1. Introduction
Osteoarthritis (OA) is the most prevalent whole-joint disorder that exists in the elderly population. Although, OA pathology is not fully understood, it is a multifactorial disorder marked by several cellular and molecular changes, such as an imbalance between cartilage anabolism and catabolism, chondrocytes hypertrophy and death, infiltration of macrophages and activation of immune responses and synovial inflammation and hypertrophy. All together, these pathological changes lead to a gradual loss of articular cartilage. Clinical symptoms, such as joint stiffness, warmth, pain, and joint effusion indicate a presence of inflammation in OA joints [
1]. Evolving comprehension of the pathology revealed that OA inflammation is particularly of a low-grade and fundamentally differs from inflammation in rheumatoid arthritis (RA). Scientific studies reported that pro-inflammatory markers in synovial fluid (SF) and blood from OA patients were modest in comparison with RA individuals but higher than healthy controls [
2]. Arthroscopy and histological investigation of OA synovium revealed a sub-acute type of inflammation, which is less pronounced than in RA synovium [
3]. Further, immune cells infiltration (like macrophages and T cells) in OA synovium is much lower, when compared to RA synovium [
4]. The gathered evidence suggests that mononuclear infiltration and over expression of inflammatory mediators in synovium are seen in early OA and predate radiographic damage in OA [
5]. Immunopathological studies in OA revealed that inflammation is primarily responsible for triggering innate immune responses that further lead to activation of metalloproteinases (MMPs), in part [
4]. Predisposition of obese and diabetic individuals to OA also suggests that systemic inflammatory factors may play an important role in the pathology [
6]. Thus, a strong involvement of inflammatory component is increasingly recognized in OA.
Considering an evolving role of inflammation in OA, a systemic blocking of conventional inflammatory pathways (such as anti-TNF or anti-IL1β agents) was thought to be effective; however, it was found that there was no benefit in generalized OA and minimally effective in erosive OA [
7,
8]. This underlines a fact that inflammatory mechanism in OA is different and not fully known. Allied to this, further illumination is also needed on the causal relationship between inflammation and cartilage degradation process to make anti-inflammatory therapies more effective in OA management. Synovial membrane and synovial fluid (SF) are the major contributors of inflammation. Key cytokines including interleukin-1β (IL-1β), tumor necrosis factor-α (TNF-α), IL-15, IL-6, IL-17 are produced by synoviocytes and macrophages, the membrane cells. Most of these cytokines including IL-1β, TNF-α and IL-15 are the chief regulators of MMPs, which are primarily responsible for cartilage loss in OA [
9]. Moreover, synovium is known to translate systemic inflammation into OA specific pathology. On the other hand, SF is a biochemical channel that transmit and receive molecular signal in the joint cavity. Therefore, focusing on synovium-SF axis can unravel inflammatory network in OA. Recent research studies showed that cartilage loss is reversible, if intervened at the early stages of the disease [
10]. Accounting non-linear progressive cartilage loss and chronic nature of OA a need is imposed to understand inflammatory milieu and MMP status at different stages of OA for the development of early-stage therapies or personalized disease management.
In this communication, we present a grade-wise pattern of key inflammatory agents and MMPs, estimated in synovium, collected from different grades of OA patients. Further, OA specific MMPs (MMP-1, MMP-13, MMP-2 and MMP-9) levels were assessed in OA SFs to find out their activity and abundance at various stages during the disease progression. To understand effect of inflammatory microenvironment on MMPs modulation, SF challenge test was performed. In this test, OA SFs of different grades were used to induce inflammation in human synoviocytes and monocytes; after the SF treatment MMPs modulation was recorded. Finally, a correlation matrix of cytokines and MMPs was developed to find out functional association between inflammation and cartilage degeneration in OA. In all, the outcomes of this multilevel analysis of inflammatory factors and MMPs in OA clinical samples are expected to enlighten the existing knowledge about the perpetuating inflammatory mediators and their effects on MMPs. Improved understanding will ultimately boost the efforts to develop the disease stage-wise or personalized management for OA.
2. Materials and Methods
2.1. Ethical Statement
The methods of this research work abide by the Declaration of Helsinki and the protocols were approved by the Institutional Ethics Committee (BVDU/MC/01; Date of approval—15 June 2015). All the study patients signed an informed written consent before participation.
2.2. Collection of Clinical Samples
2.2.1. Synovial Biopsies
Synovial biopsies (
n = 26; KL grade-I = 6; KL grade-II = 6; KL grade-III = 6; KL grade-IV = 8) were collected from OA patients, who had undergone knee arthroscopy or knee replacement surgery. OA diagnosis was performed by experienced rheumatologist based on clinical symptoms and radiographs. OA grading was determined by commonly used radiographic Kellgren–Lawrence (KL) score. According to this score system, grade-I—doubtful OA with presence of minor osteophytes of doubtful importance, grade-II—minimal OA, with definite osteophytes but unimpaired joint space, grade-III—moderate OA, with osteophytes and moderate diminution of joint space whereas grade -IV—severe OA, with greatly impaired joint space and sclerosis of subchondral bone [
11].
The collected biopsies were stored in liquid nitrogen before they were used for inflammatory biomarkers and MMPs expression analysis.
