
A KRT71 Loss-of-Function Variant Results in Inner Root Sheath Dysplasia and Recessive Congenital Hypotrichosis of Hereford Cattle

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Samples
2.2. Clinical and Histopathological Investigations
2.3. Genetic Investigations
2.3.1. SNP Genotyping and GWAS
2.3.2. Microsatellite Genotyping
2.3.3. Candidate Gene Analysis, Targeted Genotyping, Occurrence of the KRT71 Variant in a Global Control Cohort
3. Results
3.1. Clinical Phenotype
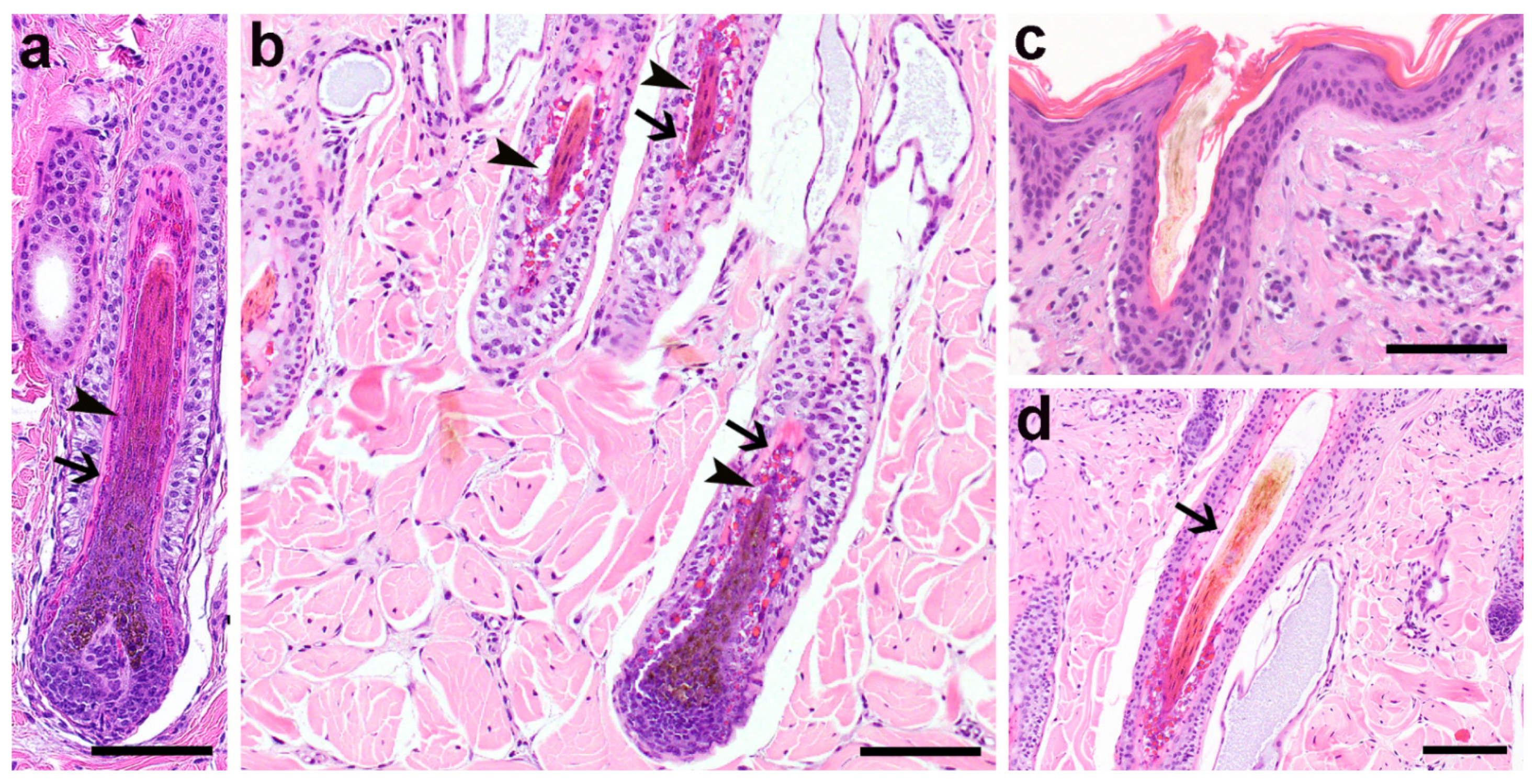
3.2. Histopathological Phenotype
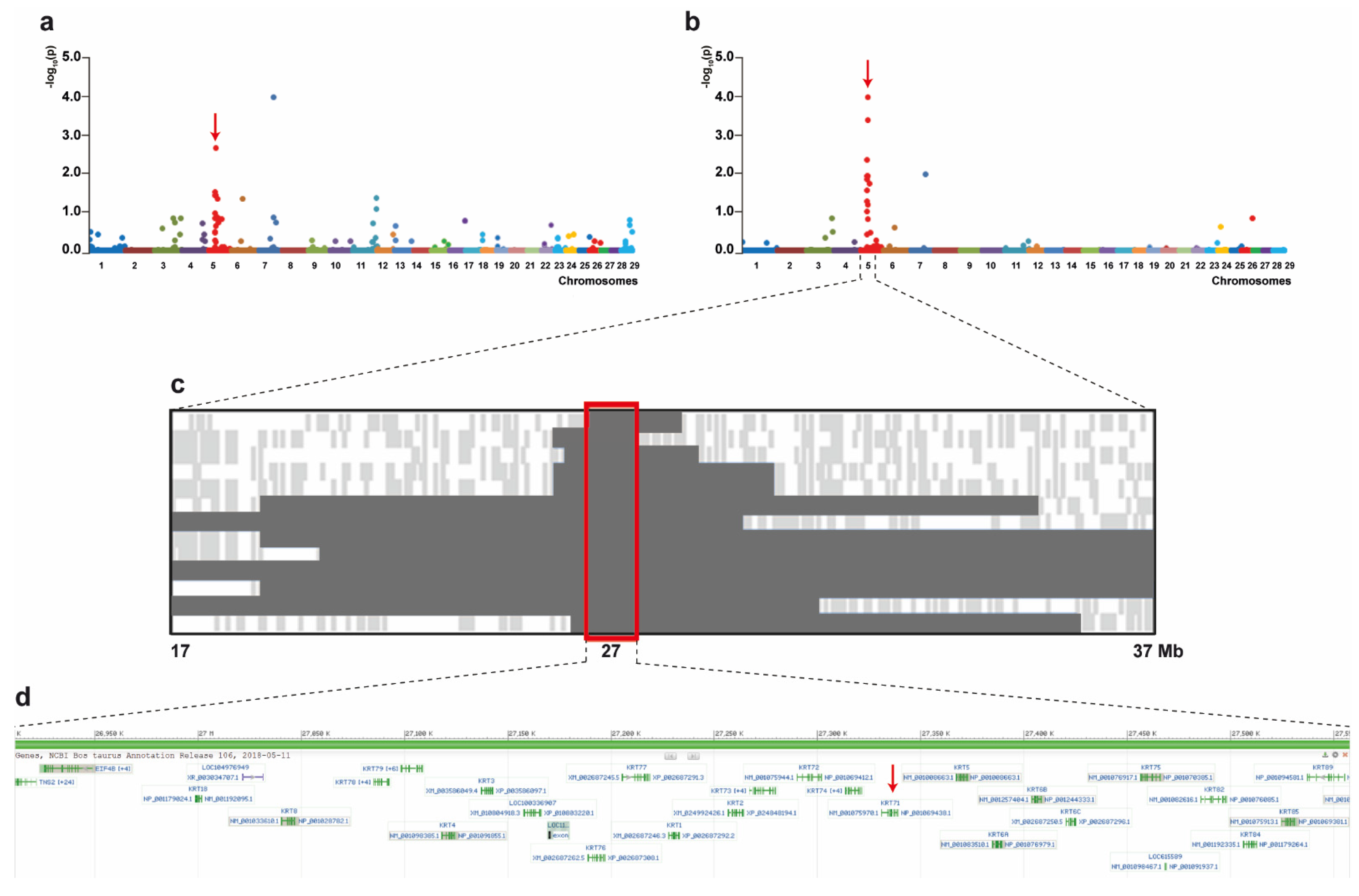
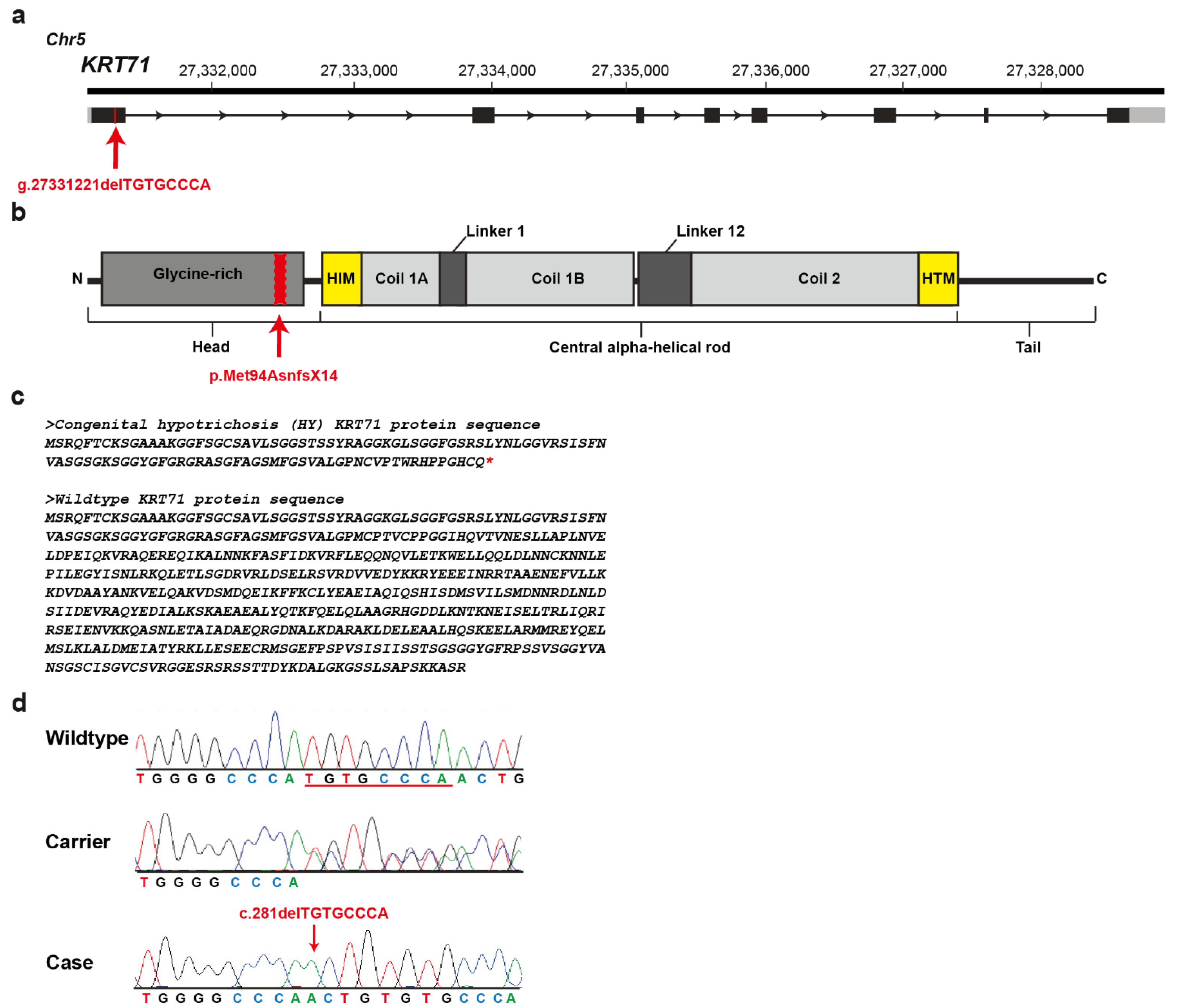
3.3. Genetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schneider, M.R.; Schmidt-Ullrich, R.; Paus, R. The Hair Follicle as a Dynamic Miniorgan. Curr. Biol. 2009, 19, R132–R142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, N.; Sawada, S.; Rogers, M.A.; Schweizer, J.; Shimomura, Y.; Tsujimoto, T.; Ito, K.; Ito, M. A novel type II cytokeratin, mK6irs, is expressed in the Huxley and Henle layers of the mouse inner root sheath. J. Investig. Dermatol. 2001, 116, 359–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porter, R.M.; Gandhi, M.; Wilson, N.J.; Wood, P.; McLean, W.H.I.; Lane, E.B. Functional analysis of keratin components in the mouse hair follicle inner root sheath. Br. J. Dermatol. 2004, 150, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Higgins, C.A.; Westgate, G.E.; Jahoda, C.A.B. Modulation in proteolytic activity is identified as a hallmark of exogen by transcriptional profiling of hair follicles. J. Investig. Dermatol. 2011, 131, 2349–2357. [Google Scholar] [CrossRef] [Green Version]
- Shimomura, Y.; Christiano, A.M. Biology and genetics of hair. Annu. Rev. Genom. Hum. Genet. 2010, 11, 109–132. [Google Scholar] [CrossRef]
