
Alternative Strategies for Multi-Stress Tolerance and Yield Improvement in Millets


1
Laboratory of Biotechnology and Molecular Biology, Department of Biology, University of North Carolina at Greensboro, 321 McIver Street, Greensboro, NC 27412, USA
2
USDA-ARS, U. S. Arid-Land Agricultural Research Center, 21881 N Cardon Ln., Maricopa, AZ 85138, USA
*
Author to whom correspondence should be addressed.
Genes 2021, 12(5), 739; https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050739
Submission received: 8 April 2021
/
Revised: 30 April 2021
/
Accepted: 10 May 2021
/
Published: 14 May 2021
(This article belongs to the Special Issue Genetics and Physiology of Multiple-Stress Tolerance in Crops)
Abstract
:Millets are important cereal crops cultivated in arid and semiarid regions of the world, particularly Africa and southeast Asia. Climate change has triggered multiple abiotic stresses in plants that are the main causes of crop loss worldwide, reducing average yield for most crops by more than 50%. Although millets are tolerant to most abiotic stresses including drought and high temperatures, further improvement is needed to make them more resilient to unprecedented effects of climate change and associated environmental stresses. Incorporation of stress tolerance traits in millets will improve their productivity in marginal environments and will help in overcoming future food shortage due to climate change. Recently, approaches such as application of plant growth-promoting rhizobacteria (PGPRs) have been used to improve growth and development, as well as stress tolerance of crops. Moreover, with the advance of next-generation sequencing technology, genome editing, using the clustered regularly interspaced short palindromic repeats (CRISPR/Cas9) system are increasingly used to develop stress tolerant varieties in different crops. In this paper, the innate ability of millets to tolerate abiotic stresses and alternative approaches to boost stress resistance were thoroughly reviewed. Moreover, several stress-resistant genes were identified in related monocots such as rice (Oryza sativa), wheat (Triticum aestivum), and maize (Zea mays), and other related species for which orthologs in millets could be manipulated by CRISPR/Cas9 and related genome-editing techniques to improve stress resilience and productivity. These cutting-edge alternative strategies are expected to bring this group of orphan crops at the forefront of scientific research for their potential contribution to global food security.
1. Introduction
Millets are considered as major cereal crops in the developing world. In the semiarid tropical areas of Asia and Africa, they are especially important because of their use as human food, as well as feed for livestock [1,2,3]. Millets are cultivated in marginal environments and represents small grain crops [4,5,6]. Finger millet (Eleusine coracana (L.) Gaertn), pearl millet (Pennisetum glaucum (L.) R. Br), kodo millet (Paspalum scrobiculatum L.), Japanese barnyard millet (Echinochloa esculneta), proso millet (Panicum miliaceum L.), foxtail millet (Setaria italica (L.) P. Beauvois), little millet (Panicum sumatrense Roth ex Roem. & Schult.), tef (Eragrostis tef (Zucc.) Trotter), and Indian barnyard millet (Echinochloa frumentacea Link) are traditionally considered as millets [7,8,9]. Pearl millet is an exception to small grain crops due to its morphological features [10,11].
One of the major attributes of millets is their high nutritional value as compared to other cereal crops [12,13,14]. The protein profile of millets shows that they contain higher amounts of methionine and other essential amino acids [15,16]. Few studies also reported that millets are rich in micronutrients and phytochemicals [17,18,19] that have health benefits. For example, antioxidant enzymes, insoluble and soluble dietary fibers, and resistant starch are abundantly found in pearl millet [15,17,18,19]. Biochemical profiling revealed that pearl millet contains 63% starch, 13% protein, 7% fats, 2% crude fibers, and about 92% dry matter [7,15,16]. Foxtail millet is used as a supplementary source of protein for other cereals because of the presence of the essential amino acid lysine [15,17,18,19]. Finger millet is rich in polyphenols and other essential phytochemicals [15,17,18,19,20], and it has a high amount of calcium, methionine, tryptophan, fiber, and sulfur-containing amino acids [21]. Finger millet contains minerals (2%), crude fibers (4%), protein (9%), and carbohydrate (81%) [22,23]. The mineral and fiber contents of finger millet are higher than those of wheat and rice. Finger millet also contains more valine, threonine, and lysine than other millet species [15,17,18,19,22]. The presence of essential nutrients and phytochemicals with health benefits make millets great resources for utilization in food industries [7,15,17,18,19,22].
Despite the great benefits of millets for humans, their yield is limited by multi-environmental stresses and the looming climate change [24,25]. Combined heat and drought stress has posed a serious threat to the productivity of these crops [26,27]. Drought stress at the seedling stage [28] and terminal drought during the reproductive stage have been shown to incur up to 60% and 40% of yield loss in pearl millet and tef, respectively [28,29,30]. In Africa, particularly in the sub-Saharan countries, about 40% of the population lives in a drier environment; however, this number is increasing and is expected to double by 2050 [31]. Drought accompanied by heat stress has a considerable effect on the physiological, cellular, and molecular functions of plants. Photosynthesis and respiration are among the major processes affected by these stresses, which determine crop yield and productivity [26,27,32]. Studies suggested that a temperature above the threshold level (28 °C to 32 °C) has very harmful effects, for example, by limiting the adaptation potential of a crop [26,27,32,33]. An increase in temperature of about 3–4 °C can reduce crop yields by up to 35% [32,34]. Reports on global warming have suggested that south Asia and Africa are regions most affected by climate change, primarily global warming [35,36]. It was reported that the average annual increase in temperature since 1980 has been 0.17 °C (0.31 °F) [32,34,37]. A direct link between decreased crops yields and combined drought and heat stress has also been reported [26,27,32,38].
In order to overcome the effects of climate change-associated stresses and to improve the yield of millets, there is a need to develop stress-tolerant and high-yielding varieties. There are several possible approaches to increase the stress tolerance and productivity of millets. For example, application of PGPRs has been used to improve yield and stress tolerance in wheat [39,40,41] and rice [42,43,44], but it remains to be tested in millets. Another approach for trait improvement is via genome editing, which has recently gained tremendous attentions due to its specific allele manipulation potential. The CRISPR/Cas9 system has emerged as a promising technique to edit plant genome/genes for stress tolerance and higher productivity. CRISPRs, along with their related proteins known as Cas, are widespread among the phyla of archaea and bacteria where they function as an adaptive immune system against phages [45]. The CRISPR loci acquire short sequences of their DNA as spacers and, thus, carry information regarding previous plasmid infections or bacteriophages upon recognition of invading DNA [46].
Conventional breeding approaches such as selection from landraces, hybridization to create new variability followed by pedigree selection, mutation breeding, and exploitation of hybrid vigor have resulted in significant improvement and release of new cultivars in different millets [47,48,49]. However, the yields of millets remain much lower than major cereals even under optimum growth conditions. Following the advent of next-generation sequencing, markers and genomics-assisted breeding approaches have been implemented to improve these group of crops. The genome sequences of foxtail millet [50], tef [51,52], pearl millet [53], finger millet [21], and proso millet (broomcorn) [54] have become available and are very useful resources for genetic improvement of these crops.
Although millets have recently gained attention for their various food and health benefits, as well adaptation to adverse environmental conditions, their productivity has remained low, and which needs to be boosted to enhance their utilization as a food crop globally. This review article highlights the combined effects of drought and heat on millets concurrent with looming climate change, as well as their inherent genetic potential as climate smart crops, and proposes potential alternative strategies for the improvement of stress tolerance and yield of these important group of crops.
2. Effects of Climate Change on Millets and Their Tolerance Strategies
Climate change and its impact on agriculture crops has been widely reviewed [55,56,57,58,59,60]. The climate is changing at an alarming rate, triggering various abiotic stresses which are affecting food crops in one way or another, which directly affects the global food supply. The major effects of climate change on millet crops include drought stress, heat stress, flooding or waterlogging stress, and lodging.
2.1. Effects of Drought Stress
The arid and semiarid lands of most developing countries are facing a problem of water scarcity, which affects the type and performance of crops grown in those areas. A study performed on wild millet (Setaria glauca), foxtail millet (Setaria italica), little millet (Panicum sumatrense), and proso millet (Panicum miliaceum) reported a significant reduction in yield when subjected to drought prior to flowering [61]. Likewise, complete yield loss was reported in two finger millet landraces subjected to drought after 4 weeks of sowing [61,62]. About 60% yield loss was recorded due to terminal drought that occurred during flowering through maturity [63]. The estimated yield loss due to drought in pearl millet was about 51% [64,65], and, in tef, it was estimated to be 40% [66,67], as shown in Table 1. Millets including tef can generally survive at very low soil moisture content. For example, it was reported that, in the Sahel region where the moisture level is extremely low, the biomass of millet and sorghum was comparable [68]. Undesirable effects of drought have been reported not only on the productivity of crops but also on their nutritional quality including grain mineral and protein content [12,61,69,70].
Plants employ various abiotic stress tolerance mechanisms [71,72] to thrive in drought-prone regions. Four major adaptation mechanisms of millets to drought-inflicted regions of the world were recently reviewed [73]. These mechanisms include (i) drought avoidance, which is the capability of plant to sustain the balance of water during stress to avoid water deficiency in tissues, (ii) drought tolerance, referring to plants’ ability to produce biomass by withstanding reduced water potential, (iii) drought escape, a state in which plants mature prior to drought stress, and (iv) drought recovery, a condition in which plants provide some yield by recovering from intermittent drought effects after moisture becomes available.
In sub-Saharan African, millets are adapted to low water potential, and grown for sustenance. These crops are presumed to guarantee food security in the future as they can be used as model for stress tolerance in crop improvement or as an alternative crop in drought-inflicted areas [74]. Drought tolerance strategies in millets also involve physiological modifications including stomatal conductance, osmotic adjustment, and cell membrane stability [75]. Among these, osmotic adjustment enables the plant leaves to maintain leaf turgor pressure (LTP) [76], even under extreme drought conditions, by retrieving and absorbing water even from dry soils [76,77]. Increased root elongation is another drought stress tolerance mechanism [78,79]. For example, the increase in root length of legumes such as cowpea (Vigna unguiculata), peanut (Arachis hypogaea), and soybean (Glycine max), when exposed to drought enables them to absorb water at greater soil depth [79]. Similarly, Ayele, et al. [80] and Debieu, et al. [81] reported that a deeper, more extensive, and broader root system in tef was shown to provide drought stress tolerance.
2.2. Effects of Heat Stress
Although most millet species are resistant to heat stress, heat induces many physiological and molecular alterations. Photosynthesis and respiration are the most sensitive processes to heat stress, which has a dramatic effect on crop productivity [82]. Significant yield loss due to heat has already been reported in many crops [83], and the annual increase in temperature due to climate change poses a threat to food security. Crop yield increases with increasing temperatures up to a threshold level (cotton 32 °C, soybean 30 °C, and maize 29 °C); however, above the threshold level, a slight increase in temperature has severe negative effects on plant growth and ultimately yield [84]. Yield loss of up to 35% has been reported due to a 3–4 °C increase in temperature (Table 1). As estimated by statistical modeling, climate change and global warming are likely to severely affect cereal production in Africa and Asia [85].
It has been established that high-temperature stress reduces transport of electrons, disrupts the function of photosystem (PS) II, and enhances the amount of ROS accumulation [86]. It also desiccates the reproductive parts and can result in plant sterility, seed abortion, reduced seed number, and shortened grain filling period [87]. Over time, through evolution, plants have developed various acclimation, avoidance, and adaptive strategies to deal with heat stress. Mechanisms of tolerance involve upregulation of the antioxidant system, transcription factors, heat-shock proteins, signaling molecules, ion transporters, and accumulation of osmoprotectants [88]. Plant membranes are prone to lipid peroxidation, and, in wheat, membrane thermal stability has been used as a selection criterion for heat tolerance [89]. Under normal conditions, cell metabolism produces ROS as a byproduct, which at high concentration can induce oxidative stress. Most plants use pathways containing antioxidants to combat ROS as protective gear against various abiotic stresses [90]. Acquired heat tolerance has been observed in cowpea [91], chickpea (Cicer arietinum) [92], and pearl millet [93]. In chickpea, identification of 18 single-nucleotide polymorphisms (SNPs) from five stress-responsive genes has been reported, which include dehydration responsive element binding (DREB), abscisic acid-, stress-, and ripening-induced (ASR), NAD+-dependent aminoaldehyde dehydrogenase (AMDH), ERECTA (ER), and cyclase-associated proteins (CAP2) promoter. These genes were highly associated with different adaptive traits under drought and heat stress [92].
2.3. The Effect of Waterlogging Stress on Millets
Waterlogging stress is the main cause of low productivity in high-precipitation areas [94]. Under waterlogging stress, the soil pores are filled with water that leads to the accumulation of toxic compounds, and inhibition of gas diffusion. This eventually affects roots, stomatal conductance, and photosynthesis [95]. Crops like wheat and maize are mostly affected by waterlogging in black clay soil (Vertisols), which has high water-holding capacity. Grain yield losses of about 18% in wild millet and 16% in proso millet were reported with waterlogging treatment that lasted from two weeks after planting through crop maturity [95], as shown in Table 1.
