
The Emerging Roles of Antioxidant Enzymes by Dietary Phytochemicals in Vascular Diseases


Department of Microbiology, School of Medicine, Kyung Hee University, Seoul 02447, Korea
*
Author to whom correspondence should be addressed.
Life 2021, 11(3), 199; https://0-doi-org.brum.beds.ac.uk/10.3390/life11030199
Submission received: 29 January 2021
/
Revised: 26 February 2021
/
Accepted: 26 February 2021
/
Published: 4 March 2021
(This article belongs to the Special Issue Frontiers in Vascular Biology)
Abstract
:Vascular diseases are major causes of death worldwide, causing pathologies including diabetes, atherosclerosis, and chronic obstructive pulmonary disease (COPD). Exposure of the vascular system to a variety of stressors and inducers has been implicated in the development of various human diseases, including chronic inflammatory diseases. In the vascular wall, antioxidant enzymes form the first line of defense against oxidative stress. Recently, extensive research into the beneficial effects of phytochemicals has been conducted; phytochemicals are found in commonly used spices, fruits, and herbs, and are used to prevent various pathologic conditions, including vascular diseases. The present review aims to highlight the effects of dietary phytochemicals role on antioxidant enzymes in vascular diseases.
1. Introduction
Vascular diseases are responsible for numerous deaths annually worldwide. The pathogenesis of vascular disease involves the activation of pro-inflammatory signaling pathways, expression of cytokines/chemokines, and elevated oxidative stress. Exposure to oxidative stress may directly injure the vasculature and induce vascular dysfunction by producing dysregulation of the immune response. Oxidative stress is caused by an imbalance between the production and accumulation of reactive oxygen species (ROS) and the capacity of antioxidant defense mechanisms favoring oxidants [1]. ROS generation may lead to cellular necrosis by damaging the mitochondria and stimulating pro-apoptotic signaling. ROS play a vital role in the progressive pathology of vascular diseases, including inflammatory responses, apoptosis, cell growth, and endothelial dysfunction [2,3]. Hence, strategies that target oxidative stress may have enormous therapeutic potential for the prevention of vascular diseases. The oxidative balance in the vasculature is tightly regulated by a wealth of pro- and antioxidant systems that contain antioxidant enzymes. A better understanding of oxidative stress and modulation of antioxidant enzymes is necessary for the broader use of pharmacological and regenerative therapies for vascular diseases. Here, we summarize the critical roles of antioxidant enzymes in vascular diseases and discuss the potential therapeutic roles of phytochemicals that target antioxidant enzymes.
2. Role of Antioxidant Enzymes
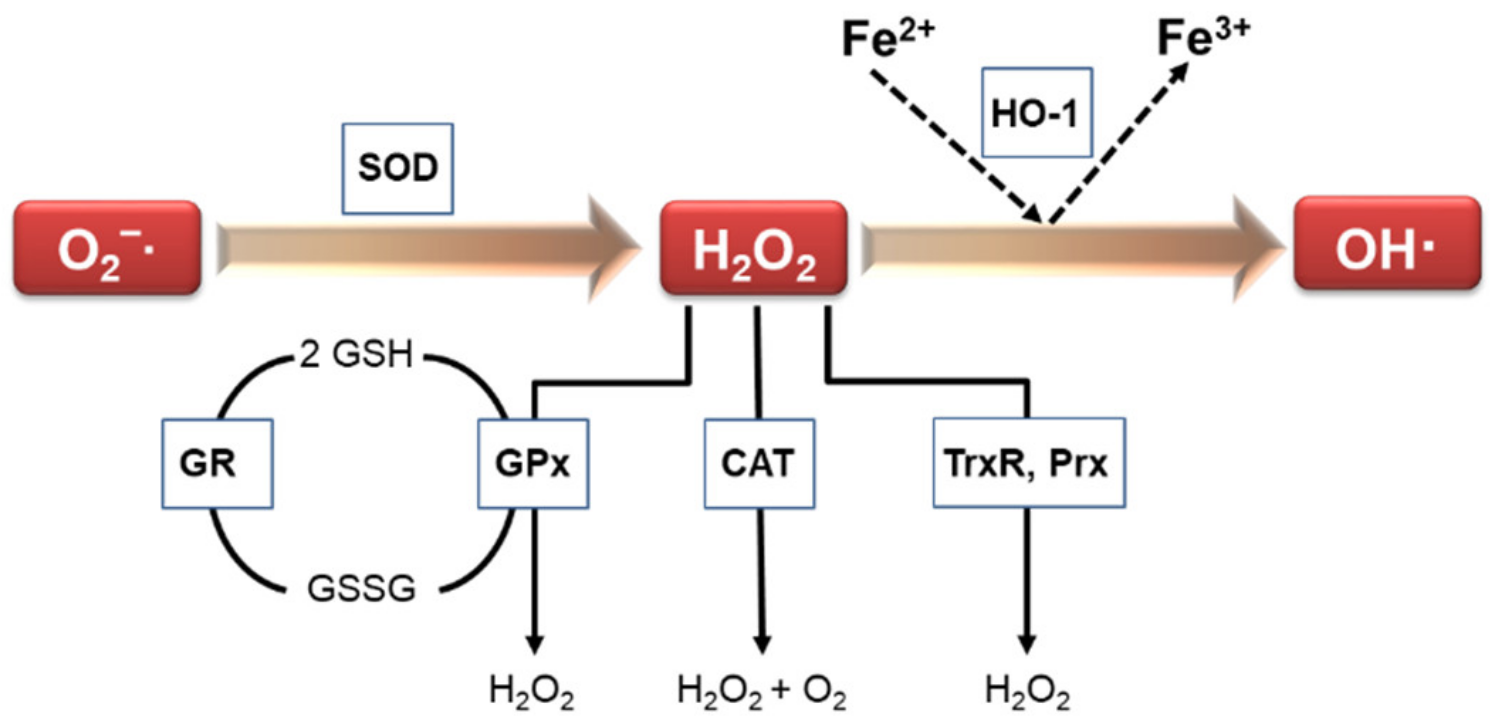
Escalated oxidative stress has been implicated in numerous human diseases, and is associated with disease severity. Vascular structures contain antioxidant defense systems that scavenge ROS [4]. Antioxidant enzymes are important as part of the cellular defense mechanism against free radical generation and in the prevention and repair of free-radical-produced molecular damage in a variety of situations [5]. Under physiological and pathophysiological conditions, various enzymes are responsible for modulating redox balance. Thus, antioxidant enzymes are vital for the maintaining the homeostasis of oxidants. Such antioxidant enzymes include catalase (CAT), glutathione reductase (GR), thioredoxin reductase (TrxR), heme oxygenase-1 (HO-1), superoxide dismutase (SOD), glutathione peroxidase (GPx), peroxiredoxin (Prx), paraoxonases (PON), and NAD(P)H: quinone oxidoreductase 1 (NQO1) (Figure 1).
2.1. Catalase (CAT)
CAT, a heme-containing tetrameric protein, is a normal component of cellular peroxisomes. CAT converts hydrogen peroxide (H2O2) into water and molecular oxygen and protects cells against excessive ROS production as well as inhibiting the accumulation of H2O2. CAT is universally expressed but is primarily located in the peroxisomes of all types of mammalian cells [6] and human vascular cells [7]. CAT is also universally expressed by neurons and glial cells in the central nervous system. Altered antioxidant enzyme activity, including decreased catalase activity in neutrophils, has been described [8,9]. Parastatidis et al. demonstrated that altered H2O2 signaling as a result of reduced CAT activity in the vasculature could be an early insult that leads to aortic dilatation [10]. Similarly, Wang et al. reported the anti-atherosclerotic effect of pterostilbene (a natural dietary compound) on experimental atherosclerosis via regulation of CAT/PTEN signaling [11].
2.2. Glutathione Reductase (GR)
Glutathione plays a fundamental antioxidant intracellular role [12] and is implicated in several metabolic processes. Oxidized glutathione molecules form disulfide bonds with each other until they are converted to reduced glutathione by GR. GR is a crucial enzyme in gene regulation, the maintenance of high rates of reduced glutathione (GSH)/oxidized disulfide (GSSG), intracellular signal transduction, elimination of free radicals and reactive oxygen species, and the preservation of intracellular redox status [13].
2.3. Thioredoxin Reductase (TrxR)
The thioredoxin system consists of thioredoxin (Trx), thioredoxin reductase (TrxR), and nicotinamide adenine dinucleotide phosphate (NADPH), which mainly regulates intracellular redox homeostasis [14]. The three mammalian genes encode different TrxR isoforms. In mice, Txnrd1 encodes cytosolic TrxR1, Txnrd2 encodes mitochondrial TrxR2 (also called TR3), and Txnrd3 encodes thioredoxin glutathione reductase (TGR), which is primarily expressed in the spermatids of the testis and seems to play a significant role in spermatogenesis [15,16].
2.4. Heme Oxygenase-1 (HO-1)
HO is the first, rate-limiting enzyme in the catalysis of cellular heme degradation and the production of carbon monoxide, biliverdin, and free iron [17]. Two functional HO isoforms have been identified in mammals: HO-1 and HO-2 [18,19]. HO-1 is ubiquitously distributed and highly inducible by a wide variety of inducers, including endotoxins, metals, oxidants, cytokines, and phytochemicals [20,21,22]. There is ample evidence to suggest that HO-1 can protect against vascular remodeling and atherogenesis [23].
2.5. Superoxide Dismutase (SOD)
SODs are widespread and are primary regulatory enzymes used by microorganisms to catalyze the conversion of superoxide into oxygen and H2O2 [24]. Mammals express three SODs isoforms: the cytoplasmic copper/zinc-dependent SOD (SOD1; Cu/ZnSOD is a 32 kDa homodimeric enzyme), the mitochondrial manganese-dependent SOD (SOD2; MnSOD is an 88 kDa homotetrameric enzyme), and the extracellular Cu/ZnSOD (SOD3; is also dependent upon copper and zinc, contains a signaling peptide sequence, and exists as a homotetramer of 135 kDa) [25,26]. Several studies have reported a relationship between SOD3 and pathological conditions that involve vascular dysfunction, such as diabetes and cardiovascular disease [26,27].
