolerance and Impacts Gibberellin and Auxin Signaling in Tomato. Int. J. Genom. 2019, 2019, 4051981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; Fujita, Y.; Maruyama, K.; Seki, M.; Hiratsu, K.; Ohme-Takagi, M.; Tran, L.S.P.; Yamaguchi-Shinozaki, K.; Shinozaki, K. A dehydration-induced NAC protein, RD26, is involved in a novel ABA-dependent stress-signaling pathway. Plant J. 2004, 39, 863–876. [Google Scholar] [CrossRef]
- Pan, Y.; Seymour, G.B.; Lu, C.; Hu, Z.; Chen, X.; Chen, G. An ethylene response factor (ERF5) promoting adaptation to drought and salt tolerance in tomato. Plant Cell Rep. 2012, 31, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Mittova, V.; Volokita, M.; Guy, M.; Tal, M. Activities of SOD and the ascorbate-glutathione cycle enzymes in subcellular compartments in leaves and roots of the cultivated tomato and its wild salt-tolerant relative Lycopersicon pennellii. Physiol. Plant 2001, 110, 42–51. [Google Scholar] [CrossRef]
- Morohashi, Y. Peroxidase activity develops in the micropylar endosperm of tomato seeds prior to radicle protrusion. J. Exp. Bot. 2002, 53, 1643–1650. [Google Scholar] [CrossRef] [Green Version]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Fukao, T.; Xu, K.; Ronald, P.C.; Bailey-Serres, J. A variable cluster of ethylene response factor-like genes regulates metabolic and developmental acclimation responses to submergence in rice. Plant Cell 2006, 18, 2021–2034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arvouet-Grand, A.; Vennat, B.; Pourrat, A.; Legret, P. Standardization of propolis extract and identification of principal constituents. J. Pharm. Belg. 1994, 49, 462–468. [Google Scholar]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Zhai, H.; Wang, F.; Si, Z.; Huo, J.; Xing, L.; An, Y.; He, S.; Liu, Q. Amyo-inositol-1-phosphate synthase gene, IbMIPS1, enhances salt and drought tolerance and stem nematode resistance in transgenic sweet potato. Plant Biotechnol. J. 2016, 14, 592–602. [Google Scholar] [CrossRef]
- Sharma, R.; De Vleesschauwer, D.; Sharma, M.K.; Ronald, P.C. Recent Advances in Dissecting Stress-Regulatory Crosstalk in Rice. Mol. Plant 2013, 6, 250–260. [Google Scholar] [CrossRef] [Green Version]
- Gerszberg, A.; Hnatuszko-Konka, K. Tomato tolerance to abiotic stress: A review of most often engineered target sequences. Plant Growth Regul. 2017, 83, 175–198. [Google Scholar] [CrossRef] [Green Version]
- Ashrafi-Dehkordi, E.; Alemzadeh, A.; Tanaka, N.; Razi, H. Meta-analysis of transcriptomic responses to biotic and abiotic stress in tomato. PeerJ 2018, 6, e4631. [Google Scholar] [CrossRef]
- Luo, X.; Chen, Z.; Gao, J.; Gong, Z. Abscisic acid inhibits root growth in Arabidopsis through ethylene biosynthesis. Plant J. 2014, 79, 44–55. [Google Scholar] [CrossRef]
- Lombardo, M.C.; LaMattina, L. Abscisic acid and nitric oxide modulate cytoskeleton organization, root hair growth and ectopic hair formation in Arabidopsis. Nitric Oxide 2018, 80, 89–97. [Google Scholar] [CrossRef]
- Dhanda, S.; Sethi, G. Inheritance of excised-leaf water loss and relative water content in bread wheat (Triticum aestivum). Euphytica 1998, 104, 39–47. [Google Scholar] [CrossRef]
- Orellana, S.; Yañez, M.; Espinoza, A.; Verdugo, I.; González, E.; Ruiz-Lara, S.; Casaretto, J.A. The transcription factor SlAREB1 confers drought, salt stress tolerance and regulates biotic and abiotic stress-related genes in tomato. Plant Cell Environ. 2010, 33, 2191–2208. [Google Scholar] [CrossRef]
- Loukehaich, R.; Wang, T.; Ouyang, B.; Ziaf, K.; Li, H.; Zhang, J.; Lu, Y.; Ye, Z. SpUSP, an annexin-interacting universal stress protein, enhances drought tolerance in tomato. J. Exp. Bot. 2012, 63, 5593–5606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, L.; Xing, X.; Sun, L.; Pan, J.; Kong, X.; Zhang, M.; Li, D. ZmLEA3, a Multifunctional Group 3 LEA Protein from Maize (Zea mays L.), is Involved in Biotic and Abiotic Stresses. Plant Cell Physiol. 2013, 54, 944–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, L.; Schumaker, K.S.; Zhu, J.-K. Cell Signaling during Cold, Drought, and Salt Stress. Plant Cell 2002, 14, S165–S183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 9, 405–410. [Google Scholar] [CrossRef]
- Zhang, H.; Han, B.; Wang, T.; Chen, S.; Li, H.; Zhang, Y.; Dai, S. Mechanisms of Plant Salt Response: Insights from Proteomics. J. Proteome Res. 2011, 11, 49–67. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. REACTIVE OXYGEN SPECIES: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Wang, X.; Jiao, Y.; Qin, Y.; Liu, X.; He, K.; Chen, C.; Ma, L.; Wang, J.; Xiong, L.; et al. Global genome expression analysis of rice in response to drought and high-salinity stresses in shoot, flag leaf, and panicle. Plant Mol. Biol. 2007, 63, 591–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhoeyen, M.E.; Bovy, A.; Collins, G.; Muir, S.; Robinson, S.; De Vos, C.H.R.; Colliver, S. Increasing antioxidant levels in tomatoes through modification of the flavonoid biosynthetic pathway. J. Exp. Bot. 2002, 53, 2099–2106. [Google Scholar] [CrossRef] [Green Version]
- Balasundram, N.; Sundram, K.; Samman, S. Phenolic compounds in plants and agri-industrial by-products: Antioxidant activity, occurrence, and potential uses. Food Chem. 2006, 99, 191–203. [Google Scholar] [CrossRef]
- Gao, J.-J.; Zhang, Z.; Peng, R.-H.; Xiong, A.-S.; Xu, J.; Zhu, B.; Yao, Q.-H. Forced expression of Mdmyb10, a myb transcription factor gene from apple, enhances tolerance to osmotic stress in transgenic Arabidopsis. Mol. Biol. Rep. 2010, 38, 205–211. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phylogenetic tree of SlGRAS10 and other known GRAS proteins. A phylogenetic tree of SlGRAS10 and other GRAS proteins was constructed to show the relationship between Arabidopsis and GRAS protein from tomato.
Figure 1.
Phylogenetic tree of SlGRAS10 and other known GRAS proteins. A phylogenetic tree of SlGRAS10 and other GRAS proteins was constructed to show the relationship between Arabidopsis and GRAS protein from tomato.

