
Brain Anatomical Mediators of GRIN2B Gene Association with Attention/Hyperactivity Problems: An Integrated Genetic-Neuroimaging Study

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Longitudinal Study Protocol
2.2. Subjects
2.3. Behavioral and Clinical Measures
Child Behavior Checklist (CBCL/6-18)
2.4. Genotyping
2.5. MRI Data Acquisition
2.6. MRI Data Processing
2.7. Statistical Analyses
2.7.1. General Linear Model (GLM) Analyses
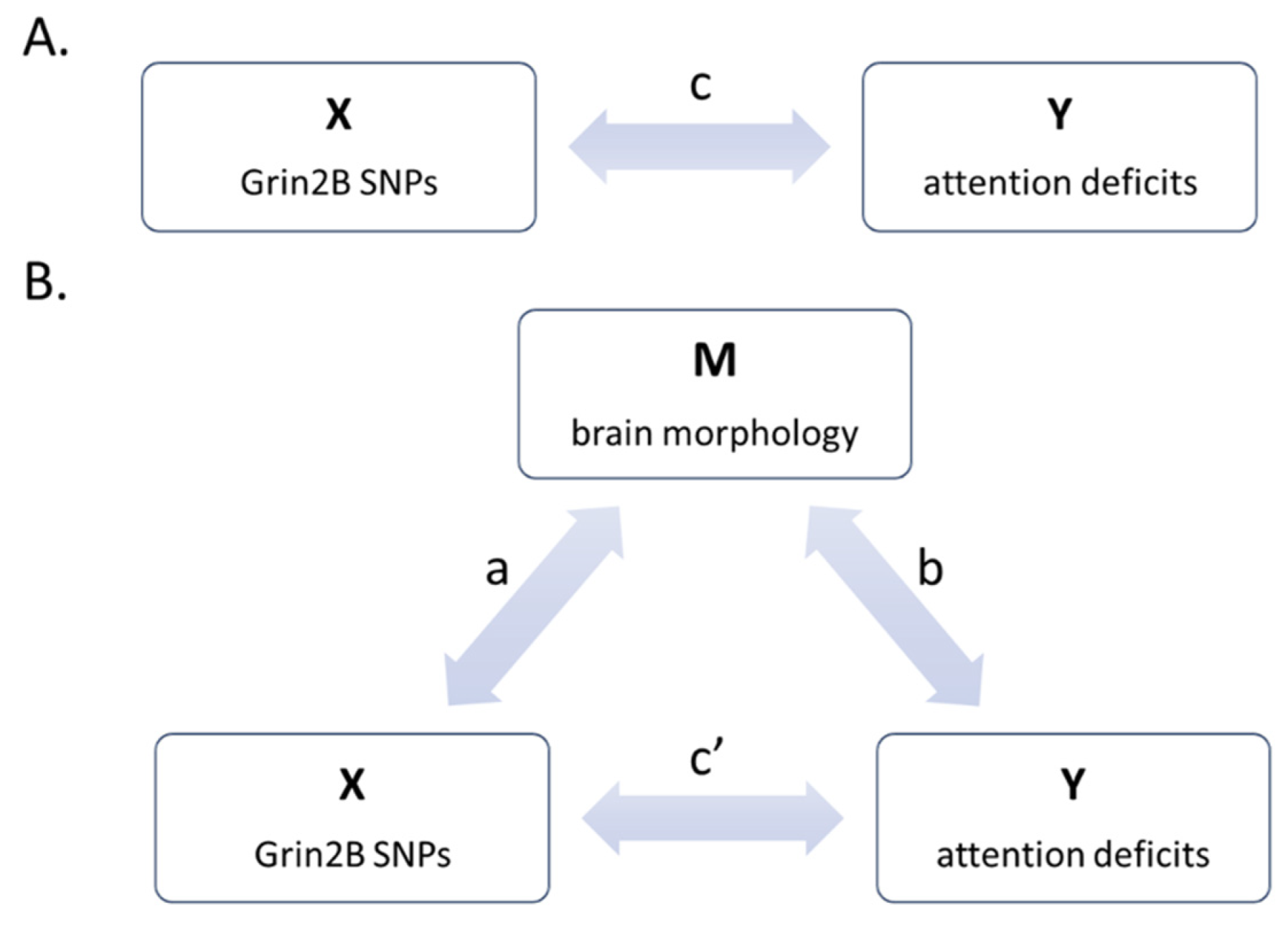
2.7.2. Mediation Analyses
3. Results
3.1. GLM Analyses
3.1.1. Design #1. GRIN2B Effects on Neuroanatomy
3.1.2. Design #2. GRIN2B Association with Attention/Hyperactivity Problems
3.1.3. Design #3. Neuroanatomy Effects on Attention/Hyperactivity Problems
3.2. Mediation Analyses
4. Discussion
4.1. GRIN2B Effect on Attention Deficits
4.2. GRIN2B Influence on Brain Structure
4.3. Brain Correlates of Attention/Hyperactivity Problems
4.4. From GRIN2B to Behavior through Neuroanatomy
4.5. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nobile, M.; Giorda, R.; Marino, C.; Carlet, O.; Pastore, V.; Vanzin, L.; Bellina, M.; Molteni, M.; Battaglia, M. Socioeconomic status mediates the genetic contribution of the dopamine receptor D4 and serotonin transporter linked promoter region repeat polymorphisms to externalization in preadolescence. Dev. Psychopathol. 2007, 19, 1147–1160. [Google Scholar] [CrossRef]
- Nobile, M.; Rusconi, M.; Bellina, M.; Marino, C.; Giorda, R.; Carlet, O.; Vanzin, L.; Molteni, M.; Battaglia, M. COMT Val158Met polymorphism and socioeconomic status interact to predict attention deficit/hyperactivity problems in children aged 10–14. Eur. Child Adolesc. Psychiatry 2009, 19, 549–557. [Google Scholar] [CrossRef] [Green Version]
- Leopold, D.R.; Christopher, M.E.; Olson, R.K.; Petrill, S.A.; Willcutt, E.G. Invariance of ADHD Symptoms Across Sex and Age: A Latent Analysis of ADHD and Impairment Ratings from Early Childhood into Adolescence. J. Abnorm. Child Psychol. 2018, 47, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Kuntsi, J.; Gayan, J.; Stevenson, J. Parents’ and teachers’ ratings of problem behaviours in children: Genetic and contrast effects. Twin Res. 2000, 3, 251–258. [Google Scholar] [PubMed]
- Rietveld, M.; Hudziak, J.; Bartels, M.; van Beijsterveldt, T.; Boomsma, D. Heritability of attention problems in children: Longitudinal results from a study of twins, age 3 to 12. J. Child Psychol. Psychiatry 2004, 45, 577–588. [Google Scholar] [CrossRef]
- Franke, B.; Faraone, S.; Asherson, P.; Buitelaar, J.; Bau, C.; Ramos-Quiroga, J.A.; Mick, E.; Grevet, E.; Johansson, S.; Havikk, J.; et al. The genetics of attention deficit/hyperactivity disorder in adults, a review. Mol. Psychiatry 2011, 17, 960–987. [Google Scholar] [CrossRef] [Green Version]
