
The Optimal Strategies to Be Adopted in Controlling the Co-Circulation of COVID-19, Dengue and HIV: Insight from a Mathematical Model




Abstract
:1. Introduction
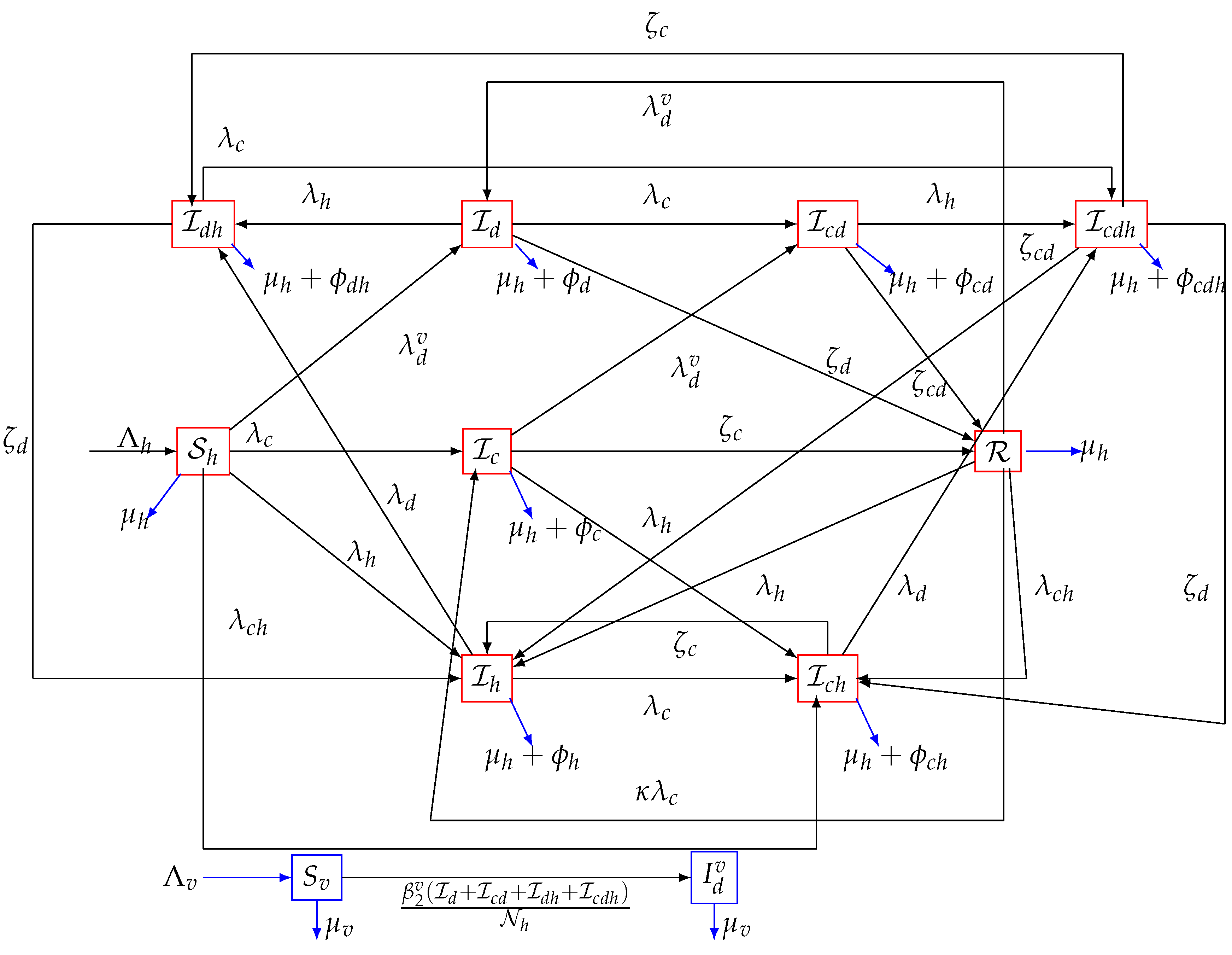
2. Model Formulation
3. Analysis of the Model
3.1. Non-Negativity of the Model Solutions
3.2. Boundedness of the Solution
3.3. The Basic Reproduction Number of the Model
3.4. Local Asymptotic Stability of the Disease Free Equilibrium (DFE) of the Model
3.5. Backward Bifurcation Analysis of the Model
3.6. Global Asymptotic Stability (GAS) of the Disease-Free Equilibrium for a Special Case
3.7. Global Asymptotic Stability (GAS) of the Endemic Equilibrium Point (EEP) of the Model (1)
4. Optimal Control Analysis
- : COVID-19 prevention control: this represents all the efforts towards COVID-19 prevention (and these include COVID-19 vaccination, face-mask usage in public, use of personal protective equipment (PPE) by health personnel, etc.);
- : Dengue prevention control: this represents all the efforts to prevent mosquito transmission of dengue disease. These include minimizing, as much as possible, the contacts between mosquitoes and humans, use of treated bed nets, and also receiving dengue vaccination;
- : HIV prevention control: This involves efforts to prevent HIV transmission via abstinence and effective condom use by sexually active individuals;
- : Control against co-infection: this involves combined efforts against all co-infections (COVID-19/dengue, COVID-19/HIV, dengue/HIV as well as COVID-19/dengue/HIV).
Existence
- (i)
- The admissible control set U is convex and closed.
- (ii)
- The state system is bounded by a linear function in the state and control variables.