- Fujimoto, A.; Farooq, M.; Fujikawa, H.; Inoue, A.; Ohyama, M.; Ehama, R.; Nakanishi, J.; Hagihara, M.; Iwabuchi, T.; Aoki, J.; et al. A missense mutation within the helix initiation motif of the keratin K71 gene underlies autosomal dominant woolly hair/hypotrichosis. J. Investig. Dermatol. 2012, 132, 2342–2349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moll, R.; Divo, M.; Langbein, L. The human keratins: Biology and pathology. Histochem. Cell Biol. 2008, 129, 705–733. [Google Scholar] [CrossRef] [Green Version]
- Coulombe, P.A.; Omary, M.B. “Hard” and “soft” principles defining the structure, function and regulation of keratin intermediate filaments. Curr. Opin. Cell Biol. 2002, 14, 110–122. [Google Scholar] [CrossRef]
- Zhang, X.; Guo, B.R.; Cai, L.Q.; Jiang, T.; Sun, L.D.; Cui, Y.; Hu, J.C.; Zhu, J.; Chen, G.; Tang, X.F.; et al. Exome sequencing identified a missense mutation of EPS8L3 in Marie Unna hereditary hypotrichosis. J. Med. Genet. 2012, 49, 727–730. [Google Scholar] [CrossRef] [Green Version]
- Pasternack, S.M.; Refke, M.; Paknia, E.; Hennies, H.C.; Franz, T.; Schäfer, N.; Fryer, A.; Van Steensel, M.; Sweeney, E.; Just, M.; et al. Mutations in SNRPE, which encodes a core protein of the spliceosome, cause autosomal-dominant hypotrichosis simplex. Am. J. Hum. Genet. 2013, 92, 81–87. [Google Scholar] [CrossRef] [Green Version]
- Betz, R.C.; Lee, Y.A.; Bygum, A.; Brandrup, F.; Bernal, A.I.; Toribio, J.; Alvarez, J.I.; Kukuk, G.M.; Ibsen, H.H.W.; Rasmussen, H.B.; et al. A gene for hypotrichosis simplex of the scalp maps to chromosome 6p21.3. Am. J. Hum. Genet. 2000, 66, 1979–1983. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.K.; Kim, E.; Baek, I.C.; Kim, B.K.; Cho, A.R.; Kim, T.Y.; Song, C.W.; Seong, J.K.; Yoon, J.B.; Stenn, K.S.; et al. Overexpression of Hr links excessive induction of Wnt signaling to Marie Unna hereditary hypotrichosis. Hum. Mol. Genet. 2009, 19, 445–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasif, N.; Naqvi, S.K.U.H.; Basit, S.; Ali, N.; Ansar, M.; Ahmad, W. Novel mutations in the keratin-74 (KRT74) gene underlie autosomal dominant woolly hair/hypotrichosis in Pakistani families. Hum. Genet. 2011, 129, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zang, D.; Jin, Y.; Wu, H.; Liu, Z.; Du, J.; Zhang, J. Mutation in ribosomal protein L21 underlies hereditary hypotrichosis simplex. Hum. Mutat. 2011, 32, 710–714. [Google Scholar] [CrossRef]