2.2.2. SF Sample Collection
SF samples (
n = 40; KL grade-I = 9; KL grade-II = 14; KL grade-III = 10, KL grade-IV = 7) were collected from different grades of OA patients by knee arthrocentesis as described [
12]. The fluid collection process was performed in minor operation theater under strict aseptic conditions to collect SFs from early grade OA patients. The fluids from advanced grade OA were collected at the time of knee replacement surgery. The collected samples were stored in −80 °C and were diluted in 1X PBS in the ratio 1:10 before they were used for the planned assays.
2.2.3. Estimation of Pro-Inflammatory Markers and MMPs in the Synovial Biopsies
IL-1β, IL-15, prostaglandin-E2 (PGE2), nerve growth factor (NGF), MMP-1, MMP-13, and TIMP-1 expression pattern was determined in the collected synovial biopsies. First, RNA isolation was performed in the collected tissues, using PureLink RNA mini kit (Invitrogen, CA, USA) as per the manufacturer’s instructions. The tissues were homogenized in 1 mL Trizol reagent for 5 min. Later, 200 µL chloroform was added in the same tube, vortexed for 15 s and kept for 3 min at RT. This step was followed by centrifugation at 12,000 rpm for 15 min at 4 °C. 400 µL supernatant was transferred in a fresh tube and equal volume of ethanol was added. This mixture was later transferred in batches to a spin cartridge and was centrifuged for 30 s at RT. Flow through was discarded and this step was repeated until the whole sample got processed. The membrane bound RNA was then subjected to thorough washing; 700 µL of Wash Buffer-I was added to spin cartridge and was centrifuged at 12,000× g at RT for 30 s. The spin cartridge was then shifted to another collection tube and washed twice by adding 500 µL of wash buffer, centrifuged at 12,000× g at RT for 30 s and the flow through was discarded. Further, the spin cartridge was centrifuged at 12,000× g at RT for 1 min to dry the membrane bound RNA; the collection tube was discarded. For the elution of the bound RNA, the spin cartridge was transferred to a fresh recovery tube and 30 µL RNAase free water was added. After a brief incubation for 2 min at RT, the spin cartridge was centrifuged at 12,000× g at RT for one minute. Quality of eluted RNA was determined by using denaturing agarose gel electrophoresis and the quantification was done by measuring absorbance at 260 nm. This RNA was later used for cDNA synthesis, as explained later in this section.
2.2.4. IL-1β Estimation in SF Samples
IL-1β estimation in the collected OA SFs were performed using commercially available ELISA kit (Abnova, Walnut, CA, USA) and following the manufacturer’s instructions. The SF samples included for IL-1β levels determination were thirty-three (KL grade-I = 8; KL grade-II = 7; KL grade-III = 6; KL grade-IV = 12). The optical density was read at 450 nm using ELISA microplate reader (Biorad, Hercules, CA, USA).
2.2.5. MMP Levels Estimation in SF Samples
Abundance of MMP-1, MMP-2, MMP-9, MMP-13 and TIMP-1 in their protein form was estimated in the collected SFs using commercially available ELISA kits (Cloud-Clone Corp, Houston, TX, USA) and as per the manufacturer’s instructions. In brief, 100 µL of standard diluent was added in antibody coated well as a blank and prepared 7 concentrations for standards; then diluted SF samples were added in antibody coated wells and incubated for 1 h at RT. In case of TIMP-1, the sample incubation period was 2 h at RT. After the incubation, the samples were removed from wells and 100 µL of detection reagent-A was added in each well. The plate was covered with a plate sealer and again incubated for 1 h at 37 °C. After incubation, each well was washed thrice with 350 µL wash buffer. After the washing procedure, 100 µL of detection reagent-B was added in each well and incubated for 30 min at 37 °C. On completion of incubation, the washing procedure was performed 5 times using washing buffer. Later, 90 µL of substrate solution was added in to each well and was incubated in dark for 10–20 min at RT. Finally, 50 µL stop solution was added and read at 450 nm immediately. MMP and TIMP levels were calculated by using standard graph and multiplied by dilution factor to get the final value.
2.3. Collagenase Activity of MMPs, Estimated in SF Samples Using FALGPA Reagent
Collagenase activity of MMPs was determined using FALGPA, a synthetic substrate, which has resemblance to the primary structure of collagen and is hydrolyzed by all known collagenases. For FALGPA assay, 200 µL of FALGPA reagent was added into a mixture of 10 µL SF sample and 90 µL of Tris buffer. This mixture was further incubated for 15 min at 25 °C. Post-incubation, the absorbance was measured at 340 nm using Perkin Elmer UV/VIS spectrophotometer. The same procedure was repeated in another tube with 100 µL of buffered collagenase enzyme, which was used as a control. FALGPA unit is hydrolysis of 1.0 micromole of FALGPA per min at 25 °C pH 8.8 in a presence of calcium ions.
2.4. SF Challenge Test on Human Synoviocytes (SW982) and Monocytes (THP1)
This experiment was planned to assess MMP specific response generated by inflamed SW982 and THP1 cells. Both the cell lines were obtained from National Centre for Cell Science (NCCS), Pune. SW982 cells were grown in culture media composed of dulbecco’s modified eagle medium (DMEM) + 10% fetal bovine serum (FBS) + 2 mmol/L L-glutamine + 100 U/mL penicillin + 100 µg/mL streptomycin. On the other hand, THP1 cells were maintained in Roswell Park Memorial Institute (RPMI) 1640 medium (containing10% FBS + 100 U/mL penicillin + 100 µg/mL streptomycin). Both the cells were maintained and grown at 95% relative humidity and 5% CO2 at 37 °C.
