
A High-Copy Suppressor Screen Reveals a Broad Role of Prefoldin-like Bud27 in the TOR Signaling Pathway in Saccharomyces cerevisiae

 , , , , and
, , , , and 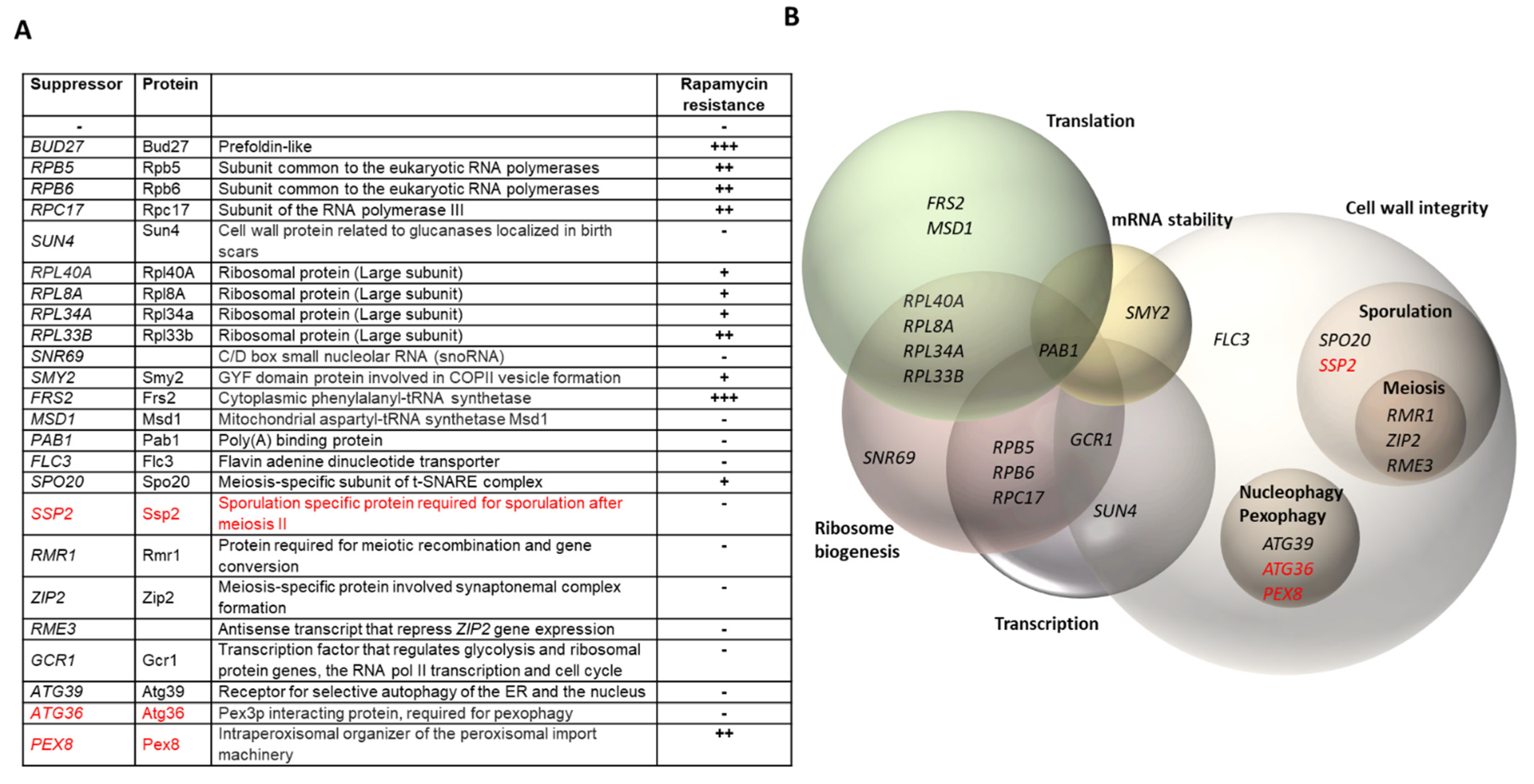{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains, Genetic Manipulations, Media, and Genetic Analyses
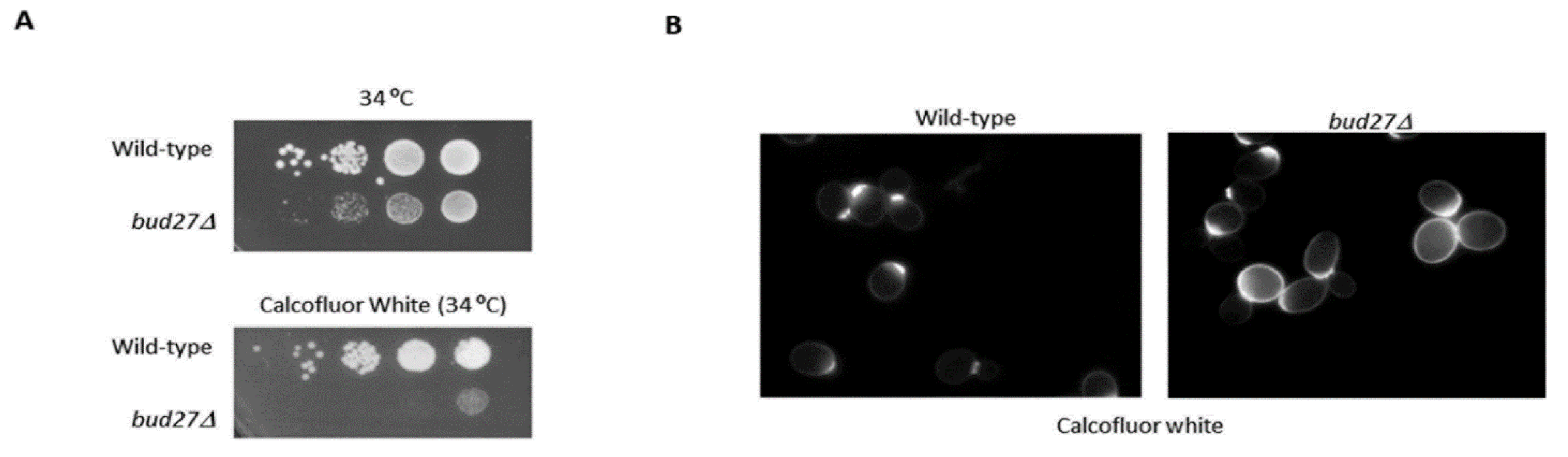
2.2. Chitin Staining with Calcofluor White and Fluorescence Microscopy
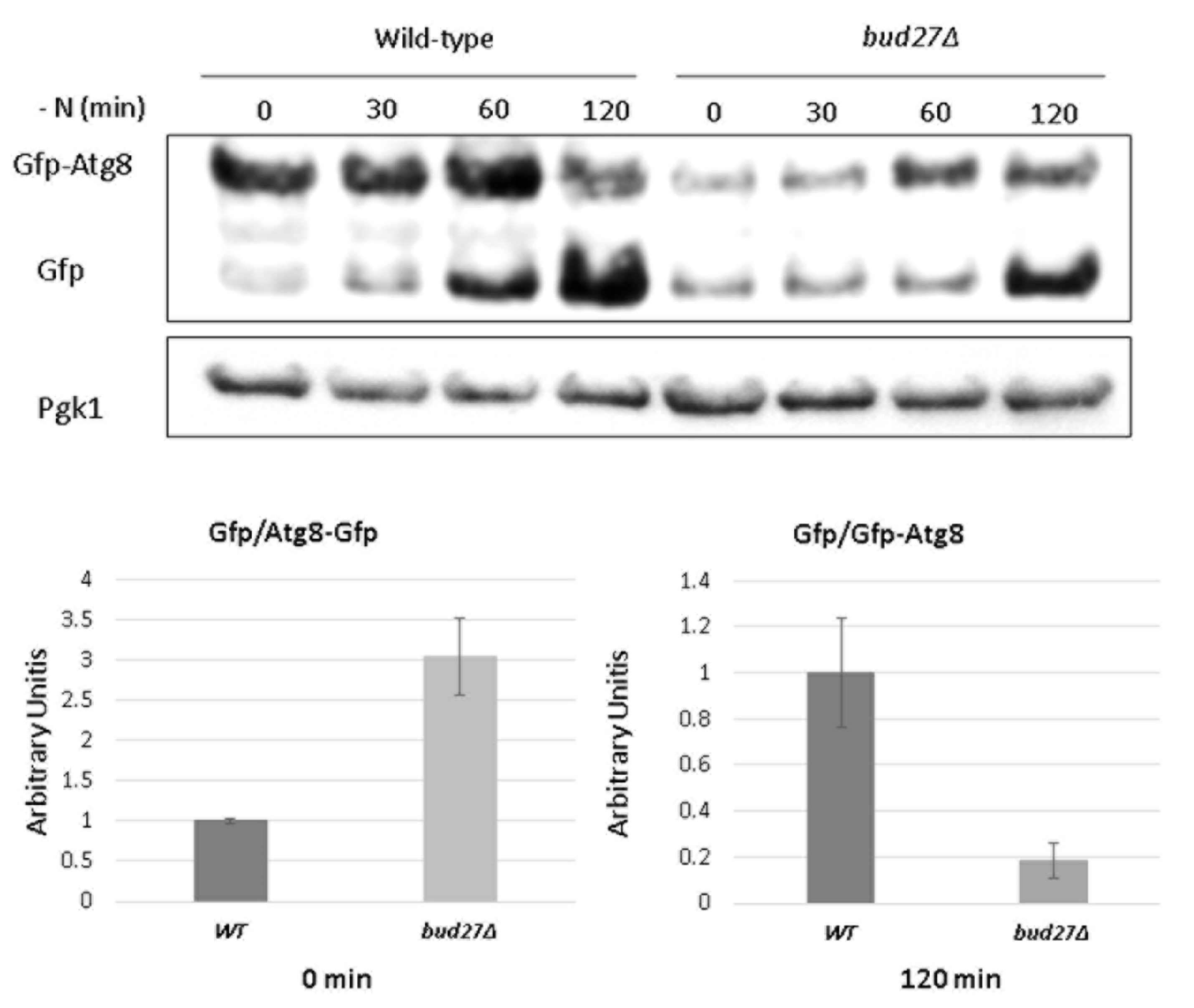
2.3. Autophagy Detection
3. Results and Discussion
3.1. Genetic Screening for Multicopy Suppressors of the Temperature-Sensitive Growth Defect of the bud27Δ Mutant
3.2. Transcriptomic Analyses Reinforce the Role of Bud27 in the TOR Signaling Pathway and in Cell Wall Integrity
3.3. Lack of Bud27 May Cause Slower Autophagy Induction
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Loewith, R.; Hall, M.N. Target of rapamycin (TOR) in nutrient signaling and growth control. Genetics 2011, 189, 1177–1201. [Google Scholar] [CrossRef] [Green Version]
- Loewith, R. A brief history of TOR. Biochem. Soc. Trans. 2011, 39, 437–442. [Google Scholar] [CrossRef] [Green Version]
- Zaman, S.; Lippman, S.I.; Zhao, X.; Broach, J.R. How Saccharomyces responds to nutrients. Annu. Rev. Genet. 2008, 42, 27–81. [Google Scholar] [CrossRef]
- Gentilella, A.; Kozma, S.C.; Thomas, G. A liaison between mTOR signaling, ribosome biogenesis and cancer. Biochim. Biophys. Acta 2015, 1849, 812–820. [Google Scholar] [CrossRef] [Green Version]
- González, A.; Hall, M.N. Nutrient sensing and TOR signaling in yeast and mammals. EMBO J. 2017, 36, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Worley, J.; Sullivan, A.; Luo, X.; Kaplan, M.E.; Capaldi, A.P. Genome-Wide Analysis of the TORC1 and Osmotic Stress Signaling Network in Saccharomyces cerevisiae. G3 (Bethesda) 2015, 6, 463–474. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Awasthi, A.; Nain, V.; Issac, B.; Puria, R. Novel insights into TOR signalling in Saccharomyces cerevisiae through Torin2. Gene 2018, 669, 15–27. [Google Scholar] [CrossRef]
- Yan, G.; Lai, Y.; Jiang, Y. TOR under stress: Targeting TORC1 by Rho1 GTPase. Cell Cycle 2012, 11, 3384–3388. [Google Scholar] [CrossRef]
- So, Y.S.; Lee, D.G.; Idnurm, A.; Ianiri, G.; Bahn, Y.S. The TOR Pathway Plays Pleiotropic Roles in Growth and Stress Responses of the Fungal Pathogen Cryptococcus neoformans. Genetics 2019, 212, 1241–1258. [Google Scholar] [CrossRef]
- Plank, M. Interaction of TOR and PKA Signaling in S. cerevisiae. Biomolecules 2022, 12, 210. [Google Scholar] [CrossRef]