- Cortese, S. The neurobiology and genetics of Attention-Deficit/Hyperactivity Disorder (ADHD): What every clinician should know. Eur. J. Paediatr. Neurol. 2012, 16, 422–433. [Google Scholar] [CrossRef]
- Dorval, K.M.; Wigg, K.G.; Crosbie, J.; Tannock, R.; Kennedy, J.L.; Ickowicz, A.; Pathare, T.; Malone, M.; Schachar, R.; Barr, C.L. Association of the glutamate receptor subunit gene GRIN2B with attention-deficit/hyperactivity disorder. Genes Brain Behav. 2006, 6, 444–452. [Google Scholar] [CrossRef] [Green Version]
- Akazawa, C.; Shigemoto, R.; Bessho, Y.; Nakanishi, S.; Mizuno, N. Differential expression of five N-methyl-D-aspartate receptor subunit mRNAs in the cerebellum of developing and adult rats. J. Comp. Neurol. 1994, 347, 150–160. [Google Scholar] [CrossRef]
- Gill, I.; Droubi, S.; Giovedi’, S.; Fedder, K.; Bury, L.A.D.; Bosco, F.; Sceniak, M.P.; Benfenati, F.; Sabo, S.L. Presynaptic NMDA receptors: Dynamics and distribution in developing axons in vitro and in vivo. J. Cell Sci. 2015, 128, 768–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monyer, H.; Burnashev, N.; Laurie, D.J.; Sakmann, B.; Seeburg, P.H. Developmental and regional expression in the rat brain and functional properties of four NMDA receptors. Neuron 1994, 12, 529–540. [Google Scholar] [CrossRef]
- Kim, J.I.; Kim, J.-W.; Park, S.; Hong, S.-B.; Lee, D.S.; Paek, S.H.; Han, D.H.; Cheong, J.H.; Kim, B.-N. The GRIN2B and GRIN2A Gene Variants Are Associated with Continuous Performance Test Variables in ADHD. J. Atten. Disord. 2016, 24, 1538–1546. [Google Scholar] [CrossRef] [PubMed]
- Mascheretti, S.; Facoetti, A.; Giorda, R.; Beri, S.; Riva, V.; Trezzi, V.; Cellino, M.R.; Marino, C. GRIN2B mediates susceptibility to intelligence quotient and cognitive impairments in developmental dyslexia. Psychiatr. Genet. 2015, 25, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Endele, S.; Rosenberger, G.; Geider, K.; Popp, B.; Tamer, C.; Stefanova, I.; Milh, M.; Kortüm, F.; Fritsch, A.; Pientka, F.K.; et al. Mutations in GRIN2A and GRIN2B encoding regulatory subunits of NMDA receptors cause variable neurodevelopmental phenotypes. Nat. Genet. 2010, 42, 1021–1026. [Google Scholar] [CrossRef]
- Hassan, A.; Heckman, M.G.; Ahlskog, J.; Wszolek, Z.K.; Serie, D.J.; Uitti, R.J.; Van Gerpen, J.A.; Okun, M.S.; Rayaprolu, S.; Ross, O.A. Association of Parkinson disease age of onset with DRD2, DRD3 and GRIN2B polymorphisms. Park. Relat. Disord. 2016, 22, 102–105. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Chen, W.; Myers, S.J.; Yuan, H.; Traynelis, S.F. Human GRIN2B variants in neurodevelopmental disorders. J. Pharmacol. Sci. 2016, 132, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Lorenzi, C.; Delmonte, D.; Pirovano, A.; Marino, E.; Bongiorno, F.; Catalano, M.; Colombo, C.; Bramanti, P.; Smeraldi, E. Searching Susceptibility Loci for Bipolar Disorder: A Sib Pair Study on Chromosome 12. Neuropsychobiology 2010, 61, 10–18. [Google Scholar] [CrossRef]