- (iii)
- The integrand of the objective functional in (26) is convex with respect to the controls.
- (iv)
- The Lagrangian is no less than where .
- (i).
- The convexity of set U is obvious since it is 4D parallelepiped [50].
- (ii).
- The control system (24) can be expressed as a linear function of control variables , with the coefficients as functions of time and state variables:withwhere,In addition, it can be deduced thatwhere .
- (iii).
- The optimal system’s Lagrangian is given bywhich is also convex.
- (iv).
- There exists constants , and , such that the Lagrangian of the problem , , , .We now establish the bound on . We note that since , so that . Now,Hence,□
5. Numerical Simulations
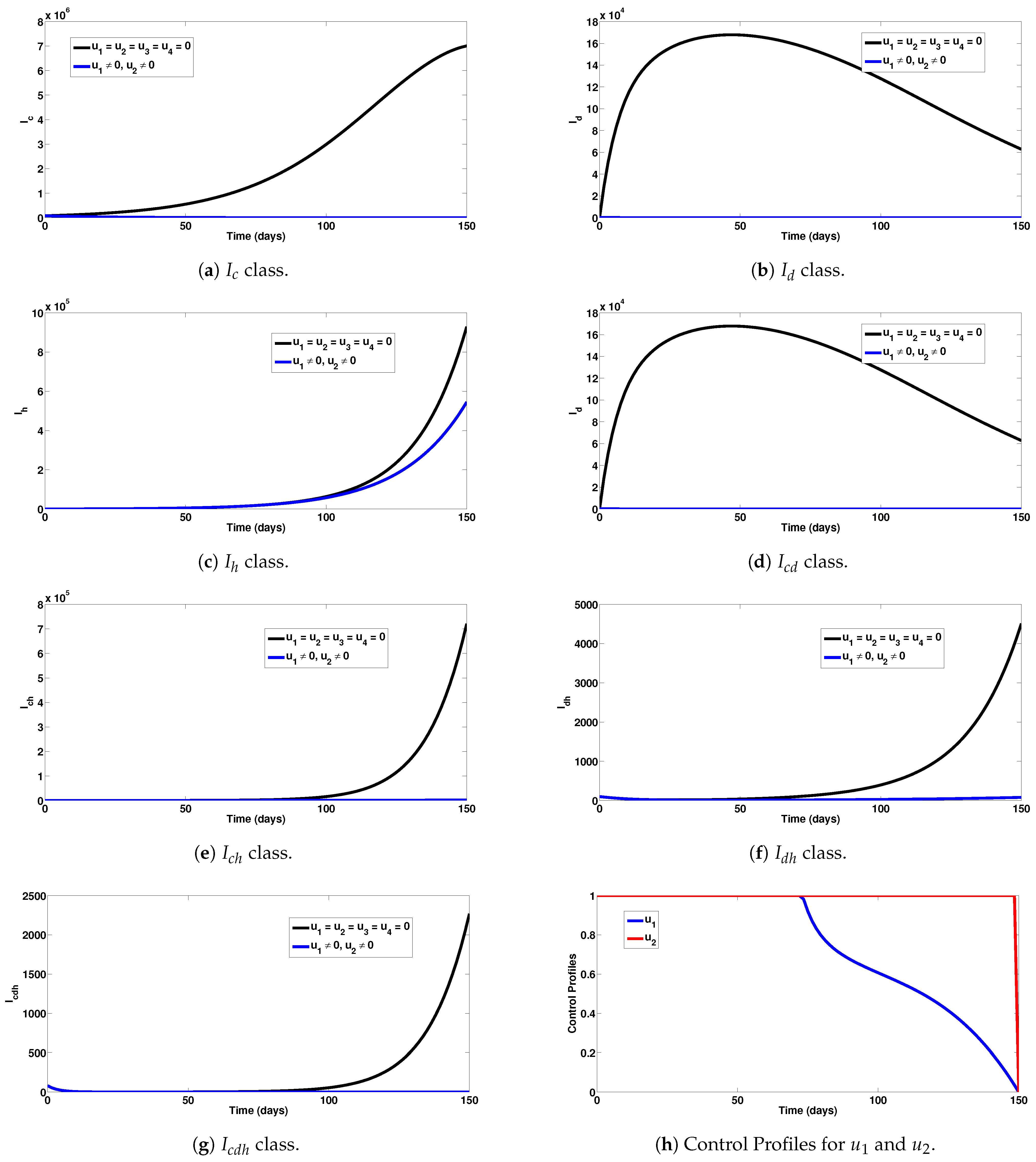
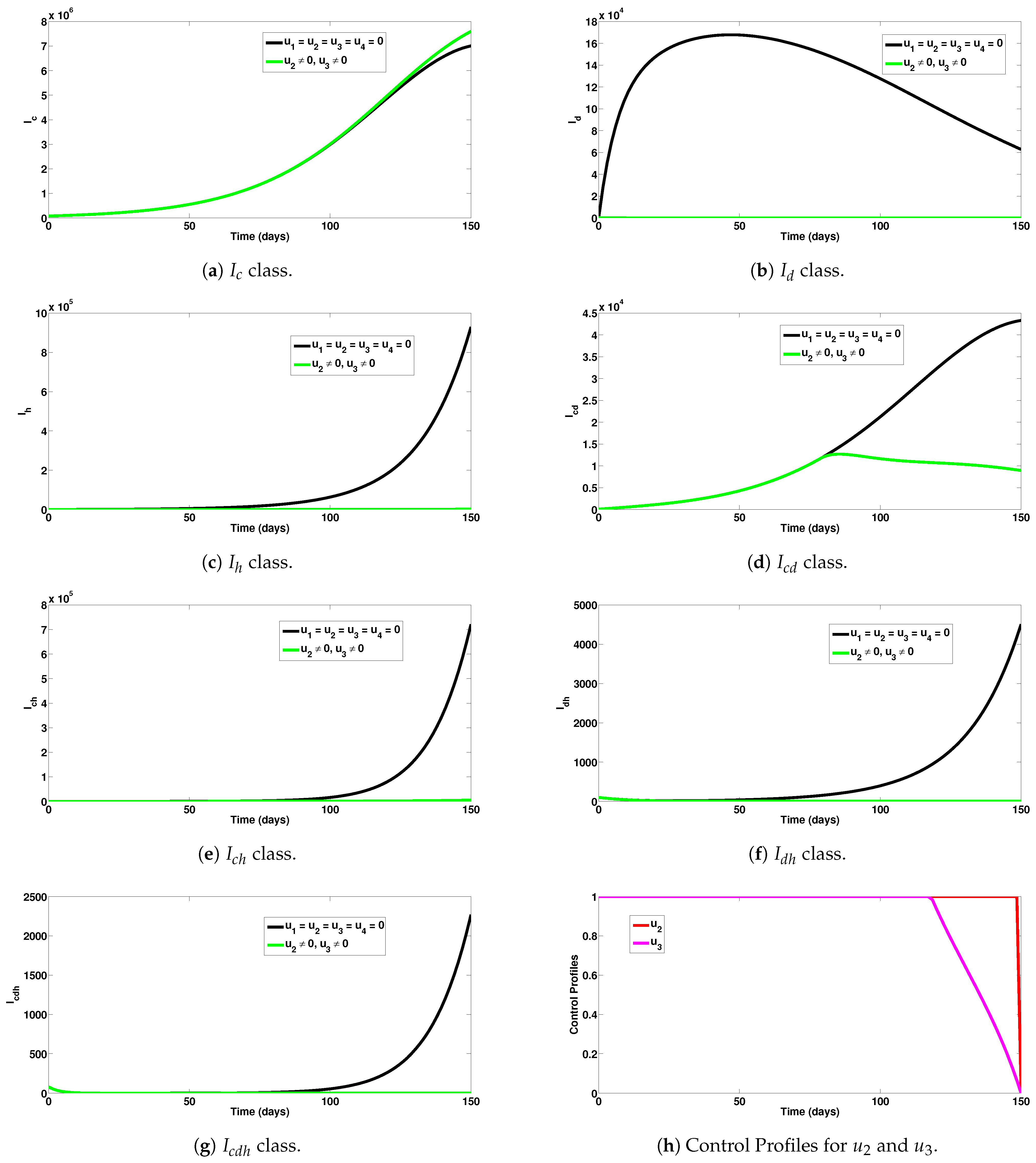
5.1. Strategy A: Assessment of COVID-19 and Dengue Combined Preventive Controls ()
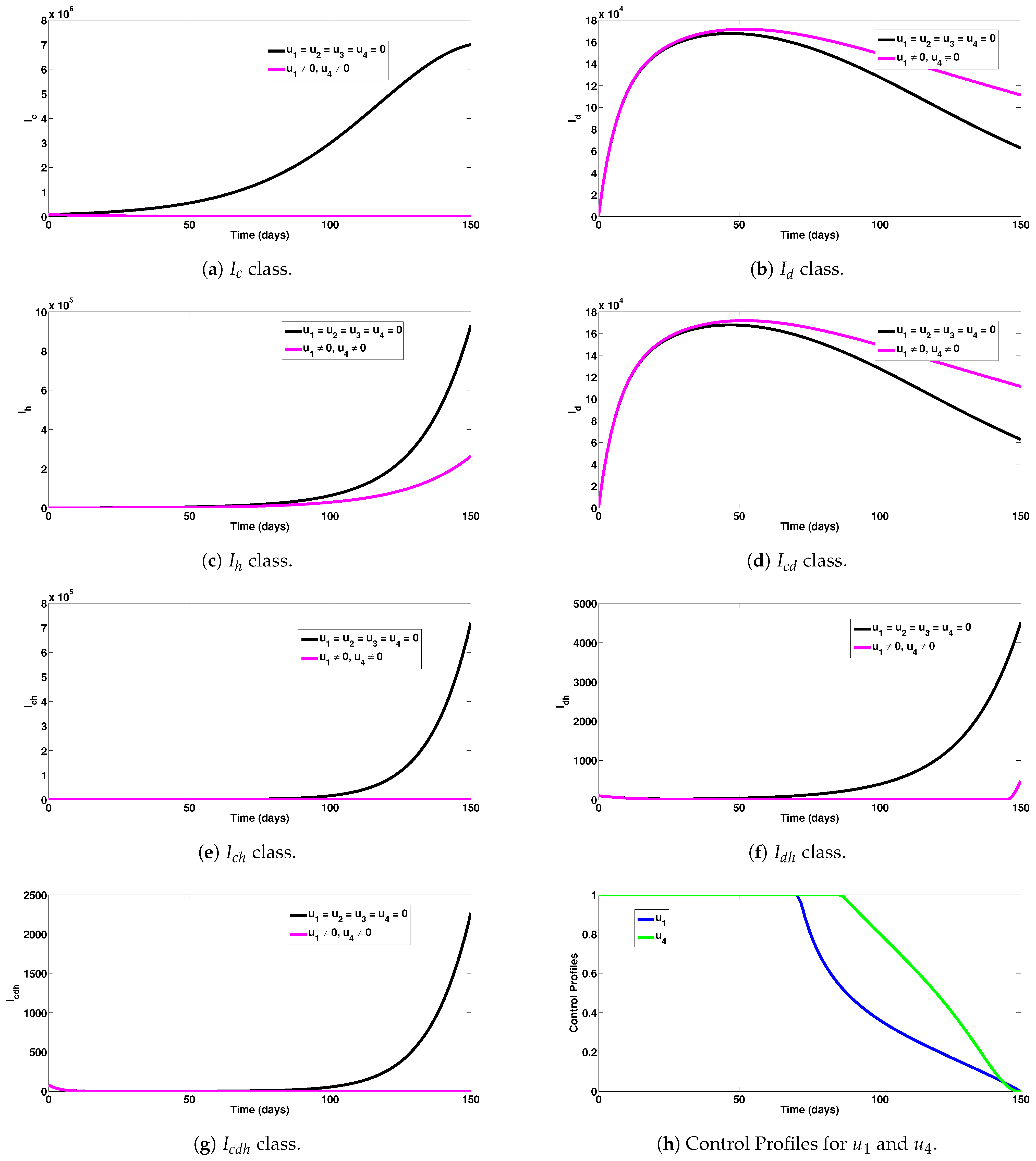
5.2. Strategy B: Assessment of COVID-19 and HIV Combined Preventive Controls ()
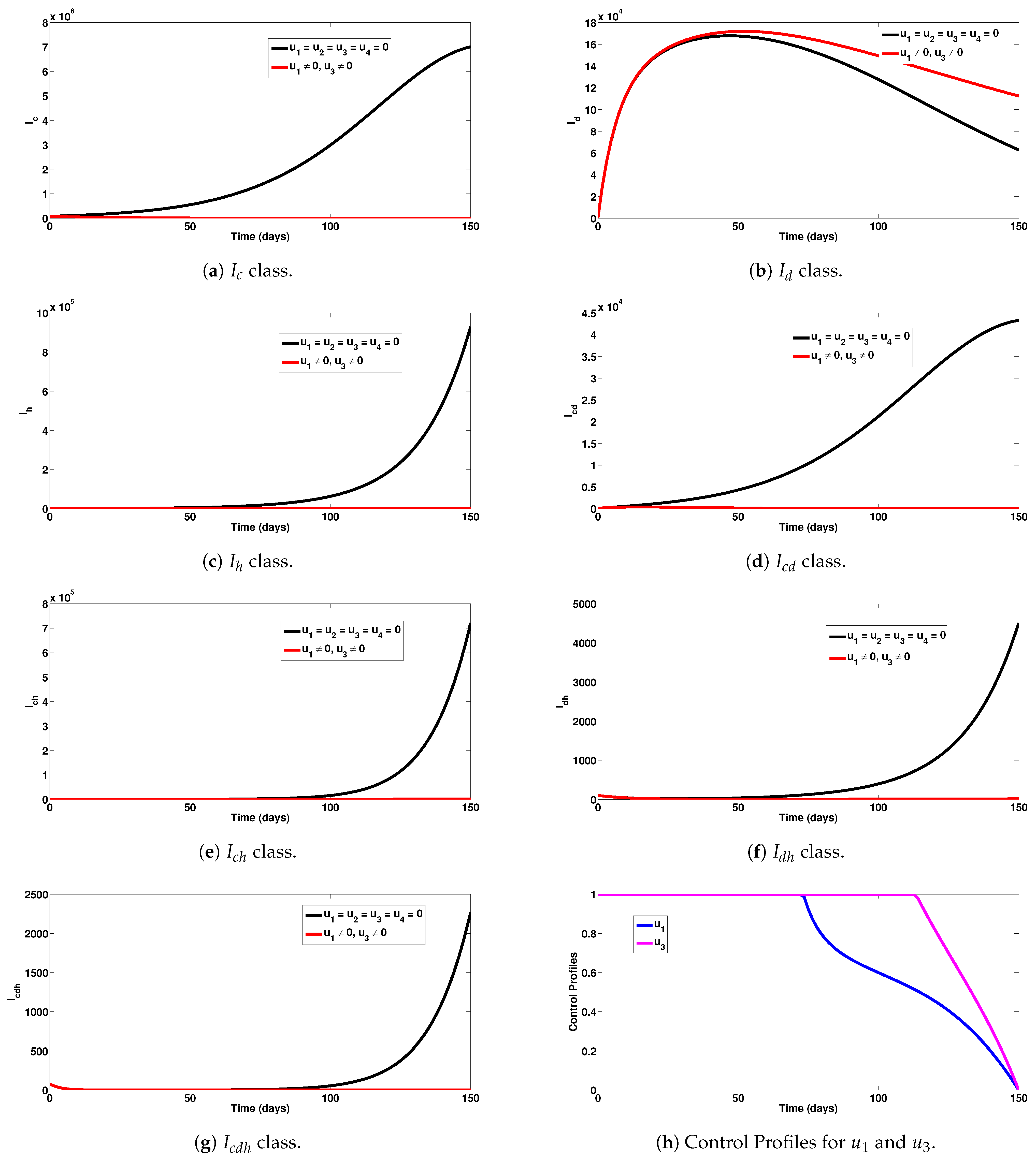
5.3. Strategy C: Assessment of COVID-19 and Co-Infection Combined Preventive Controls ()
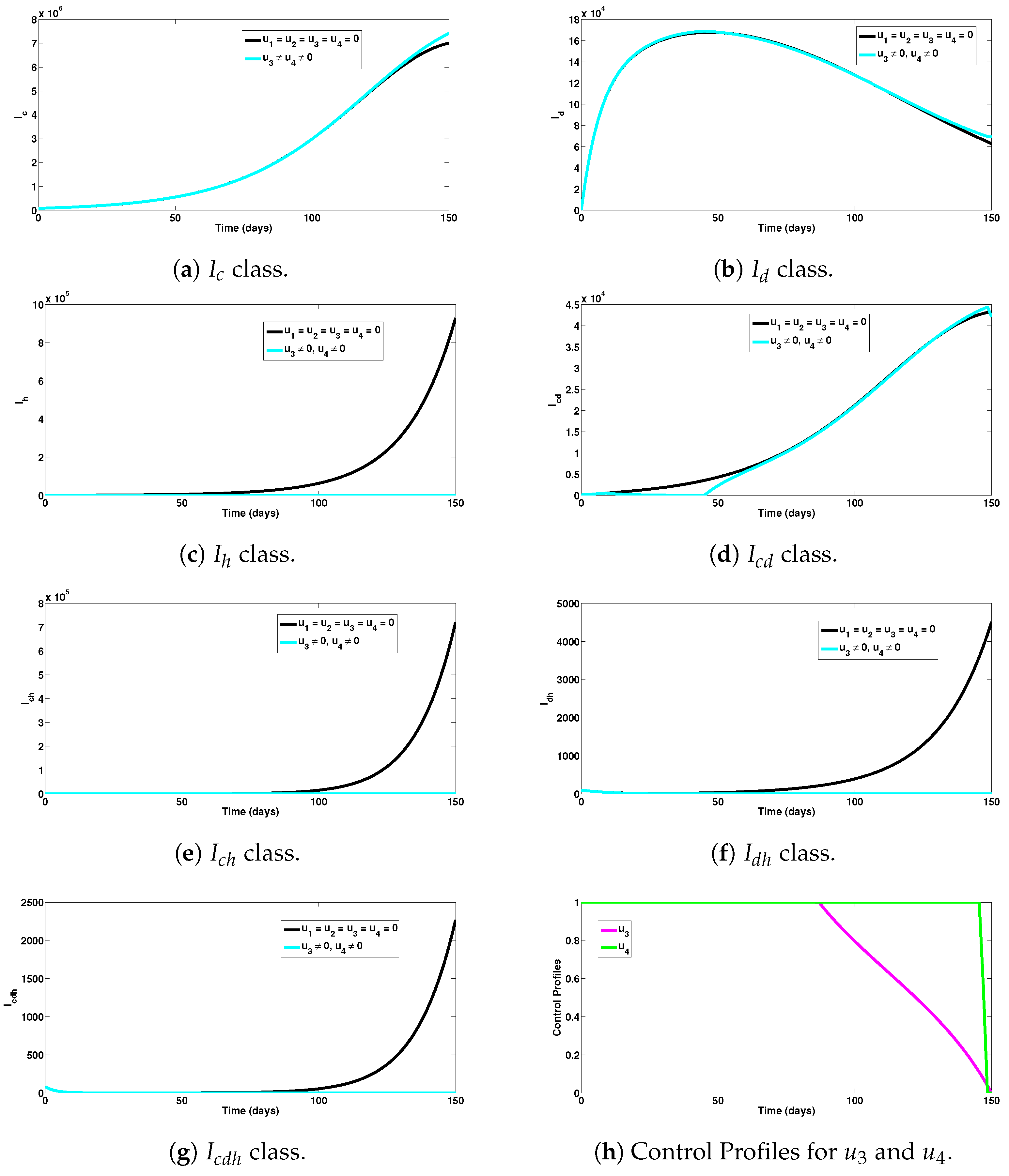
5.4. Strategy D: Assessment of Dengue and HIV Combined Preventive Controls ()
5.5. Strategy E: Assessment of HIV and Co-Infection Combined Preventive Controls ()
6. Conclusions
- (i)
- Upon implementation of the first intervention strategy (control against COVID-19 and dengue), it was observed that a significant number of single and dual infection cases were averted (as can be seen in Figure 2a–g).
- (ii)
- Under the COVID-19 and HIV prevention strategy, a good number of new single and dual infection cases were prevented (as can be observed in Figure 3a–g).
- (iii)
- Under the COVID-19 and co-infection prevention strategy, a remarkable number of new infections were averted (as presented in Figure 4a–g).
- (iv)
- Comparing all the intervention measures considered in this study, it is concluded that the strategies combining COVID-19/HIV averted the highest number of new infections. Thus, this strategy would be the most ideal and optimal to adopt for controlling the co-spread of COVID-19, dengue, and HIV.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Center-Manifold-Theory
- (A1):
- ; linearization of system (A1) in the neighbourhood of the equilibrium with φ evaluated at . The matrix A has zero eigenvalue and other eigenvalues have negative real parts;
- (A2):
- Matrix A has a right eigenvector ψ and a left eigenvector ϖ (each corresponding to the zero eigenvalue).
- (i).
- , . When with , is locally asymptotically stable and there exists an unstable equilibrium; when , is unstable and there exists a locally asymptotically stable equilibrium;
- (ii).