- Shimomura, Y.; Agalliu, D.; Vonica, A.; Luria, V.; Wajid, M.; Baumer, A.; Belli, S.; Petukhova, L.; Schinzel, A.; Brivanlou, A.H.; et al. APCDD1 is a novel Wnt inhibitor mutated in hereditary hypotrichosis simplex. Nature 2010, 464, 1043–1047. [Google Scholar] [CrossRef] [Green Version]
- Kazantseva, A.; Goltsov, A.; Zinchenko, R.; Grigorenko, A.P.; Abrukova, A.V.; Moliaka, Y.K.; Kirillov, A.G.; Guo, Z.; Lyle, S. Human hair growth deficiency is linked to a genetic defect in the phospholipase gene LIPH. Science 2006, 314, 2004–2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasternack, S.M.; Von Kügelgen, I.; Al Aboud, K.; Lee, Y.-A.; Rüschendorf, F.; Voss, K.; Hillmer, A.M.; Molderings, G.J.; Franz, T.; Ramirez, A.; et al. G protein-coupled receptor P2Y5 and its ligand LPA are involved in maintenance of human hair growth. Nat. Genet. 2008, 40, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, Y.; Sakamoto, F.; Kariya, N.; Matsunaga, K.; Ito, M. Mutations in the desmoglein 4 gene are associated with monilethrix-like congenital hypotrichosis. J. Investig. Dermatol. 2006, 126, 1281–1285. [Google Scholar] [CrossRef] [Green Version]
- Romano, M.T.; Tafazzoli, A.; Mattern, M.; Sivalingam, S.; Wolf, S.; Rupp, A.; Thiele, H.; Altmüller, J.; Nürnberg, P.; Ellwanger, J.; et al. Bi-allelic mutations in LSS, encoding lanosterol synthase, cause autosomal-recessive hypotrichosis simplex. Am. J. Hum. Genet. 2018, 103, 777–785. [Google Scholar] [CrossRef] [Green Version]
- Johansson, I. Congenital defects in mink. Vara Palsdjur. 1965, 36, 93–94. [Google Scholar]
- Buckley, R.M.; Gandolfi, B.; Creighton, E.K.; Pyne, C.A.; Bouhan, D.M.; Leroy, M.L.; Senter, D.A.; Gobble, J.R.; Abitbol, M.; Lyons, L.A. Werewolf, there wolf: Variants in hairless associated with hypotrichia and roaning in the lykoi cat breed. Genes 2020, 11, 682. [Google Scholar] [CrossRef] [PubMed]