During SF challenge test, SW982 were treated with different grades of OA SFs (5% of culture media) for 72 h, while THP1 cells were treated with 10% SF (of culture media) for 48 h to induce inflammation. Successful induction of inflammation was confirmed by estimation of nitric oxide (NO) and IL-1β. NO release was estimated from the cell culture medium, using Griess reaction method as described [
13]. IL-1β expression in the SF treated SW982 was tested by qPCR and the values were normalized against β-actin gene. SF challenge test was developed and standardized by our research group and was successfully used to study OA specific responses [
14].
2.5. cDNA Synthesis
After the SF treatment, both the cells were harvested for RNA isolation. RNA isolation was done using PureLink RNA mini kit (Invitrogen, Carlsbad, CA, USA) as per the manufacturer’s instructions. The cells without SF treatment were used as a control. Isolated RNA was used for cDNA synthesis, which was done by using a high-capacity cDNA reverse transcription kit (Invitrogen, CA, USA). As per the manufacturer’s instructions, for each 20 µL reaction, 2 µg of isolated RNA was used. Depending on the number of reactions, the reagents’ volumes were calculated. For preparation of one reaction of 2X RT master mix, 2.0 µL of 10X RT Buffer, 2.0 µL of 10X RT Random primers, 0.8 µL of 25X dNTP mix (100 mM), 1 µL of MultiScribe™ Reverse Transcriptase and 4.2 µL of nuclease-free water was used. The reaction mixture was kept on ice and mixed gently. In total, 10 µL of RNA was added into 2X RT master-mix to prepare cDNA RT reaction in a PCR tube and was mixed gently by pipetting up and down. Finally, a PCR tube was centrifuged to spin down all the contents and to remove air bubbles if any. The tube was then placed in a thermal cycler that was programmed according to manufacturer’s instructions. The reverse transcriptase run cycle was performed as described. On the completion of one cycle, the obtained cDNA was stored at −80 °C for quantitative polymerase chain reaction (qPCR) analysis.
2.6. qPCR Analysis
qPCR analysis was performed using Applied Biosystems Step One Real Time PCR System as per the manufacturer’s instructions (Applied Biosystems, Foster City, CA, USA). TaqMan gene expression assays (Applied Biosystems, Foster City, CA, USA) analyzed in the SF treated SW982 and THP1 cells were, MMP1, MMP-13. Additionally, VEGF-1 expression was determined in the SF treated THP1 cells. mRNA levels of the selected genes were calculated to the amount of β-actin using Step One Software version 2.2.2.
All the reagents, chemicals and cell/tissue culture media used in the experimental work were of analytical grade and were purchased from Sigma-Aldrich (St. Louis, MO, USA). Plastic-ware was procured from BD Biosciences and Axygen Scientific Inc., Union City, CA, USA.
2.7.Statistical Analysis
The clinical sample evaluation for the inflammatory factors and MMP estimation and in vitro studies were performed in triplicates and the data was presented as mean ± SD. Inter-grade statistical significance was calculated by One Way ANOVA followed by a Kruskal–Wallis test and Tukey’s test of significance using R software version 3.3.0.