2.6. Glutathione Peroxidase (GPx)
The GPx family comprises three evolutionary groups arising from a Cys-containing ancestor that catalyzes the reduction of H2O2. Eight isoforms of GPx have been identified, of which five are selenoproteins (GPx1-4 and GPx6). The three selenium-independent GPx enzymes rely on thiol rather than selenol chemistry. Among the GPx enzymes, GPx1 is the most abundant and ubiquitous isoform, whereas GPx6 is found as a selenoprotein only in humans [28]. The anti-atherosclerotic properties of GPx-1 in endothelial cells [29] and an inhibitory potential against lipid peroxidation of GPx-4 in mice have been reported [30]. GPx2 was first found in gastrointestinal tissues, and GPx3 is mainly synthesized in the proximal convoluted tubule cells of the kidney.
2.7. Peroxiredoxin (Px)
Prxs are a family of thiol-dependent peroxidases that neutralize reactive oxygen/nitrogen species and protect against oxidative and inflammatory stress [31]. Prx enzymes are able to form disulfide linkages following oxidation, and return to their active form following reduction by an additional enzyme, thioredoxin [32]. Six mammalian Prx isozymes (Prx1-6) have been classified based on the mechanism and the number of cysteine residues involved during catalysis [33]. Six Prx isoforms are expressed in mammals: the cytosolic isoforms Prx1, Prx2, and Prx6; the mitochondrial isoform Prx3; the secreted isoform Prx4; and Prx5, which is localized in multiple organelles [34]. Schreibelt et al. suggested that vascular Prx1 functions as an endogenous defense mechanism [35]. Guo et al. found that Prx4 is an anti-atherogenic factor that suppresses oxidative damage and apoptosis [36].
2.8. Paraoxonase (PON)
There are three different isoforms of PON: PON1, PON2, and PON3, which have multifunctional roles in numerous biological pathways, including protection against oxidative damage [37]. Among these, increasing attention has been focused on the role of PON1, which is a key functional constituent of high-density lipoprotein (HDL) particles in various human diseases, including diabetes, cardiovascular disease, cancers, aging, and several neurological disorders [38]. PON2 is an intracellular enzyme with antioxidant effects in major vascular cells [39]. Mechanistically, PON3 seems similar to PON2 [40], and transgenic PON3 expression lowers atherosclerosis and adiposity [41].
2.9. NAD (P) H: Quinone Oxidoreductase 1 (NQO1)
NQO1 is a broadly distributed FAD-dependent flavoprotein that stimulates obligatory two-electron reductions of various exogenous and endogenous quinones, quinoneimines, nitroaromatics, and azo dyes [42]. These reductions decrease quinone levels, thereby diminishing the chances of producing reactive oxygen intermediates by redox cycling and for the attenuation of intracellular thiol pools. Several studies have shown that NQO1 has been implicated in the pathogenesis of several diseases [43,44,45,46].
3. Modulation of Antioxidant Enzyme Expression
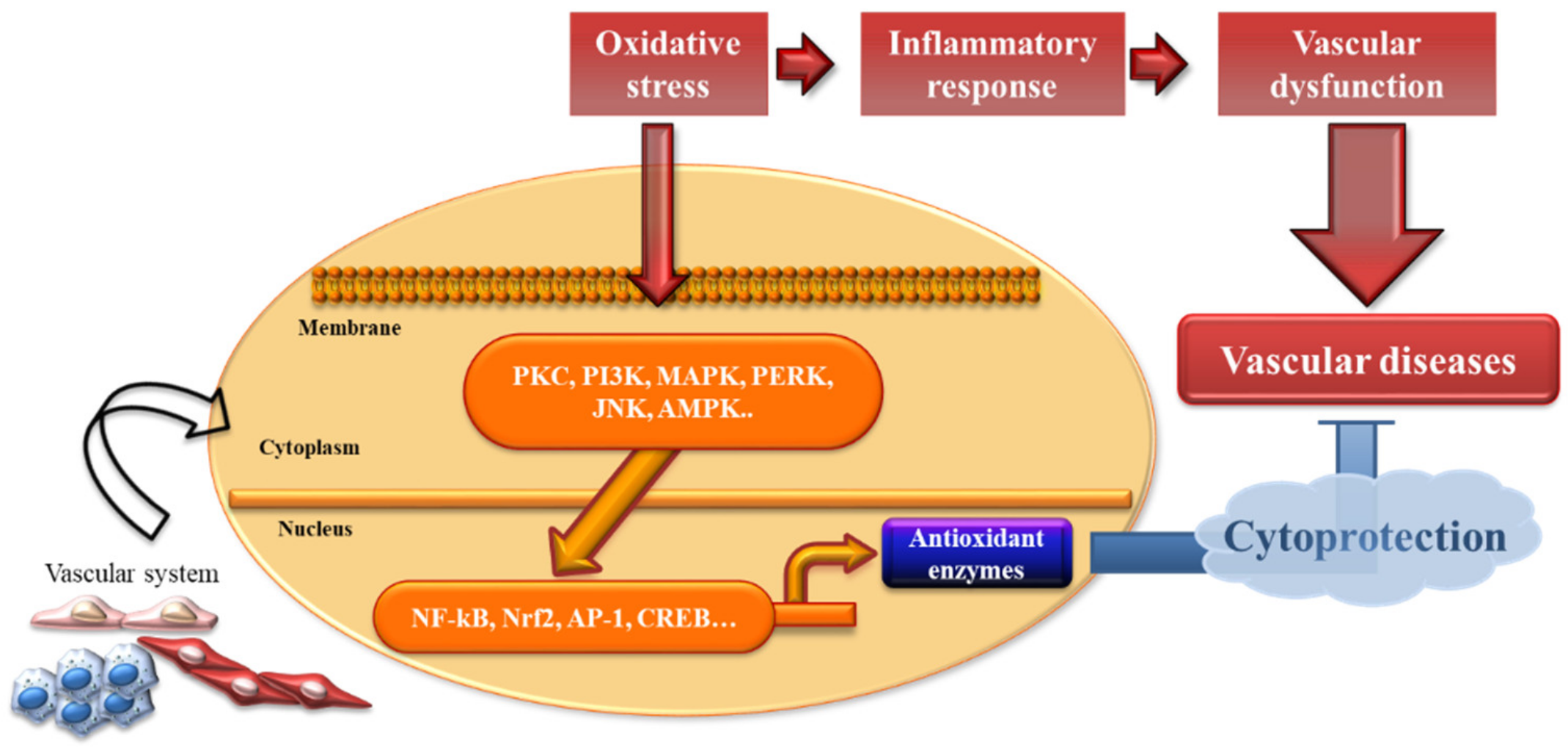
In general, oxidative stress is defined as an imbalance between pro-oxidant and antioxidant systems. Excess oxidative stress results in cellular damage due to the oxidation of innumerable essential host macromolecules. Phase II cytoprotective and detoxifying enzymes are responsible for serving as cellular guardians. To date, various signaling pathways, which mediate antioxidant enzyme regulation, have been identified, including protein kinase C (PKC), phosphatidylinositol 3-kinase (PI3K), ER-localized pancreatic endoplasmic reticulum kinase (PERK), mitogen-activated protein kinases (MAPKs), c-Jun NH2-terminal kinase (JNK), AMP-activated protein kinase (AMPK), nuclear factor E2-related factor 2 (Nrf2), activator protein-1 (AP-1), nuclear factor-κB (NF-κB), and cyclic adenosine monophosphate-response element-binding protein (CREB) [47,48,49,50,51,52,53,54,55,56] (Figure 2).
Numerous cytoprotective genes for detoxifying and antioxidative enzymes in the xenobiotic detoxification and antioxidative response pathways are induced upon exposure to oxidative stress. Nrf2 and the Kelch-like ECH-associated protein 1 (Keap1) system play a central role in antioxidant enzyme regulation [57,58]. Nrf2 is one of the best-characterized antioxidative transcription factors, with an oxidant/electrophile-sensor function. Under resting conditions, Keap1, a cytosolic repressor protein of Nrf2, binds to Nrf2 in the cytosol [59]. Upon exposure to oxidative stress, Nrf2 is separated from the Nrf2-Keap1 complex and translocates into the nucleus, where it induces transcriptional activation of cell defense genes [60]. After translocation to the nucleus, Nrf2 binds to the antioxidant or electrophile response element (ARE/EpRE) in the target gene promoter [61] as do a battery of antioxidant enzymes, including NQO1 [62], HO-1 [63], TR [64], and the Prx [65].
4. Effect of Antioxidant Enzyme Expression by Dietary Phytochemicals
Antioxidant enzymes have multifunctional roles in the development and progression of a variety of human diseases, including vascular diseases.
Dietary phytochemicals, which are abundant in fruits and vegetables, have been reported to promote health by enhancing antioxidant and anti-inflammatory abilities, as well as for their capacity for regulating a myriad of signaling mechanisms [66]. Modern scientific approaches have been utilized to identify and study various phytochemicals and have shown the potential value of phytochemicals in the field of pharmacology [67,68,69,70]. Numerous phytochemicals are recognized as inducers of antioxidant enzymes and, therefore, represent attractive candidates for use in healthcare (summarized in Table 1). However, although many studies have been conducted on the physiological activity of phytochemicals in humans, most studies have used animal subjects that have differences in applicable concentrations or metabolism. Therefore, there is an increasing demand for studies into the chemical instability, potential toxicological effects, pharmacokinetics, and pharmacodynamics of the target material to determine the appropriate concentration applicable to humans and to validate the physiological and pharmacological activity.
4.1. Anthocyanin
Fruits and vegetables are worthy sources of dietary phytochemicals, such as polyphenols, flavonoids, and carotenoids. In particular, anthocyanins, a class of flavonoids synthesized through the phenylpropanoid pathway, have pronounced antioxidant capacities in vitro and in vivo [102,103]. Anthocyanins stimulate optimal platelet function and exert antithrombotic effects [104]. They have protective effects on visual signal transduction and preclude age-related blindness by reducing the oxidative burden in mouse retinal pigment epithelium (RPE) cells [105]. Huang et al. determined the beneficial effect of anthocyanin on the increase in the levels of the antioxidant enzymes SOD, CAT, and GPx, which might have the potential to be applied to prevent eye diseases such as age-related macular degeneration (AMD) in human retinal capillary endothelial cells [71]. Another study showed that anthocyanins could ameliorate human retinal capillary endothelial function by decreasing ROS and increasing the enzyme activity of CAT and SOD and, therefore, might have the potential to avert the progression of diabetic retinopathy [72]. A recent study indicated that anthocyanins protected rodent endothelial function against high-glucose injury through antioxidant and vasodilatory mechanisms, so anthocyanins could be a promising hypotensive nutraceutical for diabetes [73].