- Gandolfi, B.; Outerbridge, C.A.; Beresford, L.G.; Myers, J.A.; Pimentel, M.; Alhaddad, H.; Grahn, J.C.; Grahn, R.A.; Lyons, L.A. The naked truth: Sphynx and Devon Rex cat breed mutations in KRT71. Mamm. Genome 2010, 21, 509–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, H.G.; Harris, A.; Dreger, D.L.; Davis, B.W.; Ostrander, E.A. The bald and the beautiful: Hairlessness in domestic dog breeds. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372. [Google Scholar] [CrossRef]
- Thomer, A.; Gottschalk, M.; Christmann, A.; Naccache, F.; Jung, K.; Hewicker-Trautwein, M.; Distl, O.; Metzger, J. An epistatic effect of KRT25 on SP6 is involved in curly coat in horses. Sci. Rep. 2018, 8, 6374. [Google Scholar] [CrossRef] [PubMed]
- Ratterree, M.S.; Baskin, G.B. Congenital hypotrichosis in a rhesus monkey. Lab. Anim. Sci. 1992, 42, 410–412. [Google Scholar]
- Pinter, A.J.; McLean, A.K. Hereditary hairlessness in the montane vole. J. Hered. 1970, 61, 112–118. [Google Scholar] [CrossRef]
- Swanson, H. The “hairless” gerbil: A new mutant. Lab. Anim. 1980, 14, 143–147. [Google Scholar] [CrossRef]
- Nixon, C. Hereditary hairlessness in the Syrian golden hamster. J. Hered. 1972, 63, 215–217. [Google Scholar] [CrossRef]
- Bolognia, J.L.; Murray, M.S.; Pawelek, J.M. Hairless pigmented guinea pigs: A new model for the study of mammalian pigmentation. Pigment Cell Res. 1990, 3, 150–156. [Google Scholar] [CrossRef]
- Lemus-Flores, C.; Ulloa-Arvizu, R.; Ramos-Kuri, M.; Estrada, F.J.; Alonso, R.A. Genetic analysis of Mexican hairless pig populations. J. Anim. Sci. 2001, 79, 3021–3026. [Google Scholar] [CrossRef] [Green Version]
- Finocchiaro, R.; Portolano, B.; Damiani, G.; Caroli, A.; Budelli, E.; Bolla, P.; Pagnacco, G. The hairless (hr) gene is involved in the congenital hypotrichosis of Valle del Belice sheep. Genet. Sel. Evol. 2003, 35, S147–S156. [Google Scholar] [CrossRef] [PubMed]