4. Discussion
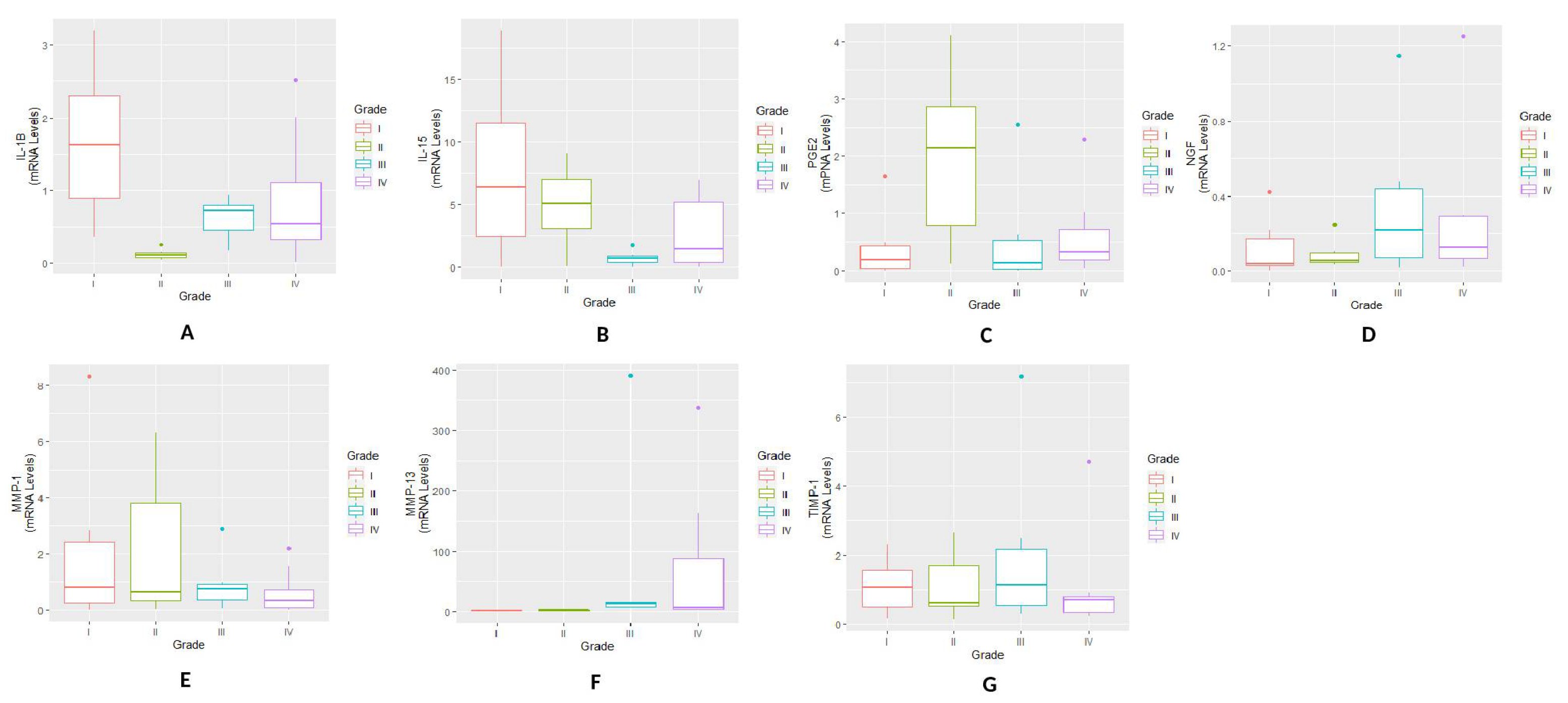
The present study investigates various molecular and cellular events along the synovium-synovial fluid axis, as it plays the unique significant biological functions of nurturing, maintaining and protecting cartilage. This axis is also known for its barrier shielding function for cartilage from various biochemical insults and obligating a protective role. All the systemic signals are received by synovium and relayed by SF and hence are expected to hold early to advance signs of the disease pathology [
9]. Gene expressions in synovium biopsies in this work formed a purposive approach to study the variation pattern of inflammatory mediators and MMPs during natural OA progression. A definite expression trend was noticed in the selected markers. Peak expressions of IL-1β and IL-15 in KL grade-I synovium indicated that the regulators of the inflammation were expressed higher at early stage that provides a biochemical trigger even before appearance of undoubted clinical signatures of OA. The highest PGE2 was found in KL grade-II samples, immediately in the next stage of peak IL-1β, the main inducer of PGE2. NGF showed comparable expressions in KL grades-I, III and IV. In a grade-wise comparison, IL-1β and IL-15 expression showed a marked significance in KL grade-I synovium. However, the other inflammatory markers and MMPs did not show any significant difference, possibly due to a smaller number of samples. Taken together, the expression trend of these pro-inflammatory markers explained persistent synovitis in all stages of OA, unlike a common claim that synovial inflammation is prevalent at early and advanced stages. Further, these results indicated that each stage of OA was marked by a presence of a particular inflammatory factor. The individual role of these mediators has been extensively studied in OA and their increased levels has been recorded in OA affected clinical samples [
15,
16,
17,
18]. Combined effect of elevated IL-β, IL-15 and PGE2 in early stages of OA provided the necessary impetus for accelerated cartilage loss at later stages of the disease as evident by relatively higher expression of MMPs in advanced staged synovial biopsies (KL grade-III and IV). Of note, excessive levels of IL-β, IL-15 and PGE2 are associated with impaired bone and cartilage remodeling. Particularly IL-1β and IL-15 are known to cause a destructive effect on articular cartilage by up-regulating MMPs [
16,
19]. PGE2 and NGF, both are pro-angiogenic factors and their high levels contribute to synovial inflammation and hyperplasia. Additionally, NGF has a strong connection with transmission of pain in OA [
18,
20]. On this background, comparable levels of TIMP−1 in all grade synovium membranes can be a natural countering against increasing MMP activity to limit the cartilage-loss. Besides, the data presented here give a context of the progressive grades of OA, which has not been adequately represented in the published literature so far.