- Sengupta, S.; Peterson, T.R.; Sabatini, D.M. Regulation of the mTOR complex 1 pathway by nutrients, growth factors, and stress. Mol. Cell 2010, 40, 310–322. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Lancaster, C.S.; Shi, B.; Guo, H.; Thimmaiah, P.; Bjornsti, M.A. TOR signaling is a determinant of cell survival in response to DNA damage. Mol. Cell Biol. 2007, 27, 7007–7017. [Google Scholar] [CrossRef] [Green Version]
- Xiao, L.; Grove, A. Coordination of ribosomal protein and ribosomal RNA gene expression in response to TOR signaling. Curr. Genom. 2009, 10, 198–205. [Google Scholar] [CrossRef] [Green Version]
- Vizoso-Vázquez, A.; Barreiro-Alonso, A.; González-Siso, M.I.; Rodríguez-Belmonte, E.; Lamas-Maceiras, M.; Cerdán, M.E. HMGB proteins involved in TOR signaling as general regulators of cell growth by controlling ribosome biogenesis. Curr. Genet. 2018, 64, 1205–1213. [Google Scholar] [CrossRef]
- Duvel, K.; Santhanam, A.; Garrett, S.; Schneper, L.; Broach, J.R. Multiple roles of Tap42 in mediating rapamycin-induced transcriptional changes in yeast. Mol. Cell 2003, 11, 1467–1478. [Google Scholar] [CrossRef]
- Farre, J.C.; Subramani, S. Mechanistic insights into selective autophagy pathways: Lessons from yeast. Nat. Rev. Mol. Cell Biol. 2016, 17, 537–552. [Google Scholar] [CrossRef]
- Diaz-Troya, S.; Perez-Perez, M.E.; Florencio, F.J.; Crespo, J.L. The role of TOR in autophagy regulation from yeast to plants and mammals. Autophagy 2008, 4, 851–865. [Google Scholar] [CrossRef]
- Cebollero, E.; Reggiori, F. Regulation of autophagy in yeast Saccharomyces cerevisiae. Biochim. Biophys. Acta 2009, 1793, 1413–1421. [Google Scholar] [CrossRef] [Green Version]
- Crespo, J.L.; Hall, M.N. Elucidating TOR signaling and rapamycin action: Lessons from Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2002, 66, 579–591. [Google Scholar] [CrossRef] [Green Version]
- Loewith, R.; Jacinto, E.; Wullschleger, S.; Lorberg, A.; Crespo, J.L.; Bonenfant, D.; Oppliger, W.; Jenoe, P.; Hall, M.N. Two TOR complexes, only one of which is rapamycin sensitive, have distinct roles in cell growth control. Mol. Cell 2002, 10, 457–468. [Google Scholar] [CrossRef]
- Inoki, K.; Ouyang, H.; Li, Y.; Guan, K.L. Signaling by target of rapamycin proteins in cell growth control. Microbiol. Mol. Biol. Rev. 2005, 69, 79–100. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.E.; Hall, M.N. The expanding TOR signaling network. Curr. Opin. Cell Biol. 2005, 17, 158–166. [Google Scholar] [CrossRef]