4.2. Baicalein
Baicalein (5, 6, 7-trihydroxyflavone) is a natural flavonoid isolated from the root of Scutellaria baicalensis, a traditional Chinese herbal medicine commonly used for the treatment of bacterial infections [106]. Baicalein has innumerable biological properties, such as scavenging of relevant toxic ROS in rodent cardiomyocytes [107], and inhibition of tumor-induced angiogenesis [108]. Chao et al. found that baicalein could protect against retinal ischemia in a rat model because of its antioxidant and anti-apoptosis functions and ability to induce HO-1 expression [74]. Shi et al. reported that baicalein ameliorates pathological modifications such as pulmonary arterial remodeling via the MAPK/NF-κB/GPx/SOD pathway in rats [75].
4.3. Berberine
Berberine is a well-identified Chinese herbal medicine, extensively used in the treatment of a wide range of inflammatory diseases [109]. Berberine is present in the roots, rhizomes, and stem bulk of plants. Berberine has a variety of biological effects, such as antibacterial, anti-inflammatory, and antioxidant effects, and has been identified as a potential therapeutic candidate for diabetic nephropathy (DN) [110,111,112]. Recent studies have demonstrated the importance of the Nrf2 pathway and expression of the Nrf2-targeted antioxidative genes NQO-1 and HO-1 because of their anti-inflammatory role. These studies suggest that berberine may be beneficial in the development of new therapeutic strategies against inflammatory diseases, such as vascular diseases [76]. Yang et al. suggested that berberine restrained lipid uptake and stimulated cholesterol efflux through Nrf2/HO-1 activation, which resulted in the repression of foam cells and the progression of atherosclerotic plaques in a mouse model [77]. Paul et al. found that berberine has potential as a therapeutic candidate for the treatment of pathologies associated with diabetes in mice [78].
4.4. Curcumin
Curcumin (diferuloylmethane) is a polyphenol found in turmeric that has the capacity to treat several chronic diseases related to the cardiovascular system, nervous system, and inflammatory conditions [113,114,115]. Curcumin plays a protective role in the endothelium by inducing HO-1 in bovine aortic endothelial cells (ECs) [116]. Takano et al. found that curcumin suppresses vascular aging and inflammation, which are associated with the elevation of HO-1 in mice [79]. Similarly, Fleenor et al. provided evidence that dietary curcumin supplementation (0.2% in chow) alters two clinically important markers of arterial dysfunction with aging, including normalization of vascular superoxide production and oxidative stress, in a mouse model [80]. Xiao et al. found that curcumin protects against acute vascular inflammation through the activation of the HO-1/Nrf2/ARE/p38 MAPK signaling pathway in rabbits, and thus, may be useful in alleviating the vascular damage that occurs as a result of acute coronary events [81]. The inhibitory effect of curcumin supplementation in high fat diet-induced vascular dysfunction by increasing antioxidant enzyme activities, thereby restraining inflammation and oxidative damage in the vascular endothelium, has been reported in rats [82]. Several studies have reported that the cardioprotective effect of curcumin in a rodent model is associated with the attenuation of oxidant stress [83,84]. Supplementation with curcumin and carotenoids led to the anticipation of low-density lipoprotein (LDL) oxidation, which can be related to an increase in HDL levels and PON1 activity, thereby reducing these cardiovascular risk factors in diabetic rats [85].
4.5. Epigallocatechin Gallate (EGCG)
EGCG belongs to the catechin family of polyphenols, and is found in fruits, vegetables, chocolate, wine, and tea [117]. EGCG is the most abundant catechin and is associated with the majority of green tea intake-related health benefits [118]. After traumatic brain injury (TBI), immediate administration of EGCG suppresses edema formation and protect against TBP-induced oxidative stress through the blockage of NADPH oxidase activation in mice [86]. Zheng et al. demonstrated that EGCG provokes Nrf2 and HO-1 via the alteration of caveolae function related to caveolin-1 displacement [87].
4.6. Fisetin
Fisetin (3,3,4,7-tetrahydroxyflavone) is a small molecular flavonoid found in many fruits, especially strawberries, and vegetables [119]. Fisetin has been shown to have antioxidative effects mainly through activation of Nrf2/ARE in both rodent and human umbilical vein endothelial cells [88,120,121], and has also been shown to prevent cell proliferation and inflammation [122,123]. Another study revealed that fisetin ameliorated hyperhomocysteinemia (HHcy)-induced endothelial dysfunction and vascular dementia in rats, with the induction of antioxidant genes, such as SOD and CAT [89]. Dong et al. found that fisetin protects against cardiac hypertrophy both in vivo and in vitro, and that repression of oxidative stress is one of the critical underlying mechanisms [90].
4.7. Myricetin
Myricetin (3, 5, 7, 3, 4, 5-hexahydroxyflavone) is found in vegetables, fruits, teas, wines, and medicinal plants, and has both anti-inflammatory and antioxidant activities [124,125]. The anti-inflammatory mechanism of myricetin may involve its ability to obstruct the production of pro-inflammatory mediators via the inhibition of the NF-κB, STAT1, Nrf2, and HO-1 pathways [91]. Guo et al. demonstrated that myricetin has a protective effect against oxidative stress in choline-induced vascular dysfunction and liver injury in mice [92].
4.8. Quercetin
Quercetin is a bioactive plant flavonol-type flavonoid that is ubiquitous in vegetables and fruits [126]. Numerous human intervention studies have been conducted to evaluate the efficacy of quercetin consumption in reducing the risk of cardiovascular diseases [127,128,129,130,131]. Quercetin has prospective free radical scavenging properties, including inhibition of cancer proliferation [132], neuroprotection [133], renoprotection [134], and anti-thrombosis [135]. In in vitro models using rodent- and human-derived cells. Chis et al. found that quercetin administration, in conjunction with modest exercise training, decreased vascular complications and tissue injuries caused by diabetes in the rat aorta [93]. Likewise, vascular ROS formation and endothelial dysfunction were suppressed by dietary quercetin in HFD-fed ApoE−/− mice, with valuable effects on atherosclerotic plaque formation [94].
4.9. Resveratrol
Resveratrol (3,5,4-trihydroxy-trans-stilbene) is a polyphenol found in plants such as peanuts and different types of berries [136,137], which exerts antioxidant, anti-inflammatory, and neuroprotective effects [138,139,140]. Several studies have shown that resveratrol is beneficial for treating and preventing memory deficits in aged rats and has beneficial cardiovascular effects [94,140,141,142]. Resveratrol upregulates the endogenous antioxidant systems, such as the SOD enzymes, in endothelial cells and cardiac myoblasts, and further decreases ROS production [95,143]. A recent study reported that resveratrol ameliorates endothelial dysfunction, memory deficits, increased oxidative stress, inflammation, and impairment of neurotrophin expression in a rat model of vascular dementia [96], and therefore, may be beneficial in DM patients because of its vasculoprotective and neuroprotective effects. In streptozotocin-induced diabetes, resveratrol ameliorates endothelial dysfunction by reducing oxidative stress [97]. Supplementation with resveratrol diminished the presence of atherosclerotic lesions and periarterial fat deposition in ApoE-deficient (apo E−/−) mice [98].
4.10. Sulforaphane
Sulforaphane is a natural phytochemical found in cruciferous vegetables, such as broccoli [144]. Sulforaphane is a potent inducer of phase II antioxidant and detoxification enzymes with anticancer, antioxidant, and anti-inflammatory properties [145,146,147]. Sulforaphane plays a protective role in the injury of human cardiovascular cells by lysophosphatidylcholine by preventing the generation of intercellular ROS [99]. Shan et al. found that the action of sulforaphane depends on inflammatory injury in human vascular endothelial cells, mediated by p38 MAPK/JNK, as well as by inducing phase 2 enzymes. [100]. Pretreatment with sulforaphane induces antioxidant defenses in the rat brain and significantly mitigates functional and behavioral deficits after stroke [101].
5. Conclusions
Vascular diseases are the leading cause of death worldwide. The underlying mechanisms of vascular disease are many and complex, including excessive generation of reactive oxygen species, oxidative/nitrosative stress, inflammatory responses, and vascular dysfunction. Various aspects of vascular pathology reflect increased oxidative stress, leading to adverse outcomes in the disease state. Mounting evidence indicates that oxidative stress is a major pathological process leading to vascular disease. Increased levels of ROS have been linked to the development and progression of vascular diseases. Cytoprotective antioxidant enzymes are highly effective at diminishing toxicity following exposure to several stressors and inducers, such as ROS.
Phytochemicals are bioactive compounds that are abundantly distributed in fruits and vegetables. The last decade has seen an increase in interest in phytochemicals. Preclinical studies have revealed several beneficial vascular effects of resveratrol, curcumin, and berberine. The advantages appear to be predominantly dependent on the antioxidant, anti-inflammatory, and antithrombotic activities of the compounds. A robust correlation between specific classes of phytochemicals and modulation of antioxidant enzymes was observed.
According to Nelson et al., natural products such as curcumin/curcuminoids, which have various physiological activities, including anti-inflammatory and antioxidant functions, are pharmacologically incompatible in human. They exhibit chemical instability; have poor absorption, distribution, metabolism, excretion, and toxicology properties; and potential toxicological effects in some studies [148]. Therefore, medicinal chemistry research such as pharmacokinetic, pharmacodynamic, and biophysical orthogonal approaches, must be addressed when studying natural bioactive compounds in vitro or in vivo to acquire validated therapeutic efficacy.
This review focuses on understanding the role of antioxidant enzymes in the pathogenesis of vascular diseases. This review has shown that dietary phytochemicals can modulate antioxidant enzyme signaling pathways and, thus, have potential therapeutic value against various chronic diseases, including vascular diseases. A better understanding of the role of antioxidant enzymes by dietary phytochemicals will provide a broader understanding of the vascular system.