Figure 2.
Expression profile analysis of SlGRAS10 in wild-type tomato plants by quantitative real-time PCR. (A) Expression levels of SlGRAS10 gene in different tissues of wild-type tomato. Root; Stem; Leaf; Flower; IMG, immature green fruit; MG, mature fruit; Br, breaker fruit; Br + 4, 4 days after breaker stage; Orange fruit; Red ripe fruit. (B) The expression level of SlGRAS10 under salt stress. (C) The expression level of SlGRAS10 under dehydration stress. (D) The expression level of SlGRAS10 under wounding stress. (E) The response of SlGRAS10 expression to phytohormones (GA3, ABA, and ACC. Asterisks show the significant differences using Student’s t-test (* p < 0.05, ** p < 0.01).
Figure 2.
Expression profile analysis of SlGRAS10 in wild-type tomato plants by quantitative real-time PCR. (A) Expression levels of SlGRAS10 gene in different tissues of wild-type tomato. Root; Stem; Leaf; Flower; IMG, immature green fruit; MG, mature fruit; Br, breaker fruit; Br + 4, 4 days after breaker stage; Orange fruit; Red ripe fruit. (B) The expression level of SlGRAS10 under salt stress. (C) The expression level of SlGRAS10 under dehydration stress. (D) The expression level of SlGRAS10 under wounding stress. (E) The response of SlGRAS10 expression to phytohormones (GA3, ABA, and ACC. Asterisks show the significant differences using Student’s t-test (* p < 0.05, ** p < 0.01).