- Martínez-Fernández, V.; Garrido-Godino, A.I.; Cuevas-Bermúdez, A.; Navarro, F. Cytoplasmic and nuclear functions for the prefoldin-like URI/Bud27. In New Research On Molecular Chaperones; Chaperones, N.R.O.M., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2015; pp. 57–73. [Google Scholar]
- Martínez-Fernández, V.; Garrido-Godino, A.I.; Cuevas-Bermúdez, A.; Navarro, F. The yeast prefoldin Bud27. Adv. Exp. Med. Biol. 2018, 1106, 109–118. [Google Scholar]
- Martinez-Fernandez, V.; Cuevas-Bermudez, A.; Gutierrez-Santiago, F.; Garrido-Godino, A.I.; Rodriguez-Galan, O.; Jordan-Pla, A.; Lois, S.; Trivino, J.C.; de la Cruz, J.; Navarro, F. Prefoldin-like Bud27 influences the transcription of ribosomal components and ribosome biogenesis in Saccharomyces cerevisiae. RNA 2020, 26, 1360–1379. [Google Scholar] [CrossRef]
- Gstaiger, M.; Luke, B.; Hess, D.; Oakeley, E.J.; Wirbelauer, C.; Blondel, M.; Vigneron, M.; Peter, M.; Krek, W. Control of nutrient-sensitive transcription programs by the unconventional prefoldin URI. Science 2003, 302, 1208–1212. [Google Scholar] [CrossRef]
- Mirón-García, M.C.; Garrido-Godino, A.I.; Martínez-Fernández, V.; Fernández-Pevida, A.; Cuevas-Bermúdez, A.; Martín-Expósito, M.; Chávez, S.; de la Cruz, J.; Navarro, F. The yeast prefoldin-like URI-orthologue Bud27 associates with the RSC nucleosome remodeler and modulates transcription. Nucleic Acids Res. 2014, 42, 9666–9676. [Google Scholar] [CrossRef]
- Vernekar, D.V.; Bhargava, P. Yeast Bud27 modulates the biogenesis of Rpc128 and Rpc160 subunits and the assembly of RNA polymerase III. Biochim. Biophys. Acta 2015, 1849, 1340–1353. [Google Scholar] [CrossRef]
- Mirón-García, M.C.; Garrido-Godino, A.I.; García-Molinero, V.; Hernández-Torres, F.; Rodríguez-Navarro, S.; Navarro, F. The prefoldin Bud27 mediates the assembly of the eukaryotic RNA polymerases in an Rpb5-dependent manner. PLoS Genet. 2013, 9, e1003297. [Google Scholar] [CrossRef] [Green Version]
- Mita, P.; Savas, J.N.; Ha, S.; Djouder, N.; Yates, J.R., 3rd; Logan, S.K. Analysis of URI nuclear interaction with RPB5 and components of the R2TP/Prefoldin-Like complex. PLoS ONE 2013, 8, e63879. [Google Scholar] [CrossRef]
- Deplazes, A.; Mockli, N.; Luke, B.; Auerbach, D.; Peter, M. Yeast Uri1p promotes translation initiation and may provide a link to cotranslational quality control. EMBO J. 2009, 28, 1429–1441. [Google Scholar] [CrossRef] [Green Version]
