
Diurnal Variations in Physiological Characteristics, Photoassimilates, and Total Ascorbate in Early and Late Sown Indian Wheat Cultivars under Exposure to Elevated Ozone



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Design
2.2. Plant Sampling
2.3. Ascorbic Acid Content
2.4. Carbohydrates
2.5. Statistical Analysis
3. Results
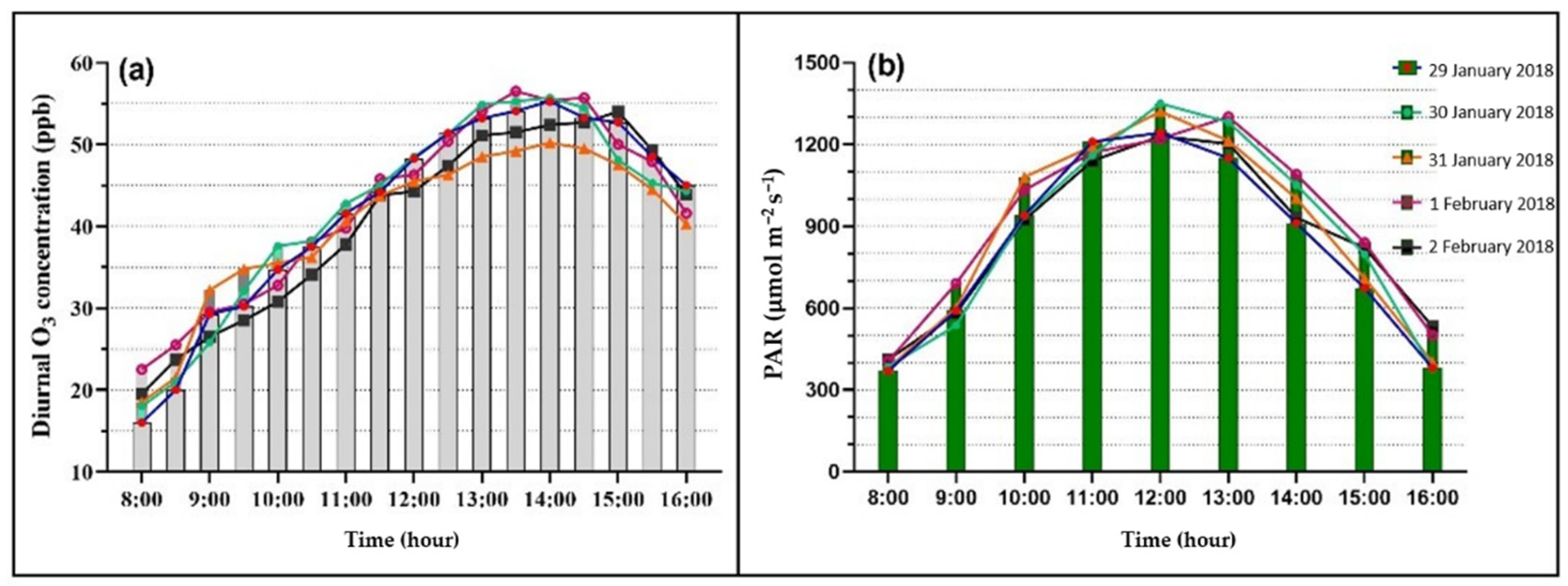
3.1. Diurnal Variations in Ambient O3 and PAR
3.2. Diurnal Gaseous Exchange and Chlorophyll Fluorescence
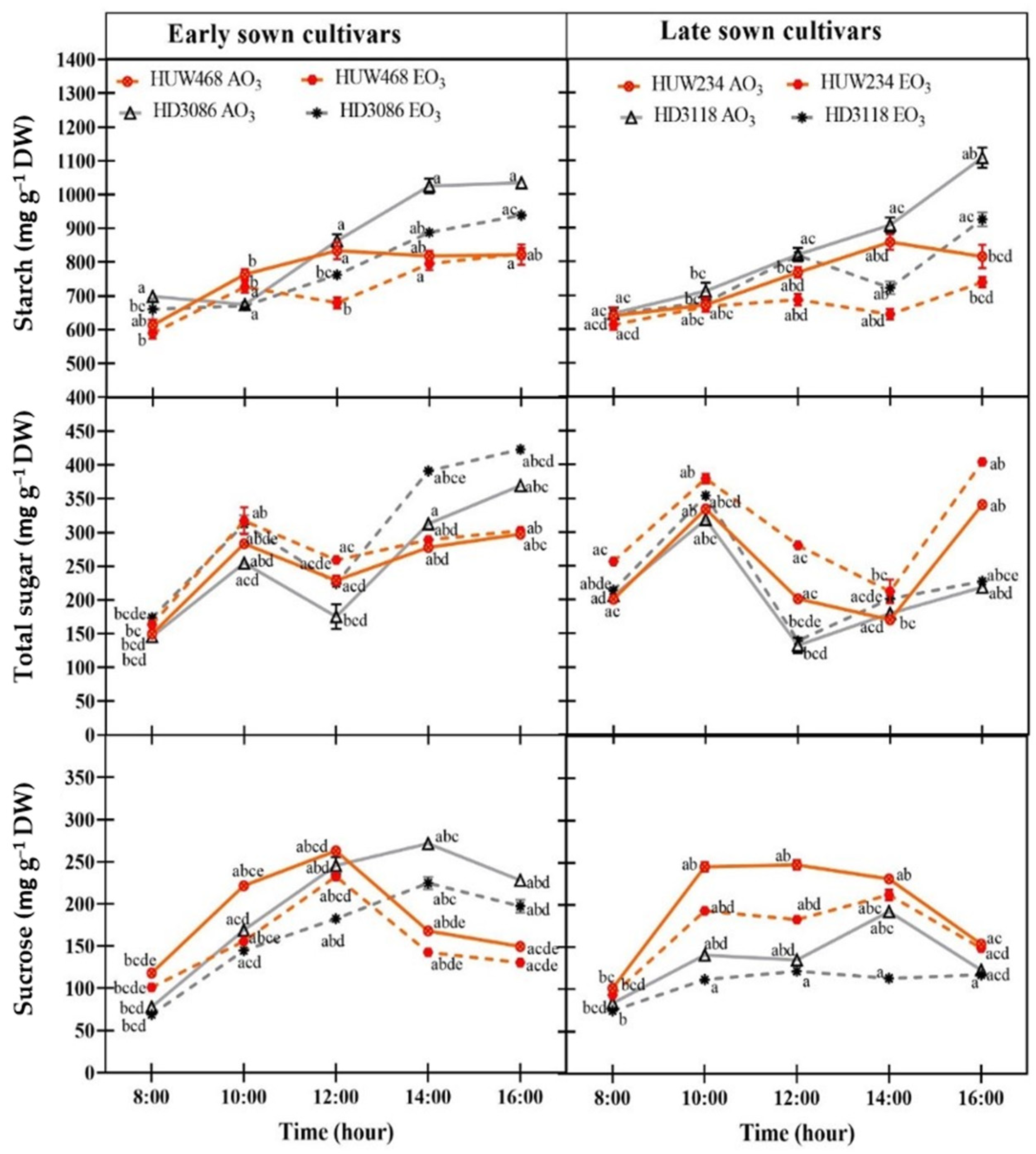
3.3. Diurnal Variations in Leaf Carbohydrate Pool
3.4. Diurnal Variations in Ascorbic Acid Content
3.5. Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feng, Z.; Pang, J.; Nouchi, I.; Kobayashi, K.; Yamakawa, T.; Zhu, J. Apoplastic ascorbate contributes to the differential ozone sensitivity in two varieties of winter wheat under fully open-air field conditions. Environ. Pollut. 2010, 158, 3539–3545. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Agrawal, M. Tropospheric Ozone and Its Impacts on Crop Plants: A Threat to Future Global Food Security; Springer: New York, NY, USA, 2018; ISBN 9783319718736. [Google Scholar]
- Mills, G.; Pleijel, H.; Malley, C.S.; Sinha, B.; Cooper, O.R.; Schultz, M.G.; Neufeld, H.S.; Simpson, D.; Sharps, K.; Feng, Z.; et al. Tropospheric ozone assessment report: Present-day tropospheric ozone distribution and trends relevant to vegetation. Elementa 2018, 6, 12. [Google Scholar] [CrossRef]
- Ren, J.; Hao, Y.; Simayi, M.; Shi, Y.; Xie, S. Spatiotemporal variation of surface ozone and its causes in Beijing, China since 2014. Atmos. Environ. 2021, 260, 118556. [Google Scholar] [CrossRef]
- Colls, J. Air Pollution, 2nd ed.; Bess Pub: London, UK, 2002. [Google Scholar]
- Wang, X.; Zheng, Q.; Feng, Z.; Xie, J.; Feng, Z.; Ouyang, Z.; Manning, W.J. Comparison of a diurnal vs. steady-state ozone exposure profile on growth and yield of oilseed rape (Brassica napus L.) in open-top chambers in the Yangtze Delta, China. Environ. Pollut. 2008, 156, 449–453. [Google Scholar] [CrossRef]
- Gaudel, A.; Cooper, O.R.; Ancellet, G.; Barret, B.; Boynard, A.; Burrows, J.P.; Clerbaux, C.; Coheur, P.F.; Cuesta, J.; Cuevas, E.; et al. Tropospheric Ozone Assessment Report: Present-day distribution and trends of tropospheric ozone relevant to climate and global atmospheric chemistry model evaluation. Elementa 2018, 6, 39. [Google Scholar] [CrossRef]
- Avnery, S.; Mauzerall, D.L.; Liu, J.; Horowitz, L.W. Global crop yield reductions due to surface ozone exposure: 2. Year 2030 potential crop production losses and economic damage under two scenarios of O3 pollution. Atmos. Environ. 2011, 45, 2297–2309. [Google Scholar] [CrossRef]