Author Contributions
Conceptualization, S.E.L. and Y.S.P.; writing—original draft preparation, S.E.L.; writing—review and editing, S.E.L. and Y.S.P.; funding acquisition, Y.S.P. Both authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Research Foundation of Korea (NRF), grant number NRF-2017M3A9E4044650 and NRF-2020R1A2C1011704.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This study was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIP) (Grant No.: NRF-2017M3A9E4044650 and NRF-2020R1A2C1011704).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kattoor, A.J.; Pothineni, N.V.K.; Palagiri, D.; Mehta, J.L. Oxidative Stress in Atherosclerosis. Curr. Atheroscler. Rep. 2017, 19, 42. [Google Scholar] [CrossRef]
- Panth, N.; Paudel, K.R.; Parajuli, K. Reactive Oxygen Species: A Key Hallmark of Cardiovascular Disease. Adv. Med. 2016, 2016, 9152732. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.C.; Sheen, J.M.; Hu, W.L.; Hung, Y.C. Polyphenols and Oxidative Stress in Atherosclerosis-Related Ischemic Heart Disease and Stroke. Oxid. Med. Cell Longev. 2017, 2017, 8526438. [Google Scholar] [CrossRef] [Green Version]
- Forstermann, U.; Xia, N.; Li, H.G. Roles of Vascular Oxidative Stress and Nitric Oxide in the Pathogenesis of Atherosclerosis. Circ. Res. 2017, 120, 713–735. [Google Scholar] [CrossRef]
- Kong, A.N.; Owuor, E.; Yu, R.; Hebbar, V.; Chen, C.; Hu, R.; Mandlekar, S. Induction of xenobiotic enzymes by the MAP kinase pathway and the antioxidant or electrophile response element (ARE/EpRE). Drug Metab. Rev. 2001, 33, 255–271. [Google Scholar] [CrossRef] [PubMed]
- Van den Bosch, H.; Schutgens, R.B.; Wanders, R.J.; Tager, J.M. Biochemistry of peroxisomes. Annu. Rev. Biochem. 1992, 61, 157–197. [Google Scholar] [CrossRef] [PubMed]
- Shingu, M.; Yoshioka, K.; Nobunaga, M.; Yoshida, K. Human vascular smooth muscle cells and endothelial cells lack catalase activity and are susceptible to hydrogen peroxide. Inflammation 1985, 9, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Dubick, M.A.; Keen, C.L.; DiSilvestro, R.A.; Eskelson, C.D.; Ireton, J.; Hunter, G.C. Antioxidant enzyme activity in human abdominal aortic aneurysmal and occlusive disease. Proc. Soc. Exp. Biol. Med. 1999, 220, 39–45. [Google Scholar] [CrossRef]
- Ramos-Mozo, P.; Madrigal-Matute, J.; Martinez-Pinna, R.; Blanco-Colio, L.M.; Lopez, J.A.; Camafeita, E.; Meilhac, O.; Michel, J.B.; Aparicio, C.; Vega de Ceniga, M.; et al. Proteomic analysis of polymorphonuclear neutrophils identifies catalase as a novel biomarker of abdominal aortic aneurysm: Potential implication of oxidative stress in abdominal aortic aneurysm progression. Arter. Thromb. Vasc. Biol. 2011, 31, 3011–3019. [Google Scholar] [CrossRef] [Green Version]
- Parastatidis, I.; Weiss, D.; Joseph, G.; Taylor, W.R. Overexpression of catalase in vascular smooth muscle cells prevents the formation of abdominal aortic aneurysms. Arter. Thromb. Vasc. Biol. 2013, 33, 2389–2396. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Wang, Y.R.; Chen, J.; Chen, Y.J.; Wang, Z.X.; Geng, M.; Xu, D.C.; Wang, Z.Y.; Li, J.H.; Xu, Z.D.; et al. Pterostilbene Attenuates Experimental Atherosclerosis through Restoring Catalase-Mediated Redox Balance in Vascular Smooth Muscle Cells. J. Agric. Food Chem. 2019, 67, 12752–12760. [Google Scholar] [CrossRef]
- Corso, C.R.; Acco, A. Glutathione system in animal model of solid tumors: From regulation to therapeutic target. Crit. Rev. Oncol. Hematol. 2018, 128, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Couto, N.; Wood, J.; Barber, J. The role of glutathione reductase and related enzymes on cellular redox homoeostasis network. Free Radic. Biol. Med. 2016, 95, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, A.; Lu, J. Thioredoxin and thioredoxin reductase: Current research with special reference to human disease. Biochem. Biophys Res. Commun. 2010, 396, 120–124. [Google Scholar] [CrossRef] [Green Version]
- Turanov, A.A.; Kehr, S.; Marino, S.M.; Yoo, M.H.; Carlson, B.A.; Hatfield, D.L.; Gladyshev, V.N. Mammalian thioredoxin reductase 1: Roles in redox homoeostasis and characterization of cellular targets. Biochem. J. 2010, 430, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Gerashchenko, M.V.; Su, D.; Gladyshev, V.N. CUG Start Codon Generates Thioredoxin/Glutathione Reductase Isoforms in Mouse Testes. J. Biol. Chem. 2010, 285, 4595–4602. [Google Scholar] [CrossRef] [Green Version]
- Ghattas, M.H.; Chuang, L.T.; Kappas, A.; Abraham, N.G. Protective effect of HO-1 against oxidative stress in human hepatoma cell line (HepG2) is independent of telomerase enzyme activity. Int. J. Biochem. Cell Biol. 2002, 34, 1619–1628. [Google Scholar] [CrossRef]
- McCoubrey, W.K.; Huang, T.J.; Maines, M.D. Isolation and characterization of a cDNA from the rat brain that encodes hemoprotein heme oxygenase-3. Eur. J. Biochem. 1997, 247, 725–732. [Google Scholar] [CrossRef]
- Abraham, N.G.; Kappas, A. Pharmacological and clinical aspects of heme oxygenase. Pharmacol. Rev. 2008, 60, 79–127. [Google Scholar] [CrossRef] [Green Version]
- Elbirt, K.K.; Whitmarsh, A.J.; Davis, R.J.; Bonkovsky, H.L. Mechanism of sodium arsenite-mediated induction of heme oxygenase-1 in hepatoma cells. Role of mitogen-activated protein kinases. J. Biol. Chem. 1998, 273, 8922–8931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pae, H.O.; Lee, Y.C.; Chung, H.T. Heme oxygenase-1 and carbon monoxide: Emerging therapeutic targets in inflammation and allergy. Recent Pat. Inflamm. Allergy Drug Discov. 2008, 2, 159–165. [Google Scholar] [CrossRef]
- Sahin, K.; Tuzcu, M.; Sahin, N.; Ali, S.; Kucuk, O. Nrf2/HO-1 signaling pathway may be the prime target for chemoprevention of cisplatin-induced nephrotoxicity by lycopene. Food Chem. Toxicol. 2010, 48, 2670–2674. [Google Scholar] [CrossRef]
- Stocker, R.; Perrella, M.A. Heme oxygenase-1—A novel drug target for atherosclerotic diseases? Circulation 2006, 114, 2178–2189. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Branicky, R.; Noe, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef]
- Yamakura, F.; Kawasaki, H. Post-translational modifications of superoxide dismutase. Biochim. Biophys. Acta 2010, 1804, 318–325. [Google Scholar] [CrossRef]
- Fukai, T.; Ushio-Fukai, M. Superoxide dismutases: Role in redox signaling, vascular function, and diseases. Antioxid. Redox Signal. 2011, 15, 1583–1606. [Google Scholar] [CrossRef] [Green Version]
- Fukai, T.; Folz, R.J.; Landmesser, U.; Harrison, D.G. Extracellular superoxide dismutase and cardiovascular disease. Cardiovasc. Res. 2002, 55, 239–249. [Google Scholar] [CrossRef]
- Kryukov, G.V.; Castellano, S.; Novoselov, S.V.; Lobanov, A.V.; Zehtab, O.; Guigo, R.; Gladyshev, V.N. Characterization of mammalian selenoproteomes. Science 2003, 300, 1439–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, A.H.; Kautz, O.; Fricke, K.; Zerr-Fouineau, M.; Demicheva, E.; Guldenzoph, B.; Bermejo, J.L.; Korff, T.; Hecker, M. Upregulation of glutathione peroxidase offsets stretch-induced proatherogenic gene expression in human endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1894–1901. [Google Scholar] [CrossRef]
- Guo, Z.; Ran, Q.; Roberts, L.J., 2nd; Zhou, L.; Richardson, A.; Sharan, C.; Wu, D.; Yang, H. Suppression of atherogenesis by overexpression of glutathione peroxidase-4 in apolipoprotein E-deficient mice. Free Radic. Biol. Med. 2008, 44, 343–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szeliga, M. Peroxiredoxins in Neurodegenerative Diseases. Antioxidants 2020, 9, 1203. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.G.; Kang, S.W.; Chang, T.S.; Jeong, W.; Kim, K. Peroxiredoxin, a novel family of peroxidases. IUBMB Life 2001, 52, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.R.; Zhu, H.; Danelisen, I. Role of Peroxiredoxins in Protecting Against Cardiovascular and Related Disorders. Cardiovasc. Toxicol. 2020, 20, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.G.; Woo, H.A.; Kil, I.S.; Bae, S.H. Peroxiredoxin Functions as a Peroxidase and a Regulator and Sensor of Local Peroxides. J. Biol. Chem. 2012, 287, 4403–4410. [Google Scholar] [CrossRef] [Green Version]