- Seripa, D.; Matera, M.G.; Franceschi, M.; Bizzarro, A.; Paris, F.; Cascavilla, L.; Rinaldi, M.; Panza, F.; Solfrizzi, V.; Daniele, A.; et al. Association Analysis of GRIN2B, Encoding N-Methyl-D-Aspartate Receptor 2B Subunit, and Alzheimer’s Disease. Dement. Geriatr. Cogn. Disord. 2008, 25, 287–292. [Google Scholar] [CrossRef]
- Meyer-Lindenberg, A.; Weinberger, D.R. Intermediate phenotypes and genetic mechanisms of psychiatric disorders. Nat. Rev. Neurosci. 2006, 7, 818–827. [Google Scholar] [CrossRef] [PubMed]
- Zayed, A.; Robinson, G.E. Understanding the Relationship Between Brain Gene Expression and Social Behavior: Lessons from the Honey Bee. Annu. Rev. Genet. 2012, 46, 591–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuswanto, C.N.; Sum, M.Y.; Thng, C.R.Z.; Bin Zhang, Y.; Yang, G.L.; Nowinski, W.L.; Sitoh, Y.Y.; Low, C.M.; Sim, K. GRIN2B Gene and Associated Brain Cortical White Matter Changes in Bipolar Disorder: A Preliminary Combined Platform Investigation. BioMed Res. Int. 2013, 2013, 635131. [Google Scholar] [CrossRef] [Green Version]
- Stein, J.; Hua, X.; Morra, J.H.; Lee, S.; Hibar, D.P.; Ho, A.J.; Leow, A.D.; Toga, A.W.; Sul, J.H.; Kang, H.M.; et al. Genome-wide analysis reveals novel genes influencing temporal lobe structure with relevance to neurodegeneration in Alzheimer’s disease. NeuroImage 2010, 51, 542–554. [Google Scholar] [CrossRef] [Green Version]
- Arnold, P.D.; MacMaster, F.; Richter, M.A.; Hanna, G.; Sicard, T.; Burroughs, E.; Mirza, Y.; Easter, P.C.; Rose, M.; Kennedy, J.L.; et al. Glutamate receptor gene (GRIN2B) associated with reduced anterior cingulate glutamatergic concentration in pediatric obsessive–compulsive disorder. Psychiatry Res. Neuroimaging 2009, 172, 136–139. [Google Scholar] [CrossRef] [Green Version]
- Dalvie, S.; Brooks, S.J.; Cardenas, V.; Fein, G.; Ramesar, R.; Stein, D.J. Genetic variation within GRIN2B in adolescents with alcohol use disorder may be associated with larger left posterior cingulate cortex volume. Acta Neuropsychiatr. 2016, 29, 252–258. [Google Scholar] [CrossRef]
- Wilens, T.E.; Zulauf, C.A. Attention deficit hyperactivity disorder, and substance use disorders. In Youth Substance Abuse and Co-Occurring Disorders; Kaminer, Y., Arlington, V.A., Eds.; American Psychiatric Publishing: Washington, DC, USA, 2015; pp. 103–129. [Google Scholar]
- Norman, L.J.; Carlisi, C.; Lukito, S.; Hart, H.; Mataix-Cols, D.; Radua, J.; Rubia, K. Structural and Functional Brain Abnormalities in Attention-Deficit/Hyperactivity Disorder and Obsessive-Compulsive Disorder. JAMA Psychiatry 2016, 73, 815–825. [Google Scholar] [CrossRef]