- Lal, S.; Venkataramani, S.; Naja, M.; Kuniyal, J.C.; Mandal, T.K.; Bhuyan, P.K.; Kumari, K.M.; Tripathi, S.N.; Sarkar, U.; Das, T.; et al. Loss of crop yields in India due to surface ozone: An estimation based on a network of observations. Environ. Sci. Pollut. Res. 2017, 24, 20972–20981. [Google Scholar] [CrossRef] [PubMed]
- Yadav, D.S.; Agrawal, S.B.; Agrawal, M. Field Crops Research Ozone flux-effect relationship for early and late sown Indian wheat cultivars: Growth, biomass, and yield. Field Crop. Res. 2021, 263, 108076. [Google Scholar] [CrossRef]
- Mills, G.; Sharps, K.; Simpson, D.; Pleijel, H.; Broberg, M.; Uddling, J.; Jaramillo, F.; Davies, W.J.; Dentener, F.; Van den Berg, M.; et al. Ozone pollution will compromise efforts to increase global wheat production. Glob. Chang. Biol. 2018, 24, 3560–3574. [Google Scholar] [CrossRef]
- Hazrati, S.; Tahmasebi-Sarvestani, Z.; Modarres-Sanavy, S.A.M.; Mokhtassi-Bidgoli, A.; Nicola, S. Effects of water stress and light intensity on chlorophyll fluorescence parameters and pigments of Aloe vera L. Plant Physiol. Biochem. 2016, 106, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Saibo, N.J.M.; Lourenço, T.; Oliveira, M.M. Transcription factors and regulation of photosynthetic and related metabolism under environmental stresses. Ann. Bot. 2009, 103, 609–623. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, E.; Holopainen, T. Responses of two birch (Betula pendula Roth) clones to different ozone profiles with similar AOT40 exposure. Atmos. Environ. 2001, 35, 5245–5254. [Google Scholar] [CrossRef]
- Feng, Z.Z.; Yao, F.F.; Chen, Z.; Wang, X.K.; Zheng, Q.W.; Feng, Z.W. Response of gas exchange and yield components of field-grown Triticum aestivum L. to elevated ozone in China. Photosynthetica 2007, 45, 441–446. [Google Scholar] [CrossRef]
- Meyer, U.; Köllner, B.; Willenbrink, J.; Krause, G.H.M. Effects of different ozone exposure regimes on photosynthesis, assimilates and thousand grain weight in spring wheat. Agric. Ecosyst. Environ. 2000, 78, 49–55. [Google Scholar] [CrossRef]
- Meyer, U.; Köllner, B.; Willenbrink, J.; Krause, G.H.M. Physiological changes on agricultural crops induced by different ambient ozone exposure regimes. I. Effects on photosynthesis and assimilate allocation in spring wheat. New Phytol. 1997, 136, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Rai, R.; Agrawal, M.; Agrawal, S.B. Effects of ambient O3 on wheat during reproductive development: Gas exchange, photosynthetic pigments, chlorophyll fluorescence, and carbohydrates. Photosynthetica 2011, 49, 285–294. [Google Scholar] [CrossRef]