Plants have various mechanisms to cope with waterlogging stress [96], which are induced by hypoxia (reduced oxygen level) or anoxia (complete absence of oxygen). Plants respond to diminished oxygen by carrying out anaerobic respiration, which has also been reported in finger millet (Eleusine coracana) [97]. Anaerobic metabolism is not as efficient as aerobic metabolism, but ATP produced through fermentation supports the cell for a short period. This mechanism requires more sugar than aerobic metabolism; thus, alterations in carbohydrate metabolism are observed in waterlogging tolerant species such as finger millet and rice [98]. During waterlogging stress, spongy tissue containing air gaps, i.e., aerenchyma, which allow gases to move to roots from stems, are formed in tolerant plants [99]. These spaces are developed without (schizogenous) or with (lysigenous) cell death [100]. Studies showed that, in stressed sunflower (Helianthus annuus), aerenchyma (lysigenous) are developed within 2 days of the onset of stress [101]. Another strategy employed by waterlogging-tolerant species includes the formation of adventitious roots [96]. Development of adventitious roots has been observed in finger millet [102] and sorghum (Sorghum bicolor) [103]. Other crops such as mung beans (Vigna radiata) are known to escape logging through fast growth [104]. Additionally, waterlogging-tolerant plant possess abundant solubilized sugar [105], while plants such as tef respond to waterlogging stress by enhancing the activity of nitrogen reductase in the shoots [106].
2.4. Lodging Effects on Millet Yield
Lodging is the permanent bending of the stem from an upright position that is a common problem in millet crops. Forces such as irrigation water, rain, wind, or their combination can induce lodging. For example, lubrication of soil by rain water in combination with wind can push the plants toward the soil [107]. The stems and roots are two main targets of lodging stress, which are described in the literature as stem lodging and root lodging, respectively [108,109]. In stem lodging, the stems tilt toward the soil or break, whereas root lodging is associated with a change in the angle between the stem and soil due to the wind force on the stem or crown bending/root disanchoring [110]. Pinthus [107] reported that, before the stem breaking, more force is exerted to the plant part interacting with the soil. Contrarily, stem flexibility enhances the swing time of the stem, which increases the damage due to minor wind force, thus leading to lodging stress in most crops.
Several studies have reported the negative impacts of lodging stress on yields of tef and foxtail millet crops (Table 1). In tef, the leaf and stem architecture are very delicate, and the plant is susceptive to root lodging [111]. Lodging stress can also reduce yield in foxtail millet Tian, et al. [112]). Another study performed by Opole [113] showed excess fertilizer to cause lodging stress in finger millet, which ultimately reduces yield. Pearl millet is resistant to lodging, as well as various other biotic and abiotic stresses [114].
Generally, most millet crops are resistant to abiotic stresses including lodging under low-input conditions as compared to other cereal crops. However, new strategies need to be developed, or existing techniques need to be improved to enhance lodging tolerance in millet crops to benefit from fertilizer application to increase yields. Reducing plant height by genetic manipulation or exogenous application of chemicals is a common strategy to overcome lodging stress in cereals. For example, during the green revolution, the genes (Rht dwarfing genes) responsible for reducing plant height were introduced into wheat to enhance input responsiveness, e.g., to nitrogen fertilizer [115]. Other crop management practices that can reduce lodging are changing the seed sowing date, tilling practices, and increasing the intra-row space or reducing the number of plants in a row [107,109,110]. Moreover, application of silicon amendments has been shown to increase the yield of millets and other crops such as rice and sugar cane [116,117,118,119,120]. In rice, Si treatment was reported to strengthen the stem by increasing silica deposition in the shoot, increasing the thickness or strength of the culm wall and vascular bundle, and enhancing stem stability [121].
Another potential strategy to reduce plant height and improve lodging tolerance is the inhibition of plant growth regulators (PGRs) such as gibberellic acid (GA). GA inhibitors play a crucial role in the development of dwarf, semi-dwarf, and sturdier plants that can withstand lodging stress [110,122]. Semi-dwarfism and a reduction in plant height are caused by shorter internodes [123]. During the green revolution era, altered GA deficiency was considered the key in producing semi-dwarf rice and wheat cultivars with high yielding potential that boosted productivity of the two cereals [124]. Some major inhibitors of GA that have been utilized for the regulation of plant architecture include daminozide, mepiquat-Cl, chlormequat-Cl, and paclobutrazol (PBZ). PBZ (α-tert-Butyl-β-(4-chlorobenzyl)-1H-1,2,4-triazole-1-ethanol) inhibits GA precursor ent-kaurene conversion to ent-kaurenoic acid [125]. Some effects of PBZ include plant height reduction, enhanced nutrient uptake, and increased seed yield [126].
In millet crops, different approaches have been employed to mitigate or reduce lodging stress and improve yield. For example, in tef and finger millet, application of PBZ has been shown to reduce plant height and lodging stress [127]. Jency, et al. [128] reported that mutation in the kodo millet CO3 variety by gamma radiation or ethyl methane sulfonate (EMS) could produce nonlodging mutants. They also developed mutants (named second mutants or M2) which showed higher lodging tolerance due to photosynthetic efficiency (PhE) and culm thickness. Mutation of the alpha-tubulin-1 gene in tef by EMS produced the lodging-tolerant cultivar ‘Kegne’ [129].

{kind=link}
{kind=link}
Table 1.
Effect of different abiotic stresses such as drought, heat, waterlogging, and lodging stresses on the yield of millet crops. Data represent percent yield loss in kg·ha−1 in response to a particular abiotic stress.
Table 1.
Effect of different abiotic stresses such as drought, heat, waterlogging, and lodging stresses on the yield of millet crops. Data represent percent yield loss in kg·ha−1 in response to a particular abiotic stress.
| Stress | Finger Millet | Foxtail Millet | Wild Millet | Kodo Millet | Pearl Millet | tef | Little Millet | Proso Millet | References |
|---|---|---|---|---|---|---|---|---|---|
| Drought | 61% | 20.3% | 30.1% | 60.1% | 69–77% | 80.5% | 64% | [28,62,130] | |
| Waterlogging | 42.14 kg/ha | 42.84 kg/ha | 18.14 kg/ha | [131] | |||||
| Lodging | 41.2% to 51.1% | 30–35% | [109,132,133] | ||||||
| Heat | 75% at 36/26 °C and 84% at 38/28 °C | 60% at 38/28 °C | 70 to 75% at 36/26 °C | [134,135,136] |
3. Alternative Strategies for Enhancing Stress Resilience in Crops
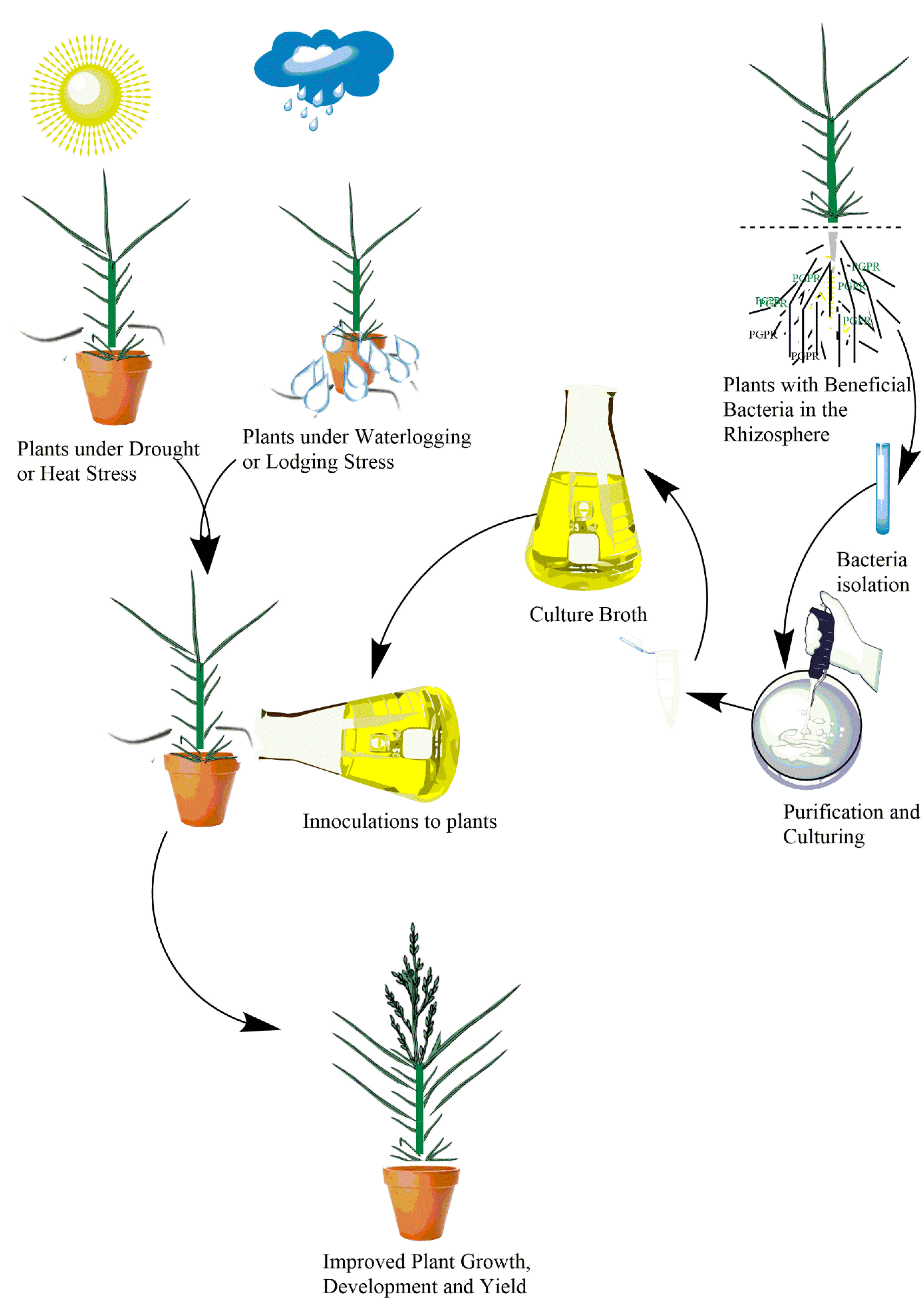
3.1. Application of Plant Growth-Promoting Rhizobacteria (PGPRs)
The beneficial effect of PGPRs in improving abiotic stress tolerance and yield has been revealed in various crops [137,138]. PGPRs have been used to mitigate abiotic stresses and improve productivity in economically important crops including rice [139], soybean [140], lettuce [141], tomato [142], maize [143], and wheat [144,145,146]. One mechanism via which PGPRs improve plant performance is via the biosynthesis of essential plant growth regulators such as gibberellic acid (GA) and indole acetic acid (IAA). PGPRs were also found to trigger plant defense mechanisms and the biosynthesis of other growth regulators such as jasmonic acid and salicylic acid [147,148]. It was also found to help plants with nutrient acquisition from the soil in stress conditions [149].
3.1.1. IAA Synthesis
Inoculation of IAA-synthesizing bacteria in various plant species resulted in enhanced root growth along with the formation of root hairs and lateral roots [150], resulting in improved nutrient and water uptake [151] and providing support to plants to tolerate osmotic stress [152]. Drought tolerance of plants increased manyfold due to IAA-synthesizing Azospirillum [150]. Production of hormones by bacteria and their activity in stimulating endogenous hormones contribute significantly to improving resistance [153]. Nitric oxide (NO) produced by Azospirillum brasilense is involved in IAA signaling, which assists tomato (Solanum lycopersicum) plants in the formation of adventitious root [154]. Association of A. brasilense (strain Cd) to bean (Phaseolus vulgaris L.) during drought condition led to an enhancement in specific root length and root projection area as compared to control without A. brasilense inoculation [155].
3.1.2. PGPR Effects on Root Morphology under Drought
Cell membranes play a great role in maintaining the physiological status of plant cells. Rhizobacteria influence processes that take place in the cell membrane. For example, in wheat seedlings, Azospirillum brasilense reduces the cell membrane potential, while, in cowpea, it was shown to decrease the phospholipid level of cell membranes and cause fluctuations in proton efflux [156]. Water deficit was reported to reduce phosphatidylethanolamine, alter root phospholipid composition, and improve phosphatidylcholine [157]; however, introduction of Azospirillum to wheat seedlings prevented these alterations, although lower phosphatidylethanolamine unsaturation and higher phosphatidylcholine were detected [158]. Alterations conferring elasticity in the cell membrane of roots due to bacterium-mediated changes is the primary mechanism for increased resistance to osmotic deficit [150]. The stability of cell membranes in plants is enhanced by the presence of PGPRs, which activate the antioxidant defense system, leading to increased resistance in plants against drought [159].
3.1.3. Activity of ACC Deaminase-Synthesizing Rhizobacteria
Under stress conditions, endogenous ethylene maintains homoeostasis, leading to diminished shoot and root growth. Aminocyclopropane-1-carboxylic acid (ACC) is a precursor for ethylene biosynthesis [160], which is acted upon by bacterial ACC deaminase to impart energy and nitrogen to the plant [161]. In addition, the removal of ACC enables the bacteria to reduce ethylene toxicity, promoting growth and ameliorating stress [162]. Achromobacter piechaudii strain ARV8 which produces ACC deaminase was shown to improve the weights of pepper (Capsicum annuum) and tomato seedlings, as well as drop ethylene synthesis during saline stress [163]. Colonization of PGPRs from water-deficient areas due to tandem dry episodes are more stress-adapting and plant growth-promoting compared to bacteria colonized from sites where water is abundantly available [163]. Treatment of tomato seedlings with A. piechaudii ARV8 obtained from an arid site promoted growth as compared to seedlings treated with GR12-2 of P. putida, which was obtained from grass rhizosphere where water is abundant [164].