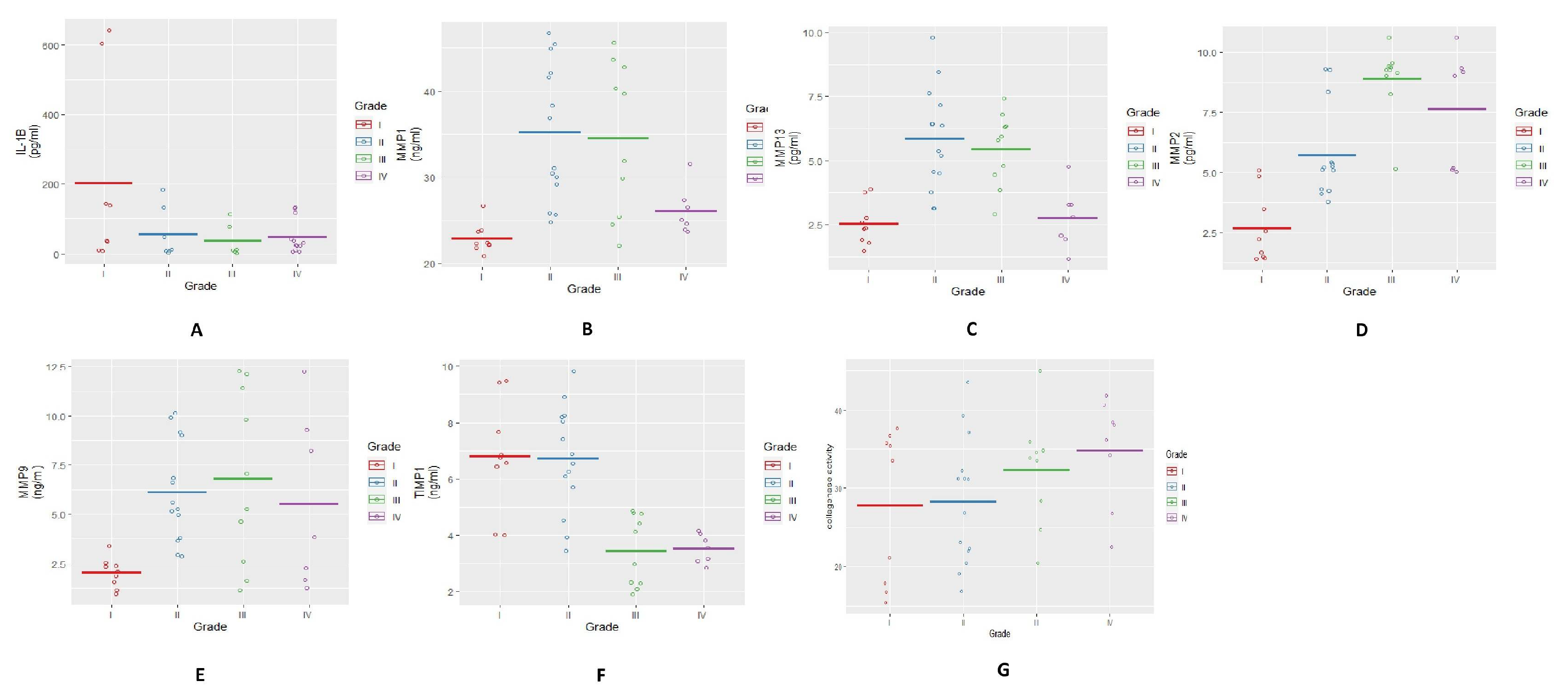
FALGPA and ELISA assays represented the collagenolytic MMPs activity and their protein abundance in SF, respectively. All MMP protein levels were noted higher in moderate grade SFs (KL grade-II and III) whereas, their collagenolytic activity as measured through FALGPA assay was found steadily increased in SF with advanced grades. TIMP is a natural inhibitor secreted by tissues and its abundance is considered to be in repulsion with the concentration of MMPs. In our observation here, TIMP abundance clearly decreased in SF of KL grade-III and IV with concomitant increase in MMPs (
Figure 2F,G). In this study a dichotomy was detected in abundance of TIMP−1 in SF and its expression in synovium. Here, a significant reduction of TIMP−1 was noted in late OA SFs (KL grade-III and IV), whereas its expression was consistent in the synovium from all the grades. Although, several explanations for this finding are possible, it is likely that TIMP−1 concentration in SF is not contributed only by the synovium. This may be recorded as limitation of present data that other tissues and other TIMPs were not studied, which certainly contribute to ECM turnover [
21]. Furthermore, TIMPs are not the only inhibitors for MMPs; Tchetverikov et al. (2005) showed α2-macroglobulin played a leading role in scavenging activated MMPs. However, the increased proMMP/TIMP ratio in favor of MMPs in their study, correlate with our observation. It may be interesting to note the similarities among progressive OA, knee injury and inflammatory arthritis [
22]. A meta-analysis covering a total of 1408 studies designed to study relationship between expression of MMPs and pathogenesis of OA, revealed higher expression of MMP-1, MMP-2 and MMP-9 in OA, as well as ethnic differences in their expression except for MMP-9 [
23]. Further, FALGPA and ELISA assay outcomes were also corresponded with our previous results of glycosaminoglycan (GAG) estimation in OA SFs of different grades. Considering GAG as a direct product of MMP action on cartilage, the highest GAG in KL grade-II and III SFs was a clear indication of the maximum activity of MMPs at these grades [
24]. Sachdeva et al. [
25] reported similar observation on the ethnically similar population as in the present study, wherein MMP-9 and MMP-13 were studied in SF of OA patients. The expression of both the MMPs was reported high in KL grade-II and KL grade-III SFs and subsides in KL grade-IV.