- Burkey, K.O.; Neufeld, H.S.; Souza, L.; Chappelka, A.H.; Davison, A.W. Seasonal profiles of leaf ascorbic acid content and redox state in ozone-sensitive wildflowers. Environ. Pollut. 2006, 143, 427–434. [Google Scholar] [CrossRef]
- Wang, L.; Pang, J.; Feng, Z.; Zhu, J.; Kobayashi, K. Diurnal variation of apoplastic ascorbate in winter wheat leaves in relation to ozone detoxification. Environ. Pollut. 2015, 207, 413–419. [Google Scholar] [CrossRef]
- Bellini, E.; De Tullio, M.C. Ascorbic acid and ozone: Novel perspectives to explain an elusive relationship. Plants 2019, 8, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burkey, K.O.; Wei, C.; Eason, G.; Ghosh, P.; Fenner, G.P. Antioxidant metabolite levels in ozone-sensitive and tolerant genotypes of snap bean. Physiol. Plant. 2000, 110, 195–200. [Google Scholar] [CrossRef] [Green Version]
- Geiger, D.R.; Servaites, J.C. Diurnal regulation of photosynthetic carbon metabolism in C3 plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1994, 45, 235–256. [Google Scholar] [CrossRef]
- Xu, H.; Chen, S.B.; Biswas, D.K.; Li, Y.G.; Jiang, G.M. Photosynthetic and yield responses of an old and a modern winter wheat cultivars to short-term ozone exposure. Photosynthetica 2009, 47, 247–254. [Google Scholar] [CrossRef]
- Singh, A.A.; Fatima, A.; Mishra, A.K.; Chaudhary, N.; Mukherjee, A.; Agrawal, M.; Agrawal, S.B. Assessment of ozone toxicity among 14 Indian wheat cultivars under field conditions: Growth and productivity. Environ. Monit. Assess. 2018, 190. [Google Scholar] [CrossRef] [PubMed]
- Yadav, D.S.; Rai, R.; Mishra, A.K.; Chaudhary, N.; Mukherjee, A.; Agrawal, S.B.; Agrawal, M. ROS production and its detoxification in early and late sown cultivars of wheat under future O 3 concentration. Sci. Total Environ. 2019, 659, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Keller, T.; Schwager, H. Air pollution and ascorbic acid. Eur. J. For. Pathol. 1977, 7, 338–350. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.; Hamilton, J.; Roberts, P.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Biochem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Yadav, D.S.; Mishra, A.K.; Rai, R.; Chaudhary, N.; Mukherjee, A.; Agrawal, S.B.; Agrawal, M. Responses of an old and a modern Indian wheat cultivar to future O3 level: Physiological, yield and grain quality parameters. Environ. Pollut. 2020, 259, 113939. [Google Scholar] [CrossRef]