3.1.4. Volatile Compounds and Drought Tolerance
Soil microbes when interacting with plant roots produce chemical compounds which are either organic volatile and inorganic volatile compounds in the form of gases which diffuse through the gaps in the soil particles or nonvolatile compounds (siderophores and phytohormones) [165]. These compounds play vital roles in the food chain of microbes, as well as in promoting plant growth, by improving plant biomass and defense systems against the plant pathogens through induced systemic resistance [166,167,168,169]. In plants, exposure to multiple stresses at the same time requires the functioning of volatiles [170,171]. These molecules introduced during stress conditions take part in signaling for generating systemic and priming effects within the same and nearby plants [172,173]. Enrichment with the AZP2 strain of Bacillus thuringiensis in wheat seedlings led to an increased biomass of plant and a five-fold increased survival rate during drought conditions because of a major decline in volatile emissions and photosynthesis enhancement [145]. These findings prove that introduction of bacteria contributes to stress resistance in plants [145]. These molecules stand as major participants for assessing drought and its mitigation through rapid, noninvasive techniques [145]. Growth of P. chlororaphis O6 in roots reduced loss of water by regulating stomata pores through a volatile metabolite known as 2R,3R-butanediol, whereas bacteria lacking such metabolite production did not show any sign of drought tolerance. This volatile also assists in introducing resistance in Arabidopsis during times of stress [174].
Given their application in improving plant performance under stress conditions in different crop plants as mentioned above, PGPRs have great potential to boost productivity of millets under abiotic stress conditions. A graphical illustration of the potential applications of PGPRs in millet crops is presented in Figure 1.
3.2. Application of CRISPR/Cas9 to Improve Stress Resilience in Millets
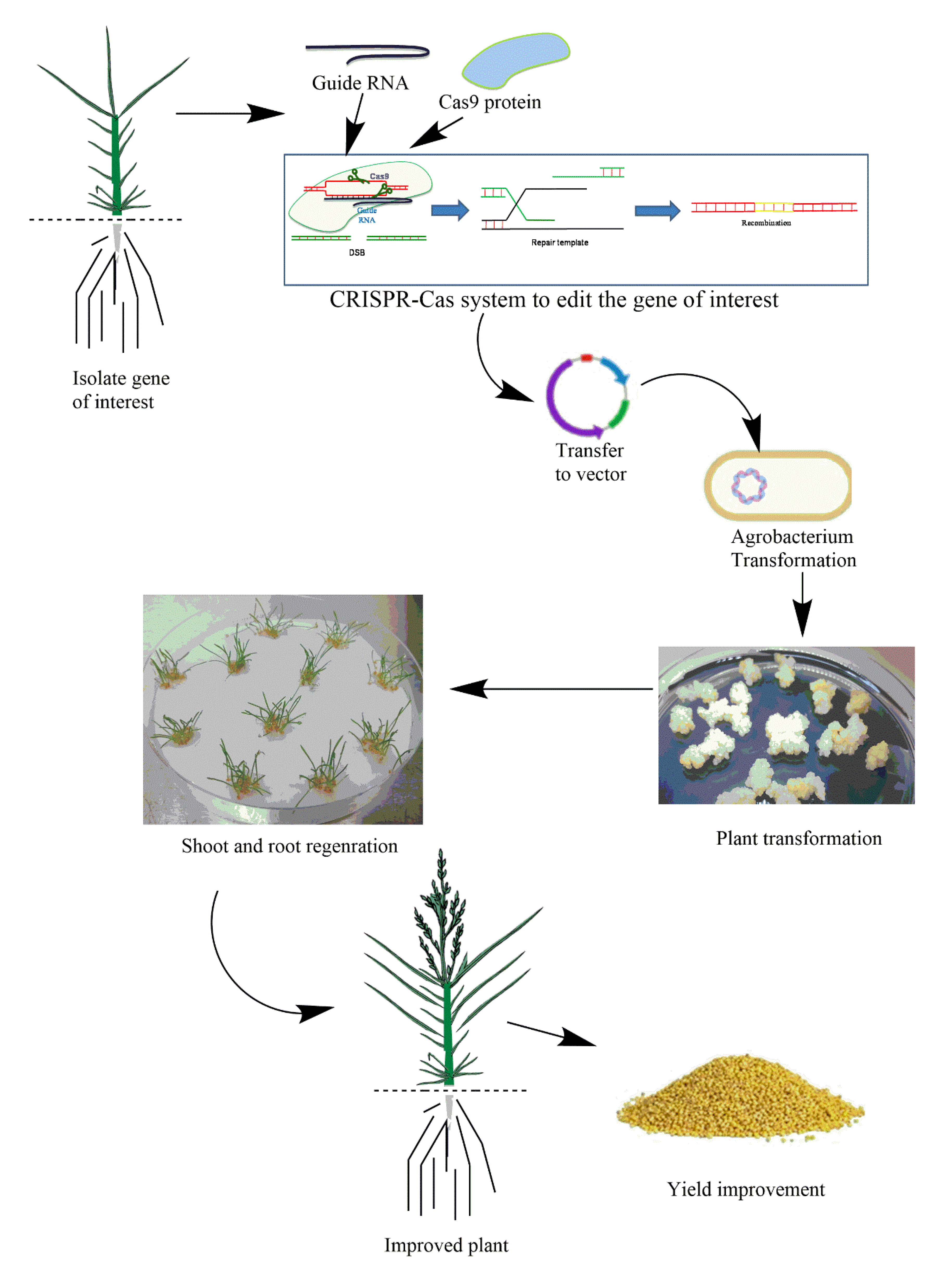
Site-specific nuclease (SSN)-based genome editing, developed in the last decade, has enabled effective and precise gene modification in plant and animal systems. The SSNs create double-strand breaks (DSBs) in their target DNA. These breaks are repaired via pathways such as homology-directed recombination (HDR) or nonhomologous end joining (NHEJ) that lead to mutations such as substitutions and insertion/deletions in the target regions [175]. Genome-editing techniques are used to produce mutants with defined phenotypes in contrast to the transgenic approach, in which the foreign DNA is randomly inserted into the genome and may or may not produce the desired phenotype [176]. Thus, the genome-editing technique is becoming a potent tool in crop breeding and functional genomics. Plants carrying edited genomes have the advantage of carrying modified DNA for a particular trait [177], while new varieties developed using this method can be used directly, unlike transgenic plants, with fewer concerns for consumers. Genome-edited plants carrying new alleles can also be used in breeding programs because of lower regulatory protocols as compared to genetically modified ones [178].
The CRISPR/Cas9 genome-editing technique has been used in more than 20 plant species [179] for improving various traits such as biotic and abiotic stress resilience and yield improvement [180,181]. Selection of a target gene is crucial to achieve the improvement of a desired trait. There are mainly two categories of genes that can be targeted for trait improvement: regulatory and structural genes. Proteins encoded by structural genes directly affect a trait, for example, abiotic stress tolerance [182], while regulatory genes act indirectly by controlling expressions of other genes that may also be involved in other cellular processes [183]. Furthermore, cis-regulatory sequences also play a crucial role in controlling abiotic stress tolerance [182]. In plants, the CRISPR/Cas9 system has been successfully employed in species such as cotton [184], maize [185,186], rice [187,188], and wheat [188,189] for abiotic stress mitigation. However, application of the CRISPR technique is limited to a few plant species, and it has focused on improvement of traits such as biotic stresses (diseases and insect pests), whereas its application for improving abiotic stress resilience and crop yield is limited. Recently, CRISPR/Cas9 was used to improve heat tolerance by targeting the SlAGAMOUS-LIKE 6 (SIAGL6) gene, which showed enhanced fruiting ability under heat stress in tomato [190]. CRISPR/Cas9 was also used for drought tolerance in maize by regulating the ARGOS gene without affecting the yield of the crop [181,191]. CRISPR/Cas9 technology has tremendous potential in developing multi-stress-resilient crops via simultaneous expression of many structural and regulatory genes in crop plants. Multiple gene editing via CRISPR/Cas9 has been performed for some crops including cotton [192], maize [185], wheat [193], and rice [187]. Single-base editing system of CRISPR-Cas has further widened its applications for the improvement of many important traits in crop plants [194,195]. Single-nucleotide changes in an essential domain of a gene may lead to loss-of-function mutation. It is thought that this technique may replace the traditional plant breeding approaches which were mostly based on the presence of populations with enough genetic variations for introducing desired traits to particular crop cultivars [196,197]. The base-editing technique of CRISPR/Cas9 can produce allelic variants in a particular population, thus leading to a desirable trait which can be identified by gRNA sequencing [198]. Thus, underutilized orphan crops such as millet could benefit from the immense potential of CRISPR/Cas9 genome-editing technology for environmental stress resilience and yield improvement. Table 2 summarizes the list of candidate genes which can be edited in millet crops through the CRISPR/Cas9 technique.
As incorporation of millet crops in food security programs is attracting increasing interest, the improvement of millets via the application of CRISPR/Cas9 and other gene-transformation technologies is being considered by the scientific community. For example, Mamidi, et al. [236] reported the genome assembly of Setaria viridis for the identification of important loci for traits such as loss of shattering and leaf angle, which are considered important yield predictors in many grass crops. They further validated the Less Shattering1 (SvLes1) gene through CRISPR/Cas9 to control seed shattering. In other studies, Agrobacterium-mediated transformation was previously performed in foxtail millet (Setaria italica) for downregulation of phosphate transporters (SiPHT1;2, SiPHT1;3, and SiPHT1;4) [237]. They reported significant reductions in inorganic and total phosphate in root and shoot tissues, as well as an increase in the number of roots and hairy roots for the nonredundant roles. Furthermore, foxtail millet was the first millet crop to be sequenced [50]. The genome sequence of pearl millet has also been published [53]. Another study reported the successful production of transgenic finger millet (Eleusine coracana (L.) Gaertn.) plants through Agrobacterium [238]. The Agrobacterium-mediated transformation protocol has been developed for finger millet, and four cultivars of finger millet have been successfully regenerated through Agrobacterium transformation techniques [239]. Johnson, et al. [240] performed genome-wide population studies of three important millet crops: proso millet, little millet, and kodo millet. They identified various SNIPs: 3461 in kodo millet, 2245 in little millet, and 1882 for proso millet. These findings could help in genome editing for stress resilience and crop improvement.
Recently, a foxtail millet mutant was generated by EMS-induced mutagenesis [241]. A point mutation named ‘Xiaomi’ in the light receptor gene phytochrome C (PHYC), which is essential in photoperiodic flowering and has rapid cycling time, was developed [242]. A CRISPR/Cas9 system that can be used to edit millet crops to improve stress resilience and yield is illustrated in Figure 2. We recently summarized tef homologs of major abiotic stress-responsive genes identified in related monocots such as rice, wheat, and maize, and we suggest those genes for CRISPR/Cas9 editing in tef for stress resilience and crop improvement [243].
4. Conclusions and Future Prospects
Millet crops have vital nutritional benefits as compared to other cereals. Nearly all millet crops have innate mechanisms to cope with certain environmental stresses such as heat, drought, lodging, and waterlogging, yet these stresses remain a threat to millet production with increasing impact of climate change. Techniques such as PGPRs and CRISPR/Cas9 are being used in other crops to lessen the impact of abiotic stresses, as well as to improve the productivity of crops. In this paper, we reviewed available literature on the subject matter and projected that the use of PGPRs and CRISPR/Cas9 will not only enable the plants to grow well in adverse conditions but also improve their yield significantly. Some candidate genes that can be targeted for manipulation by the CRISPR/Cas9 system to improve the growth and yield of millet crops have been suggested. Genomics, transcriptomics, metabolomics, proteomics, and other fields of study will also complement the alternative strategy we put forward. Improvement of millets has lagged behind that of major food crops and deserves increased attention from geneticists, biotechnologists, breeders, germplasm conservationists, etc. to improve global food security amidst climate change that is increasingly affecting the productivity of staple crops.