- Schreibelt, G.; van Horssen, J.; Haseloff, R.F.; Reijerkerk, A.; van der Pol, S.M.; Nieuwenhuizen, O.; Krause, E.; Blasig, I.E.; Dijkstra, C.D.; Ronken, E.; et al. Protective effects of peroxiredoxin-1 at the injured blood-brain barrier. Free Radic. Biol. Med. 2008, 45, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Yamada, S.; Tanimoto, A.; Ding, Y.; Wang, K.Y.; Shimajiri, S.; Murata, Y.; Kimura, S.; Tasaki, T.; Nabeshima, A.; et al. Overexpression of peroxiredoxin 4 attenuates atherosclerosis in apolipoprotein E knockout mice. Antioxid. Redox Signal. 2012, 17, 1362–1375. [Google Scholar] [CrossRef] [Green Version]
- Moya, C.; Manez, S. Paraoxonases: Metabolic role and pharmacological projection. Naunyn. Schmiedebergs Arch. Pharmacol. 2018, 391, 349–359. [Google Scholar] [CrossRef]
- Mahrooz, A. Pharmacological Interactions of Paraoxonase 1 (PON1): A HDL-Bound Antiatherogenic Enzyme. Curr. Clin. Pharm. 2016, 11, 259–264. [Google Scholar] [CrossRef]
- Horke, S.; Witte, I.; Wilgenbus, P.; Kruger, M.; Strand, D.; Forstermann, U. Paraoxonase-2 reduces oxidative stress in vascular cells and decreases endoplasmic reticulum stress-induced caspase activation. Circulation 2007, 115, 2055–2064. [Google Scholar] [CrossRef]
- Schweikert, E.M.; Devarajan, A.; Witte, I.; Wilgenbus, P.; Amort, J.; Forstermann, U.; Shabazian, A.; Grijalva, V.; Shih, D.M.; Farias-Eisner, R.; et al. PON3 is upregulated in cancer tissues and protects against mitochondrial superoxide-mediated cell death. Cell Death Differ. 2012, 19, 1549–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, D.M.; Xia, Y.R.; Wang, X.P.; Wang, S.S.; Bourquard, N.; Fogelman, A.M.; Lusis, A.J.; Reddy, S.T. Decreased obesity and atherosclerosis in human paraoxonase 3 transgenic mice. Circ. Res. 2007, 100, 1200–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinkova-Kostova, A.T.; Talalay, P. NAD(P)H:quinone acceptor oxidoreductase 1 (NQO1), a multifunctional antioxidant enzyme and exceptionally versatile cytoprotector. Arch. Biochem. Biophys. 2010, 501, 116–123. [Google Scholar] [CrossRef] [Green Version]
- SantaCruz, K.S.; Yazlovitskaya, E.; Collins, J.; Johnson, J.; DeCarli, C. Regional NAD(P)H: Quinone oxidoreductase activity in Alzheimer’s disease. Neurobiol. Aging 2004, 25, 63–69. [Google Scholar] [CrossRef]
- Dick, F.D.; de Palma, G.; Ahmadi, A.; Osborne, A.; Scott, N.W.; Prescott, G.J.; Bennett, J.; Semple, S.; Dick, S.; Mozzoni, P.; et al. Gene-environment interactions in parkinsonism and Parkinson’s disease: The Geoparkinson study. Occup. Environ. Med. 2007, 64, 673–680. [Google Scholar] [CrossRef] [Green Version]
- Van Muiswinkel, F.L.; de Vos, R.A.I.; Bol, J.G.J.M.; Andringa, G.; Steur, E.N.H.J.; Ross, D.; Siegel, D.; Drukarch, B. Expression of NAD(P)H: Quinone oxidoreductase in the normal and Parkinsonian substantia nigra. Neurobiol. Aging 2004, 25, 1253–1262. [Google Scholar] [CrossRef]
- Oh, C.J.; Park, S.; Kim, J.Y.; Kim, H.J.; Jeoung, N.H.; Choi, Y.K.; Go, Y.; Park, K.G.; Lee, I.K. Dimethylfumarate attenuates restenosis after acute vascular injury by cell-specific and Nrf2-dependent mechanisms. Redox Biol. 2014, 2, 855–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zrelli, H.; Matsuoka, M.; Kitazaki, S.; Zarrouk, M.; Miyazaki, H. Hydroxytyrosol reduces intracellular reactive oxygen species levels in vascular endothelial cells by upregulating catalase expression through the AMPK-FOXO3a pathway. Eur. J. Pharmacol. 2011, 660, 275–282. [Google Scholar] [CrossRef]
- Kumar, S.; Sud, N.; Fonseca, F.V.; Hou, Y.; Black, S.M. Shear stress stimulates nitric oxide signaling in pulmonary arterial endothelial cells via a reduction in catalase activity: Role of protein kinase C delta. Am. J. Physiol. Lung Cell Mol. Physiol. 2010, 298, L105–L116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Pan, S.; Berk, B.C. Glutaredoxin mediates Akt and eNOS activation by flow in a glutathione reductase-dependent manner. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 1283–1288. [Google Scholar] [CrossRef] [Green Version]
- Hojo, Y.; Saito, Y.; Tanimoto, T.; Hoefen, R.J.; Baines, C.P.; Yamamoto, K.; Haendeler, J.; Asmis, R.; Berk, B.C. Fluid shear stress attenuates hydrogen peroxide-induced c-Jun NH2-terminal kinase activation via a glutathione reductase-mediated mechanism. Circ. Res. 2002, 91, 712–718. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.E.; Son, G.W.; Park, H.R.; Jin, Y.H.; Park, C.S.; Park, Y.S. Induction of thioredoxin reductase 1 by crotonaldehyde as an adaptive mechanism in human endothelial cells. Mol. Cell. Toxicol. 2015, 11, 433–439. [Google Scholar] [CrossRef]
- Tsai, C.Y.; Wang, C.C.; Lai, T.Y.; Tsu, H.N.; Wang, C.H.; Liang, H.Y.; Kuo, W.W. Antioxidant effects of diallyl trisulfide on high glucose-induced apoptosis are mediated by the PI3K/Akt-dependent activation of Nrf2 in cardiomyocytes. Int. J. Cardiol. 2013, 168, 1286–1297. [Google Scholar] [CrossRef]
- Hu, R.; Wang, M.Q.; Ni, S.H.; Wang, M.; Liu, L.Y.; You, H.Y.; Wu, X.H.; Wang, Y.J.; Lu, L.; Wei, L.B. Salidroside ameliorates endothelial inflammation and oxidative stress by regulating the AMPK/NF-kappaB/NLRP3 signaling pathway in AGEs-induced HUVECs. Eur. J. Pharmacol. 2020, 867, 172797. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, P.; Feng, X.; Ma, C. Salidroside suppressing LPS-induced myocardial injury by inhibiting ROS-mediated PI3K/Akt/mTOR pathway in vitro and in vivo. J. Cell. Mol. Med. 2017, 21, 3178–3189. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Shi, S.; Cai, X.; Li, H.; Wang, L.; Li, H.; Xu, C. DR1 activation reduces the proliferation of vascular smooth muscle cells by JNK/c-Jun dependent increasing of Prx3. Mol. Cell. Biochem. 2018, 440, 157–165. [Google Scholar] [CrossRef]
- Arii, K.; Suehiro, T.; Ota, K.; Ikeda, Y.; Kumon, Y.; Osaki, F.; Inoue, M.; Inada, S.; Ogami, N.; Takata, H.; et al. Pitavastatin induces PON1 expression through p44/42 mitogen-activated protein kinase signaling cascade in Huh7 cells. Atherosclerosis 2009, 202, 439–445. [Google Scholar] [CrossRef] [Green Version]
- Randle, L.E.; Goldring, C.E.; Benson, C.A.; Metcalfe, P.N.; Kitteringham, N.R.; Park, B.K.; Williams, D.P. Investigation of the effect of a panel of model hepatotoxins on the Nrf2-Keap1 defence response pathway in CD-1 mice. Toxicology 2008, 243, 249–260. [Google Scholar] [CrossRef]
- Chapple, S.J.; Siow, R.C.; Mann, G.E. Crosstalk between Nrf2 and the proteasome: Therapeutic potential of Nrf2 inducers in vascular disease and aging. Int. J. Biochem. Cell Biol. 2012, 44, 1315–1320. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Tong, K.I.; Yamamoto, M. Molecular mechanism activating Nrf2-Keap1 pathway in regulation of adaptive response to electrophiles. Free Radic. Biol. Med. 2004, 36, 1208–1213. [Google Scholar] [CrossRef]
- Suzuki, T.; Yamamoto, M. Molecular basis of the Keap1-Nrf2 system. Free Radic. Biol. Med. 2015, 88, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Kensler, T.W.; Wakabayashi, N.; Biswal, S. Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 89–116. [Google Scholar] [CrossRef] [PubMed]
- Aghagolzadeh, P.; Radpour, R.; Bachtler, M.; van Goor, H.; Smith, E.R.; Lister, A.; Odermatt, A.; Feelisch, M.; Pasch, A. Hydrogen sulfide attenuates calcification of vascular smooth muscle cells via KEAP1/NRF2/NQO1 activation. Atherosclerosis 2017, 265, 78–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, E.S.; Yoon, J.J.; Han, B.H.; Jeong, D.H.; Lee, Y.J.; Kang, D.G.; Lee, H.S. Ligustilide attenuates vascular inflammation and activates Nrf2/HO-1 induction and, NO synthesis in HUVECs. Phytomedicine 2018, 38, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhang, Y.J.; Huang, X.; Xie, Y.; Qu, Y.; Long, H.Y.; Gu, N.; Jiang, W.M. Z-Ligustilide protects vascular endothelial cells from oxidative stress and rescues high fat diet-induced atherosclerosis by activating multiple NRF2 downstream genes. Atherosclerosis 2019, 284, 110–120. [Google Scholar] [CrossRef]
- Yang, Y.Y.; Li, X.Y.; Peng, L.Y.; An, L.; Sun, N.Y.; Hu, X.W.; Zhou, P.; Xu, Y.; Li, P.; Chen, J. Tanshindiol C inhibits oxidized low-density lipoprotein induced macrophage foam cell formation via a peroxiredoxin 1 dependent pathway. Biochim Biophys Acta. Mol. Basis Dis. 2018, 1864, 882–890. [Google Scholar] [CrossRef]
- Lu, B.Y.; Li, M.Q.; Yin, R. Phytochemical Content, Health Benefits, and Toxicology of Common Edible Flowers: A Review (2000–2015). Crit. Rev. Food Sci. Nutr. 2016, 56, S130–S148. [Google Scholar] [CrossRef]
- Tapsell, L.C.; Hemphill, I.; Cobiac, L.; Patch, C.S.; Sullivan, D.R.; Fenech, M.; Roodenrys, S.; Keogh, J.B.; Clifton, P.M.; Williams, P.G.; et al. Health benefits of herbs and spices: The past, the present, the future. Med. J. Aust. 2006, 185, S1–S24. [Google Scholar] [CrossRef] [Green Version]