- Vongcharoen, K.; Santanoo, S.; Banterng, P.; Jogloy, S.; Vorasoot, N.; Theerakulpisut, P. Diurnal and seasonal variations in the photosynthetic performance and chlorophyll fluorescence of cassava ‘rayong 9’ under irrigated and rainfed conditions. Photosynthetica 2019, 57, 268–285. [Google Scholar] [CrossRef] [Green Version]
- Betzelberger, A.M.; Yendrek, C.R.; Sun, J.; Leisner, C.P.; Nelson, R.L.; Ort, D.R.; Ainsworth, E.A. Ozone exposure response for U.S. soybean cultivars: Linear reductions in photosynthetic potential, biomass, and yield. Plant Physiol. 2012, 160, 1827–1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohotti, A.J.; Lawlor, D.W. Diurnal variation of photosynthesis and photoinhibition in tea: Effects of irradiance and nitrogen supply during growth in the field. J. Exp. Bot. 2002, 53, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.F.; Zheng, J.H.; Zhang, Y.Y.; Hu, W.H.; Mao, W.H.; Zhou, Y.H.; Yu, J.Q. Diurnal variations in gas exchange, chlorophyll fluorescence quenching and light allocation in soybean leaves: The cause for midday depression in CO2 assimilation. Sci. Hortic. 2006, 110, 214–218. [Google Scholar] [CrossRef]
- Santanoo, S.; Vongcharoen, K.; Banterng, P.; Vorasoot, N.; Jogloy, S.; Roytrakul, S.; Theerakulpisut, P. Seasonal Variation in Diurnal Photosynthesis and Chlorophyll Fluorescence of Four Genotypes of Cassava (Manihot esculenta Crantz) under Irrigation Conditions in a Tropical Savanna Climate. Agronomy 2019, 9, 206. [Google Scholar] [CrossRef] [Green Version]
- Calatayud, A.; Alvarado, J.W.; Barreno, E. Changes in chlorophyll a fluorescence, lipid peroxidation, and detoxificant system in potato plants grown under filtered and non-filtered air in open-top chambers. Photosynthetica 2001, 39, 507–513. [Google Scholar] [CrossRef]
- Guidi, L.; Degl’Innocenti, E. Ozone effects on high light-induced photoinhibition in Phaseolus vulgaris. Plant Sci. 2008, 174, 590–596. [Google Scholar] [CrossRef]
- Heath, R.L.; Lefohn, A.S.; Musselman, R.C. Temporal processes that contribute to nonlinearity in vegetation responses to ozone exposure and dose. Atmos. Environ. 2009, 43, 2919–2928. [Google Scholar] [CrossRef]
- Musselman, R.C.; Lefohn, A.S.; Massman, W.J.; Heath, R.L. A critical review and analysis of the use of exposure- and flux-based ozone indices for predicting vegetation effects. Atmos. Environ. 2006, 40, 1869–1888. [Google Scholar] [CrossRef]