Figure 3.
Morphological modifications were displayed by SlGRAS10 down-regulated transgenic lines. (A) The down-regulation of SlGRAS10 exhibiting decreased plant height. Photos were taken 40 days after sowing. Scale Bar = 1 mm. (B) Leaves of the wild-type (upper row) and the SlGRAS10-RNAi lines (lower row). Scale Bar = 1 mm. (C) The expression levels of SlGRAS10 in transgenic lines and wild-type. Each value represents the mean ± SE of three independent replicates. Asterisks show the significant differences using Student’s t-test (* p < 0.05, ** p < 0.01). (D) The plant height of SlGRAS10 transgenic three lines and wild-type plants. The plant height was measured every week from 20-day after sowing. Each value represents the mean ± SE of three independent replicates (n = 15).
Figure 3.
Morphological modifications were displayed by SlGRAS10 down-regulated transgenic lines. (A) The down-regulation of SlGRAS10 exhibiting decreased plant height. Photos were taken 40 days after sowing. Scale Bar = 1 mm. (B) Leaves of the wild-type (upper row) and the SlGRAS10-RNAi lines (lower row). Scale Bar = 1 mm. (C) The expression levels of SlGRAS10 in transgenic lines and wild-type. Each value represents the mean ± SE of three independent replicates. Asterisks show the significant differences using Student’s t-test (* p < 0.05, ** p < 0.01). (D) The plant height of SlGRAS10 transgenic three lines and wild-type plants. The plant height was measured every week from 20-day after sowing. Each value represents the mean ± SE of three independent replicates (n = 15).

Figure 4.
Down-regulation of SlGRAS10 decreased leaf size and internode length. (A) Internode length of the first to third nodes from about 35-days-old SlGRAS10-RNAi transgenic and wild-type plants. (B) The total mature compound leaf length of SlGRAS10-RNAi and wild-type plants. (C) The length of the apical leaflet from the mature compound leaves from transgenic and wild-type plants. (D) The width of the apical leaflet from the mature compound leaves from transgenic and wild-type plants. The date represents the mean ± SE of three independent replicates (n = 15). *, ** indicates a significant difference (p < 0.05, p < 0.01) between transgenic and wild-type plants.
Figure 4.
Down-regulation of SlGRAS10 decreased leaf size and internode length. (A) Internode length of the first to third nodes from about 35-days-old SlGRAS10-RNAi transgenic and wild-type plants. (B) The total mature compound leaf length of SlGRAS10-RNAi and wild-type plants. (C) The length of the apical leaflet from the mature compound leaves from transgenic and wild-type plants. (D) The width of the apical leaflet from the mature compound leaves from transgenic and wild-type plants. The date represents the mean ± SE of three independent replicates (n = 15). *, ** indicates a significant difference (p < 0.05, p < 0.01) between transgenic and wild-type plants.

Figure 5.
Down-regulation of SlGRAS10 enhances seedling growth to salt and drought stress. (A) The figure shows the growth performance of wild-type and SlGRAS10-RNAi-10. RNAi-11 and RNAi-15 seedlings under salt and drought stress after 15 days. Scale bar = 1mm (B) Root length of SlGRAS10-RNAi three lines and wild-type under control, salt stress, and drought treatments (n = 20). (C) Shoot lengths of SlGRAS10-RNAi and wild-type under control, salt stress, and drought treatments (n = 20). (D) Seedling weight of SlGRAS10-RNAi and after control, salt stress, and drought treatments (n = 20). The date represents the mean ± SE of three independent replicates. *, ** indicates a significant difference (p < 0.05, p < 0.01) between transgenic and wild-type plants.
Figure 5.
Down-regulation of SlGRAS10 enhances seedling growth to salt and drought stress. (A) The figure shows the growth performance of wild-type and SlGRAS10-RNAi-10. RNAi-11 and RNAi-15 seedlings under salt and drought stress after 15 days. Scale bar = 1mm (B) Root length of SlGRAS10-RNAi three lines and wild-type under control, salt stress, and drought treatments (n = 20). (C) Shoot lengths of SlGRAS10-RNAi and wild-type under control, salt stress, and drought treatments (n = 20). (D) Seedling weight of SlGRAS10-RNAi and after control, salt stress, and drought treatments (n = 20). The date represents the mean ± SE of three independent replicates. *, ** indicates a significant difference (p < 0.05, p < 0.01) between transgenic and wild-type plants.