- Murgiano, L.; Shirokova, V.; Welle, M.M.; Jagannathan, V.; Plattet, P.; Oevermann, A.; Pienkowska-Schelling, A.; Gallo, D.; Gentile, A.; Mikkola, M.; et al. Hairless streaks in cattle implicate TSR2 in early hair follicle formation. PLoS Genet. 2015, 11, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craft, W.A.; Blizzard, W.L. The inheritance of semi-hairless ness in cattle. J. Hered. 1934, 25, 385–390. [Google Scholar] [CrossRef]
- Bracho, G.A.; Beeman, K.; Johnson, J.L.; Leipold, H.W. Further studies of congenital hypotrichosis in Hereford cattle. Zent. Vet. 1984, 31, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Olson, T.A.; Hargrove, D.D.; Leipold, H.W. Occurrence of Hypotrichosis in Polled Hereford Cattle. Bov. Pract. 1985, 20, 4–8. [Google Scholar] [CrossRef]
- Jayasekara, M.U.; Leipold, H.W.; Cook, J.E. Pathological changes in congenital hypotrichosis in Hereford cattle. Zentralbl. Veterinarmed. A 1979, 26, 744–753. [Google Scholar] [CrossRef]
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef] [Green Version]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Temnykh, S.; DeClerck, G.; Lukashova, A.; Lipovich, L.; Cartinhour, S.; McCouch, S. Computational and experimental analysis of microsatellites in rice (Oryza sativa L.): Frequency, length variation, transposon associations, and genetic marker potential. Genome Res. 2001, 11, 1441–1452. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.; Bridges, S.; Magbanua, Z.V.; Peterson, D.G. Computational Approaches and Tools Used in Identification of Dispersed Repetitive DNA Sequences. Trop. Plant Biol. 2008, 1, 85–96. [Google Scholar] [CrossRef]
- Boutin-Ganache, I.; Raposo, M.; Raymond, M.; Deschepper, C.F. M13-tailed primers improve the readability and usability of microsatellite analyses performed with two different allele-sizing methods. Biotechniques 2001, 31, 24–28. [Google Scholar] [CrossRef] [Green Version]