- García-López, M.C.; Mirón-García, M.C.; Garrido-Godino, A.I.; Mingorance, C.; Navarro, F. Overexpression of SNG1 causes 6-azauracil resistance in Saccharomyces cerevisiae. Curr. Genet. 2010, 56, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Krick, R.; Bremer, S.; Welter, E.; Schlotterhose, P.; Muehe, Y.; Eskelinen, E.L.; Thumm, M. Cdc48/p97 and Shp1/p47 regulate autophagosome biogenesis in concert with ubiquitin-like Atg8. J. Cell Biol. 2010, 190, 965–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stettler, S.; Chiannilkulchai, N.; Hermann-Le Denmat, S.; Lalo, D.; Lacroute, F.; Sentenac, A.; Thuriaux, P. A general suppressor of RNA polymerase I, II and III mutations in Saccharomyces cerevisiae. Mol. Gen. Genet. 1993, 239, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Montella-Manuel, S.; Pujol-Carrion, N.; Mechoud, M.A.; de la Torre-Ruiz, M.A. Bulk autophagy induction and life extension is achieved when iron is the only limited nutrient in Saccharomyces cerevisiae. Biochem. J. 2021, 478, 811–837. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Moir, R.D.; McIntosh, K.B.; Willis, I.M. TOR Signaling Regulates Ribosome and tRNA Synthesis via LAMMER/Clk and GSK-3 Family Kinases. Mol. Cell 2012, 45, 836–843. [Google Scholar] [CrossRef] [Green Version]
- Aquino, G.R.R.; Krogh, N.; Hackert, P.; Martin, R.; Gallesio, J.D.; van Nues, R.W.; Schneider, C.; Watkins, N.J.; Nielsen, H.; Bohnsack, K.E.; et al. RNA helicase-mediated regulation of snoRNP dynamics on pre-ribosomes and rRNA 2’-O-methylation. Nucleic Acids Res. 2021, 49, 4066–4084. [Google Scholar] [CrossRef]
- Okano, A.; Wan, K.; Kanda, K.; Yabuki, Y.; Funato, K.; Mizuta, K. SMY2 and SYH1 suppress defects in ribosome biogenesis caused by ebp2 mutations. Biosci. Biotechnol. Biochem. 2015, 79, 1481–1483. [Google Scholar] [CrossRef]
- Sezen, B. Reduction of Saccharomyces cerevisiae Pom34 protein level by SESA network is related to membrane lipid composition. FEMS Yeast Res. 2015, 15, fov089. [Google Scholar] [CrossRef] [Green Version]
- Sanni, A.; Mirande, M.; Ebel, J.P.; Boulanger, Y.; Waller, J.P.; Fasiolo, F. Structure and expression of the genes encoding the alpha and beta subunits of yeast phenylalanyl-tRNA synthetase. J. Biol. Chem. 1988, 263, 15407–15415. [Google Scholar] [CrossRef]
- Gampel, A.; Tzagoloff, A. Homology of aspartyl- and lysyl-tRNA synthetases. Proc. Natl. Acad. Sci. USA 1989, 86, 6023–6027. [Google Scholar] [CrossRef] [Green Version]
- Brune, C.; Munchel, S.E.; Fischer, N.; Podtelejnikov, A.V.; Weis, K. Yeast poly(A)-binding protein Pab1 shuttles between the nucleus and the cytoplasm and functions in mRNA export. RNA 2005, 11, 517–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brengues, M.; Parker, R. Accumulation of polyadenylated mRNA, Pab1p, eIF4E, and eIF4G with P-bodies in Saccharomyces cerevisiae. Mol. Biol. Cell 2007, 18, 2592–2602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moqtaderi, Z.; Geisberg, J.V.; Struhl, K. Extensive structural differences of closely related 3’ mRNA isoforms: Links to Pab1 binding and mRNA stability. Mol. Cell 2018, 72, 849–861.