- Bellina, M.; Brambilla, P.; Garzitto, M.; Negri, G.A.L.; Molteni, M.; Nobile, M. The ability of CBCL DSM-oriented scales to predict DSM-IV diagnoses in a referred sample of children and adolescents. Eur. Child Adolesc. Psychiatry 2012, 22, 235–246. [Google Scholar] [CrossRef]
- Bianchi, V.; Brambilla, P.; Garzitto, M.; Colombo, P.; Fornasari, L.; Bellina, M.; Bonivento, C.; Tesei, A.; Piccin, S.; Conte, S.; et al. Latent classes of emotional and behavioural problems in epidemiological and referred samples and their relations to DSM-IV diagnoses. Eur. Child Adolesc. Psychiatry 2016, 26, 549–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufman, J.; Birmaher, B.; Brent, D.A.; Ryan, N.D.; Rao, U. K-SADS-PL. J. Am. Acad. Child Adolesc. Psychiatry 2000, 39, 1208. [Google Scholar] [CrossRef]
- Achenbach, T.M.; Rescorla, L. Manual for the ASEBA School-Age Forms & Profiles; ASEBA: Burlington, VT, USA, 2001. [Google Scholar]
- Barratt, W. The Barratt Simplified Measure of Social Status (BSMSS): Measuring SES; Unpublished manuscript; Indiana State University: Terre Haute, IN, USA, 2006. [Google Scholar]
- Wechsler, D. Manual for the Wechsler Intelligence Scale for Children; revised; Psychological Corporation: San Antonio, TX, USA, 1974. [Google Scholar]
- Achenbach, T.M.; Verhulst, F.C.; Baron, G.D.; Althaus, M.; Veriiulst, F.C. A Comparison of Syndromes Derived from the Child Behavior Checklist for American and Dutch Boys Aged 6–11 and 12–16. J. Child Psychol. Psychiatry 1987, 28, 437–453. [Google Scholar] [CrossRef]
- Verhulst, F.C.; Achenbach, T.M.; Althaus, M.; Akkerhuis, G.W. A Comparison of Syndromes Derived from the Child Behavior Checklist for American and Dutch Girls Aged 6–11 And 12–16. J. Child Psychol. Psychiatry 1988, 29, 879–895. [Google Scholar] [CrossRef]
- Fischl, B. FreeSurfer. NeuroImage 2012, 62, 774–781. [Google Scholar] [CrossRef] [Green Version]
- Desikan, R.S.; Ségonne, F.; Fischl, B.; Quinn, B.T.; Dickerson, B.C.; Blacker, D.; Buckner, R.L.; Dale, A.M.; Maguire, R.P.; Hyman, B.T.; et al. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. NeuroImage 2006, 31, 968–980. [Google Scholar] [CrossRef]
- Preacher, K.J.; Hayes, A.F. Asymptotic and resampling strategies for assessing and comparing indirect effects in multiple mediator models. Behav. Res. Methods 2008, 40, 879–891. [Google Scholar] [CrossRef]
- DiCiccio, T.J.; Efron, B. Bootstrap confidence intervals. Stat. Sci. 1996, 11, 189–228. [Google Scholar] [CrossRef]
- Riva, V.; Battaglia, M.; Nobile, M.; Cattaneo, F.; Lazazzera, C.; Mascheretti, S.; Giorda, R.; Mérette, C.; Émond, C.; Maziade, M.; et al. GRIN2B predicts attention problems among disadvantaged children. Eur. Child Adolesc. Psychiatry 2014, 24, 827–836. [Google Scholar] [CrossRef]