- , . When with , is unstable; when , 0 is locally asymptotically stable equilibrium, and there exists an unstable equilibrium;
- (iii).
- , . When with , is unstable and there exists a locally asymptotically stable equilibrium; when , is stable and an unstable equilibrium appears;
- (iv).
- , . When φ changes from negative to positive, changes its stability from stable to unstable. Correspondingly, an unstable equilibrium becomes locally asymptotically stable.
Appendix B. Pontryagin’s Maximum Principle
References
- Aguiar, D.; Lobrinus, J.A.; Schibler, M.; Fracasso, T.; Lardi, C. Inside the lungs of COVID-19 disease. Int. J. Leg. Med. 2020, 134, 1271–1274. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, S.; Liu, J.; Zhang, Z.; Wan, X.; Huang, B.; Chen, Y.; Zhang, Y. COVID-19: Immunopathogenesis and Immunotherapeutics. Signal Transduct. Target. Ther. 2020, 5, 128. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, Q.; Inchakalody, V.P.; Merhi, M.; Mestiri, S.; Taib, N.; Moustafa Abo El-Ella, D.; Bedhiafi, T.; Raza, A.; Al-Zaidan, L.; Mohsen, O.M.; et al. Emerging COVID-19 variants and their impact on SARS-CoV-2 diagnosis, therapeutics and vaccines. Ann. Med. 2022, 54, 524–540. [Google Scholar] [CrossRef] [PubMed]
- Dhama, K.; Nainu, F.; Frediansyah, A.; Yatoo, M.I.; Mohapatra, R.K.; Chakraborty, S.; Zhou, H.; Islam, M.R.; Mamada, S.S.; Kusuma, H.I.; et al. Global emerging Omicron variant of SARS-CoV-2: Impacts, challenges and strategies. J. Infect. Public Health 2022, 16, 4–14. [Google Scholar] [CrossRef]
- WHO. COVID-19 Webpage. 2020. Available online: https://covid19.who.int (accessed on 14 May 2023).
- Bosmuller, H.; Matter, M.; Fend, F.; Tzankov, A. The pulmonary pathology of COVID-19. Virchows Arch. 2021, 478, 137–150. [Google Scholar] [CrossRef]
- Sood, A.; Bedi, O. Histopathological and molecular links of COVID-19 with novel clinical manifestations for the management of coronavirus-like complications. Inflammopharmacology 2022, 30, 1219–1257. [Google Scholar] [CrossRef]
- Xu, L.; Liu, J.; Lu, M.; Yang, D.; Zheng, X. Liver injury during highly pathogenic human coronavirus infections. Liver Int. 2020, 40, 998–1004. [Google Scholar] [CrossRef]
- Nielsen, D.G. The relationship of interacting immunological components in dengue pathogenesis. Virol. J. 2009, 6, 211. [Google Scholar] [CrossRef]
- Halstead, S.B. Controversies in dengue pathogenesis. Paediatr. Int. Child Health 2012, 32 (Suppl. 1), 5–9. [Google Scholar] [CrossRef]
- Kok, B.H.; Lim, H.T.; Lim, C.P.; Lai, N.S.; Leow, C.Y.; Leow, C.H. Dengue virus infection? A review of Pathogenesis, Vaccines, Diagnosis and Therapy. Virus Res. 2022, 324, 199018. [Google Scholar] [CrossRef]
- Setiati, T.E.; Wagenaar, J.F.P.; de Kruif, M.; Mairuhu, A. Changing epidemiology of dengue haemorrhagic fever in Indonesia. Dengue Bull. 2006, 30, 1–4. [Google Scholar]
- Azhar, M.; Saeed, U.; Piracha, Z.Z.; Amjad, A.; Ahmed, A.; Batool, S.I.; Jabeen, M.; Fatima, A.; Noreen, F.; Uppal, R.; et al. SARS-CoV-2 related HIV, HBV, RSV, VZV, Enteric viruses, Influenza, DENV, S. Aureus TB Coinfections. Arch. Pathol. Clin. Res. 2021, 5, 26–33. [Google Scholar]
- WHO HIV/AIDS Webpage. Available online: https://www.who.int/news-room/fact-sheets/detail/hiv-aids (accessed on 26 May 2023).
- WHO HIV/AIDS Data 2021. Available online: https://www.who.int/data/gho/data/themes/hiv-aids (accessed on 26 May 2023).
- Hariyanto, T.I.; Rosalind, J.; Christian, K.; Kurniawan, A. Human immunodeficiency virus and mortality from coronavirus disease 2019: A systematic review and meta-analysis. South. Afr. J. HIV Med. 2021, 22, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, M.A.; Lynch, K.L.; Yun, C.; Glidden, D.V.; Peluso, M.J.; Henrich, T.J.; Gandhi, M.; Brown, L.B. SARS-CoV-2 seroprevalence, and IgG concentration and pseudovirus neutralising antibody titres after infection, compared by HIV status: A matched case-control observational study. Lancet HIV 2021, 8, E334–E341. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.aidsmap.com/about-hiv/covid-19-and-coronavirus-people-living-hiv (accessed on 26 May 2023).