Author Contributions
M.N. and A.L.-O. conceived the idea. M.N. drafted the manuscript. D.D.S. and A.L.-O. thoroughly edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This manuscript was support by the University of North Carolina at Greensboro (Grant # 133504 to A.L.-O.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not Relevant.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Taylor, J.R. Sorghum and Millets: Taxonomy, History, Distribution, and Production. In Sorghum and Millets; Elsevier: Amsterdam, The Netherlands, 2019; pp. 1–21. [Google Scholar]
- Alavi, S.; Mazumdar, S.D.; Taylor, J.R. Modern convenient sorghum and millet food, beverage and animal feed products, and their technologies. In Sorghum and Millets; Elsevier: Amsterdam, The Netherlands, 2019; pp. 293–329. [Google Scholar]
- Dias-Martins, A.M.; Pessanha, K.L.F.; Pacheco, S.; Rodrigues, J.A.S.; Carvalho, C.W.P. Potential use of pearl millet (Pennisetum glaucum (L.) R. Br.) in Brazil: Food security, processing, health benefits and nutritional products. Food Res. Int. 2018, 109, 175–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.M.; Arora, S.; Mirza, N.; Pande, A.; Lata, C.; Puranik, S.; Kumar, J.; Kumar, A. Finger millet: A “certain” crop for an “uncertain” future and a solution to food insecurity and hidden hunger under stressful environments. Front. Plant Sci. 2017, 8, 643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muthamilarasan, M.; Prasad, M. Small millets for enduring food security amidst pandemics. Trends Plant Sci. 2020, 26, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, L.; Smale, M. Village seed systems and the biological diversity of millet crops in marginal environments of India. Euphytica 2007, 155, 167–182. [Google Scholar] [CrossRef]
- Jukanti, A.; Gowda, C.L.; Rai, K.; Manga, V.; Bhatt, R. Crops that feed the world 11. Pearl Millet (Pennisetum glaucum L.): An important source of food security, nutrition and health in the arid and semi-arid tropics. Food Secur. 2016, 8, 307–329. [Google Scholar] [CrossRef]
- Madella, M.; Lancelotti, C.; García-Granero, J.J. Millet microremains—An alternative approach to understand cultivation and use of critical crops in Prehistory. Archaeol. Anthropol. Sci. 2016, 8, 17–28. [Google Scholar] [CrossRef]
- Tadele, Z. Orphan crops: Their importance and the urgency of improvement. Planta 2019, 250, 677–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javorski, M.; Castan, D.O.C.; Silva, S.S.d.; Guilhien Gomes-Junior, F.; Cicero, S.M. Image analysis to evaluate the physiological potential and morphology of pearl millet seeds. J. Seed Sci. 2018, 40, 127–134. [Google Scholar] [CrossRef]
- Govindaraj, M.; Rai, K.N.; Kanatti, A.; Upadhyaya, H.D.; Shivade, H.; Rao, A.S. Exploring the genetic variability and diversity of pearl millet core collection germplasm for grain nutritional traits improvement. Sci. Rep. 2020, 10, 21177. [Google Scholar] [CrossRef] [PubMed]
- Sarita, E.S.; Singh, E. Potential of millets: Nutrients composition and health benefits. J. Sci. Innov. Res. 2016, 5, 46–50. [Google Scholar]
- Chanyalew, S.; Ferede, S.; Damte, T.; Fikre, T.; Genet, Y.; Kebede, W.; Tolossa, K.; Tadele, Z.; Assefa, K. Significance and prospects of an orphan crop tef. Planta 2019, 250, 753–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Tomer, V.; Kaur, A.; Kumar, V.; Gupta, K. Millets: A solution to agrarian and nutritional challenges. Agric. Food Secur. 2018, 7, 31. [Google Scholar] [CrossRef]
- Nithiyanantham, S.; Kalaiselvi, P.; Mahomoodally, M.F.; Zengin, G.; Abirami, A.; Srinivasan, G. Nutritional and functional roles of millets—A review. J. Food Biochem. 2019, 43, e12859. [Google Scholar] [CrossRef]
- Dayakar Rao, B.; Bhaskarachary, K.; Arlene Christina, G.; Sudha Devi, G.; Vilas, A.T.; Tonapi, A. Nutritional and Health Benefits of Millets; ICAR_Indian Institute of Millets Research (IIMR): Hyderabad, Indian, 2017; p. 112. [Google Scholar]
- Bandyopadhyay, T.; Jaiswal, V.; Prasad, M. Nutrition potential of foxtail millet in comparison to other millets and major cereals. In The Foxtail Millet Genome; Springer: Berlin/Heidelberg, Germany, 2017; pp. 123–135. [Google Scholar]
- Nakarani, U.M.; Singh, D.; Suthar, K.P.; Karmakar, N.; Faldu, P.; Patil, H.E. Nutritional and phytochemical profiling of nutracereal finger millet (Eleusine coracana L.) genotypes. Food Chem. 2021, 341, 128271. [Google Scholar] [CrossRef] [PubMed]
- Disseka, W.K.; Faulet, M.B.; Koné, F.M.T.; Gnanwa, M.J.; Kouamé, L.P. Phytochemical composition and functional properties of millet (Pennisetum glaucum) flours fortified with sesame (Sesamum indicum) and moringa (Moringa oleifera) as a weaning food. Adv. Res. 2018, 15, 1–11. [Google Scholar] [CrossRef]
- Bora, P.; Ragaee, S.; Marcone, M. Effect of parboiling on decortication yield of millet grains and phenolic acids and in vitro digestibility of selected millet products. Food Chem. 2019, 274, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Hittalmani, S.; Mahesh, H.; Shirke, M.D.; Biradar, H.; Uday, G.; Aruna, Y.; Lohithaswa, H.; Mohanrao, A.J.B.g. Genome and transcriptome sequence of finger millet (Eleusine coracana (L.) Gaertn.) provides insights into drought tolerance and nutraceutical properties. BMC Genom. 2017, 18, 465. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; David, J.; Thompkinson, D.; Seelam, B.S.; Rajput, H.; Morya, S. Effect of roasting on functional and phytochemical constituents of finger millet (Eleusine coracana L.). Pharma Innov. J. 2018, 7, 414–418. [Google Scholar]
- Sharma, D.; Jamra, G.; Singh, U.M.; Sood, S.; Kumar, A. Calcium biofortification: Three pronged molecular approaches for dissecting complex trait of calcium nutrition in finger millet (Eleusine coracana) for devising strategies of enrichment of food crops. Front. Plant Sci. 2017, 7, 2028. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Boote, K.; Kadiyala, M.; Nedumaran, S.; Gupta, S.; Srinivas, K.; Bantilan, M. An assessment of yield gains under climate change due to genetic modification of pearl millet. Sci. Total Environ. 2017, 601, 1226–1237. [Google Scholar] [CrossRef] [Green Version]
- Ullah, A.; Ahmad, I.; Ahmad, A.; Khaliq, T.; Saeed, U.; Habib-ur-Rahman, M.; Hussain, J.; Ullah, S.; Hoogenboom, G. Assessing climate change impacts on pearl millet under arid and semi-arid environments using CSM-CERES-Millet model. Environ. Sci. Pollut. Res. 2019, 26, 6745–6757. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, B.; Singh, S.; West, C.P.; Ritchie, G.L.; Trostle, C.L. Effect of deficit irrigation on physiology and forage yield of forage sorghum, pearl millet, and corn. Crop Sci. 2020, 60, 2167–2179. [Google Scholar] [CrossRef]
- Yadav, T.; Kumar, A.; Yadav, R.; Yadav, G.; Kumar, R.; Kushwaha, M. Salicylic acid and thiourea mitigate the salinity and drought stress on physiological traits governing yield in pearl millet-wheat. Saudi J. Biol. Sci. 2020, 27, 2010–2017. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, A.; Tsuji, W.; An, P.; Inanaga, S.; Murata, K. Effect of pre-and post-heading water deficit on growth and grain yield of four millets. Plant Prod. Sci. 2012, 15, 323–331. [Google Scholar] [CrossRef] [Green Version]
- Winkel, T.; Renno, J.-F.; Payne, W. Effect of the timing of water deficit on growth, phenology and yield of pearl millet (Pennisetum glaucum (L.) R. Br.) grown in Sahelian conditions. J. Exp. Bot. 1997, 48, 1001–1009. [Google Scholar] [CrossRef] [Green Version]
- Abraha, M.T.; Hussein, S.; Laing, M.; Assefa, K. Genetic management of drought in tef: Current status and future research directions. Glob. J. Crop Soil Sci. Plant Breed. 2015, 3, 156–161. [Google Scholar]
- Rockström, J.; Falkenmark, M. Agriculture: Increase water harvesting in Africa. Nat. News 2015, 519, 283. [Google Scholar] [CrossRef] [PubMed]
- Bita, C.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlenker, W.; Roberts, M.J. Estimating the Impact of Climate Change on Crop Yields: The Importance of Nonlinear Temperature Effects; National Bureau of Economic Research: Cambridge, MA, USA, 2008. [Google Scholar]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, G.C.; Rosegrant, M.W.; Koo, J.; Robertson, R.; Sulser, T.; Zhu, T.; Ringler, C.; Msangi, S.; Palazzo, A.; Batka, M. Climate Change: Impact on Agriculture and Costs of Adaptation; International Food Policy Research Institute: Washington, DC, USA, 2009; Volume 21. [Google Scholar]
- Warren, R.; Price, J.; Graham, E.; Forstenhaeusler, N.; VanDerWal, J. The projected effect on insects, vertebrates, and plants of limiting global warming to 1.5 C rather than 2 C. Science 2018, 360, 791–795. [Google Scholar] [CrossRef] [Green Version]
- Young, G.H.; Gagen, M.H.; Loader, N.J.; McCarroll, D.; Grudd, H.; Jalkanen, R.; Kirchhefer, A.; Robertson, I. Cloud Cover Feedback Moderates Fennoscandian Summer Temperature Changes Over the Past 1,000 Years. Geophys. Res. Lett. 2019, 46, 2811–2819. [Google Scholar] [CrossRef] [Green Version]
- Alhaithloul, H.A.S. Impact of Combined Heat and Drought Stress on the Potential Growth Responses of the Desert Grass Artemisia sieberi alba: Relation to Biochemical and Molecular Adaptation. Plants 2019, 8, 416. [Google Scholar] [CrossRef] [Green Version]
- Islam, F.; Yasmeen, T.; Ali, Q.; Ali, S.; Arif, M.S.; Hussain, S.; Rizvi, H. Influence of Pseudomonas aeruginosa as PGPR on oxidative stress tolerance in wheat under Zn stress. Ecotoxicol. Environ. Saf. 2014, 104, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Barnawal, D.; Bharti, N.; Pandey, S.S.; Pandey, A.; Chanotiya, C.S.; Kalra, A. Plant growth-promoting rhizobacteria enhance wheat salt and drought stress tolerance by altering endogenous phytohormone levels and TaCTR1/TaDREB2 expression. Physiol. Plant. 2017, 161, 502–514. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, A.; Shahbaz, M.; Asadullah, A.I.; Marghoob, M.U.; Imtiaz, M.; Mubeen, F. Potential of salt tolerant PGPR in growth and yield augmentation of wheat (Triticum aestivum L.) under saline conditions. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.; Chandrasekhar, C. Effect of PGPR on growth promotion of rice (Oryza sativa L.) under salt stress. Asian J. Plant Sci. Res. 2014, 4, 62–67. [Google Scholar]
- Ji, J.; Yuan, D.; Jin, C.; Wang, G.; Li, X.; Guan, C. Enhancement of growth and salt tolerance of rice seedlings (Oryza sativa L.) by regulating ethylene production with a novel halotolerant PGPR strain Glutamicibacter sp. YD01 containing ACC deaminase activity. Acta Physiol. Plant. 2020, 42, 42. [Google Scholar] [CrossRef]
- Joshi, B.; Chaudhary, A.; Singh, H.; Kumar, P.A. Prospective evaluation of individual and consortia plant growth promoting rhizobacteria for drought stress amelioration in rice (Oryza sativa L.). Plant Soil 2020, 457, 225–240. [Google Scholar] [CrossRef]
- Barrangou, R.; Doudna, J.A. Applications of CRISPR technologies in research and beyond. Nat. Biotechnol. 2016, 34, 933. [Google Scholar] [CrossRef]
- Mohanraju, P.; Makarova, K.S.; Zetsche, B.; Zhang, F.; Koonin, E.V.; Van der Oost, J. Diverse evolutionary roots and mechanistic variations of the CRISPR-Cas systems. Science 2016, 353, aad5147. [Google Scholar] [CrossRef] [Green Version]
- Vetriventhan, M.; Azevedo, V.C.; Upadhyaya, H.; Nirmalakumari, A.; Kane-Potaka, J.; Anitha, S.; Ceasar, S.A.; Muthamilarasan, M.; Bhat, B.V.; Hariprasanna, K. Genetic and genomic resources, and breeding for accelerating improvement of small millets: Current status and future interventions. Nucleus 2020, 63, 217–239. [Google Scholar] [CrossRef]
- Serba, D.D.; Yadav, R.S.; Varshney, R.K.; Gupta, S.; Mahalingam, G.; Srivastava, R.K.; Gupta, R.; Perumal, R.; Tesso, T.T. Genomic Designing of Pearl Millet: A Resilient Crop for Arid and Semi-arid Environments. In Genomic Designing of Climate-Smart Cereal Crops; Springer: Berlin/Heidelberg, Germany, 2020; pp. 221–286. [Google Scholar]