- Wu, K.; Hanna, G.; Easter, P.; Kennedy, J.L.; Rosenberg, D.R.; Arnold, P.D. Glutamate system genes and brain volume alterations in pediatric obsessive-compulsive disorder: A preliminary study. Psychiatry Res. Neuroimaging 2013, 211, 214–220. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.I.; Yoo, J.H.; Kim, D.; Jeong, B.; Kim, B.-N. The effects of GRIN2B and DRD4 gene variants on local functional connectivity in attention-deficit/hyperactivity disorder. Brain Imaging Behav. 2017, 12, 247–257. [Google Scholar] [CrossRef]
- Greicius, M.D.; Krasnow, B.; Reiss, A.L.; Menon, V. Functional connectivity in the resting brain: A network analysis of the default mode hypothesis. Proc. Natl. Acad. Sci. USA 2003, 100, 253–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismaylova, E.; Di Sante, J.; Gouin, J.-P.; Pomares, F.; Vitaro, F.; Tremblay, R.E.; Booij, L. Associations Between Daily Mood States and Brain Gray Matter Volume, Resting-State Functional Connectivity and Task-Based Activity in Healthy Adults. Front. Hum. Neurosci. 2018, 12, 168. [Google Scholar] [CrossRef] [Green Version]
- Fransson, P. Spontaneous low-frequency BOLD signal fluctuations: An fMRI investigation of the resting-state default mode of brain function hypothesis. Hum. Brain Mapp. 2005, 26, 15–29. [Google Scholar] [CrossRef]
- Fransson, P. How default is the default mode of brain function? Further evidence from intrinsic BOLD signal fluctuations. Neuropsychologia 2006, 44, 2836–2845. [Google Scholar] [CrossRef]
- Leech, R.; Sharp, D.J. The role of the posterior cingulate cortex in cognition and disease. Brain 2014, 137, 12–32. [Google Scholar] [CrossRef] [Green Version]
- Gusnard, D.A.; Akbudak, E.; Shulman, G.L.; Raichle, M.E. Medial prefrontal cortex and self-referential mental activity: Relation to a default mode of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, 4259–4264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonuga-Barke, E.J.; Castellanos, F.X. Spontaneous attentional fluctuations in impaired states and pathological conditions: A neurobiological hypothesis. Neurosci. Biobehav. Rev. 2007, 31, 977–986. [Google Scholar] [CrossRef] [PubMed]
- Leech, R.; Kamourieh, S.; Beckmann, C.F.; Sharp, D.J. Fractionating the Default Mode Network: Distinct Contributions of the Ventral and Dorsal Posterior Cingulate Cortex to Cognitive Control. J. Neurosci. 2011, 31, 3217–3224. [Google Scholar] [CrossRef]
- Leech, R.; Braga, R.; Sharp, D.J. Echoes of the Brain within the Posterior Cingulate Cortex. J. Neurosci. 2012, 32, 215–222. [Google Scholar] [CrossRef] [Green Version]
- Vincent, J.L.; Kahn, I.; Snyder, A.Z.; Raichle, M.E.; Buckner, R.L. Evidence for a Frontoparietal Control System Revealed by Intrinsic Functional Connectivity. J. Neurophysiol. 2008, 100, 3328–3342. [Google Scholar] [CrossRef] [Green Version]
- Spreng, R.N. The fallacy of a “task-negative” network. Front. Psychol. 2012, 3, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, C.; Uddin, L.; Biswal, B.B.; Castellanos, F.X.; Milham, M.P. Competition between functional brain networks mediates behavioral variability. NeuroImage 2008, 39, 527–537. [Google Scholar] [CrossRef]