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brambilla, M.; Martani, F.; Bertacchi, S.; Vitangeli, I.; Branduardi, P. The Saccharomyces cerevisiae poly (A) binding protein (Pab1): Master regulator of mRNA metabolism and cell physiology. Yeast 2019, 36, 23–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ash, M.R.; Faelber, K.; Kosslick, D.; Albert, G.I.; Roske, Y.; Kofler, M.; Schuemann, M.; Krause, E.; Freund, C. Conserved beta-hairpin recognition by the GYF domains of Smy2 and GIGYF2 in mRNA surveillance and vesicular transport complexes. Structure 2010, 18, 944–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Protchenko, O.; Rodriguez-Suarez, R.; Androphy, R.; Bussey, H.; Philpott, C.C. A screen for genes of heme uptake identifies the FLC family required for import of FAD into the endoplasmic reticulum. J. Biol. Chem. 2006, 281, 21445–21457. [Google Scholar] [CrossRef] [Green Version]
- Sickmann, A.; Reinders, J.; Wagner, Y.; Joppich, C.; Zahedi, R.; Meyer, H.E.; Schonfisch, B.; Perschil, I.; Chacinska, A.; Guiard, B.; et al. The proteome of Saccharomyces cerevisiae mitochondria. Proc. Natl. Acad. Sci. USA 2003, 100, 13207–13212. [Google Scholar] [CrossRef] [Green Version]
- Mouassite, M.; Camougrand, N.; Schwob, E.; Demaison, G.; Laclau, M.; Guerin, M. The ‘SUN’ family: Yeast SUN4/SCW3 is involved in cell septation. Yeast 2000, 16, 905–919. [Google Scholar] [CrossRef]
- Tatjer, L.; Gonzalez, A.; Serra-Cardona, A.; Barcelo, A.; Casamayor, A.; Arino, J. The Saccharomyces cerevisiae Ptc1 protein phosphatase attenuates G2-M cell cycle blockage caused by activation of the cell wall integrity pathway. Mol. Microbiol. 2016, 101, 671–687. [Google Scholar] [CrossRef]
- Ahmed, K.; Carter, D.E.; Lajoie, P. Hyperactive TORC1 sensitizes yeast cells to endoplasmic reticulum stress by compromising cell wall integrity. FEBS Lett. 2019, 593, 1957–1973. [Google Scholar] [CrossRef]
- Levin, D.E. Cell wall integrity signaling in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2005, 69, 262–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, H.; Morishita, M.; Schwartz, C.L.; Coluccio, A.; Engebrecht, J.; Neiman, A.M. Phospholipase D and the SNARE Sso1p are necessary for vesicle fusion during sporulation in yeast. J. Cell Sci. 2006, 119 Pt 7, 1406–1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Agarwal, S.; Roeder, G.S. SSP2 