- Serba, D.D.; Yadav, R.S. Genomic tools in pearl millet breeding for drought tolerance: Status and prospects. Front. Plant Sci. 2016, 7, 1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennetzen, J.L.; Schmutz, J.; Wang, H.; Percifield, R.; Hawkins, J.; Pontaroli, A.C.; Estep, M.; Feng, L.; Vaughn, J.N.; Grimwood, J. Reference genome sequence of the model plant Setaria. Nat. Biotechnol. 2012, 30, 555–561. [Google Scholar] [CrossRef] [Green Version]
- Cannarozzi, G.; Plaza-Wüthrich, S.; Esfeld, K.; Larti, S.; Wilson, Y.S.; Girma, D.; de Castro, E.; Chanyalew, S.; Blösch, R.; Farinelli, L. Genome and transcriptome sequencing identifies breeding targets in the orphan crop tef (Eragrostis tef). BMC Genom. 2014, 15, 581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanBuren, R.; Wai, C.M.; Wang, X.; Pardo, J.; Yocca, A.E.; Wang, H.; Chaluvadi, S.R.; Han, G.; Bryant, D.; Edger, P.P. Exceptional subgenome stability and functional divergence in the allotetraploid Ethiopian cereal teff. Nat. Commun. 2020, 11, 884. [Google Scholar] [CrossRef] [Green Version]
- Varshney, R.K.; Shi, C.; Thudi, M.; Mariac, C.; Wallace, J.; Qi, P.; Zhang, H.; Zhao, Y.; Wang, X.; Rathore, A. Pearl millet genome sequence provides a resource to improve agronomic traits in arid environments. Nat. Biotechnol. 2017, 35, 969–976. [Google Scholar] [CrossRef] [Green Version]
- Zou, C.; Li, L.; Miki, D.; Li, D.; Tang, Q.; Xiao, L.; Rajput, S.; Deng, P.; Peng, L.; Jia, W. The genome of broomcorn millet. Nat. Commun. 2019, 10, 436. [Google Scholar] [CrossRef] [Green Version]
- Mulla, S.; Singh, S.K.; Singh, K.; Praveen, B. Climate Change and Agriculture: A Review of Crop Models. In Global Climate Change and Environmental Policy; Springer: Berlin/Heidelberg, Germany, 2020; pp. 423–435. [Google Scholar]
- Avendaño, A.R.; Garcia, D.P. Selection of Key Characteristics for Crops to Deal with Climate Change Through Quality Function Deployment. In Frontiers in Water-Energy-Nexus-Nature-Based Solutions, Advanced Technologies and Best Practices for Environmental Sustainability; Springer: Berlin/Heidelberg, Germany, 2020; pp. 75–77. [Google Scholar]
- Anderson, R.; Bayer, P.E.; Edwards, D. Climate change and the need for agricultural adaptation. Curr. Opin. Plant Biol. 2020, 56, 197–202. [Google Scholar] [CrossRef]
- Zilli, M.; Scarabello, M.; Soterroni, A.C.; Valin, H.; Mosnier, A.; Leclère, D.; Havlík, P.; Kraxner, F.; Lopes, M.A.; Ramos, F.M. The impact of climate change on Brazil’s agriculture. Sci. Total. Environ. 2020, 740, 139384. [Google Scholar] [CrossRef]
- Olabanji, M.F.; Ndarana, T.; Davis, N. Impact of Climate Change on Crop Production and Potential Adaptive Measures in the Olifants Catchment, South Africa. Climate 2021, 9, 6. [Google Scholar] [CrossRef]
- Vesco, P.; Kovacic, M.; Mistry, M.; Croicu, M. Climate variability, crop and conflict: Exploring the impacts of spatial concentration in agricultural production. J. Peace Res. 2021, 58, 98–113. [Google Scholar] [CrossRef]
- Tadele, Z. Drought Adaptation in Millets; InTech: London, UK, 2016. [Google Scholar]
- Maqsood, M.; Ali, S.A. Effects of drought on growth, development, radiation use efficiency and yield of finger millet (Eleucine coracana). Pak. J. Bot. 2007, 39, 123. [Google Scholar]
- Hadebe, S.; Modi, A.; Mabhaudhi, T. Drought tolerance and water use of cereal crops: A focus on sorghum as a food security crop in sub-Saharan Africa. J. Agron. Crop Sci. 2017, 203, 177–191. [Google Scholar] [CrossRef]
- Vadez, V.; Hash, T.; Kholova, J. II. 1.5 Phenotyping pearl millet for adaptation to drought. Front. Physiol. 2012, 3, 386. [Google Scholar] [CrossRef] [Green Version]
- Ashok, S.; Senthil, A.; Sritharan, N.; Punitha, S.; Divya, K.; Ravikesavan, R. Yield Potential of Small Millets under Drought Condition. Madras Agric. J. 2018, 105. [Google Scholar] [CrossRef]
- Mengistu, D.K.; Mekonnen, L.S. Integrated agronomic crop managements to improve tef productivity under terminal drought. Water Stress. Vienna Tech. 2011, 1, 235–254. [Google Scholar]
- Wondewosen, S.; Alemayehu, B.; Hussen, M. Genetic variation for grain yield and yield related traits in tef [Eragrostis tef (Zucc.) Trotter] under moisture stress and non-stress environments. Am. J. Plant Sci. 2012, 3, 2012. [Google Scholar]
- Sultan, B.; Roudier, P.; Quirion, P.; Alhassane, A.; Muller, B.; Dingkuhn, M.; Ciais, P.; Guimberteau, M.; Traore, S.; Baron, C. Assessing climate change impacts on sorghum and millet yields in the Sudanian and Sahelian savannas of West Africa. Environ. Res. Lett. 2013, 8, 014040. [Google Scholar] [CrossRef]
- Saleh, A.S.; Zhang, Q.; Chen, J.; Shen, Q. Millet grains: Nutritional quality, processing, and potential health benefits. Compr. Rev. Food Sci. Food Saf. 2013, 12, 281–295. [Google Scholar] [CrossRef]
- Ravindran, G. Studies on millets: Proximate composition, mineral composition, and phytate and oxalate contents. Food Chem. 1991, 39, 99–107. [Google Scholar] [CrossRef]
- Mickelbart, M.V.; Hasegawa, P.M.; Bailey-Serres, J. Genetic mechanisms of abiotic stress tolerance that translate to crop yield stability. Nat. Rev. Genet. 2015, 16, 237–251. [Google Scholar] [CrossRef] [PubMed]
- Duque, A.S.; de Almeida, A.M.; da Silva, A.B.; da Silva, J.M.; Farinha, A.P.; Santos, D.; Fevereiro, P.; de Sousa Araújo, S. Abiotic stress responses in plants: Unraveling the complexity of genes and networks to survive. In Abiotic Stress-Plant Responses and Applications in Agriculture; InTech: London, UK, 2013; pp. 49–101. [Google Scholar]
- Tadele, Z. Abiotic and Biotic Stress in Plants—Recent Advances and Future Perspectives; InTech: London, UK, 2016; p. 768. [Google Scholar]
- Plaza-Wüthrich, S.; Blösch, R.; Rindisbacher, A.; Cannarozzi, G.; Tadele, Z. Gibberellin Deficiency Confers Both Lodging and Drought Tolerance in Small Cereals. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusaka, M.; Lalusin, A.G.; Fujimura, T. The maintenance of growth and turgor in pearl millet (Pennisetum glaucum [L.] Leeke) cultivars with different root structures and osmo-regulation under drought stress. Plant Sci. 2005, 168, 1–14. [Google Scholar] [CrossRef]
- Debieu, M.; Sine, B.; Passot, S.; Grondin, A.; Akata, E.; Gangashetty, P.; Vadez, V.; Gantet, P.; Fonceka, D.; Cournac, L. Response to early drought stress and identification of QTLs controlling biomass production under drought in pearl millet. PLoS ONE 2018, 13, e0201635. [Google Scholar] [CrossRef] [Green Version]
- Chivenge, P.; Mabhaudhi, T.; Modi, A.; Mafongoya, P. The potential role of neglected and underutilised crop species as future crops under water scarce conditions in Sub-Saharan Africa. Int. J. Environ. Res. Public Health 2015, 12, 5685–5711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiferaw, B.; Baker, D.A. An evaluation of drought screening techniques for Eragrostis tef. Trop. Sci. 1996, 36, 74–85. [Google Scholar]
- Dichio, B.; Xiloyannis, C.; Angelopoulos, K.; Nuzzo, V.; Bufo, S.A.; Celano, G. Drought-induced variations of water relations parameters in Olea europaea. Plant Soil 2003, 257, 381–389. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S. Plant drought stress: Effects, mechanisms and management. In Sustainable Agriculture; Springer: Berlin/Heidelberg, Germany, 2009; pp. 153–188. [Google Scholar]
- Merrill, S.D.; Tanaka, D.L.; Hanson, J.D. Root length growth of eight crop species in Haplustoll soils. Soil Sci. Soc. Am. J. 2002, 66, 913–923. [Google Scholar] [CrossRef]
- Ayele, M.; Blum, A.; Nguyen, H.T. Diversity for osmotic adjustment and root depth in TEF [Eragrostis tef (Zucc) Trotter]. Euphytica 2001, 121, 237–249. [Google Scholar] [CrossRef]
- Barnabás, B.; Jäger, K.; Fehér, A. The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ. 2008, 31, 11–38. [Google Scholar] [CrossRef]
- Deryng, D.; Conway, D.; Ramankutty, N.; Price, J.; Warren, R. Global crop yield response to extreme heat stress under multiple climate change futures. Environ. Res. Lett. 2014, 9, 034011. [Google Scholar] [CrossRef] [Green Version]
- Lipiec, J.; Doussan, C.; Nosalewicz, A.; Kondracka, K. Effect of drought and heat stresses on plant growth and yield: A review. Int. Agrophysics 2013, 27, 463–477. [Google Scholar] [CrossRef]
- Ray, D.K.; West, P.C.; Clark, M.; Gerber, J.S.; Prishchepov, A.V.; Chatterjee, S. Climate change has likely already affected global food production. PLoS ONE 2019, 14, e0217148. [Google Scholar] [CrossRef] [PubMed]
- Asthir, B. Mechanisms of heat tolerance in crop plants. Biol. Plant. 2015, 59, 620–628. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Prueger, J.H. Temperature extremes: Effect on plant growth and development. Weather Clim. Extrem. 2015, 10, 4–10. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef]
- Gupta, N.; Agarwal, S.; Agarwal, V.; Nathawat, N.; Gupta, S.; Singh, G. Effect of short-term heat stress on growth, physiology and antioxidative defence system in wheat seedlings. Acta Physiol. Plant. 2013, 35, 1837–1842. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Ehlers, J.; Hall, A. Heat tolerance of contrasting cowpea lines in short and long days. Field Crops Res. 1998, 55, 11–21. [Google Scholar] [CrossRef]
- Thudi, M.; Upadhyaya, H.D.; Rathore, A.; Gaur, P.M.; Krishnamurthy, L.; Roorkiwal, M.; Nayak, S.N.; Chaturvedi, S.K.; Basu, P.S.; Gangarao, N. Genetic dissection of drought and heat tolerance in chickpea through genome-wide and candidate gene-based association mapping approaches. PLoS ONE 2014, 9, e96758. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.; Hafeez, M. Thermotolerance of pearl millet and maize at early growth stages: Growth and nutrient relations. Biol. Plant. 2004, 48, 81–86. [Google Scholar] [CrossRef]
- Linkemer, G.; Board, J.E.; Musgrave, M.E. Waterlogging effects on growth and yield components in late-planted soybean. Crop Sci. 1998, 38, 1576–1584. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, A.; An, P.; Murata, K.; Inanaga, S. Effect of pre-and post-heading waterlogging on growth and grain yield of four millets. Plant Prod. Sci. 2016, 19, 348–359. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.A.; Uddin, S.N. Mechanisms of waterlogging tolerance in wheat: Morphological and metabolic adaptations under hypoxia or anoxia. Aust. J. Crop Sci. 2011, 5, 1094. [Google Scholar]
- Kulkarni, S.; Chavan, P. Rice Plants Subjected to Waterlogging Stress. Int. J. Bot. 2013, 9, 80–85. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, S.; Chavan, P. Influence of waterlogging on carbohydrate metabolism in ragi and rice roots. J. Stress Physiol. Biochem. 2013, 9, 199–205. [Google Scholar]
- Parent, C.; Capelli, N.; Berger, A.; Crèvecoeur, M.; Dat, J.F. An overview of plant responses to soil waterlogging. Plant Stress 2008, 2, 20–27. [Google Scholar]
- Yin, D.; Chen, S.; Chen, F.; Guan, Z.; Fang, W. Morpho-anatomical and physiological responses of two Dendranthema species to waterlogging. Environ. Exp. Bot. 2010, 68, 122–130. [Google Scholar] [CrossRef]
- Ni, X.-L.; Gui, M.-Y.; Tan, L.-L.; Zhu, Q.; Liu, W.-Z.; Li, C.-X. Programmed cell death and aerenchyma formation in water-logged sunflower stems and its promotion by ethylene and ROS. Front. Plant Sci. 2018, 9, 1928. [Google Scholar] [CrossRef]
- Kulkarn, S.; Chavan, P. Study of effect of waterlogging on root anatomy of ragi and rice. Am. J. Plant Physiol. 2014, 9, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Promkhambut, A.; Younger, A.; Polthanee, A.; Akkasaeng, C. Morphological and physiological responses of sorghum (Sorghum bicolor L. Moench) to waterlogging. Asian J. Plant Sci. 2010, 9, 183. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Pal, M.; Joshi, R.; Sairam, R. Yield, growth and physiological responses of mung bean [Vigna radiata (L.) Wilczek] genotypes to waterlogging at vegetative stage. Physiol. Mol. Biol. Plants 2013, 19, 209–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sairam, R.; Kumutha, D.; Ezhilmathi, K.; Deshmukh, P.; Srivastava, G. Physiology and biochemistry of waterlogging tolerance in plants. Biol. Plant. 2008, 52, 401. [Google Scholar] [CrossRef]