- Mesulam, M.; Nobre, A.C.; Kim, Y.-H.; Parrish, T.; Gitelman, D. Heterogeneity of Cingulate Contributions to Spatial Attention. NeuroImage 2001, 13, 1065–1072. [Google Scholar] [CrossRef]
- Wagner, A.D.; Shannon, B.J.; Kahn, I.; Buckner, R.L. Parietal lobe contributions to episodic memory retrieval. Trends Cogn. Sci. 2005, 9, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Maddock, R. The retrosplenial cortex and emotion: New insights from functional neuroimaging of the human brain. Trends Neurosci. 1999, 22, 310–316. [Google Scholar] [CrossRef]
- Nobile, M.; Rusconi, M.; Bellina, M.; Marino, C.; Giorda, R.; Carlet, O.; Vanzin, L.; Molteni, M.; Battaglia, M. The influence of family structure, the TPH2 G-703T and the 5-HTTLPR serotonergic genes upon affective problems in children aged 10–14 years. J. Child Psychol. Psychiatry 2009, 50, 317–325. [Google Scholar] [CrossRef]
- Plomin, R. Child Development and Molecular Genetics: 14 Years Later. Child Dev. 2012, 84, 104–120. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Males (n = 42) | Females (n = 16) | Total (n = 58) | |
|---|---|---|---|
| Wave 0 | |||
| Age (years) 1 | 8.55 ± 2.51 | 9.38 ± 2.07 | 8.78 ± 2.43 |
| SES 1 | 49.05 ± 18.59 | 36.25 ± 21.18 | 45.52 ± 20.17 |
| CBCL Attention Problems 1 | 63.10 ± 9.40 | 66.56 ± 10.37 | 64.05 ± 9.80 |
| FSIQ 1 | 108.81 ± 16.39 | 109.38 ± 13.93 | 108.96 ± 15.65 |
| K-SADS-PL DIAGNOSIS 2 | |||
| ADHD | N = 9 (21.4%) | N = 4 (25.0%) | N = 13 (22.4%) |
| Any behavioral disorder | N = 10 (23.8%) | N = 0 (0.0%) | N = 10 (17.2%) |
| Any mood disorder | N = 12 (28.6%) | N = 6 (37.5%) | N = 18 (31.0%) |
| Any anxiety disorder | N = 19 (45.2%) | N = 9 (56.3%) | N = 28 (48.3%) |
| Other diagnoses | N = 7 (16.7%) | N = 5 (31.3%) | N = 12 (20.7%) |
| No current diagnosis | N = 2 (4.8%) | N = 1 (6.3%) | N = 3 (5.2%) |
| Comorbidities | 1 diagnosis: N = 25 (59.5%) | 1 diagnosis: N = 8 (50.0%) | 1 diagnosis: N = 33 (56.9%) |
| 2 diagnoses: N = 13 (31.0%) | 2 diagnoses: N = 5 (31.3%) | 2 diagnoses: N = 18 (31.0%) | |
| 3 diagnoses: N = 2 (4.8%) | 3 diagnoses: N = 2 (12.5%) | 3 diagnoses: N = 4 (6.9%) | |
| Wave 1 | |||
| Age (years) 1 | 14.12 ± 2.09 | 15.58 ± 2.24 | 14.52 ± 2.23 |
| CBCL Attention Problems1 | 59.48 ± 7.86 | 62.06 ± 7.24 | 60.19 ± 7.78 |
| Wave 2 | |||
| Age (years) 1 | 15.80 ± 2.30 | 17.14 ± 2.43 | 16.17 ± 2.41 |
| CBCL Attention Problems 1 | 57.83 ± 6.88 | 59.00 ± 4.98 | 58.16 ± 6.43 |
| GRIN2B SNP | Allele | Frequency 1 | Hardy-Weinberg Equilibrium |
|---|---|---|---|
| rs5796555 | - | 0.71 | 0.201 |
| A | 0.29 | ||
| rs1012586 | G | 0.66 | 0.744 |
| C | 0.34 | ||
| rs2268119 | A | 0.73 | 0.213 |
| T | 0.27 | ||
| rs2216128 | A | 0.74 | 0.146 |
| G | 0.26 | ||