- Wheelan, S.J.; Church, D.M.; Ostell, J.M. Spidey: A tool for mRNA-to-genomic alignments. Genome Res. 2001, 11, 1952–1957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, B.J.; Daetwyler, H.D. 1000 Bull Genomes Project to Map Simple and Complex Genetic Traits in Cattle: Applications and Outcomes. Annu. Rev. Anim. Biosci. 2019, 7, 89–102. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, D.; Häfliger, I.M.; Leuthard, F.; Schumaker, B.; Steadman, L.; Murphy, B.; Drögemüller, C.; Leeb, T. X-linked hypohidrotic ectodermal dysplasia in crossbred beef cattle due to a large deletion in EDA. Animals 2021, 11, 657. [Google Scholar] [CrossRef] [PubMed]
- Snider, N.T.; Omary, M.B. Post-translational modifications of intermediate filament proteins: Mechanisms and functions. Nat. Rev. Mol. Cell Biol. 2014, 15, 163–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arin, M.J. The molecular basis of human keratin disorders. Hum. Genet. 2009, 125, 355–373. [Google Scholar] [CrossRef]
- Langbein, L.; Rogers, M.A.; Praetzel, S.; Winter, H.; Schweizer, J. K6irs1, K6irs2, K6irs3, and K6irs4 represent the inner-root-sheath-specific type II epithelial keratins of the human hair follicle. J. Investig. Dermatol. 2003, 120, 512–522. [Google Scholar] [CrossRef] [Green Version]
- Genovese, D.W.; Johnson, T.L.; Lamb, K.E.; Gram, W.D. Histological and dermatoscopic description of sphynx cat skin. Vet. Dermatol. 2014, 25, 523-e90. [Google Scholar] [CrossRef]
- Fairfield, H.; Srivastava, A.; Ananda, G.; Liu, R.; Kircher, M.; Lakshminarayana, A.; Harris, B.S.; Karst, S.Y.; Dionne, L.A.; Kane, C.C.; et al. Exome sequencing reveals pathogenic mutations in 91 strains of mice with Mendelian disorders. Genome Res. 2015, 25, 948–957. [Google Scholar] [CrossRef] [Green Version]
- Runkel, F.; Klaften, M.; Koch, K.; Böhnert, V.; Büssow, H.; Fuchs, H.; Franz, T.; De Angelis, M.H. Morphologic and molecular characterization of two novel Krt71 (Krt2-6g) mutations: Krt71rco12 and Krt71rco13. Mamm. Genome 2006, 17, 1172–1182. [Google Scholar] [CrossRef]