- Masyeni, S.; Santoso, M.S.; Widyaningsih, P.D.; Asmara, D.W.; Nainu, F.; Harapan, H.; Sasmono, R.T. Serological cross-reaction and coinfection of dengue and COVID-19 in Asia: Experience from Indonesia. Int. J. Infect. Dis. 2021, 102, 152–154. [Google Scholar] [CrossRef]
- COVID-19: Massive Impact on Lower-Income Countries Threatens More Disease Outbreaks Gavi, the Vaccine Alliance. 2021. Available online: https://www.gavi.org/news/media-room/covid-19-massive-impact-lower-income-countries-threatens-more-disease-outbreaks (accessed on 7 April 2022).
- Impact of COVID-19 on Vaccine Supplies. UNICEF Supply Division. 2020. Available online: https://www.unicef.org/supply/stories/impact-covid-19-vaccine-supplies (accessed on 7 April 2022).
- Darling, K.E.; Diserens, E.A.; N’garambe, C.; Ansermet-Pagot, A.; Masserey, E.; Cavassini, M.; Bodenmann, P. A cross-sectional survey of attitudes to HIV risk and rapid HIV testing among clients of sex workers in Switzerland. Sex Transm. Infect. 2012, 88, 462–464. [Google Scholar] [CrossRef]
- Vezzani, D.; Velazquez, S.M.; Schweigmann, N. Seasonal pattern of abundance of Aedes Aegypti (Diptera: Culicidae) Buenos Aires City, Argentina. Mem. Inst. Oswaldo Cruz 2004, 99, 351–356. [Google Scholar] [CrossRef]
- Pan American Health Organization; World Health Organization. PAHO, PLISA Health Information Platform for the Americas. 2019. Available online: http://www.paho.org/data/index.php/en/ (accessed on 26 April 2022).
- López, M.S.; Jordan, D.I.; Blatter, E.; Walker, E.; Gómez, A.A.; Müller, G.V.; Mendicino, D.; Robert, M.A.; Estallo, E.L. Dengue emergence in the temperate Argentinian province of Santa Fe, 2009–2020. Sci. Data 2021, 8, 134. [Google Scholar] [CrossRef]
- Khan, M.A.; Shah, S.W.; Ullah, S.; Gómez-Aguilar, J.F. A dynamical model of asymptomatic carrier zika virus with optimal control strategies. Nonlinear Anal. Real World Appl. 2019, 50, 144–170. [Google Scholar] [CrossRef]
- Jan, R.; Khan, M.A.; Gómez-Aguilar, J.F. Asymptomatic carriers in transmission dynamics of dengue with control interventions. Optim. Control Appl. Methods 2020, 41, 430–447. [Google Scholar] [CrossRef]
- Ullah, S.; Khan, M.A.; Gómez-Aguilar, J.F. Mathematical formulation of hepatitis B virus with optimal control analysis. Optim. Control Appl. Methods 2019, 40, 529–544. [Google Scholar] [CrossRef]
- Omame, A.; Rwezaura, H.; Diagne, M.L.; Inyama, S.C.; Tchuenche, J.M. COVID-19 and dengue co-infection in Brazil: Optimal control and cost-effectiveness analysis. Eur. Phys. J. Plus 2021, 136, 1090. [Google Scholar] [CrossRef]
- Hezam, I.M. COVID-19 and Chikungunya: An optimal control model with consideration of social and environmental factors. J. Ambient. Intell. Humaniz. Comput. 2022. [Google Scholar] [CrossRef] [PubMed]
- Omame, A.; Abbas, M.; Onyenegecha, C.P. Backward bifurcation and optimal control in a co-infection model for SARS-CoV-2 and ZIKV. Results Phys. 2022, 37, 105481. [Google Scholar] [CrossRef]
- Omame, A.; Okuonghae, D. A co-infection model for oncogenic Human papillomavirus and tuberculosis with optimal control and cost-effectiveness analysis. Optim. Control Appl. Methods 2021, 42, 1081–1101. [Google Scholar] [CrossRef]
- Bonyah, E.; Khan, M.A.; Okosun, K.O.; Gómez-Aguilar, J.F. On the co-infection of dengue fever and Zika virus. Optim. Control Appl. Methods 2019, 40, 394–421. [Google Scholar] [CrossRef]
- Asamoah, J.K.K.; Yankson, E.; Okyere, E.; Sun, G.-Q.; Jin, Z.; Jan, R.; Fatmawati. Optimal control and cost-effectiveness analysis for dengue fever model with asymptomatic and partial immune individuals. Results Phys. 2021, 31, 104919. [Google Scholar] [CrossRef]
- Ojo, M.M.; Benson, T.O.; Peter, O.J.; Goufo, E.F.D. Nonlinear optimal control strategies for a mathematical model of COVID-19 and influenza co-infection. Physica A 2022, 607, 128173. [Google Scholar] [CrossRef]
- Abidemi, A.; Ackora-Prah, J.; Fatoyinbo, H.O.; Asamoah, J.K.K. Lyapunov stability analysis and optimization measures for a dengue disease transmission model. Phys. A Stat. Mech. Appl. 2022, 602, 127646. [Google Scholar] [CrossRef]
- Bi, K.; Chen, Y.; Wu, C.-H.; Ben-Arieh, D. A memetic algorithm for solving optimal control problems of Zika virus epidemic with equilibriums and backward bifurcation analysis. Commun. Nonlinear Sci. Numer. Simul. 2020, 84, 105176. [Google Scholar] [CrossRef]
- Chemaitelly, H.; Nagelkerke, N.; Ayoub, H.H.; Coyle, P.; Tang, P.; Yassine, H.M.; Al-Khatib, H.A.; Smarri, M.K.; Hasan, M.R.; Al-Kanaani, Z.; et al. Duration of immune protection of SARS-CoV-2 natural infection against reinfection. J. Travel Med. 2022, 29, Taac109. [Google Scholar] [CrossRef]
- Salvo, C.P.; Lella, N.D.; Lopez, F.S.; Hugo, J.; Zito, J.G.; Vilela, A. Coinfection Dengue Y SARS-CoV-2 en Paciente HIV Positivo. Medicina 2020, 80, 94–96. [Google Scholar]
- Omame, A.; Okuonghae, D.; Nwajeri, U.K.; Onyenegecha, C.P. A fractional-order multi-vaccination model for COVID-19 with non-singular kernel. Alex. Eng. J. 2022, 61, 6089–6104. [Google Scholar] [CrossRef]
- Available online: https://www.indexmundi.com/argentina/demographics_profile.html (accessed on 2 December 2021).