- Triggiani, V.; Resta, F.; Guastamacchia, E.; Sabba, C.; Licchelli, B.; Ghiyasaldin, S.; Tafaro, E. Role of antioxidants, essential fatty acids, carnitine, vitamins, phytochemicals and trace elements in the treatment of diabetes mellitus and its chronic complications. Endocr Metab. Immune Disord. Drug Targets 2006, 6, 77–93. [Google Scholar] [CrossRef]
- Yin, R.; Kuo, H.C.; Hudlikar, R.; Sargsyan, D.; Li, S.; Wang, L.; Wu, R.; Kong, A.N. Gut microbiota, dietary phytochemicals and benefits to human health. Curr. Pharm. Rep. 2019, 5, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Vinayagam, R.; Xiao, J.; Xu, B. An insight into anti-diabetic properties of dietary phytochemicals. Phytochem. Rev. 2017, 16, 535–553. [Google Scholar] [CrossRef]
- Huang, W.Y.; Wu, H.; Li, D.J.; Song, J.F.; Xiao, Y.D.; Liu, C.Q.; Zhou, J.Z.; Sui, Z.Q. Protective Effects of Blueberry Anthocyanins against H2O2-Induced Oxidative Injuries in Human Retinal Pigment Epithelial Cells. J. Agric. Food Chem. 2018, 66, 1638–1648. [Google Scholar] [CrossRef]
- Huang, W.; Yan, Z.; Li, D.; Ma, Y.; Zhou, J.; Sui, Z. Antioxidant and Anti-Inflammatory Effects of Blueberry Anthocyanins on High Glucose-Induced Human Retinal Capillary Endothelial Cells. Oxid. Med. Cell Longev. 2018, 2018, 1862462. [Google Scholar] [CrossRef]
- Huang, W.; Hutabarat, R.P.; Chai, Z.; Zheng, T.; Zhang, W.; Li, D. Antioxidant Blueberry Anthocyanins Induce Vasodilation via PI3K/Akt Signaling Pathway in High-Glucose-Induced Human Umbilical Vein Endothelial Cells. Int. J. Mol. Sci. 2020, 21, 1575. [Google Scholar] [CrossRef] [Green Version]
- Chao, H.M.; Chuang, M.J.; Liu, J.H.; Liu, X.Q.; Ho, L.K.; Pan, W.H.T.; Zhang, X.M.; Liu, C.M.; Tsai, S.K.; Kong, C.W.; et al. Baicalein Protects Against Retinal Ischemia by Antioxidation, Antiapoptosis, Downregulation of HIF-1 alpha, VEGF, and MMP-9 and Upregulation of HO-1. J. Ocul. Pharmacol. Ther. 2013, 29, 539–549. [Google Scholar] [CrossRef] [Green Version]
- Shi, R.Z.; Wei, Z.H.; Zhu, D.Y.; Fu, N.J.; Wang, C.; Yin, S.; Liang, Y.Q.; Xing, J.F.; Wang, X.N.; Wang, Y. Baicalein attenuates monocrotaline-induced pulmonary arterial hypertension by inhibiting vascular remodeling in rats. Pulm. Pharmacol. Ther. 2018, 48, 124–135. [Google Scholar] [CrossRef]
- Mo, C.F.; Wang, L.; Zhang, J.; Numazawa, S.; Tang, H.; Tang, X.Q.; Han, X.J.; Li, J.H.; Yang, M.; Wang, Z.; et al. The Crosstalk Between Nrf2 and AMPK Signal Pathways Is Important for the Anti-Inflammatory Effect of Berberine in LPS-Stimulated Macrophages and Endotoxin-Shocked Mice. Antioxid. Redox Signal. 2014, 20, 574–588. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.J.; Liu, F.; Feng, N.; Ding, X.S.; Chen, Y.; Zhu, S.X.; Yang, L.C.; Feng, X.F. Berberine Attenuates Cholesterol Accumulation in Macrophage Foam Cells by Suppressing AP-1 Activity and Activation of the Nrf2/HO-1 Pathway. J. Cardiovasc. Pharmacol. 2020, 75, 45–53. [Google Scholar] [CrossRef]
- Paul, M.; Hemshekhar, M.; Kemparaju, K.; Girish, K.S. Berberine mitigates high glucose-potentiated platelet aggregation and apoptosis by modulating aldose reductase and NADPH oxidase activity. Free Radic. Biol. Med. 2019, 130, 196–205. [Google Scholar] [CrossRef]
- Takano, K.; Tatebe, J.; Washizawa, N.; Morita, T. Curcumin Inhibits Age-Related Vascular Changes in Aged Mice Fed a High-Fat Diet. Nutrients 2018, 10, 1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleenor, B.S.; Sindler, A.L.; Marvi, N.K.; Howell, K.L.; Zigler, M.L.; Yoshizawa, M.; Seals, D.R. Curcumin ameliorates arterial dysfunction and oxidative stress with aging. Exp. Gerontol. 2013, 48, 269–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Xia, J.; Wu, S.; Lv, Z.; Huang, S.; Huang, H.; Su, X.; Cheng, J.; Ke, Y. Curcumin Inhibits Acute Vascular Inflammation through the Activation of Heme Oxygenase-1. Oxid Med. Cell Longev. 2018, 2018, 3295807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, I.J.; Chen, C.W.; Tsai, S.Y.; Wang, P.Y.; Owaga, E.; Hsieh, R.H. Curcumin supplementation ameliorated vascular dysfunction and improved antioxidant status in rats fed a high-sucrose, high-fat diet. Appl. Physiol. Nutr. Me 2018, 43, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Ilyas, E.I.; Nur, B.M.; Laksono, S.P.; Bahtiar, A.; Estuningtyas, A.; Vitasyana, C.; Kusmana, D.; Suyatna, F.D.; Tadjudin, M.K.; Freisleben, H.J. Effects of Curcumin on Parameters of Myocardial Oxidative Stress and of Mitochondrial Glutathione Turnover in Reoxygenation after 60 Minutes of Hypoxia in Isolated Perfused Working Guinea Pig Hearts. Adv. Pharmacol Sci. 2016, 2016, 6173648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Salazar, A.; Molina-Jijon, E.; Correa, F.; Zarco-Marquez, G.; Calderon-Oliver, M.; Tapia, E.; Zazueta, C.; Pedraza-Chaverri, J. Curcumin protects from cardiac reperfusion damage by attenuation of oxidant stress and mitochondrial dysfunction. Cardiovasc. Toxicol. 2011, 11, 357–364. [Google Scholar] [CrossRef]
- Assis, R.P.; Arcaro, C.A.; Gutierres, V.O.; Oliveira, J.O.; Costa, P.I.; Baviera, A.M.; Brunetti, I.L. Combined Effects of Curcumin and Lycopene or Bixin in Yoghurt on Inhibition of LDL Oxidation and Increases in HDL and Paraoxonase Levels in Streptozotocin-Diabetic Rats. Int. J. Mol. Sci. 2017, 18, 332. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Wang, B.; Cao, S.H.; Wang, Y.Q. Epigallocatechin-3-Gallate (EGCG) Attenuates Traumatic Brain Injury by Inhibition of Edema Formation and Oxidative Stress. Korean J. Physiol. Pha. 2015, 19, 491–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Morris, A.; Sunkara, M.; Layne, J.; Toborek, M.; Hennig, B. Epigallocatechin-gallate stimulates NF-E2-related factor and heme oxygenase-1 via caveolin-1 displacement. J. Nutr. Biochem. 2012, 23, 163–168. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Li, X.L.; Liu, X.; Wang, C.; Zhou, D.S.; Ma, Q.; Zhou, W.H.; Hu, Z.Y. Antinociceptive effects of fisetin against diabetic neuropathic pain in mice: Engagement of antioxidant mechanisms and spinal GABAA receptors. Pharmacol. Res. 2015, 102, 286–297. [Google Scholar] [CrossRef]
- Kumar, B.H.; Reddy, R.A.; Kumar, J.M.; Kumar, B.D.; Diwan, P.V. Effects of fisetin on hyperhomocysteinemia-induced experimental endothelial dysfunction and vascular dementia. Can. J. Physiol. Pharmacol. 2017, 95, 32–42. [Google Scholar] [CrossRef] [Green Version]
- Dong, B.; Liu, C.; Xue, R.C.; Wang, Y.; Sun, Y.; Liang, Z.M.; Fan, W.D.; Jiang, J.Z.; Zhao, J.J.; Su, Q.; et al. Fisetin inhibits cardiac hypertrophy by suppressing oxidative stress. J. Nutr. Biochem. 2018, 62, 221–229. [Google Scholar] [CrossRef]
- Cho, B.O.; Yin, H.H.; Park, S.H.; Byun, E.B.; Ha, H.Y.; Jang, S.I. Anti-inflammatory activity of myricetin from Diospyros lotus through suppression of NF-B and STAT1 activation and Nrf2-mediated HO-1 induction in lipopolysaccharide-stimulated RAW264.7 macrophages. Biosci. Biotech. Bioch. 2016, 80, 1520–1530. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.J.; Meng, Y.H.; Zhao, Y.; Hu, Y.Y.; Ren, D.Y.; Yang, X.B. Myricetin derived from Hovenia dulcis Thunb. ameliorates vascular endothelial dysfunction and liver injury in high choline-fed mice. Food Funct. 2015, 6, 1620–1634. [Google Scholar] [CrossRef] [PubMed]
- Chis, I.C.; Coseriu, A.; Simedrea, R.; Oros, A.; Nagy, A.L.; Clichici, S. In Vivo Effects of Quercetin in Association with Moderate Exercise Training in Improving Streptozotocin-Induced Aortic Tissue Injuries. Molecules 2015, 20, 21770–21786. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Tian, R.; Lu, N. Quercetin Inhibited Endothelial Dysfunction and Atherosclerosis in Apolipoprotein E-Deficient Mice: Critical Roles for NADPH Oxidase and Heme Oxygenase-1. J. Agric. Food Chem. 2020, 68, 10875–10883. [Google Scholar] [CrossRef] [PubMed]
- Spanier, G.; Xu, H.; Xia, N.; Tobias, S.; Deng, S.; Wojnowski, L.; Forstermann, U.; Li, H. Resveratrol Reduces Endothelial Oxidative Stress by Modulating the Gene Expression of Superoxide Dismutase 1 (Sod1), Glutathione Peroxidase 1 (Gpx1) and Nadph Oxidase Subunit (Nox4). J. Physiol. Pharmacol. 2009, 60, 111–116. [Google Scholar] [PubMed]
- Gocmez, S.S.; Sahin, T.D.; Yazir, Y.; Duruksu, G.; Eraldemir, F.C.; Polat, S.; Utkan, T. Resveratrol prevents cognitive deficits by attenuating oxidative damage and inflammation in rat model of streptozotocin diabetes induced vascular dementia. Physiol. Behav. 2019, 201, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Hammad, A.S.A.; Ahmed, A.F.; Heeba, G.H.; Taye, A. Heme oxygenase-1 contributes to the protective effect of resveratrol against endothelial dysfunction in STZ-induced diabetes in rats. Life Sci. 2019, 239, 117065. [Google Scholar] [CrossRef] [PubMed]