- Peters, T.; Sedlmeier, R.; Büssow, H.; Runkel, F.; Lüers, G.H.; Korthaus, D.; Fuchs, H.; Hrabé De Angelis, M.; Stumm, G.; Russ, A.P.; et al. Alopecia in a novel mouse model RCO3 is caused by mK6irs1 deficiency. J. Investig. Dermatol. 2003, 121, 674–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuca, T.; Marron, B.M.; Jacinto, J.G.P.; Paris, J.M.; Gerspach, C.; Beever, J.E.; Drögemüller, C. A nonsense variant in hephaestin like 1 (HEPHL1) is responsible for congenital hypotrichosis in Belted Galloway cattle. Genes. 2021, 12, 643. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| wt/wt | wt/del | del/del | |
|---|---|---|---|
| HY-affected calves | |||
| Swiss cases b | 2 | ||
| US cases | 4 a | 13 | |
| Obligate carriers c | |||
| Swiss b | 2 | ||
| US | 22 | ||
| Unrelated normal Hereford cattle | |||
| Swiss b | 21 | 8 | |
| US | 197 | 9 | |
| Global cohort included in the 1000 Bull Genomes Project | 117 | ||
| Normal control cattle from various different breeds | 3993 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jacinto, J.G.P.; Markey, A.D.; Veiga, I.M.B.; Paris, J.M.; Welle, M.; Beever, J.E.; Drögemüller, C. A KRT71 Loss-of-Function Variant Results in Inner Root Sheath Dysplasia and Recessive Congenital Hypotrichosis of Hereford Cattle. Genes 2021, 12, 1038. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12071038
Jacinto JGP, Markey AD, Veiga IMB, Paris JM, Welle M, Beever JE, Drögemüller C. A KRT71 Loss-of-Function Variant Results in Inner Root Sheath Dysplasia and Recessive Congenital Hypotrichosis of Hereford Cattle. Genes. 2021; 12(7):1038. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12071038
Chicago/Turabian StyleJacinto, Joana G. P., Alysta D. Markey, Inês M. B. Veiga, Julia M. Paris, Monika Welle, Jonathan E. Beever, and Cord Drögemüller. 2021. "A KRT71 Loss-of-Function Variant Results in Inner Root Sheath Dysplasia and Recessive Congenital Hypotrichosis of Hereford Cattle" Genes 12, no. 7: 1038. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12071038