- Cannarozzi, G.; Weichert, A.; Schnell, M.; Ruiz, C.; Bossard, S.; Blösch, R.; Plaza-Wüthrich, S.; Chanyalew, S.; Assefa, K.; Tadele, Z. Waterlogging affects plant morphology and the expression of key genes in tef (Eragrostis tef). Plant Direct 2018, 2, e00056. [Google Scholar] [CrossRef] [PubMed]
- Pinthus, M.J. Lodging in wheat, barley, and oats: The phenomenon, its causes, and preventive measures. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 1974; Volume 25, pp. 209–263. [Google Scholar]
- Piñera-Chavez, F.; Berry, P.; Foulkes, M.; Jesson, M.; Reynolds, M. Avoiding lodging in irrigated spring wheat. I. Stem and root structural requirements. Field Crops Res. 2016, 196, 325–336. [Google Scholar] [CrossRef]
- Van Delden, S.; Vos, J.; Ennos, A.; Stomph, T. Analysing lodging of the panicle bearing cereal teff (Eragrostis tef). New Phytol. 2010, 186, 696–707. [Google Scholar] [CrossRef]
- Berry, P.; Sterling, M.; Spink, J.; Baker, C.; Sylvester-Bradley, R.; Mooney, S.; Tams, A.; Ennos, A. Understanding and reducing lodging in cereals. Adv. Agron. 2004, 84, 215–269. [Google Scholar]
- Ben-Zeev, S.; Rabinovitz, O.; Orlov-Levin, V.; Chen, A.; Graff, N.; Goldwasser, Y.; Saranga, Y. Less is more: Lower sowing rate of irrigated tef (Eragrostis tef) alters plant morphology and reduces lodging. Agronomy 2020, 10, 570. [Google Scholar] [CrossRef]
- Tian, B.; Wang, J.; Zhang, L.; Li, Y.; Wang, S.; Li, H. Assessment of resistance to lodging of landrace and improved cultivars in foxtail millet. Euphytica 2010, 172, 295–302. [Google Scholar] [CrossRef]
- Opole, R.A. Effect of Environmental Stress and Management on Grain and Biomass Yield of Finger Millet (Eleusine coracana (L.) Gaertn.); Kansas State University: Manhattan, KS, USA, 2012. [Google Scholar]
- Shivhare, R.; Lata, C. Exploration of Genetic and Genomic Resources for Abiotic and Biotic Stress Tolerance in Pearl Millet. Front. Plant Sci. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Würschum, T.; Langer, S.M.; Longin, C.F.H.; Tucker, M.R.; Leiser, W.L. A modern Green Revolution gene for reduced height in wheat. Plant J. 2017, 92, 892–903. [Google Scholar] [CrossRef] [Green Version]
- Pardo, J.; Wai, C.M.; Chay, H.; Madden, C.F.; Hilhorst, H.W.; Farrant, J.M.; VanBuren, R. Intertwined signatures of desiccation and drought tolerance in grasses. Proc. Natl. Acad. Sci. USA 2020, 117, 10079–10088. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, J.R.; Koetz, M.; Bonfim-Silva, E.M.; da Silva, T.J.A. Production and accumulation of silicon (Si) in rice plants under silicate fertilization and soil water tensions. Aust. J. Crop Sci. 2016, 10, 244–250. [Google Scholar]
- Liang, Y.; Nikolic, M.; Bélanger, R.; Gong, H.; Song, A. Effect of silicon on crop growth, yield and quality. In Silicon in Agriculture; Springer: Berlin/Heidelberg, Germany, 2015; pp. 209–223. [Google Scholar]
- Agostinho, F.B.; Tubana, B.S.; Martins, M.S.; Datnoff, L.E. Effect of different silicon sources on yield and silicon uptake of rice grown under varying phosphorus rates. Plants 2017, 6, 35. [Google Scholar] [CrossRef]
- Ligaba-Osena, A.; Guo, W.; Choi, S.C.; Limmer, M.A.; Seyfferth, A.L.; Hankoua, B.B. Silicon enhances biomass and grain yield in an ancient crop tef [Eragrostis tef (Zucc.) Trotter]. Front. Plant Sci. 2020, 11, 608503. [Google Scholar] [CrossRef] [PubMed]
- Fallah, A. Silicon effect on lodging parameters of rice plants under hydroponic culture. Int. J. Agric. Sci. 2012, 2, 630–634. [Google Scholar]
- Rademacher, W. Growth retardants: Effects on gibberellin biosynthesis and other metabolic pathways. Annu. Rev. Plant Biol. 2000, 51, 501–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanvicente, P.; Lazarevitch, S.; Blouet, A.; Guckert, A. Morphological and anatomical modifications in winter barley culm after late plant growth regulator treatment. Eur. J. Agron. 1999, 11, 45–51. [Google Scholar] [CrossRef]
- Hedden, P. The genes of the Green Revolution. Trends Genet. 2003, 19, 5–9. [Google Scholar] [CrossRef]
- Hedden, P.; Graebe, J.E. Inhibition of gibberellin biosynthesis by paclobutrazol in cell-free homogenates of Cucurbita maxima endosperm and Malus pumila embryos. J. Plant Growth Regul. 1985, 4, 111–122. [Google Scholar] [CrossRef]
- Bizuayehu, D.; Getachew, A. Paclobutrazol as a plant growth regulator. Chem. Biol. Technol. Agric. 2021, 8. [Google Scholar] [CrossRef]
- Jency, J.P.; Rajasekaran, R.; Singh, R.K.; Muthurajan, R.; Prabhakaran, J.; Mehanathan, M.; Prasad, M.; Ganesan, J. Induced mutagenesis enhances lodging resistance and photosynthetic efficiency of kodomillet (Paspalum scrobiculatum). Agronomy 2020, 10, 227. [Google Scholar] [CrossRef] [Green Version]
- Jöst, M.; Esfeld, K.; Burian, A.; Cannarozzi, G.; Chanyalew, S.; Kuhlemeier, C.; Assefa, K.; Tadele, Z. Semi-dwarfism and lodging tolerance in tef (Eragrostis tef) is linked to a mutation in the α-Tubulin 1 gene. J. Exp. Bot. 2015, 66, 933–944. [Google Scholar] [CrossRef] [Green Version]
- Bidinger, F.; Mahalakshmi, V.; Rao, G.D.P. Assessment of drought resistance in pearl millet (Pennisetum americanum (L.) Leeke). II. Estimation of genotype response to stress. Aust. J. Agric. Res. 1987, 38, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Sahu, R.K. Study on Technological Gap, Yield Gap and Utilization Pattern of Small Millets among the Tribals in the Bastar Plateau Zone of Chhattisgarh. Doctoral Dissertation, Indira Gandhi Krishi Vishwavidyalaya, Raipur, India, 2010. Corpus ID: 132183334. [Google Scholar]
- Assefa, K.; Yu, J.K.; Zeid, M.; Belay, G.; Tefera, H.; Sorrells, M. Breeding tef [Eragrostis tef (Zucc.) trotter]: Conventional and molecular approaches. Plant Breed. 2011, 130, 1–9. [Google Scholar] [CrossRef]
- Tian, B.; Luan, S.; Zhang, L.; Liu, Y.; Zhang, L.; Li, H. Penalties in yield and yield associated traits caused by stem lodging at different developmental stages in summer and spring foxtail millet cultivars. Field Crops Res. 2018, 217, 104–112. [Google Scholar] [CrossRef]
- Opole, R.; Prasad, P.; Djanaguiraman, M.; Vimala, K.; Kirkham, M.; Upadhyaya, H. Thresholds, sensitive stages and genetic variability of finger millet to high temperature stress. J. Agron. Crop Sci. 2018, 204, 477–492. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Perumal, R.; Ciampitti, I.; Gupta, S.; Prasad, P. Quantifying pearl millet response to high temperature stress: Thresholds, sensitive stages, genetic variability and relative sensitivity of pollen and pistil. Plant Cell Environ. 2018, 41, 993–1007. [Google Scholar] [CrossRef] [Green Version]
- Aidoo, M.K.; Bdolach, E.; Fait, A.; Lazarovitch, N.; Rachmilevitch, S. Tolerance to high soil temperature in foxtail millet (Setaria italica L.) is related to shoot and root growth and metabolism. Plant Physiol. Biochem. 2016, 106, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D.; Davranov, K.; Wirth, S.; Hashem, A.; Abd_Allah, E.F. Impact of soil salinity on the plant-growth—Promoting and biological control abilities of root associated bacteria. Saudi J. Biol. Sci. 2017, 24, 1601–1608. [Google Scholar] [CrossRef]
- Gamalero, E.; Berta, G.; Massa, N.; Glick, B.R.; Lingua, G. Interactions between Pseudomonas putida UW4 and Gigaspora rosea BEG9 and their consequences for the growth of cucumber under salt-stress conditions. J. Appl. Microbiol. 2010, 108, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Bal, H.B.; Nayak, L.; Das, S.; Adhya, T.K. Isolation of ACC deaminase producing PGPR from rice rhizosphere and evaluating their plant growth promoting activity under salt stress. Plant Soil 2013, 366, 93–105. [Google Scholar] [CrossRef]
- Tewari, S.; Arora, N.K.; Miransari, M. Plant growth promoting rhizobacteria to alleviate soybean growth under abiotic and biotic stresses. In Abiotic and Biotic Stresses in Soybean Production; Elsevier: Amsterdam, The Netherlands, 2016; pp. 131–155. [Google Scholar]
- Fasciglione, G.; Casanovas, E.M.; Yommi, A.; Sueldo, R.J.; Barassi, C.A. Azospirillum improves lettuce growth and transplant under saline conditions. J. Sci. Food Agric. 2012, 92, 2518–2523. [Google Scholar] [CrossRef]
- Palaniyandi, S.; Damodharan, K.; Yang, S.; Suh, J. Streptomyces sp. strain PGPA39 alleviates salt stress and promotes growth of ‘Micro Tom’tomato plants. J. Appl. Microbiol. 2014, 117, 766–773. [Google Scholar] [CrossRef]
- Curá, J.; Franz, D.; Filosofía, J.; Balestrasse, K.; Burgueño, L. Inoculation with Azospirillum sp. and Herbaspirillum sp. bacteria increases the tolerance of maize to drought stress. Microorganisms 2017, 5, 41. [Google Scholar] [CrossRef] [Green Version]
- Asaf, S.; Khan, A.L.; Khan, M.A.; Imran, Q.M.; Yun, B.-W.; Lee, I.-J. Osmoprotective functions conferred to soybean plants via inoculation with Sphingomonas sp. LK11 and exogenous trehalose. Microbiol. Res. 2017, 205, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Timmusk, S.; Abd El-Daim, I.A.; Copolovici, L.; Tanilas, T.; Kännaste, A.; Behers, L.; Nevo, E.; Seisenbaeva, G.; Stenström, E.; Niinemets, Ü. Drought-Tolerance of Wheat Improved by Rhizosphere Bacteria from Harsh Environments: Enhanced Biomass Production and Reduced Emissions of Stress Volatiles. PLoS ONE 2014, 9, e96086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol. Res. 2016, 184, 13–24. [Google Scholar] [CrossRef]
- Castillo, P.; Molina, R.; Andrade, A.; Vigliocco, A.; Alemano, S.; Cassán, F.D. Phytohormones and other plant growth regulators produced by PGPR: The genus Azospirillum. In Handbook for Azospirillum; Springer: Berlin/Heidelberg, Germany, 2015; pp. 115–138. [Google Scholar]
- Jogaiah, S.; Abdelrahman, M.; Tran, L.S.P.; Ito, S.I. Different mechanisms of Trichoderma virens-mediated resistance in tomato against Fusarium wilt involve the jasmonic and salicylic acid pathways. Mol. Plant Pathol. 2018, 19, 870–882. [Google Scholar] [CrossRef] [Green Version]
- Ghassemi-Golezani, K.; Farhangi-Abriz, S. Foliar sprays of salicylic acid and jasmonic acid stimulate H+-ATPase activity of tonoplast, nutrient uptake and salt tolerance of soybean. Ecotoxicol. Environ. Saf. 2018, 166, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Dimkpa, C.; Weinand, T.; Asch, F. Plant–rhizobacteria interactions alleviate abiotic stress conditions. Plant Cell Environ. 2009, 32, 1682–1694. [Google Scholar] [CrossRef] [PubMed]
- Mantelin, S.; Touraine, B. Plant growth-promoting bacteria and nitrate availability: Impacts on root development and nitrate uptake. J. Exp. Bot. 2004, 55, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Egamberdieva, D.; Kucharova, Z. Selection for root colonising bacteria stimulating wheat growth in saline soils. Biol. Fertil. Soils 2009, 45, 563–571. [Google Scholar] [CrossRef]
- Cassán, F.; Bottini, R.; Schneider, G.; Piccoli, P. Azospirillum brasilense and Azospirillum lipoferum hydrolyze conjugates of GA20 and metabolize the resultant aglycones to GA1 in seedlings of rice dwarf mutants. Plant Physiol. 2001, 125, 2053–2058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molina-Favero, C.; Creus, C.M.; Simontacchi, M.; Puntarulo, S.; Lamattina, L. Aerobic nitric oxide production by Azospirillum brasilense Sp245 and its influence on root architecture in tomato. Mol. Plant-Microbe Interact. 2008, 21, 1001–1009. [Google Scholar] [CrossRef] [Green Version]
- German, M.A.; Burdman, S.; Okon, Y.; Kigel, J. Effects of Azospirillum brasilense on root morphology of common bean (Phaseolus vulgaris L.) under different water regimes. Biol. Fertil. Soils 2000, 32, 259–264. [Google Scholar] [CrossRef]
- Bashan, Y.; Holguin, G.; De-Bashan, L.E. Azospirillum-plant relationships: Physiological, molecular, agricultural, and environmental advances (1997–2003). Can. J. Microbiol. 2004, 50, 521–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sueldo, R.; Invernati, A.; Plaza, S.; Barassi, C. Osmotic stress in wheat seedlings: Effects on fatty acid composition and phospholipid turnover in coleoptiles. Cereal Res. Commun. 1996, 24, 77–84. [Google Scholar]