- Do, G.M.; Kwon, E.Y.; Kim, H.J.; Jeon, S.M.; Ha, T.Y.; Park, T.; Choi, M.S. Long-term effects of resveratrol supplementation on suppression of atherogenic lesion formation and cholesterol synthesis in apo E-deficient mice. Biochem. Biophys. Res. Commun. 2008, 374, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Tian, S.; Liu, X.; He, C.; Ding, Z.; Shan, Y. Sulforaphane protected the injury of human vascular endothelial cell induced by LPC through up-regulating endogenous antioxidants and phase II enzymes. Food Funct 2015, 6, 1984–1991. [Google Scholar] [CrossRef] [PubMed]
- Shan, Y.J.; Zhao, R.F.; Geng, W.; Lin, N.; Wang, X.X.; Du, X.Y.; Wang, S.R. Protective Effect of Sulforaphane on Human Vascular Endothelial Cells Against Lipopolysaccharide-Induced Inflammatory Damage. Cardiovasc. Toxicol. 2010, 10, 139–145. [Google Scholar] [CrossRef]
- Alfieri, A.; Srivastava, S.; Siow, R.C.M.; Cash, D.; Modo, M.; Duchen, M.R.; Fraser, P.A.; Williams, S.C.R.; Mann, G.E. Sulforaphane preconditioning of the Nrf2/HO-1 defense pathway protects the cerebral vasculature against blood-brain barrier disruption and neurological deficits in stroke. Free Radic. Biol. Med. 2013, 65, 1012–1022. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Wang, X.; Gao, J.; Guo, Y.; Huang, Z.; Du, Y. The Tomato Hoffman’s Anthocyaninless Gene Encodes a bHLH Transcription Factor Involved in Anthocyanin Biosynthesis That Is Developmentally Regulated and Induced by Low Temperatures. PLoS ONE 2016, 11, e0151067. [Google Scholar] [CrossRef] [Green Version]
- Veljkovic, M.; Pavlovic, D.R.; Stojiljkovic, N.; Ilic, S.; Jovanovic, I.; Poklar Ulrih, N.; Rakic, V.; Velickovic, L.; Sokolovic, D. Bilberry: Chemical Profiling, in Vitro and in Vivo Antioxidant Activity and Nephroprotective Effect against Gentamicin Toxicity in Rats. Phytother Res. 2017, 31, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Rechner, A.R.; Kroner, C. Anthocyanins and colonic metabolites of dietary polyphenols inhibit platelet function. Thromb. Res. 2005, 116, 327–334. [Google Scholar] [CrossRef]
- Yu, C.C.; Nandrot, E.F.; Dun, Y.; Finnemann, S.C. Dietary antioxidants prevent age-related retinal pigment epithelium actin damage and blindness in mice lacking alphavbeta5 integrin. Free Radic. Biol. Med. 2012, 52, 660–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.; Guo, M.; Li, Q.; Peng, L.; Liu, H.; Zhang, L.; Bai, X.; Wang, Y.; Li, J.; Cai, C. Plasma metabolomic profiling to reveal antipyretic mechanism of Shuang-huang-lian injection on yeast-induced pyrexia rats. PLoS ONE 2014, 9, e100017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, W.T.; Shao, Z.H.; Yin, J.J.; Mehendale, S.; Wang, C.Z.; Qin, Y.M.; Li, J.; Chen, W.J.; Chien, C.T.; Becker, L.B.; et al. Comparative effects of flavonoids on oxidant scavenging and ischemia-reperfusion injury in cardiomyocytes. Eur. J. Pharmacol. 2007, 566, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Wartenberg, M.; Budde, P.; de Marees, M.; Grunheck, F.; Tsang, S.Y.; Huang, Y.; Chen, Z.Y.; Hescheler, J.; Sauer, H. Inhibition of tumor-induced angiogenesis and matrix-metalloproteinase expression in confrontation cultures of embryoid bodies and tumor spheroids by plant ingredients used in traditional Chinese medicine. Lab. Invest. 2003, 83, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.L.; Chan, K.G.; Pusparajah, P.; Duangjai, A.; Saokaew, S.; Mehmood Khan, T.; Lee, L.H.; Goh, B.H. Rhizoma Coptidis: A Potential Cardiovascular Protective Agent. Front Pharm. 2016, 7, 362. [Google Scholar] [CrossRef] [Green Version]
- Lan, T.; Liu, W.; Xie, X.; Huang, K.; Peng, J.; Huang, J.; Shen, X.; Liu, P.; Yang, H.; Huang, H. Berberine suppresses high glucose-induced TGF-beta1 and fibronectin synthesis in mesangial cells through inhibition of sphingosine kinase 1/AP-1 pathway. Eur. J. Pharmacol. 2012, 697, 165–172. [Google Scholar] [CrossRef]
- Hsu, Y.Y.; Tseng, Y.T.; Lo, Y.C. Berberine, a natural antidiabetes drug, attenuates glucose neurotoxicity and promotes Nrf2-related neurite outgrowth. Toxicol. Appl. Pharm. 2013, 272, 787–796. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, Y.; Xu, J.; Xue, Z.; Zhang, M.; Pang, X.; Zhang, X.; Zhao, L. Modulation of gut microbiota by berberine and metformin during the treatment of high-fat diet-induced obesity in rats. Sci. Rep. 2015, 5, 14405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, M.S.; Fleenor, B.S. The emerging role of curcumin for improving vascular dysfunction: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 2790–2799. [Google Scholar] [CrossRef] [PubMed]
- Sarraf, P.; Parohan, M.; Javanbakht, M.H.; Ranji-Burachaloo, S.; Djalali, M. Short-term curcumin supplementation enhances serum brain-derived neurotrophic factor in adult men and women: A systematic review and dose-response meta-analysis of randomized controlled trials. Nutr. Res. 2019, 69, 1–8. [Google Scholar] [CrossRef]
- Yang, M.; Akbar, U.; Mohan, C. Curcumin in Autoimmune and Rheumatic Diseases. Nutrients 2019, 11, 1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motterlini, R.; Foresti, R.; Bassi, R.; Green, C.J. Curcumin, an antioxidant and anti-inflammatory agent, induces heme oxygenase-1 and protects endothelial cells against oxidative stress. Free Radic. Biol. Med. 2000, 28, 1303–1312. [Google Scholar] [CrossRef]
- Yamagata, K. Protective Effect of Epigallocatechin Gallate on Endothelial Disorders in Atherosclerosis. J. Cardiovasc. Pharmacol. 2020, 75, 292–298. [Google Scholar] [CrossRef]
- Chen, D.; Wan, S.B.; Yang, H.J.; Yuan, J.; Chan, T.H.; Dou, Q.P. Egcg, Green Tea Polyphenols and Their Synthetic Analogs and Prodrugs for Human Cancer Prevention and Treatment. Adv. Clin. Chem. 2011, 53, 155–177. [Google Scholar] [CrossRef]
- Khan, N.; Syed, D.N.; Ahmad, N.; Mukhtar, H. Fisetin: A dietary antioxidant for health promotion. Antioxid. Redox Signal. 2013, 19, 151–162. [Google Scholar] [CrossRef]
- Maurya, B.K.; Trigun, S.K. Fisetin Modulates Antioxidant Enzymes and Inflammatory Factors to Inhibit Aflatoxin-B1 Induced Hepatocellular Carcinoma in Rats. Oxid. Med. Cell Longev. 2016, 2016, 1972793. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.E.; Jeong, S.I.; Yang, H.; Park, C.S.; Jin, Y.H.; Park, Y.S. Fisetin induces Nrf2-mediated HO-1 expression through PKC-delta and p38 in human umbilical vein endothelial cells. J. Cell Biochem. 2011, 112, 2352–2360. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.A.; Lee, S.; Kim, D.E.; Kim, M.; Kwon, B.M.; Han, D.C. Fisetin, a dietary flavonoid, induces apoptosis of cancer cells by inhibiting HSF1 activity through blocking its binding to the hsp70 promoter. Carcinogenesis 2015, 36, 696–706. [Google Scholar] [CrossRef] [PubMed]
- Currais, A.; Prior, M.; Dargusch, R.; Armando, A.; Ehren, J.; Schubert, D.; Quehenberger, O.; Maher, P. Modulation of p25 and inflammatory pathways by fisetin maintains cognitive function in Alzheimer’s disease transgenic mice. Aging Cell 2014, 13, 379–390. [Google Scholar] [CrossRef]
- Choi, H.N.; Kang, M.J.; Lee, S.J.; Kim, J.I. Ameliorative effect of myricetin on insulin resistance in mice fed a high-fat, high-sucrose diet. Nutr. Res. Pract. 2014, 8, 544–549. [Google Scholar] [CrossRef] [PubMed]
- Bennett, C.J.; Caldwell, S.T.; McPhail, D.B.; Morrice, P.C.; Duthie, G.G.; Hartley, R.C. Potential therapeutic antioxidants that combine the radical scavenging ability of myricetin and the lipophilic chain of vitamin E to effectively inhibit microsomal lipid peroxidation. Bioorg. Med. Chem. 2004, 12, 2079–2098. [Google Scholar] [CrossRef]
- Eid, H.M.; Haddad, P.S. The Antidiabetic Potential of Quercetin: Underlying Mechanisms. Curr. Med. Chem. 2017, 24, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Edwards, R.L.; Lyon, T.; Litwin, S.E.; Rabovsky, A.; Symons, J.D.; Jalili, T. Quercetin reduces blood pressure in hypertensive subjects. J. Nutr. 2007, 137, 2405–2411. [Google Scholar] [CrossRef]