| rs11609779 | C | 0.84 | 0.546 |
| T | 0.16 | ||
| rs2192973 | G | 0.78 | 0.115 |
| A | 0.22 |
| GRIN2B SNP | Allele | FS Feature | Brain Region | T 1 | p | pcorr 2 |
|---|---|---|---|---|---|---|
| rs5796555 | “-/A” and “A/A” | Volume | Left inferior parietal | 3.72 | <0.001 | <0.05 |
| Left isthmus cingulate | 3.62 | <0.001 | <0.05 | |||
| Left middle temporal | 3.90 | <0.001 | <0.05 | |||
| Left pars orbitalis | 3.72 | <0.001 | <0.05 | |||
| Left precuneus | 3.78 | <0.001 | <0.05 | |||
| Left rostral middle frontal | 3.59 | <0.001 | <0.05 | |||
| Right caudal ACC | 4.32 | <0.0001 | <0.01 | |||
| Right inferior parietal | 4.14 | <0.001 | <0.01 | |||
| Right middle temporal | 3.43 | 0.001 | <0.05 | |||
| Right pars orbitalis | 3.72 | <0.001 | <0.05 | |||
| Right rostral ACC | 3.95 | <0.001 | <0.01 | |||
| Right rostral middle frontal | 3.49 | <0.001 | <0.05 | |||
| Right transverse temporal | 3.40 | 0.001 | <0.05 | |||
| rs2268119 | “A/T” and “T/T” | Area | Left lateral orbitofrontal | 3.40 | 0.001 | <0.05 |
| Right lateral occipital | 3.98 | <0.001 | <0.01 | |||
| rs2216128 | “G/C” and “C/C” | Right isthmus cingulate | 3.42 | 0.001 | <0.05 |
| Mediation Analysis 1 | Mediation Analysis 2 | ||||||
|---|---|---|---|---|---|---|---|
| Parameter | Value | 95% CI | p | Parameter | Value | 95% CI | p |
| a | –0.45 | [–0.26 0.26] | <0.001 | a | 0.48 | [–0.26 0.26] | <0.001 |
| b | –0.25 | [–0.27 0.26] | 0.037 | b | −0.18 | [–0.25 0.29] | 0.08 |
| c | 0.31 | [–0.27 0.26] | 0.011 | c | 0.31 | [–0.27 0.26] | 0.011 |
| ab | 0.11 | [–0.04 0.05] | <0.001 | ab | n.e. | n.e. | n.e. |
| c’ | 0.20 | [–0.27 0.27] | 0.09 | c’ | n.e. | n.e. | n.e. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nobile, M.; Maggioni, E.; Mauri, M.; Garzitto, M.; Piccin, S.; Bonivento, C.; Giorda, R.; Girometti, R.; Tomasino, B.; Molteni, M.; et al. Brain Anatomical Mediators of GRIN2B Gene Association with Attention/Hyperactivity Problems: An Integrated Genetic-Neuroimaging Study. Genes 2021, 12, 1193. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12081193
Nobile M, Maggioni E, Mauri M, Garzitto M, Piccin S, Bonivento C, Giorda R, Girometti R, Tomasino B, Molteni M, et al. Brain Anatomical Mediators of GRIN2B Gene Association with Attention/Hyperactivity Problems: An Integrated Genetic-Neuroimaging Study. Genes. 2021; 12(8):1193. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12081193
Chicago/Turabian StyleNobile, Maria, Eleonora Maggioni, Maddalena Mauri, Marco Garzitto, Sara Piccin, Carolina Bonivento, Roberto Giorda, Rossano Girometti, Barbara Tomasino, Massimo Molteni, and et al. 2021. "Brain Anatomical Mediators of GRIN2B Gene Association with Attention/Hyperactivity Problems: An Integrated Genetic-Neuroimaging Study" Genes 12, no. 8: 1193. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12081193