- Pereyra, M.; Zalazar, C.; Barassi, C. Root phospholipids in Azospirillum-inoculated wheat seedlings exposed to water stress. Plant Physiol. Biochem. 2006, 44, 873–879. [Google Scholar] [CrossRef] [PubMed]
- Gusain, Y.S.; Singh, U.; Sharma, A. Bacterial mediated amelioration of drought stress in drought tolerant and susceptible cultivars of rice (Oryza sativa L.). Afr. J. Biotechnol. 2015, 14, 764–773. [Google Scholar]
- Polko, J.K.; Kieber, J.J. 1-aminocyclopropane 1-carboxylic acid and its emerging role as an ethylene-independent growth regulator. Front. Plant Sci. 2019, 10, 1602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etesami, H.; Noori, F.; Ebadi, A.; Samani, N.R. Alleviation of Stress-Induced Ethylene-Mediated Negative Impact on Crop Plants by Bacterial ACC Deaminase: Perspectives and Applications in Stressed Agriculture Management. Plant Microbiomes Sustain. Agric. 2020, 25, 287–315. [Google Scholar]
- Glick, B.R. Modulation of plant ethylene levels by the bacterial enzyme ACC deaminase. FEMS Microbiol. Lett. 2005, 251, 1–7. [Google Scholar] [CrossRef]
- Mayak, S.; Tirosh, T.; Glick, B.R. Plant growth-promoting bacteria that confer resistance to water stress in tomatoes and peppers. Plant Sci. 2004, 166, 525–530. [Google Scholar] [CrossRef]
- Lifshitz, R.; Kloepper, J.W.; Scher, F.M.; Tipping, E.M.; Laliberté, M. Nitrogen-fixing pseudomonads isolated from roots of plants grown in the Canadian High Arctic. Appl. Env. Microbiol. 1986, 51, 251–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kai, M.; Haustein, M.; Molina, F.; Petri, A.; Scholz, B.; Piechulla, B. Bacterial volatiles and their action potential. Appl. Microbiol. Biotechnol. 2009, 81, 1001–1012. [Google Scholar] [CrossRef]
- Ryu, C.-M.; Farag, M.A.; Hu, C.-H.; Reddy, M.S.; Kloepper, J.W.; Paré, P.W. Bacterial volatiles induce systemic resistance in Arabidopsis. Plant Physiol. 2004, 134, 1017–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, C.-M.; Farag, M.A.; Paré, P.W.; Kloepper, J.W. Invisible signals from the underground: Bacterial volatiles elicit plant growth promotion and induce systemic resistance. Plant Pathol. J. 2005, 21, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Farag, M.A.; Zhang, H.; Ryu, C.-M. Dynamic chemical communication between plants and bacteria through airborne signals: Induced resistance by bacterial volatiles. J. Chem. Ecol. 2013, 39, 1007–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoro, M.V.; Zygadlo, J.; Giordano, W.; Banchio, E. Volatile organic compounds from rhizobacteria increase biosynthesis of essential oils and growth parameters in peppermint (Mentha piperita). Plant Physiol. Biochem. 2011, 49, 1177–1182. [Google Scholar] [CrossRef]
- Loreto, F.; Schnitzler, J.-P. Abiotic stresses and induced BVOCs. Trends Plant Sci. 2010, 15, 154–166. [Google Scholar] [CrossRef]
- Holopainen, J.K.; Gershenzon, J. Multiple stress factors and the emission of plant VOCs. Trends Plant Sci. 2010, 15, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Heil, M.; Bueno, J.C.S. Within-plant signaling by volatiles leads to induction and priming of an indirect plant defense in nature. Proc. Natl. Acad. Sci. USA 2007, 104, 5467–5472. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, D.K.; Johri, B.N.; Prakash, A. Volatiles as priming agents that initiate plant growth and defence responses. Curr. Sci. 2008, 94, 595–604. [Google Scholar]
- Cho, S.M.; Kang, B.R.; Han, S.H.; Anderson, A.J.; Park, J.-Y.; Lee, Y.-H.; Cho, B.H.; Yang, K.-Y.; Ryu, C.-M.; Kim, Y.C. 2R, 3R-butanediol, a bacterial volatile produced by Pseudomonas chlororaphis O6, is involved in induction of systemic tolerance to drought in Arabidopsis thaliana. Mol. Plant-Microbe Interact. 2008, 21, 1067–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, M.; Sakina, A.; Wani, S.H.; Shikari, A.B.; Tripathi, P.; Zaid, A.; Galla, A.; Abdelrahman, M.; Sharma, M.; Singh, A.K. Harnessing genome editing techniques to engineer disease resistance in plants. Front. Plant Sci. 2019, 10, 550. [Google Scholar] [CrossRef] [Green Version]
- Malzahn, A.; Lowder, L.; Qi, Y. Plant genome editing with TALEN and CRISPR. Cell Biosci. 2017, 7, 21. [Google Scholar] [CrossRef] [Green Version]
- Waltz, E. With a free pass, CRISPR-edited plants reach market in record time. Nat. Biotechnol. 2018. [Google Scholar] [CrossRef]
- Ricroch, A.; Clairand, P.; Harwood, W. Use of CRISPR systems in plant genome editing: Toward new opportunities in agriculture. Emerg. Top. Life Sci. 2017, 1, 169–182. [Google Scholar]
- Jaganathan, D.; Ramasamy, K.; Sellamuthu, G.; Jayabalan, S.; Venkataraman, G. CRISPR for crop improvement: An update review. Front. Plant Sci. 2018, 9, 985. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Gao, H.; Wang, H.; Lafitte, H.R.; Archibald, R.L.; Yang, M.; Hakimi, S.M.; Mo, H.; Habben, J.E. ARGOS 8 variants generated by CRISPR-Cas9 improve maize grain yield under field drought stress conditions. Plant Biotechnol. J. 2017, 15, 207–216. [Google Scholar] [CrossRef] [Green Version]
- Zafar, S.A.; Zaidi, S.S.-e.-A.; Gaba, Y.; Singla-Pareek, S.L.; Dhankher, O.P.; Li, X.; Mansoor, S.; Pareek, A. Engineering abiotic stress tolerance via CRISPR/Cas-mediated genome editing. J. Exp. Bot. 2020, 71, 470–479. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.J. Artificial trans-acting small interfering RNA: A tool for plant biology study and crop improvements. Planta 2014, 239, 1139–1146. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Lu, X.; Shu, N.; Wang, S.; Wang, J.; Wang, D.; Guo, L.; Ye, W. Targeted mutagenesis in cotton (Gossypium hirsutum L.) using the CRISPR/Cas9 system. Sci. Rep. 2017, 7, 44304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Char, S.N.; Neelakandan, A.K.; Nahampun, H.; Frame, B.; Main, M.; Spalding, M.H.; Becraft, P.W.; Meyers, B.C.; Walbot, V.; Wang, K. An Agrobacterium-delivered CRISPR/Cas9 system for high-frequency targeted mutagenesis in maize. Plant Biotechnol. J. 2017, 15, 257–268. [Google Scholar] [CrossRef]
- Svitashev, S.; Schwartz, C.; Lenderts, B.; Young, J.K.; Cigan, A.M. Genome editing in maize directed by CRISPR–Cas9 ribonucleoprotein complexes. Nat. Commun. 2016, 7, 13274. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Guo, D.; Zhang, J.; Huang, Q.; Qin, G.; Zhang, X.; Wan, J.; Gu, H.; Qu, L.-J. Targeted mutagenesis in rice using CRISPR-Cas system. Cell Res. 2013, 23, 1233–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, Q.; Wang, Y.; Li, J.; Gao, C. Genome editing in rice and wheat using the CRISPR/Cas system. Nat. Protoc. 2014, 9, 2395–2410. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-León, S.; Gil-Humanes, J.; Ozuna, C.V.; Giménez, M.J.; Sousa, C.; Voytas, D.F.; Barro, F. Low-gluten, nontransgenic wheat engineered with CRISPR/Cas9. Plant Biotechnol. J. 2018, 16, 902–910. [Google Scholar] [CrossRef] [PubMed]
- Klap, C.; Yeshayahou, E.; Bolger, A.M.; Arazi, T.; Gupta, S.K.; Shabtai, S.; Usadel, B.; Salts, Y.; Barg, R. Tomato facultative parthenocarpy results from Sl AGAMOUS-LIKE 6 loss of function. Plant Biotechnol. J. 2017, 15, 634–647. [Google Scholar] [CrossRef] [PubMed]
- Martignago, D.; Rico-Medina, A.; Blasco-Escámez, D.; Fontanet-Manzaneque, J.B.; Caño-Delgado, A.I. Drought Resistance by Engineering Plant Tissue-Specific Responses. Front. Plant Sci. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Long, L.; Tian, X.; Xu, F.; Liu, J.; Singh, P.K.; Botella, J.R.; Song, C. Genome editing in cotton with the CRISPR/Cas9 system. Front. Plant Sci. 2017, 8, 1364. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Wang, S.; Liang, Z.; Shi, W.; Gao, C.; Xia, G. From genetic stock to genome editing: Gene exploitation in wheat. Trends Biotechnol. 2018, 36, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Leal, D.; Lemmon, Z.H.; Man, J.; Bartlett, M.E.; Lippman, Z.B. Engineering quantitative trait variation for crop improvement by genome editing. Cell 2017, 171, 470–480.e8. [Google Scholar] [CrossRef] [Green Version]
- Mahas, A.; Mahfouz, M. Engineering virus resistance via CRISPR–Cas systems. Curr. Opin. Virol. 2018, 32, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Kaur, R.; Singh, A. Recent advances in CRISPR/Cas mediated genome editing for crop improvement. Plant Biotechnol. Rep. 2017, 11, 193–207. [Google Scholar] [CrossRef]
- Bhat, S.; Srinivasan, S. Molecular and genetic analyses of transgenic plants:: Considerations and approaches. Plant Sci. 2002, 163, 673–681. [Google Scholar] [CrossRef]
- Eid, A.; Alshareef, S.; Mahfouz, M.M. CRISPR base editors: Genome editing without double-stranded breaks. Biochem. J. 2018, 475, 1955–1964. [Google Scholar] [CrossRef]
- Ning, Y.; Jantasuriyarat, C.; Zhao, Q.; Zhang, H.; Chen, S.; Liu, J.; Liu, L.; Tang, S.; Park, C.H.; Wang, X. The SINA E3 ligase OsDIS1 negatively regulates drought response in rice. Plant Physiol. 2011, 157, 242–255. [Google Scholar] [CrossRef] [Green Version]
- Sharma, G.; Giri, J.; Tyagi, A.K. Rice OsiSAP7 negatively regulates ABA stress signalling and imparts sensitivity to water-deficit stress in Arabidopsis. Plant Sci. 2015, 237, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Xie, K.; Xiong, L. Conserved miR164-targeted NAC genes negatively regulate drought resistance in rice. J. Exp. Bot. 2014, 65, 2119–2135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, K.H.; Van Ha, C.; Nishiyama, R.; Watanabe, Y.; Leyva-González, M.A.; Fujita, Y.; Tran, U.T.; Li, W.; Tanaka, M.; Seki, M. Arabidopsis type B cytokinin response regulators ARR1, ARR10, and ARR12 negatively regulate plant responses to drought. Proc. Natl. Acad. Sci. USA 2016, 113, 3090–3095. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-C.; Wu, Y.-R.; Huang, X.-H.; Sun, J.; Xie, Q. AtPUB19, a U-box E3 ubiquitin ligase, negatively regulates abscisic acid and drought responses in Arabidopsis thaliana. Mol. Plant 2011, 4, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Yao, H.; Zhao, H.; Zhou, J.; Luo, X.; Huang, Y.; Li, C.; Chen, H. Tartary buckwheat FtMYB10 encodes an R2R3-MYB transcription factor that acts as a novel negative regulator of salt and drought response in transgenic Arabidopsis. Plant Physiol. Biochem. 2016, 109, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.-C.; Hsieh, E.-J.; Chen, J.-H.; Chen, H.-Y.; Lin, T.-P. Arabidopsis RGLG2, functioning as a RING E3 ligase, interacts with AtERF53 and negatively regulates the plant drought stress response. Plant Physiol. 2012, 158, 363–375. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Jia, H.; Chen, X.; Hao, L.; An, H.; Guo, X. The cotton WRKY transcription factor GhWRKY17 functions in drought and salt stress in transgenic Nicotiana benthamiana through ABA signaling and the modulation of reactive oxygen species production. Plant Cell Physiol. 2014, 55, 2060–2076. [Google Scholar] [CrossRef] [Green Version]
- Satapathy, L.; Kumar, D.; Kumar, M.; Mukhopadhyay, K. Functional and DNA–protein binding studies of WRKY transcription factors and their expression analysis in response to biotic and abiotic stress in wheat (Triticum aestivum L.). 3 Biotech. 2018, 8, 40. [Google Scholar] [CrossRef]
- Ma, J.; Gao, X.; Liu, Q.; Shao, Y.; Zhang, D.; Jiang, L.; Li, C. Overexpression of TaWRKY146 increases drought tolerance through inducing stomatal closure in Arabidopsis thaliana. Front. Plant Sci. 2017, 8, 2036. [Google Scholar] [CrossRef]
- Wang, Y.; Tong, X.; Qiu, J.; Li, Z.; Zhao, J.; Hou, Y.; Tang, L.; Zhang, J. A phosphoproteomic landscape of rice (Oryza sativa) tissues. Physiol. Plant. 2017, 160, 458–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soltani Najafabadi, M. Improving rice (Oryza sativa L.) drought tolerance by suppressing a NF-YA transcription factor. Iran. J. Biotechnol. 2012, 10, 40–48. [Google Scholar]