Higher levels of pro-inflammatory factors in SF have been reported by many workers, including some proteomic study wherein proteins and peptides were detected, which may act as trigger for cellular and tissue inflammation. In a study on 100 subjects, Saetan et al. (2014) reported higher VEGF expression in synovial tissue and corresponding increased levels in SF and serum [
26]. However, these SF proteins need to be tested for their potential to induce biologically relevant inflammation in joint tissue individually as well as collectively to elucidate the underlining pathological inflammatory mechanism. The novelty of the present data is the SF challenge test that was designed to study the dynamics of modulation in inflammatory microenvironment at joint in terms of classical inflammatory mediators and MMPs expression. These experiments were based on our previous learning that OA SF holds inflammatory milieu of various cytokines and chemokines and can be used to study the disease specific responses on cell lines [
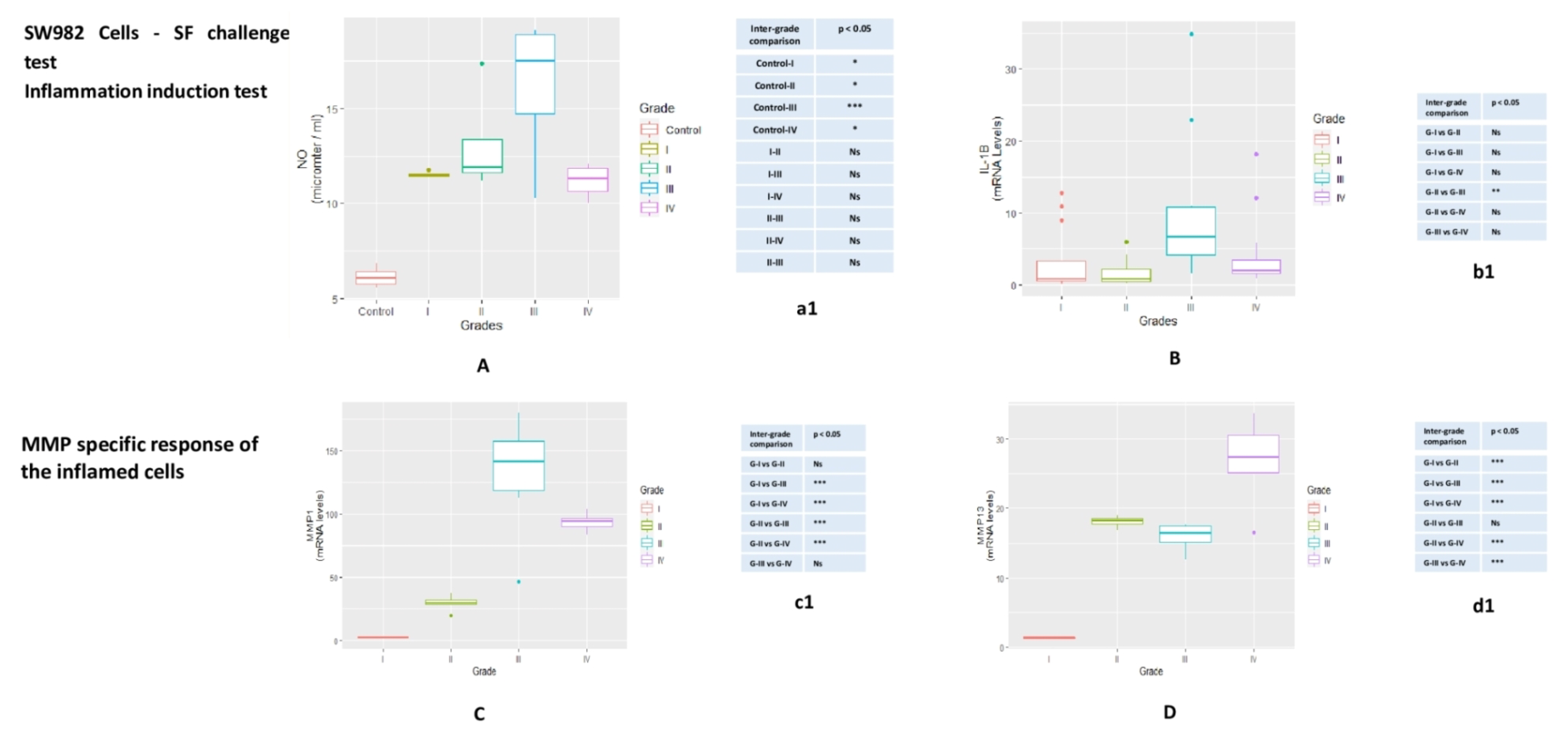
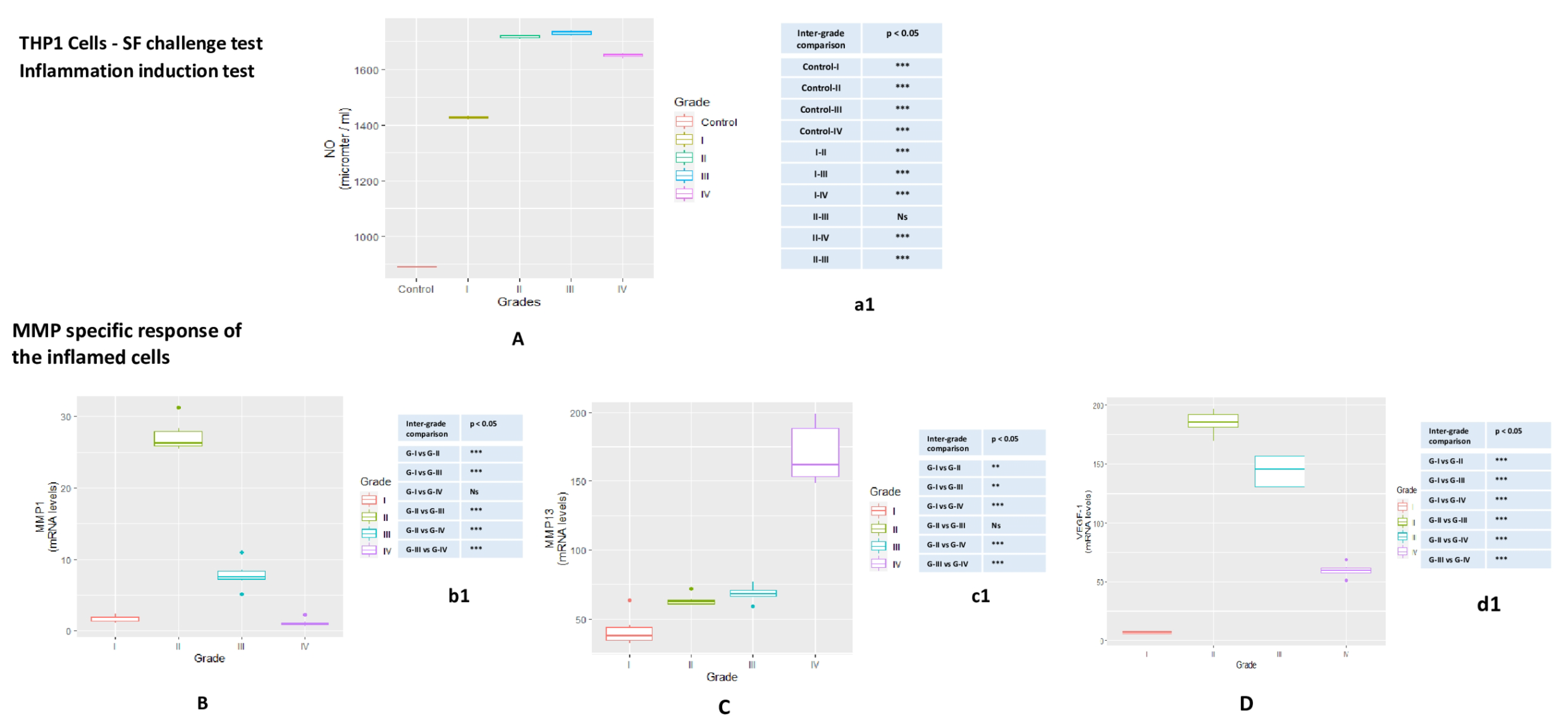
14]. Most of the etiopathology of OA spins around the synovium-SF-cartilage axis, wherein SF occupies the central position. The SF challenge test was carried out with the SF obtained from OA patients of different grades and with two different types of cells. The advantage of this biological assay was its physiological relevance; the cells are subjected to their natural inflammatory micro-environment as in OA joints and hence are expected to mimic their response as in vivo conditions. The SF challenge test was performed using human monocytes and synoviocytes, the two major cell types of synovium, which are involved in the synovial inflammation. Successful induction of inflammation in the SF treated cells was confirmed by estimation of NO and mRNA levels of IL-1β (
Figure 3A,B and
Figure 4A). NO is a known inflammatory marker produced by the cells under stress. In the present work, SF treatment in both the cell lines showed a significant NO release as compared to untreated cells (
Figure 3, Table a1 and