- Loke, W.M.; Hodgson, J.M.; Proudfoot, J.M.; McKinley, A.J.; Puddey, I.B.; Croft, K.D. Pure dietary flavonoids quercetin and (-)-epicatechin augment nitric oxide products and reduce endothelin-1 acutely in healthy men. Am. J. Clin. Nutr. 2008, 88, 1018–1025. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.H.; Park, E.; Lee, H.J.; Kim, M.O.; Cha, Y.J.; Kim, J.M.; Lee, H.; Shin, M.J. Effects of daily quercetin-rich supplementation on cardiometabolic risks in male smokers. Nutr. Res. Pract. 2011, 5, 28–33. [Google Scholar] [CrossRef] [Green Version]
- Pfeuffer, M.; Auinger, A.; Bley, U.; Kraus-Stojanowic, I.; Laue, C.; Winkler, P.; Rufer, C.E.; Frank, J.; Boesch-Saadatmandi, C.; Rimbach, G.; et al. Effect of quercetin on traits of the metabolic syndrome, endothelial function and inflammation in men with different APOE isoforms. Nutr. Metab. Cardiovas 2013, 23, 403–409. [Google Scholar] [CrossRef]
- Perez, A.; Gonzalez-Manzano, S.; Jimenez, R.; Perez-Abud, R.; Haro, J.M.; Osuna, A.; Santos-Buelga, C.; Duarte, J.; Perez-Vizcaino, F. The flavonoid quercetin induces acute vasodilator effects in healthy volunteers: Correlation with beta-glucuronidase activity. Pharmacol. Res. 2014, 89, 11–18. [Google Scholar] [CrossRef]
- Dobrydnev, A.V.; Tkachuka, T.M.; Atamaniuka, V.P.; Popova, M.V. Quercetin-Amino Acid Conjugates are Promising Anti-Cancer Agents in Drug Discovery Projects. Mini-Rev. Med. Chem. 2020, 20, 107–122. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.W.; Liu, X.L.; Kong, L.; Zhang, M.Y.; Chen, Y.J.; Zhu, X.; Hao, Y.C. Neuroprotection of quercetin on central neurons against chronic high glucose through enhancement of Nrf2/ARE/glyoxalase-1 pathway mediated by phosphorylation regulation. Biomed. Pharmacother. 2019, 109, 2145–2154. [Google Scholar] [CrossRef] [PubMed]
- Muthuraman, A.; Kaur, P.; Kaur, P.; Singh, H.; Boparai, P.S. Ameliorative potential of vitamin P and digoxin in ischemic-reperfusion induced renal injury using the Langendorff apparatus. Life Sci. 2015, 124, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Stainer, A.R.; Sasikumar, P.; Bye, A.P.; Unsworth, A.J.; Holbrook, L.M.; Tindall, M.; Lovegrove, J.A.; Gibbins, J.M. The Metabolites of the Dietary Flavonoid Quercetin Possess Potent Antithrombotic Activity, and Interact with Aspirin to Enhance Antiplatelet Effects. TH Open 2019, 3, e244–e258. [Google Scholar] [CrossRef] [PubMed]
- Abbott, J.A.; Medina-Bolivar, F.; Martin, E.M.; Engelberth, A.S.; Villagarcia, H.; Clausen, E.C.; Carrier, D.J. Purification of Resveratrol, Arachidin-1, and Arachidin-3 from Hairy Root Cultures of Peanut (Arachis hypogaea) and Determination of Their Antioxidant Activity and Cytotoxicity. Biotechnol. Prog. 2010, 26, 1344–1351. [Google Scholar] [CrossRef]
- Lyons, M.M.; Yu, C.W.; Toma, R.B.; Cho, S.Y.; Reiboldt, W.; Lee, J.; van Breemen, R.B. Resveratrol in raw and baked blueberries and bilberries. J. Agric. Food Chem. 2003, 51, 5867–5870. [Google Scholar] [CrossRef]
- Tian, X.; Liu, Y.L.; Ren, G.W.; Yin, L.; Liang, X.Y.; Geng, T.; Dang, H.Q.; An, R.F. Resveratrol limits diabetes-associated cognitive decline in rats by preventing oxidative stress and inflammation and modulating hippocampal structural synaptic plasticity. Brain Res. 2016, 1650, 1–9. [Google Scholar] [CrossRef]
- Molino, S.; Dossena, M.; Buonocore, D.; Ferrari, F.; Venturini, L.; Ricevuti, G.; Verri, M. Polyphenols in dementia: From molecular basis to clinical trials. Life Sci. 2016, 161, 69–77. [Google Scholar] [CrossRef]
- Yazir, Y.; Utkan, T.; Gacar, N.; Aricioglu, F. Resveratrol exerts anti-inflammatory and neuroprotective effects to prevent memory deficits in rats exposed to chronic unpredictable mild stress. Physiol. Behav. 2015, 138, 297–304. [Google Scholar] [CrossRef]
- Tiwari, V.; Chopra, K. Resveratrol abrogates alcohol-induced cognitive deficits by attenuating oxidative-nitrosative stress and inflammatory cascade in the adult rat brain. Neurochem. Int. 2013, 62, 861–869. [Google Scholar] [CrossRef]
- Gocmez, S.S.; Gacar, N.; Utkan, T.; Gacar, G.; Scarpace, P.J.; Tumer, N. Protective effects of resveratrol on aging-induced cognitive impairment in rats. Neurobiol. Learn. Mem. 2016, 131, 131–136. [Google Scholar] [CrossRef]
- Tanno, M.; Kuno, A.; Yano, T.; Miura, T.; Hisahara, S.; Ishikawa, S.; Shimamoto, K.; Horio, Y. Induction of Manganese Superoxide Dismutase by Nuclear Translocation and Activation of SIRT1 Promotes Cell Survival in Chronic Heart Failure. J. Biol. Chem. 2010, 285, 8375–8382. [Google Scholar] [CrossRef] [Green Version]
- Tortorella, S.M.; Royce, S.G.; Licciardi, P.V.; Karagiannis, T.C. Dietary Sulforaphane in Cancer Chemoprevention: The Role of Epigenetic Regulation and HDAC Inhibition. Antioxid. Redox Signal. 2015, 22, 1382–1424. [Google Scholar] [CrossRef] [Green Version]
- Dinkova-Kostova, A.T.; Kostov, R.V. Glucosinolates and isothiocyanates in health and disease. Trends Mol. Med. 2012, 18, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Bhakkiyalakshmi, E.; Sireesh, D.; Rajaguru, P.; Paulmurugan, R.; Ramkumar, K.M. The emerging role of redox-sensitive Nrf2-Keap1 pathway in diabetes. Pharmacol. Res. 2015, 91, 104–114. [Google Scholar] [CrossRef]
- Cheung, K.L.; Kong, A.N. Molecular targets of dietary phenethyl isothiocyanate and sulforaphane for cancer chemoprevention. AAPS J 2010, 12, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Nelson, K.M.; Dahlin, J.L.; Bisson, J.; Graham, J.; Pauli, G.F.; Walters, M.A. The Essential Medicinal Chemistry of Curcumin. J. Med. Chem. 2017, 60, 1620–1637. [Google Scholar] [CrossRef]
Figure 1.
Signaling pathways involved in antioxidant enzymes.

Figure 2.
Antioxidant enzymes in vascular protection. Oxidative stress and inflammation in the vascular system response to multiple kinds of inducers, subsequently leading to several pathological conditions. This review supposed that induction of antioxidant enzyme expression provides cytoprotection against vascular injury and preserves vascular function.
Figure 2.
Antioxidant enzymes in vascular protection. Oxidative stress and inflammation in the vascular system response to multiple kinds of inducers, subsequently leading to several pathological conditions. This review supposed that induction of antioxidant enzyme expression provides cytoprotection against vascular injury and preserves vascular function.


{kind=link}
{kind=link}
Table 1.
Summary of phytochemicals with modulating Nrf2 activity and antioxidant enzymes.
| Phytochemical | Effects | Altered Antioxidant enzyme |
|---|---|---|
| Anthocyanin | Prevent eye disease | SOD, CAT, GPx [71] |
| Anti-diabetic effect | CAT, SOD [72] | |
| Hypotensive effect | HO-1, SOD [73] | |
| Baicalein | Anti-ischemic effect | HO-1 [74] |
| Cardiopulmonary protective effect | GPx, SOD [75] | |
| Berberine | Anti-inflammation effect | NQO-1, HO-1 [76] |
| Anti-atherosclerotic effect | HO-1 [77] | |
| Anti-diabetic effect | GR [78] | |
| Curcumin | Anti-atherosclerotic effect | HO-1 [79] |
| Cardio-protective effect | SOD [80] | |
| Anti-inflammation effect | HO-1 [81] | |
| Vasculoprotective effect | CAT [82] | |
| Cardio-protective effect | GPx, GR [83] | |
| Cardio-protective effect | GR, SOD [84] | |
| Anti-diabetic effect | PON1 [85] | |
| EGCG | Neuroprotective effect | SOD, GPx [86] |
| Anti-inflammation effect | HO-1 [87] | |
| Fisetin | Anti-inflammation effect | HO-1 [88] |
| Neuroprotective effect | SOD, CAT [89] | |
| Anti-hypertrophic effect | SOD, CAT, HO-1 [90] | |
| Myricetin | Anti-inflammation effect | HO-1 [91] |
| Anti-oxidative effect | SOD, GPx [92] | |
| Quercetin | Anti-diabetic effect | SOD, CAT [93] |
| Vasculoprotective effect | HO-1 [94] | |
| Resveratrol | Vasculoprotective effect | SOD, GPx [95] |
| Neuroprotective effect | SOD, HO-1 [96] | |
| Vasculoprotective effect | HO-1 [97] | |
| Anti-atherosclerotic effect | PON [98] | |
| Sulforaphane | Cardio-protective effect | GPx, SOD, TrxR [99] |
| Anti-inflammation effect | TrxR, HO-1 [100] | |
| Anti-ischemic effect | HO-1 [101] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lee, S.E.; Park, Y.S. The Emerging Roles of Antioxidant Enzymes by Dietary Phytochemicals in Vascular Diseases. Life 2021, 11, 199. https://0-doi-org.brum.beds.ac.uk/10.3390/life11030199
AMA Style
Lee SE, Park YS. The Emerging Roles of Antioxidant Enzymes by Dietary Phytochemicals in Vascular Diseases. Life. 2021; 11(3):199. https://0-doi-org.brum.beds.ac.uk/10.3390/life11030199
Chicago/Turabian StyleLee, Seung Eun, and Yong Seek Park. 2021. "The Emerging Roles of Antioxidant Enzymes by Dietary Phytochemicals in Vascular Diseases" Life 11, no. 3: 199. https://0-doi-org.brum.beds.ac.uk/10.3390/life11030199
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.