- Fatima, A.; Singh, A.A.; Mukherjee, A.; Agrawal, M.; Agrawal, S.B. Ascorbic acid and thiols as potential biomarkers of ozone tolerance in tropical wheat cultivars. Ecotoxicol. Environ. Saf. 2019, 171, 701–708. [Google Scholar] [CrossRef]
- Wang, J.; Zeng, Q.; Zhu, J.; Chen, C.; Liu, G.; Tang, H. Apoplastic antioxidant enzyme responses to chronic free-air ozone exposure in two different ozone-sensitive wheat cultivars. Plant Physiol. Biochem. 2014, 82, 183–193. [Google Scholar] [CrossRef]
- Dolker, T.; Agrawal, M. Negative impacts of elevated ozone on dominant species of semi-natural grassland vegetation in Indo-Gangetic plain. Ecotoxicol. Environ. Saf. 2019, 182, 109404. [Google Scholar] [CrossRef]
- Severino, J.F.; Stich, K.; Soja, G. Ozone stress and antioxidant substances in Trifolium repens and Centaurea jacea leaves. Environ. Pollut. 2007, 146, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.M.; Britz, S.J. Ascorbate-dehydroascorbate level and redox status in leaflets of field-grown soybeans exposed to elevated ozone. Int. J. Plant Sci. 2001, 162, 119–125. [Google Scholar] [CrossRef]
- Ansari, N.; Yadav, D.S.; Agrawal, M.; Agrawal, S.B. The impact of elevated ozone on growth, secondary metabolites, production of reactive oxygen species and antioxidant response in an anti-diabetic plant Costus pictus. Funct. Plant Biol. 2021, 48, 597–610. [Google Scholar] [CrossRef]
- Shinohara, Y.; Suzuki, Y. Effects of Light and Nutritional Conditions on the Ascorbic Acid Content of Lettuce. J. Jpn. Soc. Hortic. Sci. 1981, 50, 239–246. [Google Scholar] [CrossRef]
- Sideris, C.P.; Young, H.Y.; Chun, H.H.Q. Diurnal changes and growth rates as associated with ascorbic acid, titratable acidity, carbohydrate and nitrogeous fractions in the leaves of Ananas comosus (L.) Merr. Plant Physiol. 1948, 23, 38–69. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, T.; Maruta, T.; Yoshimura, K.; Smirnoff, N. Biosynthesis and regulation of ascorbic acid in plants. In Antioxidants and Antioxidant Enzymes in Higher Plants; Springer: Cham, Switzerland, 2018; pp. 163–179. ISBN 9783319750880. [Google Scholar]
- Seminario, A.; Song, L.; Zulet, A.; Nguyen, H.T.; González, E.M.; Larrainzar, E. Drought stress causes a reduction in the biosynthesis of ascorbic acid in soybean plants. Front. Plant Sci. 2017, 8, 1042. [Google Scholar] [CrossRef] [Green Version]
- Dowdle, J.; Ishikawa, T.; Gatzek, S.; Rolinski, S.; Smirnoff, N. Two genes in Arabidopsis thaliana encoding GDP-L-galactose phosphorylase are required for ascorbate biosynthesis and seedling viability. Plant J. 2007, 52, 673–689. [Google Scholar] [CrossRef]