Figure 4a1). In inflamed joints, this unmitigated NO stimulates excessive MMP production and inhibits synthesis of collagen and proteoglycans [
27]. Interestingly, both the cell lines showed up-regulation of MMP-1 and MMP-13 in a similar manner; this means that moderate grade OA SFs (KL grade-II and III) caused the highest up-regulation of MMP-1, while KL grade-IV SF treatment was responsible for the maximum up-regulation of MMP-13. Additionally, SF treatment on THP1 also up-regulated VEGF−1, a pro-angiogenic factor. A surprising data published by Hoff et al. (2013) showed that OA-SF is more potent over to RA-SF in inducing VEGF in primary chondrocytes indicating that SF contains pro-inflammatory mediators capable of inducing inflammatory changes in synoviocytes on one hand and chondrocytes on the other [
28].
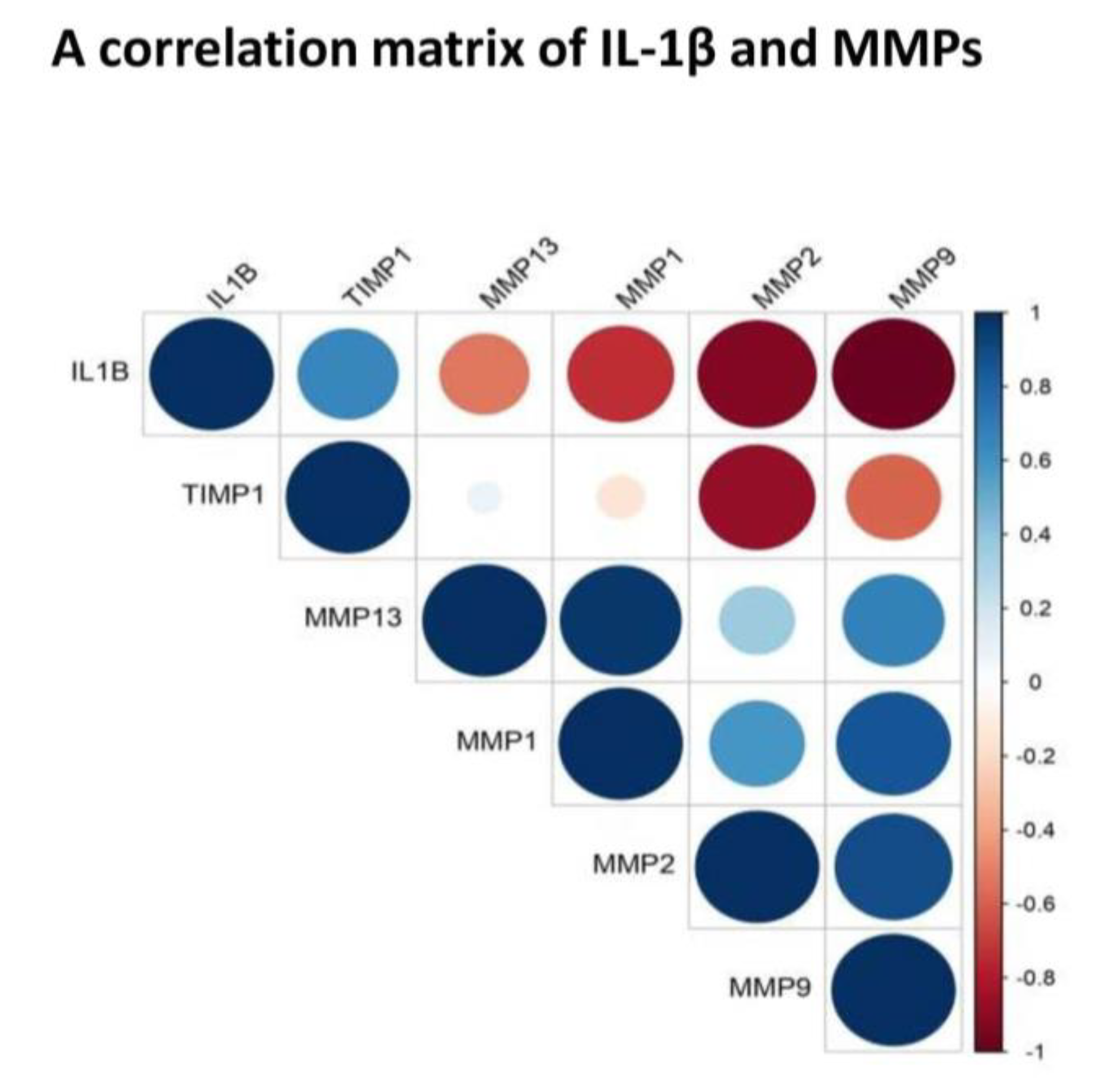
To find out a functional correlation between inflammation and cartilage-loss in OA, we plotted a correlation matrix of IL-1β and MMPs using their values in SF (
Figure 5). Of note, SF represents a “collective secretome” of all the cells in the joint that include, synovium, cartilage, meniscus, and immune cells and thus, represents a true cross-section of metabolic status at the given KL grade of the joint. Moreover, in this study, SF values yielded a consistent pattern of correlation matrix, which matched with FALGPA and SF challenge test. This correlation matrix revealed a negative correlation between IL-1β and MMPs, particularly, with MMP-1, MMP-2 and MMP-9, suggesting a clear special difference in their expression. Otherwise, IL-1β is known to induce MMPs secretion through multiple pathways for example, MMP-1 and MMP-13 are stimulated via MAPK/ERK, NF-κB and Wnt−5A signaling [
29], whereas up-regulation of Wnt−5A additionally induce MMP-9 as shown in rabbit chondrocytes [
30]. In the recent work, IL-1β was also shown to induce MMPs via up-regulation of Notch1 and NICD in chondrocytes [
31]. The studies also revealed that effects of IL-1β on synoviocytes and chondrocytes are similar in terms of strong induction of MMP-2 and MMP-13 via activation of RUNX−2 and Wnt/β-catenin as well as MAPK and NF-κB pathways [
32,