and OSW1, Two Sporulation-specific Genes Involved in Spore Morphogenesis in Saccharomyces cerevisiae. Genetics 2006, 175, 143–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neiman, A.M. Sporulation in the budding yeast Saccharomyces cerevisiae. Genetics 2011, 189, 737–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan, P.W.; Klein, F.; Leach, D.R. Novel roles for selected genes in meiotic DNA processing. PLoS Genet. 2007, 3, e222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chua, P.R.; Roeder, G.S. Zip2, a meiosis-specific protein required for the initiation of chromosome synapsis. Cell 1998, 93, 349–359. [Google Scholar] [CrossRef] [Green Version]
- Gelfand, B.; Mead, J.; Bruning, A.; Apostolopoulos, N.; Tadigotla, V.; Nagaraj, V.; Sengupta, A.M.; Vershon, A.K. Regulated antisense transcription controls expression of cell-type-specific genes in yeast. Mol. Cell Biol. 2011, 31, 1701–1709. [Google Scholar] [CrossRef] [Green Version]
- Shaw, R.J.; Wilson, J.L.; Smith, K.T.; Reines, D. Regulation of an IMP dehydrogenase gene and its overexpression in drug-sensitive transcription elongation mutants of yeast. J. Biol. Chem. 2001, 276, 32905–32916. [Google Scholar] [CrossRef] [Green Version]
- Deminoff, S.J.; Santangelo, G.M. Rap1p requires Gcr1p and Gcr2p homodimers to activate ribosomal protein and glycolytic genes, respectively. Genetics 2001, 158, 133–143. [Google Scholar] [CrossRef]
- Ravi, C.; Gowsalya, R.; Nachiappan, V. Impaired GCR1 transcription resulted in defective inositol levels, vacuolar structure and autophagy in Saccharomyces cerevisiae. Curr. Genet. 2019, 65, 995–1014. [Google Scholar] [CrossRef]
- Tornow, J.; Zeng, X.; Gao, W.; Santangelo, G.M. GCR1, a transcriptional activator in Saccharomyces cerevisiae, complexes with RAP1 and can function without its DNA binding domain. EMBO J. 1993, 12, 2431–2437. [Google Scholar] [CrossRef] [PubMed]
- Hall, D.B.; Wade, J.T.; Struhl, K. An HMG protein, Hmo1, associates with promoters of many ribosomal protein genes and throughout the rRNA gene locus in Saccharomyces cerevisiae. Mol. Cell Biol. 2006, 26, 3672–3679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mochida, K.; Oikawa, Y.; Kimura, Y.; Kirisako, H.; Hirano, H.; Ohsumi, Y.; Nakatogawa, H. Receptor-mediated selective autophagy degrades the endoplasmic reticulum and the nucleus. Nature 2015, 522, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Yorimitsu, T.; Nair, U.; Yang, Z.; Klionsky, D.J. Endoplasmic reticulum stress triggers autophagy. J. Biol. Chem. 2006, 281, 30299–30304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, R.; Bermejo, C.; Grau, C.; Perez, R.; Rodriguez-Pena, J.M.; Francois, J.; Nombela, C.; Arroyo, J. The global transcriptional response to transient cell wall damage in Saccharomyces cerevisiae and its regulation by the cell integrity signaling pathway. J. Biol. Chem. 2004, 279, 15183–15195. [Google Scholar] [CrossRef] [Green Version]