- Capell, T.; Escobar, C.; Liu, H.; Burtin, D.; Lepri, O.; Christou, P. Over-expression of the oat arginine decarboxylase cDNA in transgenic rice (Oryza sativa L.) affects normal development patterns in vitro and results in putrescine accumulation in transgenic plants. Theor. Appl. Genet. 1998, 97, 246–254. [Google Scholar] [CrossRef]
- Li, X.-Y.; Mantovani, R.; Hooft van Huijsduijnen, R.; Andre, I.; Benoist, C.; Mathis, D. Evolutionary variation of the CCAAT-binding transcription factor NF-Y. Nucleic Acids Res. 1992, 20, 1087–1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Sun, F.; Cao, H.; Peng, H.; Ni, Z.; Sun, Q.; Yao, Y. TamiR159 directed wheat TaGAMYB cleavage and its involvement in anther development and heat response. PLoS ONE 2012, 7, e48445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, G.-P.; Drenth, J.; McIntyre, C.L. TaHsfA6f is a transcriptional activator that regulates a suite of heat stress protection genes in wheat (Triticum aestivum L.) including previously unknown Hsf targets. J. Exp. Bot. 2015, 66, 1025–1039. [Google Scholar] [CrossRef] [Green Version]
- Qin, D.; Wang, F.; Geng, X.; Zhang, L.; Yao, Y.; Ni, Z.; Peng, H.; Sun, Q. Overexpression of heat stress-responsive TaMBF1c, a wheat (Triticum aestivum L.) Multiprotein Bridging Factor, confers heat tolerance in both yeast and rice. Plant Mol. Biol. 2015, 87, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Zang, X.; Geng, X.; Wang, F.; Liu, Z.; Zhang, L.; Zhao, Y.; Tian, X.; Ni, Z.; Yao, Y.; Xin, M. Overexpression of wheat ferritin gene TaFER-5B enhances tolerance to heat stress and other abiotic stresses associated with the ROS scavenging. BMC Plant Biol. 2017, 17, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zang, X.; Geng, X.; Liu, K.; Wang, F.; Liu, Z.; Zhang, L.; Zhao, Y.; Tian, X.; Hu, Z.; Yao, Y. Ectopic expression of TaOEP16-2-5B, a wheat plastid outer envelope protein gene, enhances heat and drought stress tolerance in transgenic Arabidopsis plants. Plant Sci. 2017, 258, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Geng, X.; Zhang, H.; Zhou, C.; Zhao, A.; Wang, F.; Zhao, Y.; Tian, X.; Hu, Z.; Xin, M. Isolation and characterization of heat-responsive gene TaGASR1 from wheat (Triticum aestivum L.). J. Plant Biol. 2017, 60, 57–65. [Google Scholar] [CrossRef]
- Singh, A.; Khurana, P. Molecular and functional characterization of a wheat B2 protein imparting adverse temperature tolerance and influencing plant growth. Front. Plant Sci. 2016, 7, 642. [Google Scholar] [CrossRef] [Green Version]
- Katiyar-Agarwal, S.; Agarwal, M.; Grover, A. Heat-tolerant basmati rice engineered by over-expression of hsp101. Plant Mol. Biol. 2003, 51, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Liu, J.; Wang, Y.; Huang, N.; Zhang, X.; Wang, L.; Zhang, J.; Tu, J.; Zhong, X. A dominant major locus in chromosome 9 of rice (Oryza sativa L.) confers tolerance to 48 C high temperature at seedling stage. J. Hered. 2013, 104, 287–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, S.; Lee, S.-C.; Kim, M.-K.; Koh, J.H.; Lee, S.; An, G.; Choe, S.; Kim, S.-R. T-DNA tagged knockout mutation of rice OsGSK1, an orthologue of Arabidopsis BIN2, with enhanced tolerance to various abiotic stresses. Plant Mol. Biol. 2007, 65, 453–466. [Google Scholar] [CrossRef] [PubMed]
- Yokotani, N.; Ichikawa, T.; Kondou, Y.; Matsui, M.; Hirochika, H.; Iwabuchi, M.; Oda, K. Expression of rice heat stress transcription factor OsHsfA2e enhances tolerance to environmental stresses in transgenic Arabidopsis. Planta 2008, 227, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Wang, H.; Zou, Y.; Liu, C.; Liu, Y.; Wang, Y.; Zhang, W. Over-expression of mitochondrial heat shock protein 70 suppresses programmed cell death in rice. FEBS Lett. 2011, 585, 231–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami, T.; Matsuba, S.; Funatsuki, H.; Kawaguchi, K.; Saruyama, H.; Tanida, M.; Sato, Y. Over-expression of a small heat shock protein, sHSP17. 7, confers both heat tolerance and UV-B resistance to rice plants. Mol. Breed. 2004, 13, 165–175. [Google Scholar] [CrossRef]
- Sohn, S.; Back, K. Transgenic rice tolerant to high temperature with elevated contents of dienoic fatty acids. Biol. Plant 2007, 51, 340–342. [Google Scholar] [CrossRef]
- Feng, L.; Wang, K.; Li, Y.; Tan, Y.; Kong, J.; Li, H.; Li, Y.; Zhu, Y. Overexpression of SBPase enhances photosynthesis against high temperature stress in transgenic rice plants. Plant Cell Rep. 2007, 26, 1635–1646. [Google Scholar] [CrossRef]
- Yamanouchi, U.; Yano, M.; Lin, H.; Ashikari, M.; Yamada, K. A rice spotted leaf gene, Spl7, encodes a heat stress transcription factor protein. Proc. Natl. Acad. Sci. USA 2002, 99, 7530–7535. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Wei, X.; Adedze, Y.; Sheng, Z.; Tang, S.; Hu, P.; Wang, J. Characterization and gene cloning of the rice (Oryza sativa L.) dwarf and narrow-leaf mutant dnl3. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Xue, Y.; Yang, H.; Li, H.; Sun, G.; Zhao, X.; Ding, D.; Tang, J. Genome-wide identification of miRNAs and their targets involved in the developing internodes under maize ears by responding to hormone signaling. PLoS ONE 2016, 11, e0164026. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Gu, L.; Li, P.; Song, X.; Wei, L.; CHen, Z.; Cao, X. Degradome sequencing reveals endogenous small RNA targets in rice (Oryza sativa L. ssp. indica). Front. Biol. 2010, 5, 67–90. [Google Scholar] [CrossRef]
- Yang, Y.-h.; Zhang, F.-m.; Ge, S. Evolutionary rate patterns of the Gibberellin pathway genes. BMC Evol. Biol. 2009, 9, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singla, B.; Khurana, J.P.; Khurana, P. Characterization of three somatic embryogenesis receptor kinase genes from wheat, Triticum aestivum. Plant Cell Rep. 2008, 27, 833–843. [Google Scholar] [CrossRef] [PubMed]
- Pinthus, M.; Levy, A. The relationship between the Rht 1 and Rht 2 dwarfing genes and grain weight in Triticum aestivum L. spring wheat. Theor. Appl. Genet. 1983, 66, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Spielmeyer, W.; Ellis, M.H.; Chandler, P.M. Semidwarf (sd-1), “green revolution” rice, contains a defective gibberellin 20-oxidase gene. Proc. Natl. Acad. Sci. USA 2002, 99, 9043–9048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mamidi, S.; Healey, A.; Huang, P.; Grimwood, J.; Jenkins, J.; Barry, K.; Sreedasyam, A.; Shu, S.; Lovell, J.T.; Feldman, M. A genome resource for green millet Setaria viridis enables discovery of agronomically valuable loci. Nat. Biotechnol. 2020, 38, 1203–1210. [Google Scholar] [CrossRef] [PubMed]
- Ceasar, S.A.; Baker, A.; Ignacimuthu, S. Functional characterization of the PHT1 family transporters of foxtail millet with development of a novel Agrobacterium-mediated transformation procedure. Sci. Rep. 2017, 7, 14064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceasar, S.A.; Ignacimuthu, S. Agrobacterium-mediated transformation of finger millet (Eleusine coracana (L.) Gaertn.) using shoot apex explants. Plant Cell Rep. 2011, 30, 1759–1770. [Google Scholar] [CrossRef]
- Satish, L.; Ceasar, S.A.; Ramesh, M. Improved Agrobacterium-mediated transformation and direct plant regeneration in four cultivars of finger millet (Eleusine coracana (L.) Gaertn.). Plant Cell Tissue Organ. Cult. 2017, 131, 547–565. [Google Scholar] [CrossRef]
- Johnson, M.; Deshpande, S.; Vetriventhan, M.; Upadhyaya, H.D.; Wallace, J.G. Genome-Wide Population Structure Analyses of Three Minor Millets: Kodo Millet, Little Millet, and Proso Millet. Plant Genome 2019, 12, 190021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, R.; Zhang, B. Foxtail Millet: A New Model for C4 Plants. Trends Plant Sci. 2020, 26, 199–201. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhang, H.; Li, X.; Shen, H.; Gao, J.; Hou, S.; Zhang, B.; Mayes, S.; Bennett, M.; Ma, J. A mini foxtail millet with an Arabidopsis-like life cycle as a C 4 model system. Nat. Plants 2020, 6, 1167–1178. [Google Scholar] [CrossRef] [PubMed]
- Numan, M.; Khan, A.L.; Asaf, S.; Salehin, M.; Beyene, G.; Tadele, Z.; Ligaba-Osena, A. From Traditional Breeding to Genome Editing for Boosting Productivity of the Ancient Grain Tef [Eragrostis tef (Zucc.) Trotter]. Plants 2021, 10, 628. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Potential application of PGPRs in abiotic stress mitigation and crop yield improvement in millets. PGPRs can be isolated from plants harboring the bacteria. The PGPRs are then cultured in a laboratory and applied to the soil of plant-growing substrate.
Figure 1.
Potential application of PGPRs in abiotic stress mitigation and crop yield improvement in millets. PGPRs can be isolated from plants harboring the bacteria. The PGPRs are then cultured in a laboratory and applied to the soil of plant-growing substrate.

Figure 2.
Illustration of CRISPR/Cas9 application in genome editing for yield improvement and stress mitigation. For CRISPR/Cas9 gene editing, a guide RNA (gRNA) is designed from a gene of interest, such as those listed in Table 2, and inserted into a binary vector. A bacterial CAS9 Nick-ase enzyme (Cas9) protein is also inserted into a binary vector. The gRNA and Cas9 expression cassettes are then used for Agrobacterium-mediate transformation for trait improvement. The fox-tail millet plant is used as a new model plant for CRISPR/Cas9 editing [241].
Figure 2.
Illustration of CRISPR/Cas9 application in genome editing for yield improvement and stress mitigation. For CRISPR/Cas9 gene editing, a guide RNA (gRNA) is designed from a gene of interest, such as those listed in Table 2, and inserted into a binary vector. A bacterial CAS9 Nick-ase enzyme (Cas9) protein is also inserted into a binary vector. The gRNA and Cas9 expression cassettes are then used for Agrobacterium-mediate transformation for trait improvement. The fox-tail millet plant is used as a new model plant for CRISPR/Cas9 editing [241].

Table 2.
List of candidate genes from related species for potential CRISPR/Cas9 editing in millets.
| Crop species | Candidate Genes for Editing in Millets | References |
|---|---|---|
| Candidate genes for drought tolerance | ||
| Oryza sativa | OsDIS1, OsiSAP7, OMTN2, OMTN3, OMTN4, OMTN6 | [199,200,201] |
| Arabidopsis thaliana | ARR1, ARR10, ARR12, AtPUB19, FtMYB10, RGLG2 | [202,203,204,205] |
| Nicotiana benthamiana | GhWRKY17 | [206] |
| Triticum aestivum | WRKY mRNA, TaWRKY146 | [207,208] |
| Eleusine coracana | Threonine dehydratase mRNA | [21] |
| Oryza sativa Japonica | OsCDPK7, NF-Y18, Arginine decarboxylase (ADC), CIPK12 | [209,210,211] |
| Zea mays | NF-YB | [212] |
| Candidate genes for heat tolerance | ||
| Triticum aestivum | TamiR159, TaHsfA6f, TaMBF1c, TaFER-5B, TaOEP16-2-5B, TaB2, TaGASR1 | [213,214,215,216,217,218,219] |
| Oryza sativa | hsp101, ZFP, OsWRKY11, OsGSK1, OsHsfA2e, mtHsp70, sHSP17.7, FAD7, SBPase, Sp17 (rice spotted leaf gene) | [220,221,222,223,224,225,226,227,228] |
| Candidate genes for dwarfism (lodging tolerance) | ||
| Oryza sativa Japonica | KO2 | [229] |
| Zea mays | GA regulatory factor-like (GRF) mRNA | [230] |
| Oryza sativa Indica | Growth-regulating factor 10 (GRF10) | [231] |
| Oryza granulata | GA20-oxidase (GA20ox2) | [232] |
| Triticum aestivum | BRI1, Rht1 | [233,234] |
| Oryza sativa | Sd-1 (used in green revl) | [235] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Numan, M.; Serba, D.D.; Ligaba-Osena, A. Alternative Strategies for Multi-Stress Tolerance and Yield Improvement in Millets. Genes 2021, 12, 739. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050739
AMA Style
Numan M, Serba DD, Ligaba-Osena A. Alternative Strategies for Multi-Stress Tolerance and Yield Improvement in Millets. Genes. 2021; 12(5):739. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050739
Chicago/Turabian StyleNuman, Muhammad, Desalegn D. Serba, and Ayalew Ligaba-Osena. 2021. "Alternative Strategies for Multi-Stress Tolerance and Yield Improvement in Millets" Genes 12, no. 5: 739. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050739
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.