Figure 6.
Down-regulation of SlGRAS10 enhances seedling growth to ABA stress. (A) The figure shows the growth performance of wild-type and SlGRAS10-RNAi seedlings under control and ABA stress after 15 days. Scale bar = 5 mm (B) Photographs of wild-type and SlGRAS10-RNAi-10, RNAi-11, and RNAi-15 seedlings under control and salt ABA stress after 15 days. Scale bar = 5 mm. (C) The seed Germination rate of SlGRAS10-RNAi and after control, and ABA treatments (n = 20). (D) Root length of SlGRAS10-RNAi three lines and wild-type under control, and ABA treatments (n = 20). (E) Shoot lengths of SlGRAS10-RNAi and wild-type under control, and ABA treatments (n = 20). The date represents the mean ± SE of three independent replicates. *, **, *** indicates a significant difference (p < 0.05, p < 0.01, p < 0.001) between transgenic and wild-type plants.
Figure 6.
Down-regulation of SlGRAS10 enhances seedling growth to ABA stress. (A) The figure shows the growth performance of wild-type and SlGRAS10-RNAi seedlings under control and ABA stress after 15 days. Scale bar = 5 mm (B) Photographs of wild-type and SlGRAS10-RNAi-10, RNAi-11, and RNAi-15 seedlings under control and salt ABA stress after 15 days. Scale bar = 5 mm. (C) The seed Germination rate of SlGRAS10-RNAi and after control, and ABA treatments (n = 20). (D) Root length of SlGRAS10-RNAi three lines and wild-type under control, and ABA treatments (n = 20). (E) Shoot lengths of SlGRAS10-RNAi and wild-type under control, and ABA treatments (n = 20). The date represents the mean ± SE of three independent replicates. *, **, *** indicates a significant difference (p < 0.05, p < 0.01, p < 0.001) between transgenic and wild-type plants.

Figure 7.
Down-regulation of SlGRAS10 enhances tolerance to salt and drought stress treatment. (A) Photographs of representative plants after one month of control, salt stress, drought stress, and re-watered treatments compared to control plants. Scale bar = 5 mm. (B) Total chlorophyll content of SlGRAS10-RNAi (L10, L11) and wild-type plants are shown in (A) under control, salt, and drought stresses. (C) Relative water content (RWC) of SlGRAS10-RNAi (L10, L11) and wild-type plants are shown in (A) under control, salt, and drought stresses. (D) Survival ratio of drought stress plants of WT and SlGRAS10-RNAi lines after re-watered at different time points (1 Day re-watered, 3 Days re-watered, and 6 Days re-watered) (n = 20). The date represents the mean ± SE of three independent replicates. *, **, *** indicates a significant difference (p < 0.05, p < 0.01, p < 0.001) between transgenic and wild-type plants.
Figure 7.
Down-regulation of SlGRAS10 enhances tolerance to salt and drought stress treatment. (A) Photographs of representative plants after one month of control, salt stress, drought stress, and re-watered treatments compared to control plants. Scale bar = 5 mm. (B) Total chlorophyll content of SlGRAS10-RNAi (L10, L11) and wild-type plants are shown in (A) under control, salt, and drought stresses. (C) Relative water content (RWC) of SlGRAS10-RNAi (L10, L11) and wild-type plants are shown in (A) under control, salt, and drought stresses. (D) Survival ratio of drought stress plants of WT and SlGRAS10-RNAi lines after re-watered at different time points (1 Day re-watered, 3 Days re-watered, and 6 Days re-watered) (n = 20). The date represents the mean ± SE of three independent replicates. *, **, *** indicates a significant difference (p < 0.05, p < 0.01, p < 0.001) between transgenic and wild-type plants.

Figure 8.
Down-regulation of SlGRAS10 elevates ROS scavenging ability. CAT, POD, and SOD activities and O2−, H2O2, proline, MDA, and soluble sugar contents in the leaves of SlGRAS10-RNAi (L10, L11) and WT plants under control and stress conditions. The date represents the mean ± SE of three independent replicates. *, ** indicates a significant difference (p < 0.05, p < 0.01) between transgenic and wild-type plants.
Figure 8.
Down-regulation of SlGRAS10 elevates ROS scavenging ability. CAT, POD, and SOD activities and O2−, H2O2, proline, MDA, and soluble sugar contents in the leaves of SlGRAS10-RNAi (L10, L11) and WT plants under control and stress conditions. The date represents the mean ± SE of three independent replicates. *, ** indicates a significant difference (p < 0.05, p < 0.01) between transgenic and wild-type plants.