- Barve, G.; Manjithaya, R. Cross-talk between autophagy and sporulation in Saccharomyces cerevisiae. Yeast 2021, 38, 401–413. [Google Scholar] [CrossRef]
- Meguro, S.; Zhuang, X.; Kirisako, H.; Nakatogawa, H. Pex3 confines pexophagy receptor activity of Atg36 to peroxisomes by regulating Hrr25-mediated phosphorylation and proteasomal degradation. J. Biol. Chem. 2020, 295, 16292–16298. [Google Scholar] [CrossRef]
- Motley, A.M.; Nuttall, J.M.; Hettema, E.H. Pex3-anchored Atg36 tags peroxisomes for degradation in Saccharomyces cerevisiae. EMBO J. 2012, 31, 2852–2868. [Google Scholar] [CrossRef] [Green Version]
- Motley, A.M.; Nuttall, J.M.; Hettema, E.H. Atg36: The Saccharomyces cerevisiae receptor for pexophagy. Autophagy 2012, 8, 1680–1681. [Google Scholar] [CrossRef] [Green Version]
- Wroblewska, J.P.; Cruz-Zaragoza, L.D.; Yuan, W.; Schummer, A.; Chuartzman, S.G.; de Boer, R.; Oeljeklaus, S.; Schuldiner, M.; Zalckvar, E.; Warscheid, B.; et al. Saccharomyces cerevisiae cells lacking Pex3 contain membrane vesicles that harbor a subset of peroxisomal membrane proteins. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 1656–1667. [Google Scholar] [CrossRef]
- Montella-Manuel, S.; Pujol-Carrion, N.; de la Torre-Ruiz, M.A. The Cell Wall Integrity Receptor Mtl1 Contributes to Articulate Autophagic Responses When Glucose Availability Is Compromised. J. Fungi 2021, 7, 903. [Google Scholar] [CrossRef] [PubMed]
- Bandhakavi, S.; Xie, H.; O’Callaghan, B.; Sakurai, H.; Kim, D.H.; Griffin, T.J. Hsf1 activation inhibits rapamycin resistance and TOR signaling in yeast revealed by combined proteomic and genetic analysis. PLoS ONE 2008, 3, e1598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef] [PubMed]
- Roncero, C.; Valdivieso, M.H.; Ribas, J.C.; Duran, A. Isolation and characterization of Saccharomyces cerevisiae mutants resistant to Calcofluor white. J. Bacteriol. 1988, 170, 1950–1954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapteyn, J.C.; Ram, A.F.; Groos, E.M.; Kollar, R.; Montijn, R.C.; Van Den Ende, H.; Llobell, A.; Cabib, E.; Klis, F.M. Altered extent of cross-linking of beta1,6-glucosylated mannoproteins to chitin in Saccharomyces cerevisiae mutants with reduced cell wall beta1,3-glucan content. J. Bacteriol. 1997, 179, 6279–6284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesage, G.; Shapiro, J.; Specht, C.A.; Sdicu, A.M.; Menard, P.; Hussein, S.; Tong, A.H.; Boone, C.; Bussey, H. An interactional network of genes involved in chitin synthesis in Saccharomyces cerevisiae. BMC Genet. 2005, 6, 8. [Google Scholar] [CrossRef] [Green Version]