33]. This research evidence clearly exhibited a positive biological correlation between IL-1β and MMPs although, it differentiated in time as revealed by the present data. IL-1β along with other cytokines is also known to be involved in activation, differentiation and proliferation of effector cells, which ultimately produce MMPs [
34]. Overwhelming evidence from various experiments elsewhere and in the present study suggest that cartilage-loss in OA is inflammation driven. We also observed a strong positive correlation between MMP-1 and MMP-13 as well as between MMP-2 and MMP-9 (
Figure 5). This was quite expected as both the MMPs belong to the collagenase sub-class of the family and are about 50%−55% identical in sequence. Of note, type-II collagen is a poor substrate for MMP-1, while MMP-13 is five to ten times more potent against it. However, different MMPs overtake each other’s role with comparable efficiency to degrade type-II collagen, is the main reason why MMPs inhibition is not a successful therapeutic strategy.
The strength of this study includes multilevel analysis of OA tissues (synovium and SF) of across the grades of the disease and rational validation of the outcomes using physiologically relevant in vitro model. On concluding remarks, the etiology of OA is complex and multifactorial and involves trauma, age, systemic comorbidities, and heredity. Typical to the complex non-communicable diseases, rigorous analysis of the structure and the complex dynamics of molecular networks across the various tissues, remains as a prerequisite to give a thorough understanding of the disease, which in the case of OA is still a major impediment in the discovery of potential druggable targets, and to design effective therapy. Further, there is a large person-to-person variation as seen elsewhere and in present analysis, indicating perhaps “one for all” medicine may not be a yielding strategy in OA. A personalized approach built through the combination of inhibition of degrading enzymes, reducing inflammation, immuno-modulation, limiting angiogenesis and synovitis and management of pain appears to be the plausible approach in the management of OA.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