Figure 9.
Down-regulation of SlGRAS10 elevates flavonoid accumulation. Total flavonoid contents and expression profile of flavonoid biosynthesis genes in SlGRAS10-RNAi transgenic lines (L10, L11) and WT under control, salt, and drought. The date represents the mean ± SE of three independent replicates. *, ** indicates a significant difference (p < 0.05, p < 0.01) between transgenic and wild-type plants.
Figure 9.
Down-regulation of SlGRAS10 elevates flavonoid accumulation. Total flavonoid contents and expression profile of flavonoid biosynthesis genes in SlGRAS10-RNAi transgenic lines (L10, L11) and WT under control, salt, and drought. The date represents the mean ± SE of three independent replicates. *, ** indicates a significant difference (p < 0.05, p < 0.01) between transgenic and wild-type plants.

Figure 10.
Enhanced expression of chlorophyll biosynthesis-related genes in SlGRAS10-RNAi under stress conditions. Expression analysis of SlGLK1 and SlGLK2 genes in SlGRAS10-RNAi (L10, L11) and WT after control, salt, and drought stress treatments. The date represents the mean ± SE of three independent replicates. *, **, *** indicates a significant difference (p < 0.05, p < 0.01, p < 0.001) between transgenic and wild-type plants.
Figure 10.
Enhanced expression of chlorophyll biosynthesis-related genes in SlGRAS10-RNAi under stress conditions. Expression analysis of SlGLK1 and SlGLK2 genes in SlGRAS10-RNAi (L10, L11) and WT after control, salt, and drought stress treatments. The date represents the mean ± SE of three independent replicates. *, **, *** indicates a significant difference (p < 0.05, p < 0.01, p < 0.001) between transgenic and wild-type plants.

Figure 11.
Expression of senescence-associated genes in SlGRAS10-RNAi under stress conditions. Expression analysis of SlSAG12, SlSAG13, and SlSAG15 genes in SlGRAS10-RNAi (L10, L11) and WT after control, salt, and drought stress treatments. The date represents the mean ± SE of three independent replicates. *, ** indicates a significant difference (p < 0.05, p < 0.01) between transgenic and wild-type plants.
Figure 11.
Expression of senescence-associated genes in SlGRAS10-RNAi under stress conditions. Expression analysis of SlSAG12, SlSAG13, and SlSAG15 genes in SlGRAS10-RNAi (L10, L11) and WT after control, salt, and drought stress treatments. The date represents the mean ± SE of three independent replicates. *, ** indicates a significant difference (p < 0.05, p < 0.01) between transgenic and wild-type plants.

Figure 12.
Expression analysis of ROS Scavenging related genes. Expression analysis of stress-related genes (SlAPX, SlCAT2, SlGME2, SlGST, SlLOX, SlP5CS, SlPOD, and SlSOD) in SlGRAS10-RNAi (L10, L11) and WT under control, salt and drought stresses. The date represents the mean ± SE of three independent replicates. *, **, *** indicates a significant difference (p < 0.05, p < 0.01, p < 0.001) between transgenic and wild-type plants.
Figure 12.
Expression analysis of ROS Scavenging related genes. Expression analysis of stress-related genes (SlAPX, SlCAT2, SlGME2, SlGST, SlLOX, SlP5CS, SlPOD, and SlSOD) in SlGRAS10-RNAi (L10, L11) and WT under control, salt and drought stresses. The date represents the mean ± SE of three independent replicates. *, **, *** indicates a significant difference (p < 0.05, p < 0.01, p < 0.001) between transgenic and wild-type plants.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Habib, S.; Lwin, Y.Y.; Li, N. Down-Regulation of SlGRAS10 in Tomato Confers Abiotic Stress Tolerance. Genes 2021, 12, 623. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050623
AMA Style
Habib S, Lwin YY, Li N. Down-Regulation of SlGRAS10 in Tomato Confers Abiotic Stress Tolerance. Genes. 2021; 12(5):623. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050623
Chicago/Turabian StyleHabib, Sidra, Yee Yee Lwin, and Ning Li. 2021. "Down-Regulation of SlGRAS10 in Tomato Confers Abiotic Stress Tolerance" Genes 12, no. 5: 623. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050623
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.