- Rockwell, N.C.; Wolfger, H.; Kuchler, K.; Thorner, J. ABC transporter Pdr10 regulates the membrane microenvironment of Pdr12 in Saccharomyces cerevisiae. J. Membr. Biol. 2009, 229, 27–52. [Google Scholar] [CrossRef] [Green Version]
- Queiroz, M.G.; Elsztein, C.; de Morais, M.A., Jr. The effects of the Ncw2 protein of Saccharomyces cerevisiae on the positioning of chitin in response to cell wall damage. Antonie Van Leeuwenhoek 2020, 113, 265–277. [Google Scholar] [CrossRef]
- Ishii, A.; Kurokawa, K.; Hotta, M.; Yoshizaki, S.; Kurita, M.; Koyama, A.; Nakano, A.; Kimura, Y. Role of Atg8 in the regulation of vacuolar membrane invagination. Sci. Rep. 2019, 9, 14828. [Google Scholar] [CrossRef] [Green Version]
- Cuevas-Bermúdez, A.; Garrido-Godino, A.; Navarro, F. A novel yeast chromatin-enriched fractions purification approach, yChEFs, for the chromatin-associated protein analysis used for chromatin-associated and RNA-dependent chromatin-associated proteome studies from Saccharomyces cerevisiae. Gene Rep. 2019, 16, 100450. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez-Santiago, F.; Cintas-Galán, M.; Martín-Expósito, M.; del Carmen Mota-Trujillo, M.; Cobo-Huesa, C.; Perez-Fernandez, J.; Navarro Gómez, F. A High-Copy Suppressor Screen Reveals a Broad Role of Prefoldin-like Bud27 in the TOR Signaling Pathway in Saccharomyces cerevisiae. Genes 2022, 13, 748. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050748
Gutiérrez-Santiago F, Cintas-Galán M, Martín-Expósito M, del Carmen Mota-Trujillo M, Cobo-Huesa C, Perez-Fernandez J, Navarro Gómez F. A High-Copy Suppressor Screen Reveals a Broad Role of Prefoldin-like Bud27 in the TOR Signaling Pathway in Saccharomyces cerevisiae. Genes. 2022; 13(5):748. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050748
Chicago/Turabian StyleGutiérrez-Santiago, Francisco, María Cintas-Galán, Manuel Martín-Expósito, Maria del Carmen Mota-Trujillo, Cristina Cobo-Huesa, Jorge Perez-Fernandez, and Francisco Navarro Gómez. 2022. "A High-Copy Suppressor Screen Reveals a Broad Role of Prefoldin-like Bud27 in the TOR Signaling Pathway in Saccharomyces cerevisiae" Genes 13, no. 5: 748. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050748