- Lavola, A.; Julkunen-Tiitto, R.; Paakkonen, E. Does ozone stress change the primary or secondary metabolites of birch (Betula pendula Roth.)? New Phytol. 1994, 126, 637–642. [Google Scholar] [CrossRef]
- Castagna, A.; Ranieri, A. Detoxification and repair process of ozone injury: From O3 uptake to gene expression adjustment. Environ. Pollut. 2009, 157, 1461–1469. [Google Scholar] [CrossRef] [PubMed]
- Friend, A.L.; Tomlinson, P.T. Mild ozone exposure alters 14C dynamics in foliage of Pinus taeda L. Tree Physiol. 1992, 11, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Gallie, D.R. Increasing tolerance to ozone by elevating foliar ascorbic acid confers greater protection against ozone than increasing avoidance. Plant Physiol. 2005, 138, 1673–1689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranieri, A.; D’Urso, G.; Nali, C.; Lorenzini, G.; Soldatini, G.F. Ozone stimulates apoplastic antioxidant systems in pumpkin leaves. Physiol. Plant. 1996, 97, 381–387. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Feng, Z.; Dai, L.; Shang, B.; Paoletti, E. Large variability in ambient ozone sensitivity across 19 ethylenediurea-treated Chinese cultivars of soybean is driven by total ascorbate. J. Environ. Sci. 2018, 64, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Höller, S.; Ueda, Y.; Wu, L.; Wang, Y.; Hajirezaei, M.R.; Ghaffari, M.R.; von Wirén, N.; Frei, M. Ascorbate biosynthesis and its involvement in stress tolerance and plant development in rice (Oryza sativa L.). Plant Mol. Biol. 2015, 88, 545–560. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yadav, D.S.; Jaiswal, B.; Agrawal, S.B.; Agrawal, M. Diurnal Variations in Physiological Characteristics, Photoassimilates, and Total Ascorbate in Early and Late Sown Indian Wheat Cultivars under Exposure to Elevated Ozone. Atmosphere 2021, 12, 1568. https://0-doi-org.brum.beds.ac.uk/10.3390/atmos12121568
Yadav DS, Jaiswal B, Agrawal SB, Agrawal M. Diurnal Variations in Physiological Characteristics, Photoassimilates, and Total Ascorbate in Early and Late Sown Indian Wheat Cultivars under Exposure to Elevated Ozone. Atmosphere. 2021; 12(12):1568. https://0-doi-org.brum.beds.ac.uk/10.3390/atmos12121568
Chicago/Turabian StyleYadav, Durgesh Singh, Bhavna Jaiswal, Shashi Bhushan Agrawal, and Madhoolika Agrawal. 2021. "Diurnal Variations in Physiological Characteristics, Photoassimilates, and Total Ascorbate in Early and Late Sown Indian Wheat Cultivars under Exposure to Elevated Ozone" Atmosphere 12, no. 12: 1568. https://0-doi-org.brum.beds.ac.uk/10.3390/atmos12121568