- Garba, S.M.; Gumel, A.B.; Abu Bakar, M.R. Backward bifurcations in dengue transmission dynamics. Math. Biosci. 2008, 215, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Okuneye, K.O.; Velasco-Hernandez, J.X.; Gumel, A.B. The “unholy” Chikungunya-Dengue-Zika Trinity: A Theoretical Analysis. J. Biol. Syst. 2017, 25, 545–585. [Google Scholar] [CrossRef]
- Omame, A.; Isah, M.E.; Abbas, M. An optimal control model for COVID-19, Zika, Dengue and Chikungunya co-dynamics with re-infection. Optim. Control Appl. Methods 2022, 44, 170–204. [Google Scholar] [CrossRef]
- Nwankwo, A.; Okuonghae, D. Mathematical analysis of the transmission dynamics of HIV syphilis Co-infection in the presence of treatment for syphilis. Bull. Math. Biol. 2018, 80, 437–492. [Google Scholar] [CrossRef]
- Van Den Driessche, P.; Watmough, J. Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission. Math. Biosci. 2002, 180, 29–48. [Google Scholar] [CrossRef]
- Castillo-Chavez, C.; Song, B. Dynamical models of tuberculosis and their applications. Math. Biosci. Eng. 2004, 2, 361–404. [Google Scholar] [CrossRef]
- LaSalle, J.P. The Stability of Dynamical Systems. In Regional Conferences Series in Applied Mathematics; SIAM: Philadelphia, PA, USA, 1976. [Google Scholar]
- Fleming, W.H.; Rishel, R.W. Deterministic and Stochastic Optimal Control; Springer: New York, NY, USA, 1975. [Google Scholar]
- Rector, C.R.; Chandra, S.; Dutta, J. Principles of Optimization Theory; Narosa Publishing House: New Delhi, India, 2005. [Google Scholar]
- Pontryagin, L.; Boltyanskii, V.; Gamkrelidze, R.; Mishchenko, E. The Mathematical Theory of Optimal Control Process 4; John Wiley & Sons: New York, NY, USA; London, UK, 1962. [Google Scholar]
- Lenhart, S.; Workman, J.T. Optimal Control Applied to Biological Models; Mathematical and Computational Biology Series; Chapman & Hall/CRC: Boca Raton, FL, USA, 2007. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Description | Value | Source |
|---|---|---|---|
| Contact rate for human–mosquito | |||
| spread of dengue | 0.60–0.70 day | [31] | |
| Dengue fever induced death rate | 0.05 day | [31] | |
| COVID-19 recovery rate | day | [40] | |
| COVID-19-induced death rate | 0.015 day | [40] | |
| Recruitment rate for humans | day | [41] | |
| Human natural death rate | day | [41] | |
| Recruitment rate for mosquitoes | 20,000 day | [42] | |
| Mosquito removal rate | day | [42] | |
| Dengue fever recovery rate | day | [43] | |
| COVID-19 transmission rate | day | [44] | |
| Contact rate for mosquito-human | |||
| spread of dengue | 0.3427 day | [44] | |
| HIV transmission rate | 0.3425 day | [45] | |
| COVID-19/HIV dual-transmission rate | 0.6 day | Assumed | |
| COVID-Dengue recovery rate | day | Assumed | |
| HIV induced death rate | day | [45] | |
| Co-infection death rate | day | Assumed | |
| Co-infection death rate | day | Assumed | |
| Co-infection death rate | day | Assumed | |
| Co-infection death rate | day | Assumed | |
| Re-infection rate for COVID-19 | day | [38] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Omame, A.; Raezah, A.A.; Diala, U.H.; Onuoha, C. The Optimal Strategies to Be Adopted in Controlling the Co-Circulation of COVID-19, Dengue and HIV: Insight from a Mathematical Model. Axioms 2023, 12, 773. https://0-doi-org.brum.beds.ac.uk/10.3390/axioms12080773
Omame A, Raezah AA, Diala UH, Onuoha C. The Optimal Strategies to Be Adopted in Controlling the Co-Circulation of COVID-19, Dengue and HIV: Insight from a Mathematical Model. Axioms. 2023; 12(8):773. https://0-doi-org.brum.beds.ac.uk/10.3390/axioms12080773
Chicago/Turabian StyleOmame, Andrew, Aeshah A. Raezah, Uchenna H. Diala, and Chinyere Onuoha. 2023. "The Optimal Strategies to Be Adopted in Controlling the Co-Circulation of COVID-19, Dengue and HIV: Insight from a Mathematical Model" Axioms 12, no. 8: 773. https://0-doi-org.brum.beds.ac.uk/10.3390/axioms12080773